
Isolation and Characterization of Polysaccharides from the Ascidian Styela clava

 ,
,  ,
, 

Abstract
:1. Introduction
2. Materials and Methods
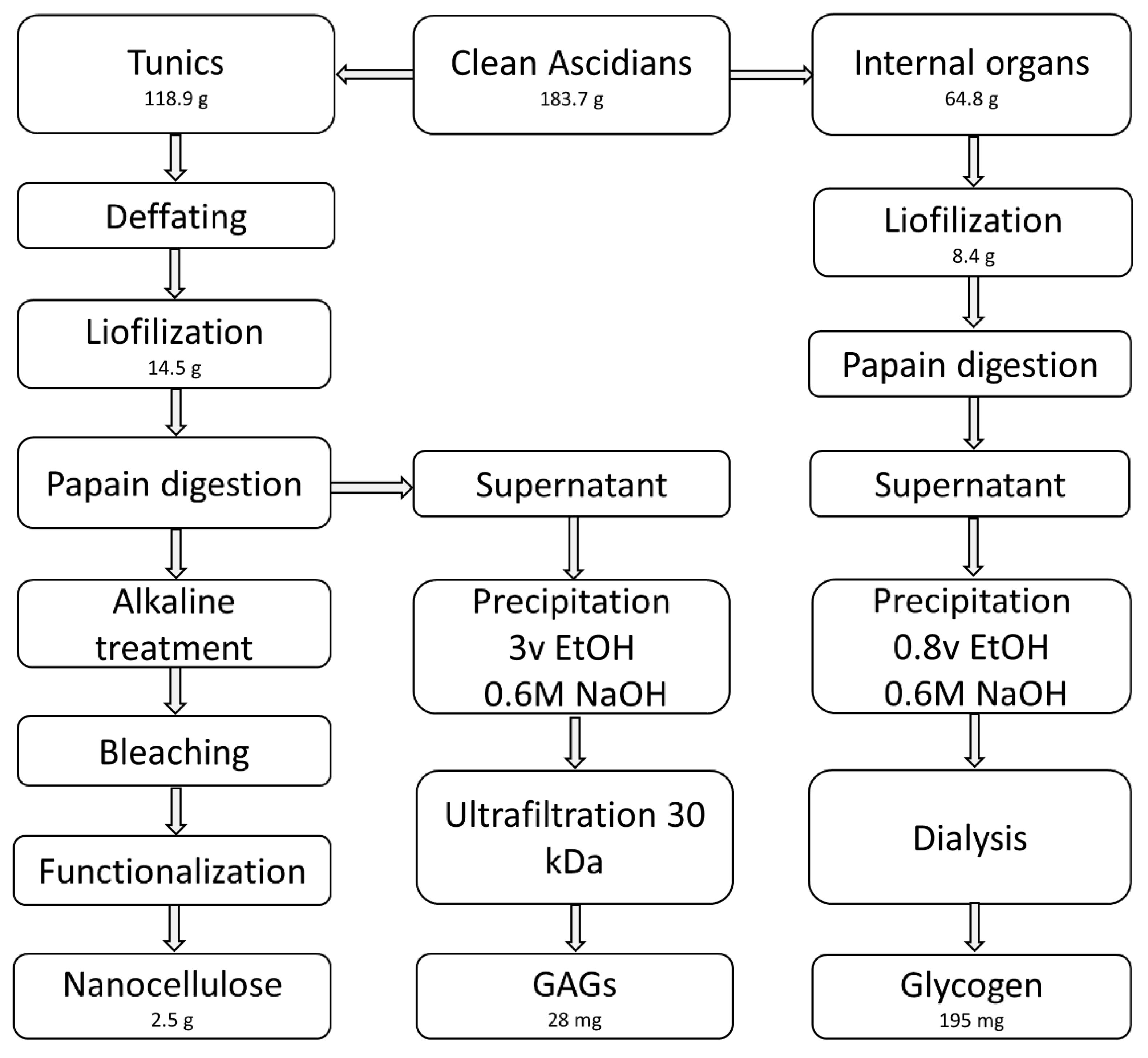
2.1. Raw Materials
2.2. Extraction and Isolation
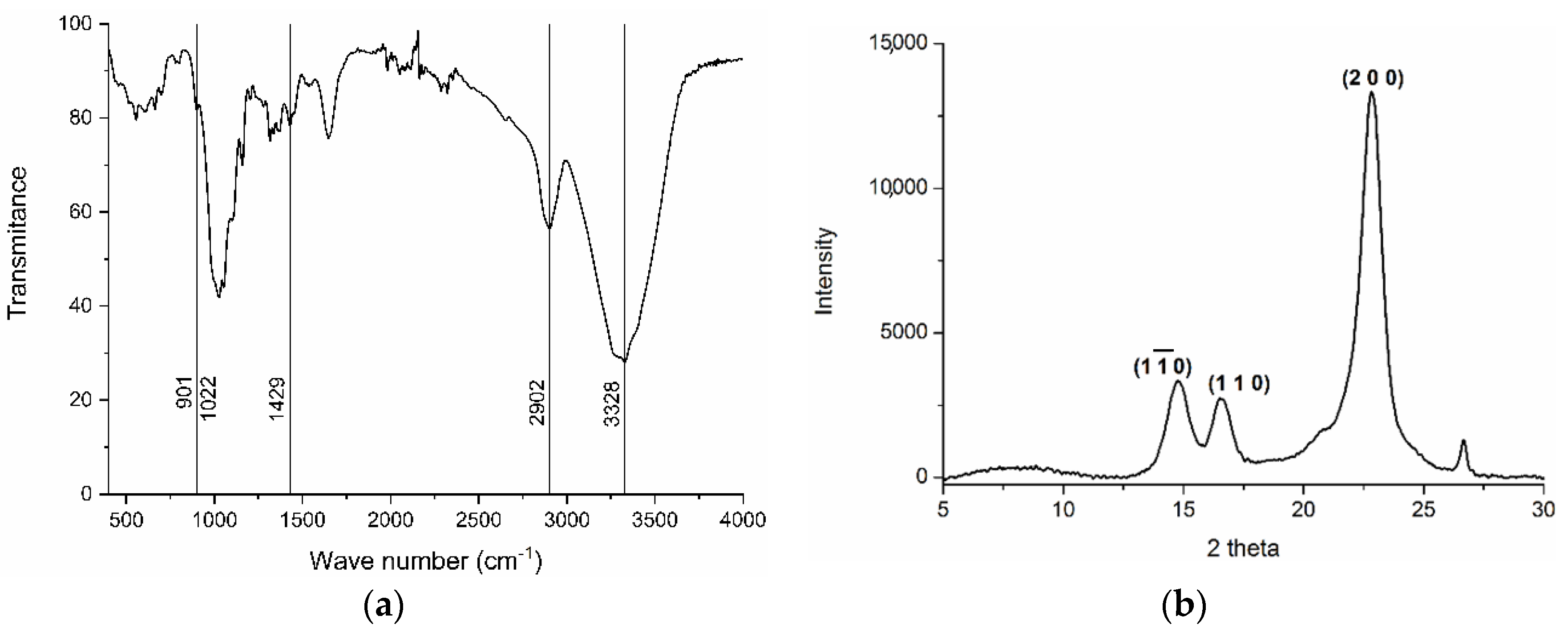
2.3. Cellulose Characterisation
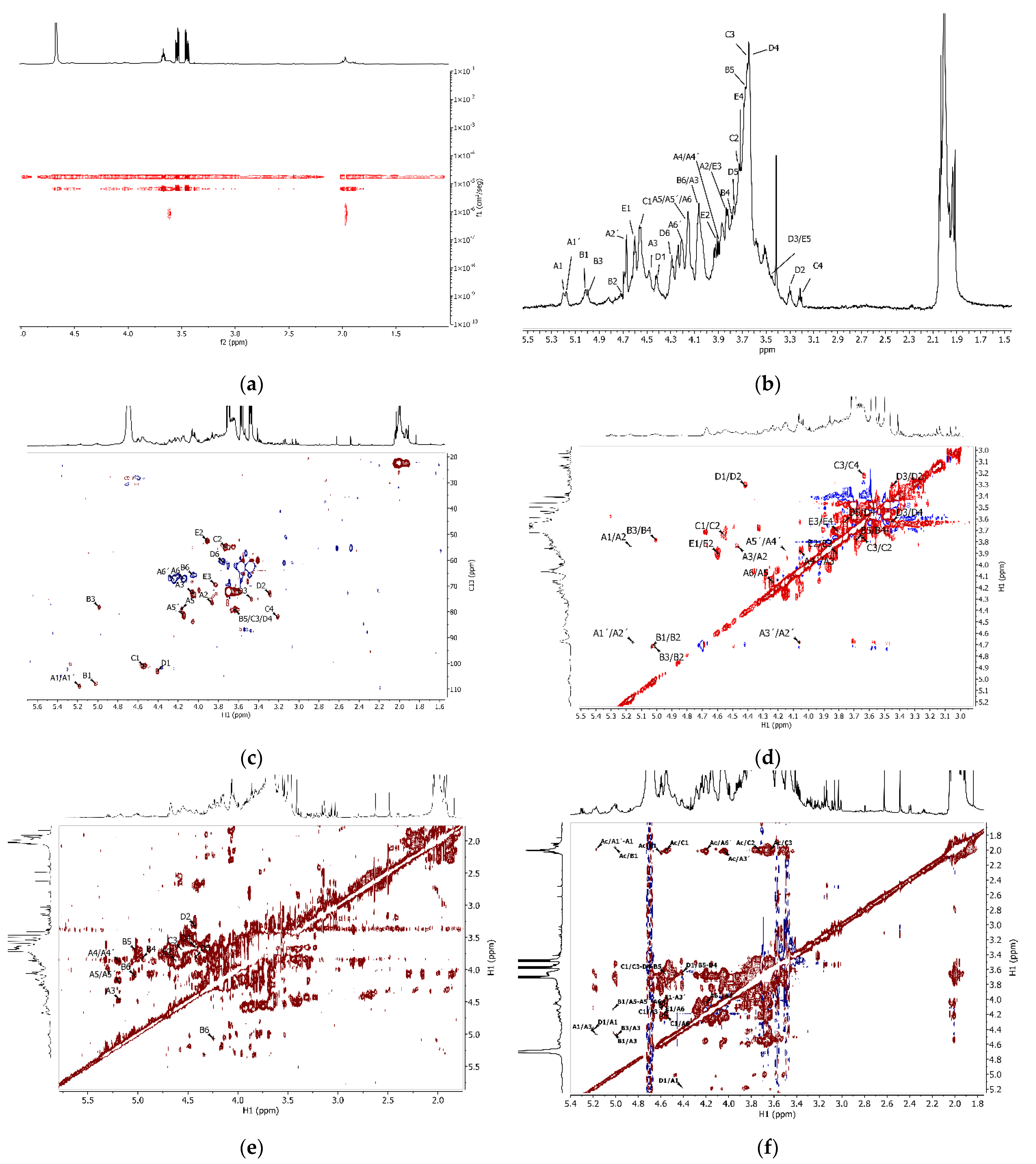
2.4. Polysaccharide Characterisation
3. Results and Discussion
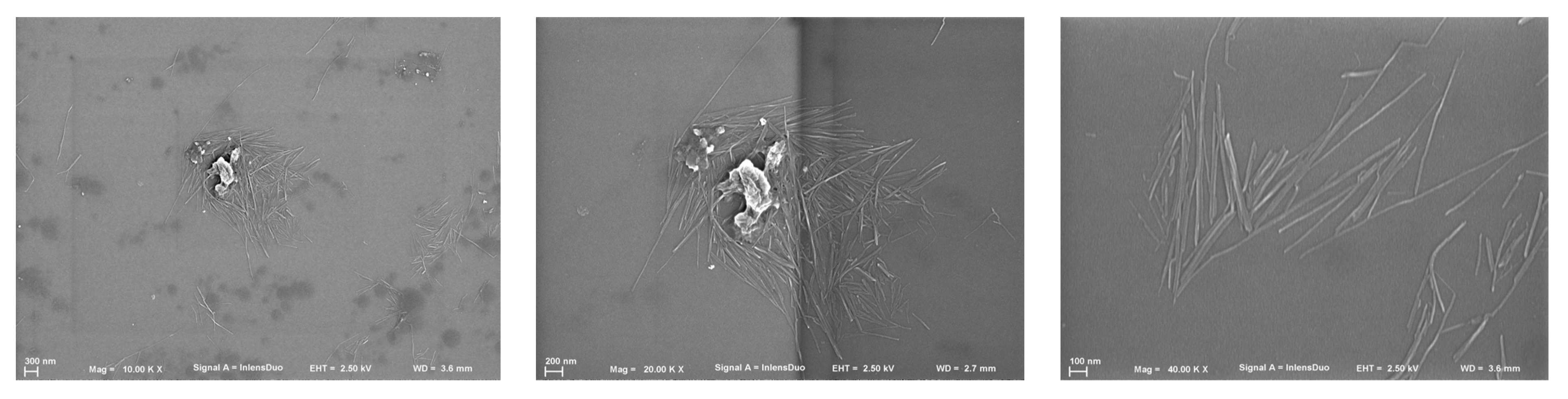
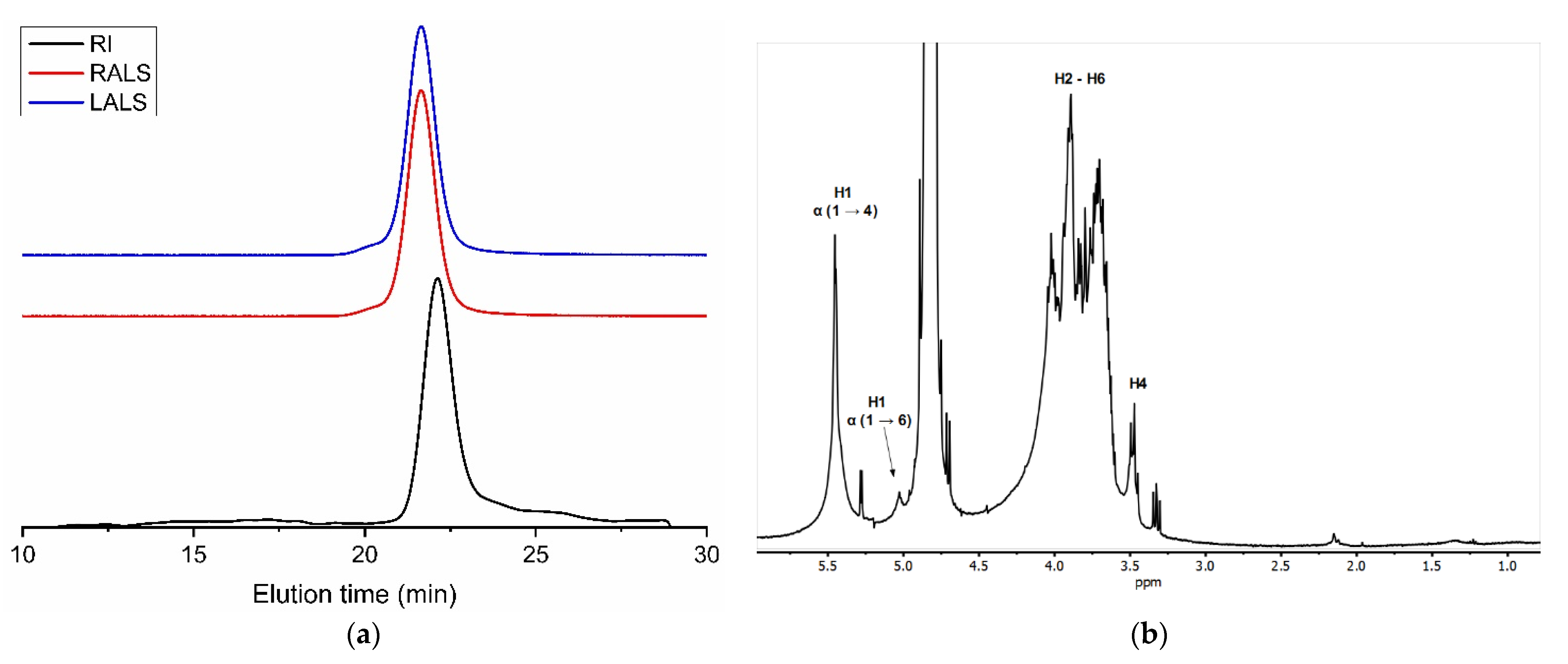
3.1. Nanocellulose
3.2. Tunic Polysaccharides
3.3. Glycogen from the Internal Organs
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Soliman, T. Forecasting the economic impacts of two biofouling invaders on aquaculture production of green-lipped mussels Perna canaliculus in New Zealand. Aquac. Environ. Interact. 2018, 10, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Lins, D.M.; de Marco, P., Jr.; Andrade, A.F.A.; Rocha, R.M. Predicting global ascidian invasions. Divers. Distrib. 2018, 24, 692–704. [Google Scholar] [CrossRef] [Green Version]
- Dupont, L.; Viard, F.; Davis, M.H.; Nishikawa, T.; Bishop, J.D.D. Pathways of spread of the introduced ascidian Styela clava (Tunicata) in Northern Europe, as revealed by microsatellite markers. Biol. Invasions 2010, 12, 2707–2721. [Google Scholar] [CrossRef]
- Lambert, G.; Karney, R.C.; Rhee, W.Y.; Carman, M.R. Wild and cultured edible tunicates: A review. Manag. Biol. Invasions 2016, 7, 59–66. [Google Scholar] [CrossRef] [Green Version]
- van den Berg, O.; Capadona, J.R.; Weder, C. Preparation of Homogeneous Dispersions of Tunicate Cellulose Whiskers in Organic Solvents. Biomacromolecules 2007, 8, 1353–1357. [Google Scholar] [CrossRef] [PubMed]
- Bacakova, L.; Pajorova, J.; Bacakova, M.; Skogberg, A.; Kallio, P.; Kolarova, K.; Svorcik, V. Versatile Application of Nanocellulose: From Industry to Skin Tissue Engineering and Wound Healing. Nanomaterials 2019, 9, 164. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.M.; Park, J.M.; Kang, T.Y.; Kim, Y.S.; Lee, S.K. Purification of squirt cellulose membrane from the cystic tunic of Styela clava and identification of its osteoconductive effect. Cellulose 2013, 20, 655–673. [Google Scholar] [CrossRef]
- Song, S.H.; Seong, K.Y.; Kim, J.E.; Go, J.; Koh, E.K.; Sung, J.E.; Son, H.J.; Jung, Y.J.; Kim, H.S.; Hong, J.T. Effects of different cellulose membranes regenerated from Styela clava tunics on wound healing. Int. J. Mol. Med. 2017, 39, 1173–1187. [Google Scholar] [CrossRef] [Green Version]
- Park, J.J.; Kim, J.E.; Yun, W.B.; Lee, M.R.; Choi, J.Y.; Song, B.R.; Son, H.J.; Lim, Y.; Kang, H.; An, B.S. Therapeutic effects of a liquid bandage prepared with cellulose powders from Styela clava tunics and Broussonetia kazinoki bark: Healing of surgical wounds on the skin of Sprague Dawley rats. Mol. Med. Rep. 2019, 19, 452–460. [Google Scholar] [CrossRef] [Green Version]
- Klemm, D.; Kramer, F.; Moritz, S.; Lindström, T.; Ankerfors, M.; Gray, D.; Dorris, A. Nanocelluloses: A New Family of Nature-Based Materials. Angew. Chem. Int. Ed. 2011, 50, 5438–5466. [Google Scholar] [CrossRef]
- Koh, E.K.; Kim, J.E.; Go, J.; Song, S.H.; Sung, J.E.; Son, H.J.; Jung, Y.J.; Kim, B.H.; Jung, Y.S.; Hwang, D.Y. Protective effects of the antioxidant extract collected from Styela clava tunics on UV radiation-induced skin aging in hairless mice. Int. J. Mol. Med. 2016, 38, 1565–1577. [Google Scholar] [CrossRef] [Green Version]
- Koh, E.K.; Kim, J.E.; Song, S.H.; Sung, J.E.; Lee, H.A.; Kim, K.S.; Hong, J.T.; Hwang, D.Y. Ethanol extracts collected from the Styela clava tunic alleviate hepatic injury induced by carbon tetrachloride (CCl4) through inhibition of hepatic apoptosis, inflammation, and fibrosis. J. Toxicol. Pathol. 2017, 30, 291–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, C.-X.; Jin, H.; Chung, Y.-S.; Shin, J.-Y.; Woo, M.-A.; Lee, K.-H.; Palmos, G.N.; Choi, B.-D.; Cho, M.-H. Chondroitin sulfate extracted from the Styela clava tunic suppresses TNF-α-induced expression of inflammatory factors, VCAM-1 and iNOS by blocking Akt/NF-κB signal in JB6 cells. Cancer Lett. 2008, 264, 93–100. [Google Scholar] [CrossRef]
- Ko, S.-C.; Jeon, Y.-J. Anti-inflammatory effect of enzymatic hydrolysates from Styela clava flesh tissue in lipopolysaccharide-stimulated RAW 264.7 macrophages and in vivo zebrafish model. Nutr. Res. Pract. 2015, 9, 219. [Google Scholar] [CrossRef] [Green Version]
- Jumeri; Kim, S.M. Antioxidant and anticancer activities of enzymatic hydrolysates of solitary tunicate (Styela clava). Food Sci. Biotechnol. 2011, 20, 1075. [Google Scholar] [CrossRef]
- Ko, S.-C.; Jung, W.-K.; Lee, S.-H.; Lee, D.H.; Jeon, Y.-J. Antihypertensive effect of an enzymatic hydrolysate from Styela clava flesh tissue in type 2 diabetic patients with hypertension. Nutr. Res. Pract. 2017, 11, 396. [Google Scholar] [CrossRef] [Green Version]
- Ko, S.-C.; Kim, J.-I.; Park, S.-J.; Jung, W.-K.; Jeon, Y.-J. Antihypertensive peptide purified from Styela clava flesh tissue stimulates glucose uptake through AMP-activated protein kinase (AMPK) activation in skeletal muscle cells. Eur. Food Res. Technol. 2016, 242, 163–170. [Google Scholar] [CrossRef]
- Kang, N.; Ko, S.-C.; Samarakoon, K.; Kim, E.-A.; Kang, M.-C.; Lee, S.-C.; Kim, J.; Kim, Y.-T.; Kim, J.-S.; Kim, H.; et al. Purification of antioxidative peptide from peptic hydrolysates of Mideodeok (Styela clava) flesh tissue. Food Sci. Biotechnol. 2013, 22, 541–547. [Google Scholar] [CrossRef]
- Ju, B.; Chen, B.; Zhang, X.; Han, C.; Jiang, A. Purification and Characterization of Bioactive Compounds from Styela clava. J. Chem. 2014, 2014, 525141. [Google Scholar] [CrossRef] [Green Version]
- Yun, S.-M.; Jang, J.-H.; Ryu, J.-E.; Choi, B.-D.; Lee, J.-S. Antibacterial sulfated alkene from a tunicate, Styela clava. Nat. Prod. Sci. 2007, 13, 132–134. [Google Scholar]
- Pavão, M.S.; Albano, R.M.; Lawson, A.M.; Mourão, P.A. Structural heterogeneity among unique sulfated L-galactans from different species of ascidians (tunicates). J. Biol. Chem. 1989, 264, 9972–9979. [Google Scholar] [CrossRef]
- Restrepo-Espinosa, D.C.; Román, Y.; Colorado-Ríos, J.; de Santana-Filho, A.P.; Sassaki, G.L.; Cipriani, T.R.; Martínez, A.; Iacomini, M.; Pavão, M.S.G. Structural analysis of a sulfated galactan from the tunic of the ascidian Microcosmus exasperatus and its inhibitory effect of the intrinsic coagulation pathway. Int. J. Biol. Macromol. 2017, 105, 1391–1400. [Google Scholar] [CrossRef]
- Pavao, M.S.G.; Aiello, K.R.M.; Werneck, C.C.; Silva, L.C.F.; Valente, A.-P.; Mulloy, B.; Colwell, N.S.; Tollefsen, D.M.; Mourão, P.A.S. Highly sulfated dermatan sulfates from ascidians structure versus anticoagulant activity of these glycosaminoglycans. J. Biol. Chem. 1998, 273, 27848–27857. [Google Scholar] [CrossRef] [Green Version]
- Karamanou, K.; Espinosa, D.C.R.; Fortuna-Costa, A.; Pavão, M.S.G. Biological function of unique sulfated glycosaminoglycans in primitive chordates. Glycoconj. J. 2017, 34, 277–283. [Google Scholar] [CrossRef]
- Ahn, S.H.; Jung, S.H.; Kang, S.J.; Jeong, T.S.; Choi, B.D. Extraction of glycosaminoglycans from Styela clava tunic. Biotechnol. Bioproc. Eng. 2003, 18, 180–185. [Google Scholar]
- AOAC Association of Official Analytical Chemistry. Methods of Analysis, 15th ed.; AOAC International: Washington DC, USA, 1997. [Google Scholar]
- Gomes, A.M.; Kozlowski, E.O.; Pomin, V.H.; de Barros, C.M.; Zaganeli, J.L.; Pavão, M.S.G. Unique Extracellular Matrix Heparan Sulfate from the Bivalve Nodipecten nodosus (Linnaeus, 1758) Safely Inhibits Arterial Thrombosis after Photochemically Induced Endothelial Lesion. J. Biol. Chem. 2010, 285, 7312–7323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vázquez, J.A.; Fraguas, J.; Novoa-Carballal, R.; Reis, R.L.; Pérez-Martín, R.I.; Valcarcel, J. Optimal isolation and characterisation of chondroitin sulfate from rabbit fish (Chimaera monstrosa). Carbohydr. Polym. 2019, 210. [Google Scholar] [CrossRef] [Green Version]
- Segal, L.; Creely, J.J.; Martin Jr, A.E.; Conrad, C.M. An empirical method for estimating the degree of crystallinity of native cellulose using the X-ray diffractometer. Text. Res. J. 1959, 29, 786–794. [Google Scholar] [CrossRef]
- Wu, D.H.; Chen, A.D.; Johnson, C.S. An Improved Diffusion-Ordered Spectroscopy Experiment Incorporating Bipolar-Gradient Pulses. J. Magn. Reson. Ser. A 1995, 115, 260–264. [Google Scholar] [CrossRef]
- Johnson, C.S. Diffusion ordered nuclear magnetic resonance spectroscopy: Principles and applications. Prog. Nucl. Magn. Reson. Spectrosc. 1999, 34, 203–256. [Google Scholar] [CrossRef]
- Koos, M.R.M.; Kummerlöwe, G.; Kaltschnee, L.; Thiele, C.M.; Luy, B. CLIP-COSY: A Clean In-Phase Experiment for the Rapid Acquisition of COSY-type Correlations. Angew. Chem. Int. Ed. 2016, 55, 7655–7659. [Google Scholar] [CrossRef]
- Bondeson, D.; Mathew, A.; Oksman, K. Optimization of the isolation of nanocrystals from microcrystalline cellulose by acid hydrolysis. Cellulose 2006, 13, 171. [Google Scholar] [CrossRef]
- Carpenter, A.W.; de Lannoy, C.-F.; Wiesner, M.R. Cellulose Nanomaterials in Water Treatment Technologies. Environ. Sci. Technol. 2015, 49, 5277–5287. [Google Scholar] [CrossRef]
- Sharma, P.R.; Chattopadhyay, A.; Sharma, S.K.; Hsiao, B.S. Efficient Removal of UO22+ from Water Using Carboxycellulose Nanofibers Prepared by the Nitro-Oxidation Method. Ind. Eng. Chem. Res. 2017, 56, 13885–13893. [Google Scholar] [CrossRef]
- Sharma, P.R.; Joshi, R.; Sharma, S.K.; Hsiao, B.S. A Simple Approach to Prepare Carboxycellulose Nanofibers from Untreated Biomass. Biomacromolecules 2017, 18, 2333–2342. [Google Scholar] [CrossRef]
- Sharma, P.R.; Chattopadhyay, A.; Sharma, S.K.; Geng, L.; Amiralian, N.; Martin, D.; Hsiao, B.S. Nanocellulose from Spinifex as an Effective Adsorbent to Remove Cadmium(II) from Water. ACS Sustain. Chem. Eng. 2018, 6, 3279–3290. [Google Scholar] [CrossRef]
- Zhan, C.; Sharma, P.R.; He, H.; Sharma, S.K.; McCauley-Pearl, A.; Wang, R.; Hsiao, B.S. Rice husk based nanocellulose scaffolds for highly efficient removal of heavy metal ions from contaminated water. Environ. Sci. Water Res. Technol. 2020, 6, 3080–3090. [Google Scholar] [CrossRef]
- Habibi, Y.; Lucia, L.A.; Rojas, O.J. Cellulose Nanocrystals: Chemistry, Self-Assembly, and Applications. Chem. Rev. 2010, 110, 3479–3500. [Google Scholar] [CrossRef] [PubMed]
- Dunlop, M.J.; Acharya, B.; Bissessur, R. Isolation of nanocrystalline cellulose from tunicates. J. Environ. Chem. Eng. 2018, 6, 4408–4412. [Google Scholar] [CrossRef]
- Dunlop, M.J.; Clemons, C.; Reiner, R.; Sabo, R.; Agarwal, U.P.; Bissessur, R.; Sojoudiasli, H.; Carreau, P.J.; Acharya, B. Towards the scalable isolation of cellulose nanocrystals from tunicates. Sci. Rep. 2020, 10, 19090. [Google Scholar] [CrossRef] [PubMed]
- Kimura, F.; Kimura, T.; Tamura, M.; Hirai, A.; Ikuno, M.; Horii, F. Magnetic Alignment of the Chiral Nematic Phase of a Cellulose Microfibril Suspension. Langmuir 2005, 21, 2034–2037. [Google Scholar] [CrossRef]
- Elazzouzi-Hafraoui, S.; Nishiyama, Y.; Putaux, J.-L.; Heux, L.; Dubreuil, F.; Rochas, C. The Shape and Size Distribution of Crystalline Nanoparticles Prepared by Acid Hydrolysis of Native Cellulose. Biomacromolecules 2008, 9, 57–65. [Google Scholar] [CrossRef]
- Zhao, Y.; Li, J. Excellent chemical and material cellulose from tunicates: Diversity in cellulose production yield and chemical and morphological structures from different tunicate species. Cellulose 2014, 21, 3427–3441. [Google Scholar] [CrossRef]
- Sun, J.X.; Sun, X.F.; Zhao, H.; Sun, R.C. Isolation and characterization of cellulose from sugarcane bagasse. Polym. Degrad. Stab. 2004, 84, 331–339. [Google Scholar] [CrossRef]
- Song, S.H.; Kim, J.E.; Lee, Y.J.; Kwak, M.H.; Sung, G.Y.; Kwon, S.H.; Son, H.J.; Lee, H.S.; Jung, Y.J.; Hwang, D.Y. Cellulose film regenerated from Styela clava tunics have biodegradability, toxicity and biocompatibility in the skin of SD rats. J. Mater. Sci. Mater. Med. 2014, 25, 1519–1530. [Google Scholar] [CrossRef] [PubMed]
- Vázquez, J.A.; Noriega, D.; Ramos, P.; Valcarcel, J.; Novoa-Carballal, R.; Pastrana, L.; Reis, R.L.; Pérez-Martín, R.I. Optimization of high purity chitin and chitosan production from Illex argentinus pens by a combination of enzymatic and chemical processes. Carbohydr. Polym. 2017, 174. [Google Scholar] [CrossRef] [PubMed]
- Vázquez, J.A.; Fraguas, J.; Novoa-Carvallal, R.; Reis, R.L.; Antelo, L.T.; Pérez-Martín, R.I.; Valcarcel, J. Isolation and chemical characterization of chondroitin sulfate from cartilage by-products of blackmouth catshark (Galeus melastomus). Mar. Drugs 2018, 16, 344. [Google Scholar] [CrossRef] [Green Version]
- Blundell, C.D.; Reed, M.A.C.; Almond, A. Complete assignment of hyaluronan oligosaccharides up to hexasaccharides. Carbohydr. Res. 2006, 341, 2803–2815. [Google Scholar] [CrossRef] [PubMed]
- Volpi, N.; Maccari, F. Purification and characterization of hyaluronic acid from the mollusc bivalve Mytilus galloprovincialis. Biochimie 2003, 85, 619–625. [Google Scholar] [CrossRef]
- Mucci, A.; Schenetti, L.; Volpi, N. 1 H and 13 C nuclear magnetic resonance identification and characterization of components of chondroitin sulfates of various origin. Carbohydr. Polym. 2000, 41, 37–45. [Google Scholar] [CrossRef]
- Santos, J.A.; Mulloy, B.; Mourão, P.A.S. Structural diversity among sulfated α-L-galactans from ascidians (tunicates). Eur. J. Biochem. 1992, 204, 669–677. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, R.G.S.; Cyr, N.; Korsch, B.; Koch, H.J.; Perlin, A.S. Carbon-13 Chemical Shifts of Furanosides and Cyclopentanols. Configurational and Conformational Influences. Can. J. Chem. 1975, 53, 1424–1433. [Google Scholar] [CrossRef]
- Zang, L.H.; Rothman, D.L.; Shulman, R.G. 1H NMR visibility of mammalian glycogen in solution. Proc. Natl. Acad. Sci. USA 1990, 87, 1678–1680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanoh, S.; Maeyama, K.; Tanaka, R.; Takahashi, T.; Aoyama, M.; Watanabe, M.; Iida, K.; Ueda, S.; Mae, M.; Takagi, K.; et al. Possible utilization of the pearl oyster phospholipid and glycogen as a cosmetic material. In More Efficient Utilization of Fish and Fisheries Products; Sakaguchi, M.B.T.-D., Ed.; Elsevier: Amsterdam, The Netherlands, 2004; Volume 42, pp. 179–190. ISBN 0167-4501. [Google Scholar]
- Vázquez, J.A.; Montemayor, M.I.; Fraguas, J.; Murado, M.A. Hyaluronic acid production by Streptococcus zooepidemicus in marine by-products media from mussel processing wastewaters and tuna peptone viscera. Microb. Cell Fact. 2010, 9, 46. [Google Scholar] [CrossRef] [Green Version]
- Gallardi, D.; Hobbs, K.; Mills, T.; Couturier, C.; Parrish, C.C.; Murray, H.M. Effects of extended ambient live holding on cultured blue mussels (Mytilus edulis L.) with reference to condition index, lipid profile, glycogen content and organoleptic testing. Aquaculture 2014, 430, 149–158. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | Ref | Length (nm) | Width (nm) | Aspect Ratio |
|---|---|---|---|---|
| Bacterial | [39] | 100–1000 | 10–50 | 20 |
| Cotton | [39] | 100–300 | 15 | 10–20 |
| S. clava (tunicate) | [40,41] | 1500 ± 600 | 18 ± 10 | 90 ± 30 |
| S. clava (tunicate) | This work | 1200 ± 600 | 19 ± 5 | 60 ± 30 |
| S. plicata (tunicate) | [40] | 1000–3000 | 20 | 50 |
| Halocinthyia roretzi (tunicate) | [42,43] | 1600 | 30 | 50 |
| Ciona intestinalis (tunicate) | [40,44] | 1300 ± 600 | 16–20 | 65 |
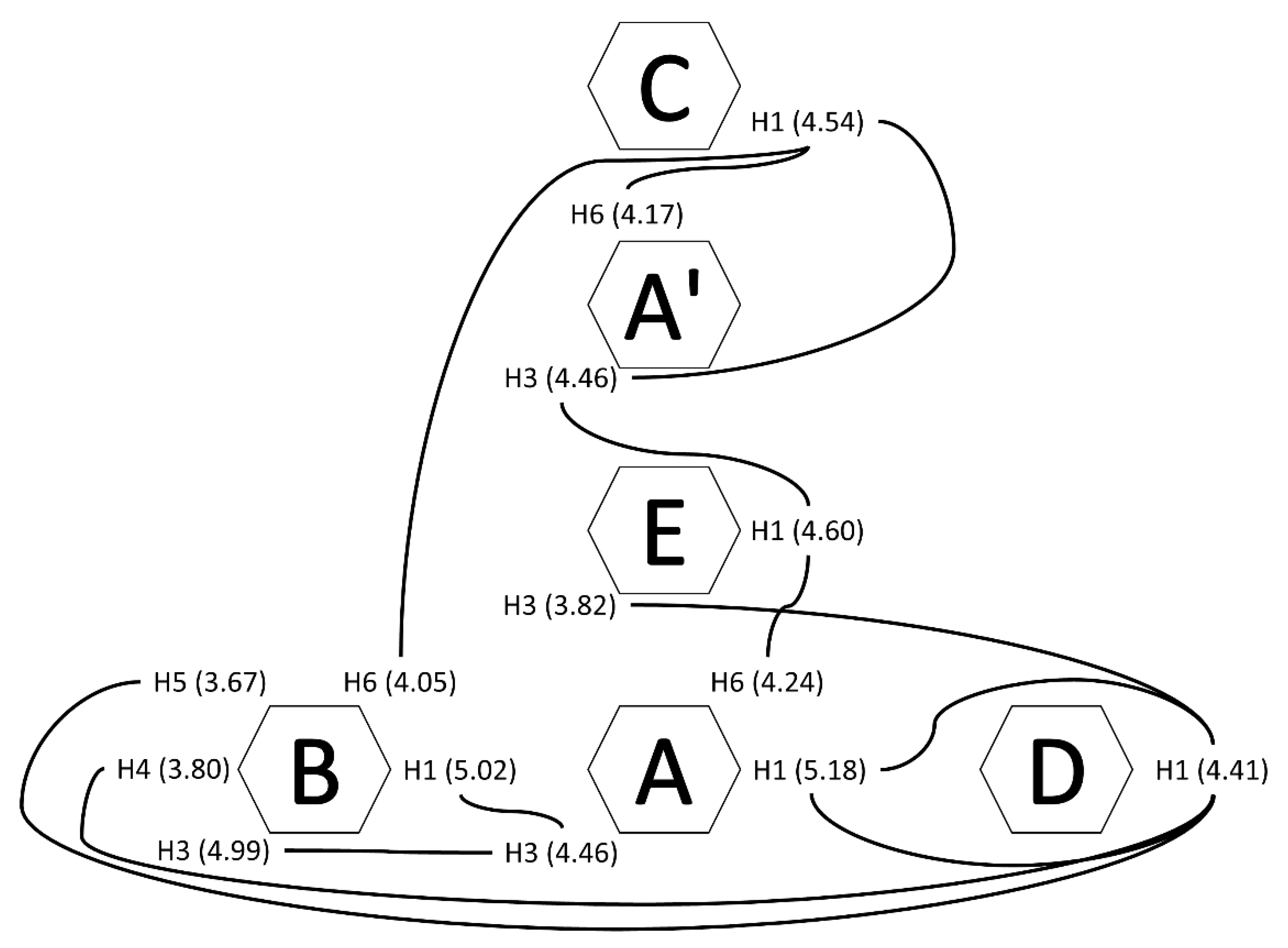
| A | A’ | B | C | D | E | |
|---|---|---|---|---|---|---|
| H1/C1 | 5.18/108.83 | 5.16/108.83 | 5.02/107.87 | 4.54/101.1 | 4.41/103.3 | 4.60/101.42 |
| H2/C2 | 3.84/76.61 | 4.68/- | 4.72/- | 3.74/55.02 | 3.31 */73.07 | 3.91 */52.44 |
| H3/C3 | 4.46 */68.18 | 4.06/72.74 | 4.99/78.22 | 3.65 */79.19 | 3.45/75.03 | 3.82/69.52 |
| H4/C4 | 3.88 */- | 3.88 */- | 3.80 */- | 3.22/82.09 | 3.63 */79.19 | 3.70/- |
| H5/C5 | 4.17 */78.87 | 4.15 */81.44 | 3.67 */79.19 | 3.48 | 3.76 */- | 3.47/- |
| H6/C6 | 4.24 */67.4 | 4.17 */67.2 | 4.05 */67.6 | - | 4.27 */66.8 | - |
| Ac | 1.99 | 1.99 | 1.97 | 2.00 | 2.04 |
| C | E | 1 GlcNAc [49] | 2 GlcNAc [50] | 3 GalNAc [51] | |
|---|---|---|---|---|---|
| H1 | 4.54 | 4.60 | 4.55 (4.71 terminal) | 4.56 | |
| H2 | 3.74 | 3.91 | 3.84 | 4.04 | |
| H3 | 3.65 | 3.82 | 3.71 | 3.88 | |
| H4 | 3.22 | 3.70 | 3.52 | 4.13 | |
| H5 | 3.48 | 3.47 | 3.48 | 3.74 | |
| H6 | 3.78/82 | ||||
| C1 | 101.1 | 101.42 | 103.32 (97.6 terminal) | 103.52 | 103.8 |
| C2 | 55.02 | 52.44 | 57.12 | 57.11 | 54.5 |
| C3 | 79.19 | 69.52 | 85.32 | 85.38 | 78.6 |
| C4 | 82.09 | 71.24 | 71.25 | 79.4 | |
| C5 | 78.13 | 78.5 | 77.5 | ||
| C6 | 63.37 |
| A | A’ | B | D | IdoA 2S | IdoA | SG1 | SG2 | SG3 | |
|---|---|---|---|---|---|---|---|---|---|
| H1 | 5.18 | 5.16 | 5.02 | 4.41 | 5.14–5.16 | 4.88–4.90 | 5.18/4.65/4.47 | 5.15/5.01/4.47 | |
| H2 | 3.84 | 4.68 | 4.72 | 3.31 | 4.14–4.17 | 3.51–3.54 | 3.87/3.83/3.31 | ||
| H3 | 4.46 | 4.06 | 4.99 | 3.45 | 4.20–4.32 | 3.90–4.06 | 4.08/3.99/3.57 | ||
| H4 | 3.88 | 3.88 | 3.80 | 3.63 | 4.03–4.06 | 4.06–4.10 | 4.81/4.76/3.78 | ||
| H5 | 4.17 | 4.15 | 3.67 | 3.76 | 4.83–4.85 | 4.70–4.80 | 4.11/4.03/4.08 | ||
| H6 | 4.24 | 4.17 | 4.05 | 4.27 | - | - | 3.70/3.75/3.39–4.11 | ||
| C1 | 108.8 | 108.8 | 107.9 | 103.3 | 101.2–106 | 105.3–105.7 | 101.3 | 102.7 | 100.7/101.9/104.6 |
| C2 | 76.6 | - | - | 73.1 | 73.8–75.2 | 71.6–72.0 | 68.6 | 69.9 | 71.9/71.7/76.0 |
| C3 | 68.2 | 72.7 | 78.2 | 75.0 | 69.0–71.0 | 73.2–73.7 | 77.7 | 78.8 | 71.5/72.4/77.1 |
| C4 | - | - | - | 79.2 | 79.2–80.7 | 82.3–82.7 | 77.1 | 78.8 | 80.9/80.8/79.6 |
| C5 | 78.9 | 81.4 | 79.2 | - | 68.0–69.7 | 72.0–72.4 | 72.6 | 73.9 | 73.3/73.6/73.4 |
| C6 | 67.4 | 67.2 | 67.6 | 66.8 | - | - | 60.7 | 62.2 | 64.0/64.4/66.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valcarcel, J.; Vázquez, J.A.; Varela, U.R.; Reis, R.L.; Novoa-Carballal, R. Isolation and Characterization of Polysaccharides from the Ascidian Styela clava. Polymers 2022, 14, 16. https://doi.org/10.3390/polym14010016
Valcarcel J, Vázquez JA, Varela UR, Reis RL, Novoa-Carballal R. Isolation and Characterization of Polysaccharides from the Ascidian Styela clava. Polymers. 2022; 14(1):16. https://doi.org/10.3390/polym14010016
Chicago/Turabian StyleValcarcel, Jesus, José Antonio Vázquez, Uxía R. Varela, Rui L. Reis, and Ramon Novoa-Carballal. 2022. "Isolation and Characterization of Polysaccharides from the Ascidian Styela clava" Polymers 14, no. 1: 16. https://doi.org/10.3390/polym14010016
APA StyleValcarcel, J., Vázquez, J. A., Varela, U. R., Reis, R. L., & Novoa-Carballal, R. (2022). Isolation and Characterization of Polysaccharides from the Ascidian Styela clava. Polymers, 14(1), 16. https://doi.org/10.3390/polym14010016