Biological Effects of Polyrotaxane Surfaces on Cellular Responses of Fibroblast, Preosteoblast and Preadipocyte Cell Lines

 , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
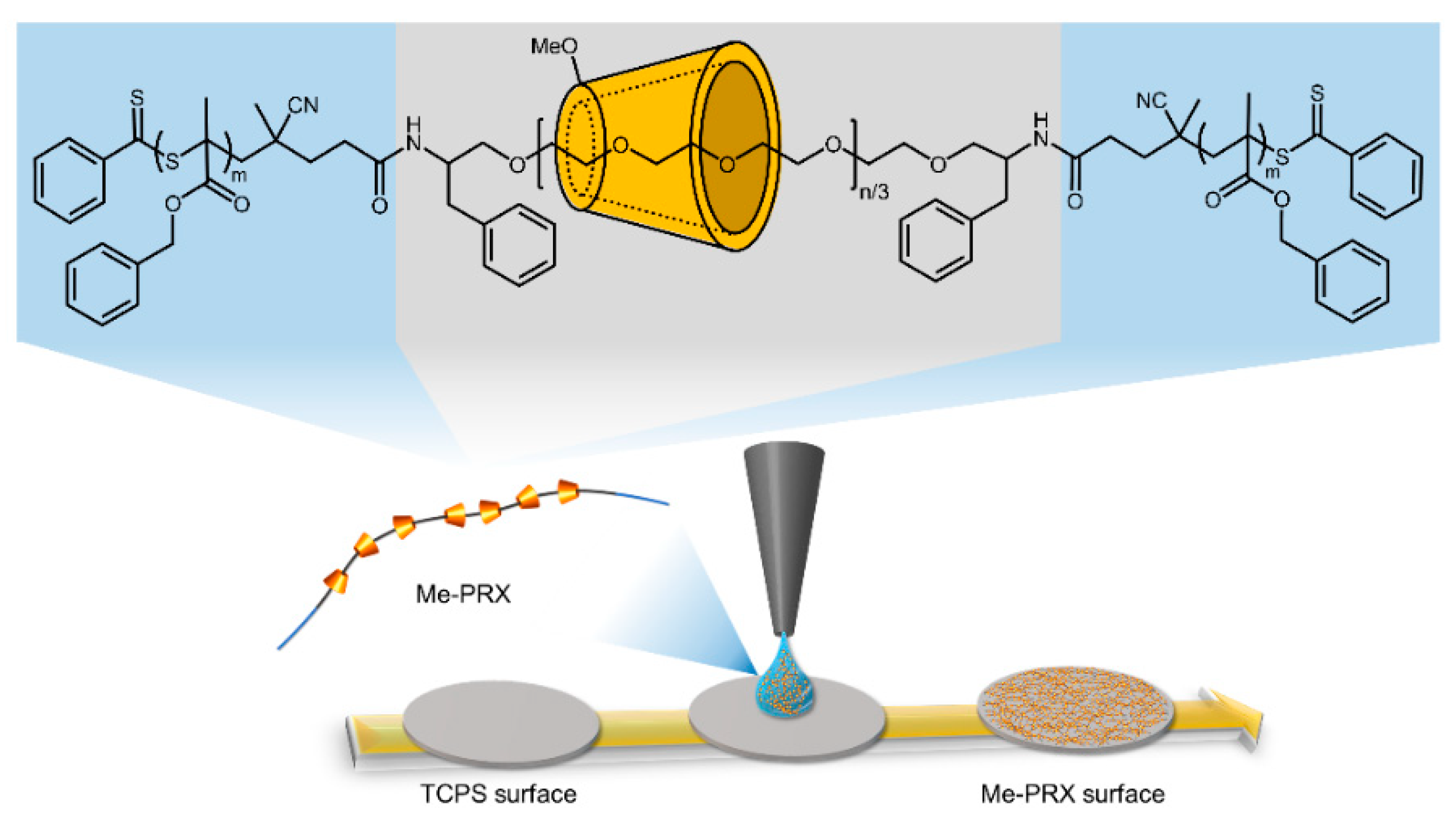
2.2. Preparation of the Me-PRX Surfaces
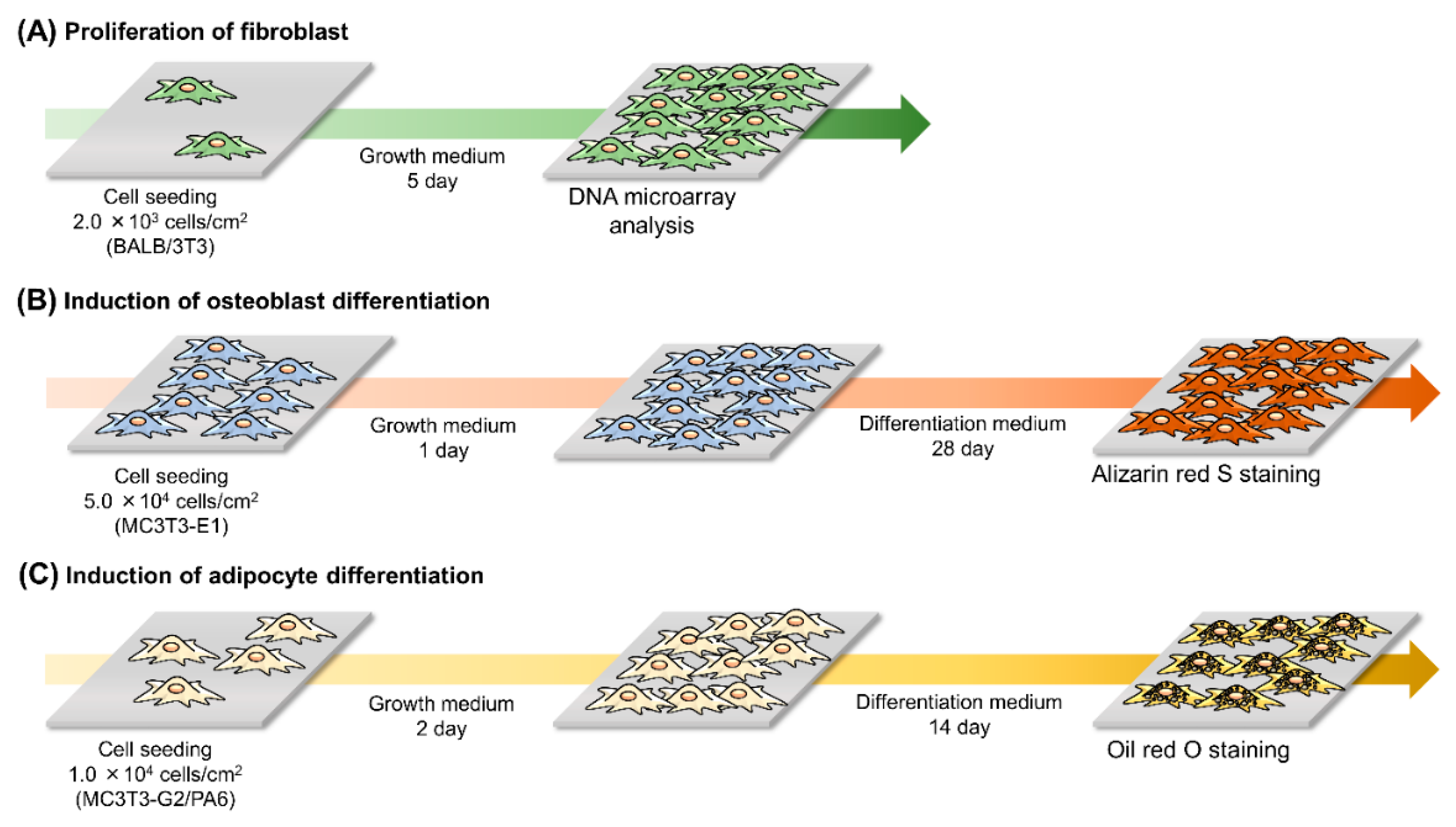
2.3. Morphology and Proliferation of the Cells
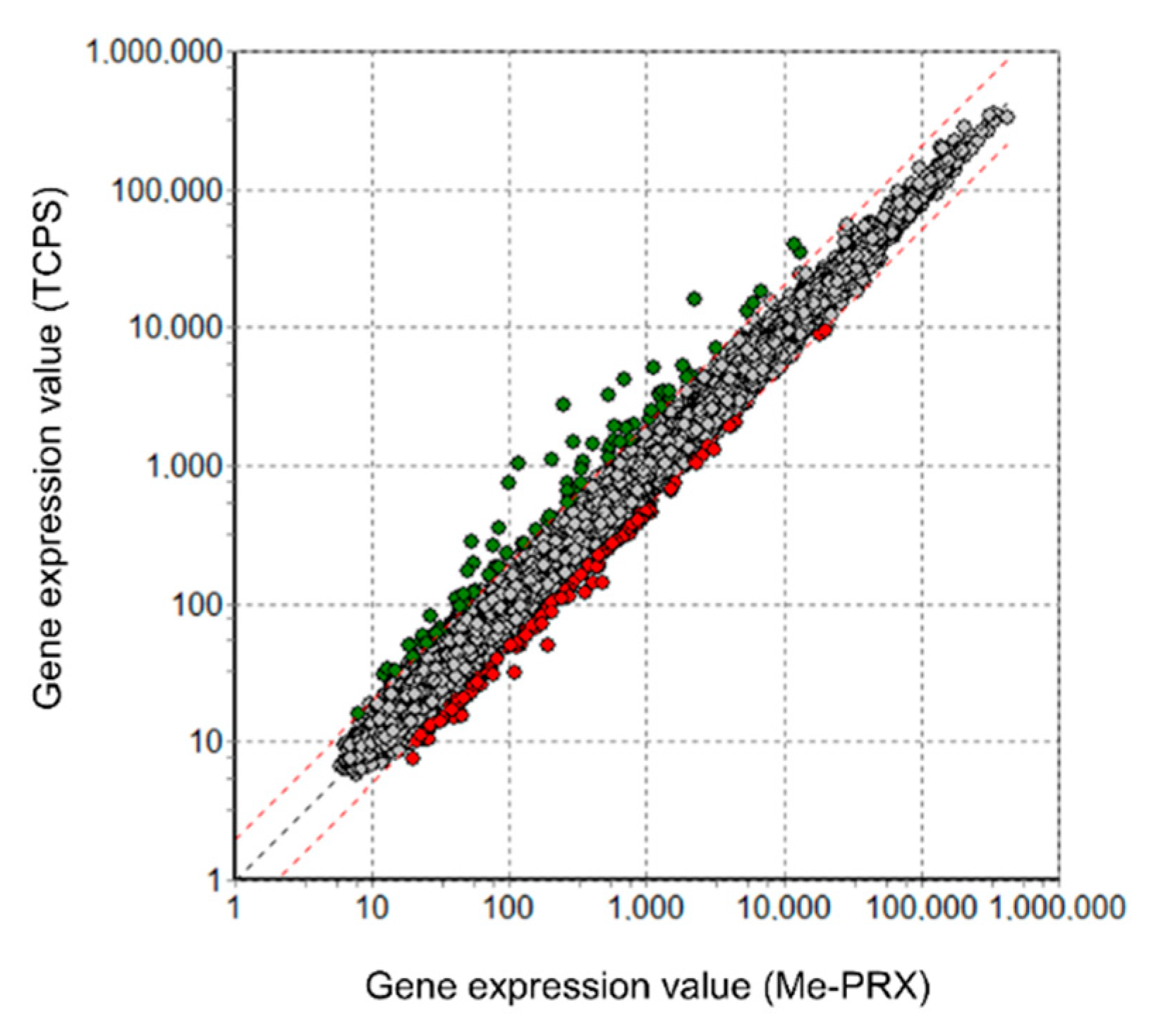
2.4. DNA Microarray Analysis
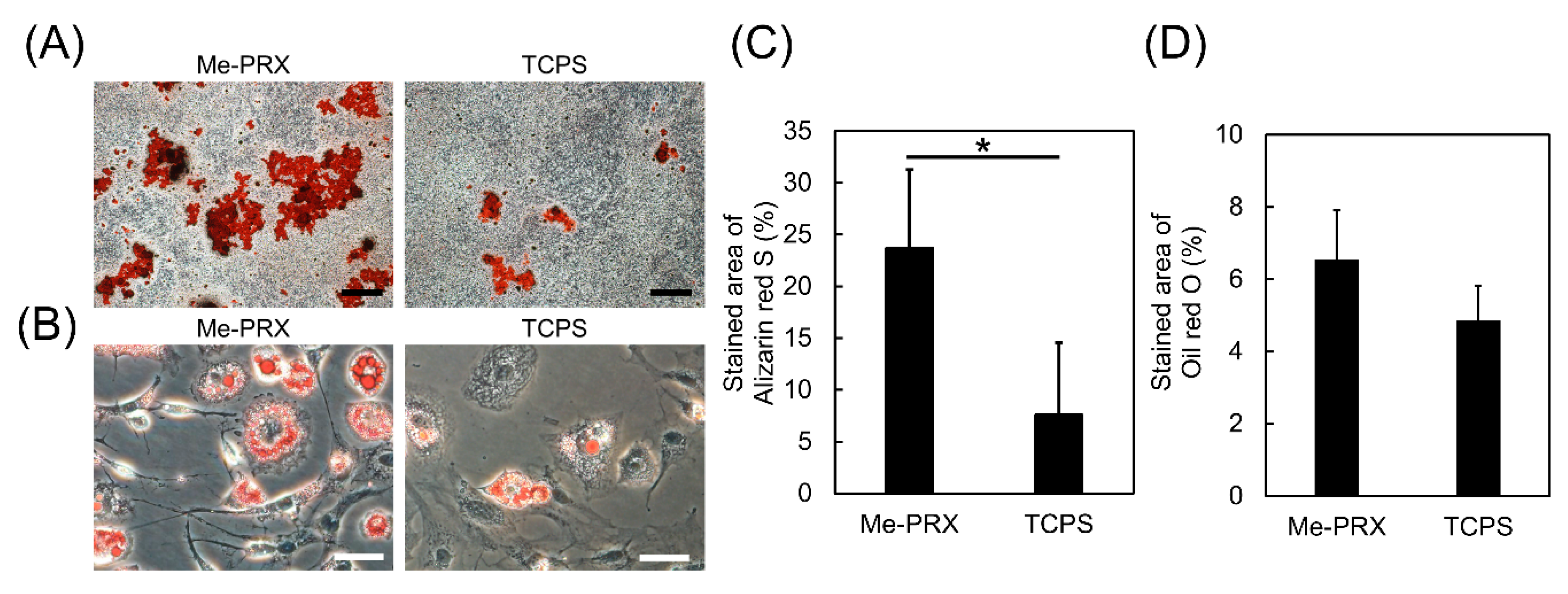
2.5. Differentiation of the Cells
2.6. Statistical Analysis
3. Results and Discussion
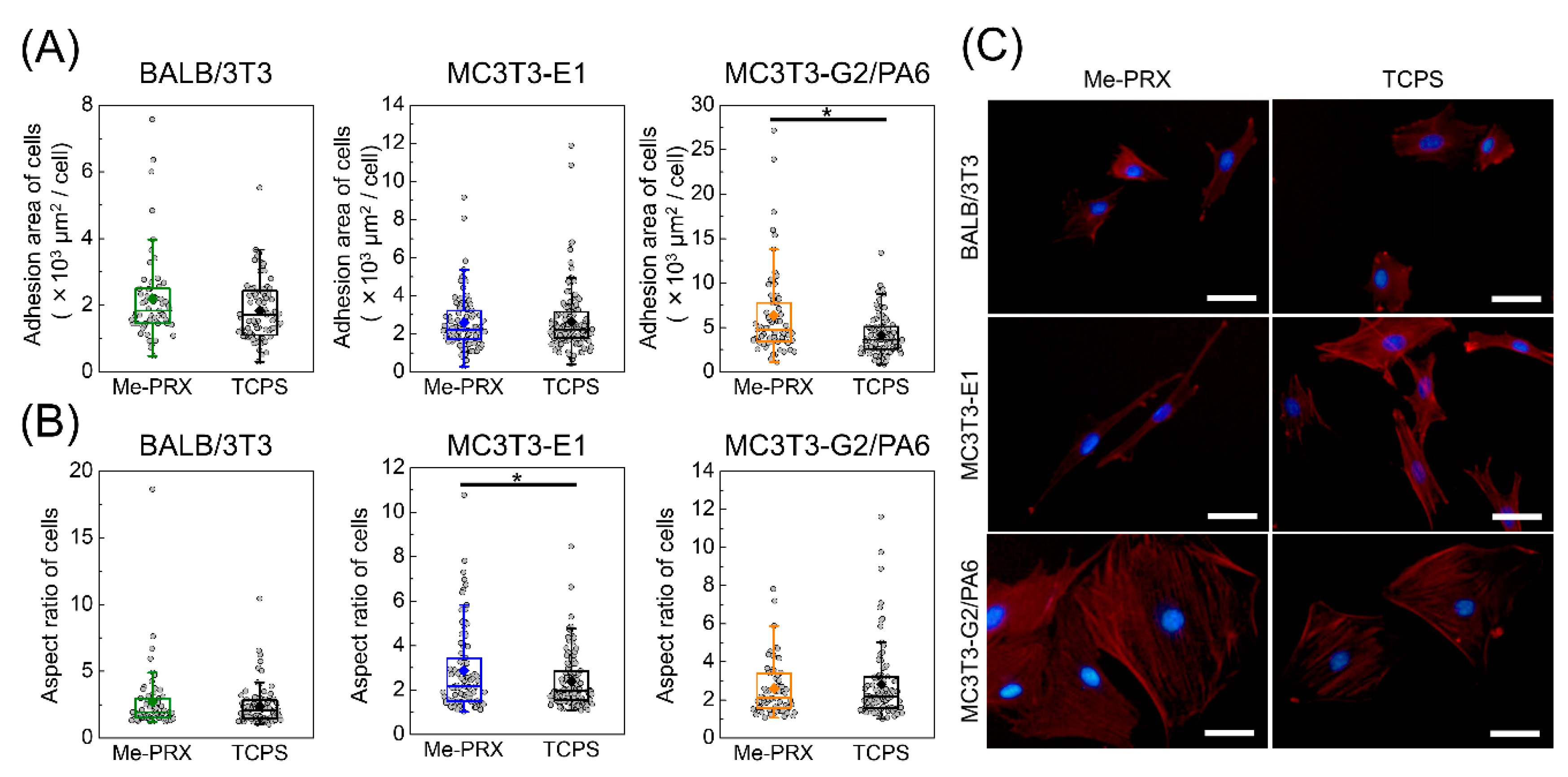
3.1. Morphology of the Cells
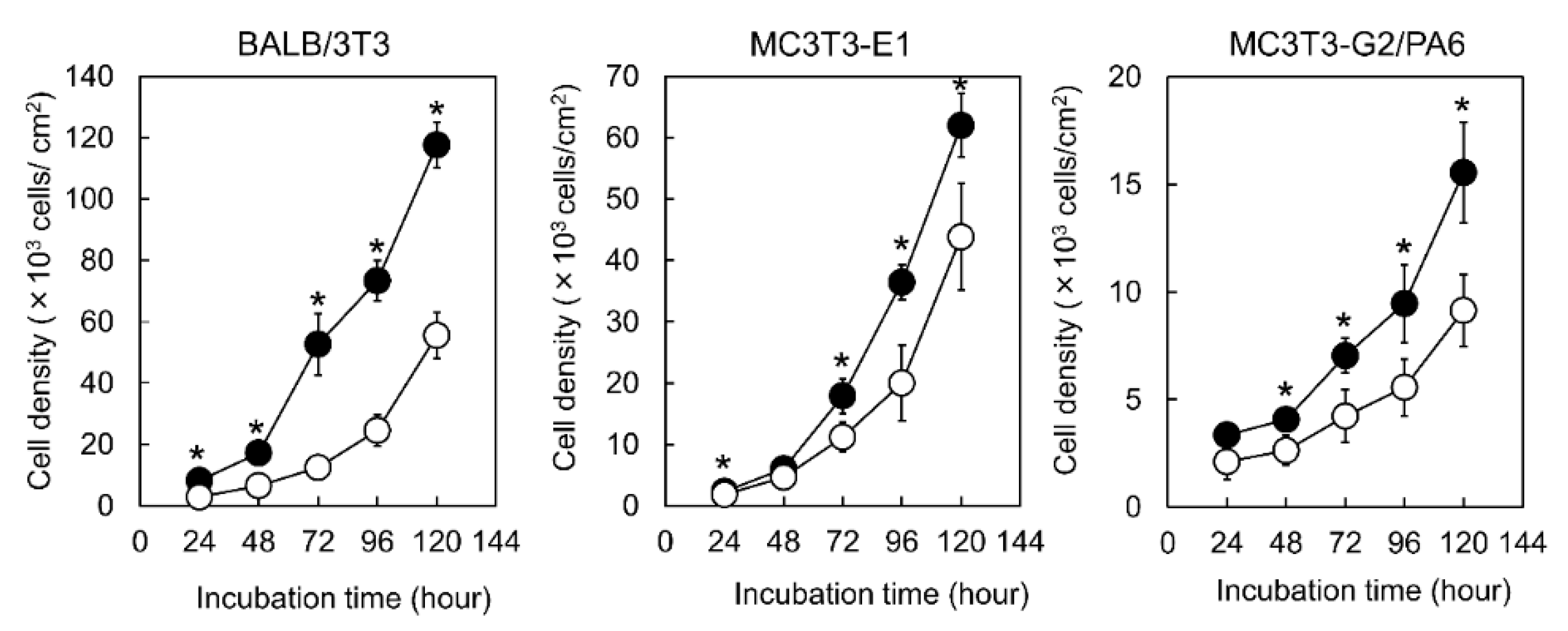
3.2. Proliferation of the Cells
3.3. Differentiation of the Cells
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Yang, Y.; Wang, K.; Gu, X.; Leong, K.W. Biophysical Regulation of Cell Behavior—Cross Talk between Substrate Stiffness and Nanotopography. Engineering 2017, 3, 36–54. [Google Scholar] [CrossRef]
- Engler, A.; Bacakova, L.; Newman, C.; Hategan, A.; Griffin, M.; Discher, D. Substrate compliance versus ligand density in cell on gel responses. Biophys. J. 2004, 86, 617–628. [Google Scholar] [CrossRef]
- Tsimbouri, P.M.; McMurray, R.J.; Burgess, K.V.; Alakpa, E.V.; Reynolds, P.M.; Murawski, K.; Kingham, E.; Oreffo, R.O.; Gadegaard, N.; Dalby, M.J. Using Nanotopography and metabolimics to identify biochemical effectors of multipotency. ACS Nano 2012, 6, 10239–10249. [Google Scholar] [CrossRef] [PubMed]
- McMurray, R.J.; Gadegaard, N.; Tshimbouri, P.M.; Burgess, K.V.; McNamara, L.E.; Tare, R.; Murawski, K.; Kingham, E.; Oreffo, R.O.; Dalby, M.J. Nanoscale surfaces for the long-term maintenance of mesenchymal stem cell phenotype and multipotency. Nat. Mater. 2011, 10, 637–644. [Google Scholar] [CrossRef] [PubMed]
- Engler, A.J.; Sen, S.; Sweeney, H.L.; Discher, D.E. Matrix elasticity directs stem cell lineage specification. Cell 2006, 126, 677–689. [Google Scholar] [CrossRef] [PubMed]
- Seo, J.H.; Yui, N. The effect of molecular mobility of supramolecular polymer surfaces on fibroblast adhesion. Biomaterials 2013, 34, 55–63. [Google Scholar] [CrossRef]
- Hyodo, K.; Arisaka, Y.; Yamaguchi, S.; Yoda, T.; Yui, N. Stimulation of Microvascular Networks on Sulfonated Polyrotaxane Surfaces with Immobilized Vascular Endothelial Growth Factor. Macromol. Biosci. 2019, 19, 1800346. [Google Scholar] [CrossRef]
- Arisaka, Y.; Yui, N. Tethered bone morphogenetic protein-2 onto sulfonated-polyrotaxane based surfaces promotes osteogenic differentiation of MC3T3-E1 cells. J. Biomater. Sci. Polym. Ed. 2017, 28, 974–998. [Google Scholar] [CrossRef]
- Tamura, A.; Yui, N. Threaded macromolecules as a versatile framework for biomaterials. Chem. Commun. 2014, 50, 13433–13446. [Google Scholar] [CrossRef]
- Harada, A.; Hashidzume, A.; Yamaguchi, H.; Takashima, Y. Polymeric rotaxanes. Chem. Rev. 2009, 109, 5974–6023. [Google Scholar] [CrossRef]
- Seo, J.H.; Kakinoki, S.; Inoue, Y.; Nam, K.; Yamaoka, T.; Ishihara, K.; Kishida, A.; Yui, N. The significance of hydrated surface molecular mobility in the control of the morphology of adhering fibroblasts. Biomaterials 2013, 34, 3206–3214. [Google Scholar] [CrossRef] [PubMed]
- Seo, J.H.; Kakinoki, S.; Yamaoka, T.; Yui, N. Directing stem cell differentiation by changing the molecular mobility of supramolecular surfaces. Adv. Healthc. Mater. 2015, 4, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Wohlrab, S.; Müller, S.; Schmidt, A.; Neubauer, S.; Kessler, H.; Leal-Egaña, A.; Scheibel, T. Cell adhesion and proliferation on RGD-modified recombinant spider silk proteins. Biomaterials 2012, 33, 6650–6659. [Google Scholar] [CrossRef]
- Khatiwala, C.B.; Peyton, S.R.; Putnam, A.J. Intrinsic mechanical properties of the extracellular matrix affect the behavior of pre-osteoblastic MC3T3-E1 cells. Am. J. Physiol. Cell Physiol. 2006, 290, 1640–1650. [Google Scholar] [CrossRef] [PubMed]
- Kramer, A.H.; Joos-Vandewalle, J.; Edkins, A.L.; Frost, C.L.; Princloo, E. Real-time monitoring of 3T3-L1 preadipocyte differentiation using a commercially available electric cell-substrate impedance sensor system. Biochem. Biophys. Res. Commun. 2014, 443, 1245–1250. [Google Scholar] [CrossRef] [PubMed]
- Sekiya-Aoyama, R.; Arisaka, Y.; Yui, N. Mobility Tuning of Polyrotaxane Surfaces to Stimulate Myocyte Differentiation. Macromol. Biosci. 2020, 20, 1900424. [Google Scholar] [CrossRef] [PubMed]
- Hoon, J.L.; Tan, M.H.; Koh, C.G. The Regulation of Cellular Responses to Mechanical cues by Rho GTPases. Cells 2016, 5, 17. [Google Scholar] [CrossRef]
- Yao, X.; Peng, R.; Ding, J. Effects of aspect ratios of stem cells on lineage commitments with and without induction media. Biomaterials 2013, 34, 930–939. [Google Scholar] [CrossRef]
- McBeath, R.; Pirone, D.M.; Nelson, C.M.; Bhadriraju, K.; Chen, C.S. Cell shape, cytoskeletal tension, and RhoA regulate stem cell lineage commitment. Dev. Cell. 2004, 6, 483–495. [Google Scholar] [CrossRef]
- Wozniak, M.A.; Chen, C.S. Mechanotransduction in development: A growing role for contractility. Nat. Rev. Mol. Cell Biol. 2009, 10, 34–43. [Google Scholar] [CrossRef]
- Chen, Y.; Xie, Y.; Xu, L.; Zhan, S.; Xiao, Y.; Gao, Y.; Wu, B.; Ge, W. Protein content and functional characteristics of serum-purified exosomes from patients with colorectal cancer revealed by quantitative proteomics. Int. J. Cancer 2017, 140, 900–913. [Google Scholar] [CrossRef] [PubMed]
- Young, J.L.; Krethmer, K.; Ondeck, M.G.; Zambon, A.C.; Engler, A.J. Mechanosensitive kinases regulate stiffness-induced cardiomyocyte maturation. Sci. Rep. 2014, 4, 6425. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Liu, W.Z.; Liu, T.; Feng, X.; Yang, N.; Zhou, H.F. Signaling pathway of MAPK/ERK in cell proliferation, differentiation, migration, senescence and apoptosis. J. Recept. Signal Transduct. Res. 2015, 35, 600–604. [Google Scholar] [CrossRef] [PubMed]
- Kilian, K.A.; Bugarija, B.; Lahn, B.T.; Mrksich, M. Geometric cues for directing the differentiation of mesenchymal stem cells. Proc. Natl. Acad. Sci. USA 2010, 107, 4872–4877. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name of Pathway | Changed Genes | Total Genes | Z Score | p-Value | Gene Symbols |
|---|---|---|---|---|---|
| Primary Focal Segmental Glomerulosclerosis | 4 | 72 | 5.803 | 0.0006 | Itga3, Itgb3, kirrel3, Tgfb1 |
| ACE Inhibitor Pathway | 2 | 8 | 9.435 | 0.0012 | Bdkrb2, Kng1 |
| Integrin-mediated Cell Adhesion | 4 | 100 | 4.718 | 0.002 | Itga3, Itga6, Itgb3, Map2k6 |
| Toll Like Receptor Signaling | 2 | 33 | 4.325 | 0.0147 | Map2k6, Traf3 |
| Spinal Cord Injury | 3 | 97 | 3.428 | 0.0156 | Selp, Gdnf, Tgfb1 |
| Focal Adhesion | 4 | 184 | 3.021 | 0.0167 | Itga3, Itga6, Itgb3, Map2k6 |
| Endochondral Ossification | 2 | 62 | 2.884 | 0.0456 | Tgfb1, Sox5 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Masuda, H.; Arisaka, Y.; Sekiya-Aoyama, R.; Yoda, T.; Yui, N. Biological Effects of Polyrotaxane Surfaces on Cellular Responses of Fibroblast, Preosteoblast and Preadipocyte Cell Lines. Polymers 2020, 12, 924. https://doi.org/10.3390/polym12040924
Masuda H, Arisaka Y, Sekiya-Aoyama R, Yoda T, Yui N. Biological Effects of Polyrotaxane Surfaces on Cellular Responses of Fibroblast, Preosteoblast and Preadipocyte Cell Lines. Polymers. 2020; 12(4):924. https://doi.org/10.3390/polym12040924
Chicago/Turabian StyleMasuda, Hiroki, Yoshinori Arisaka, Ruriko Sekiya-Aoyama, Tetsuya Yoda, and Nobuhiko Yui. 2020. "Biological Effects of Polyrotaxane Surfaces on Cellular Responses of Fibroblast, Preosteoblast and Preadipocyte Cell Lines" Polymers 12, no. 4: 924. https://doi.org/10.3390/polym12040924
APA StyleMasuda, H., Arisaka, Y., Sekiya-Aoyama, R., Yoda, T., & Yui, N. (2020). Biological Effects of Polyrotaxane Surfaces on Cellular Responses of Fibroblast, Preosteoblast and Preadipocyte Cell Lines. Polymers, 12(4), 924. https://doi.org/10.3390/polym12040924