
Purification and Crystallographic Analysis of a Novel Cold-Active Esterase (HaEst1) from Halocynthiibacter arcticus

 , , ,
, , ,  , ,
, ,  , and
, and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals and Columns
2.2. Gene Cloning and Protein Purification
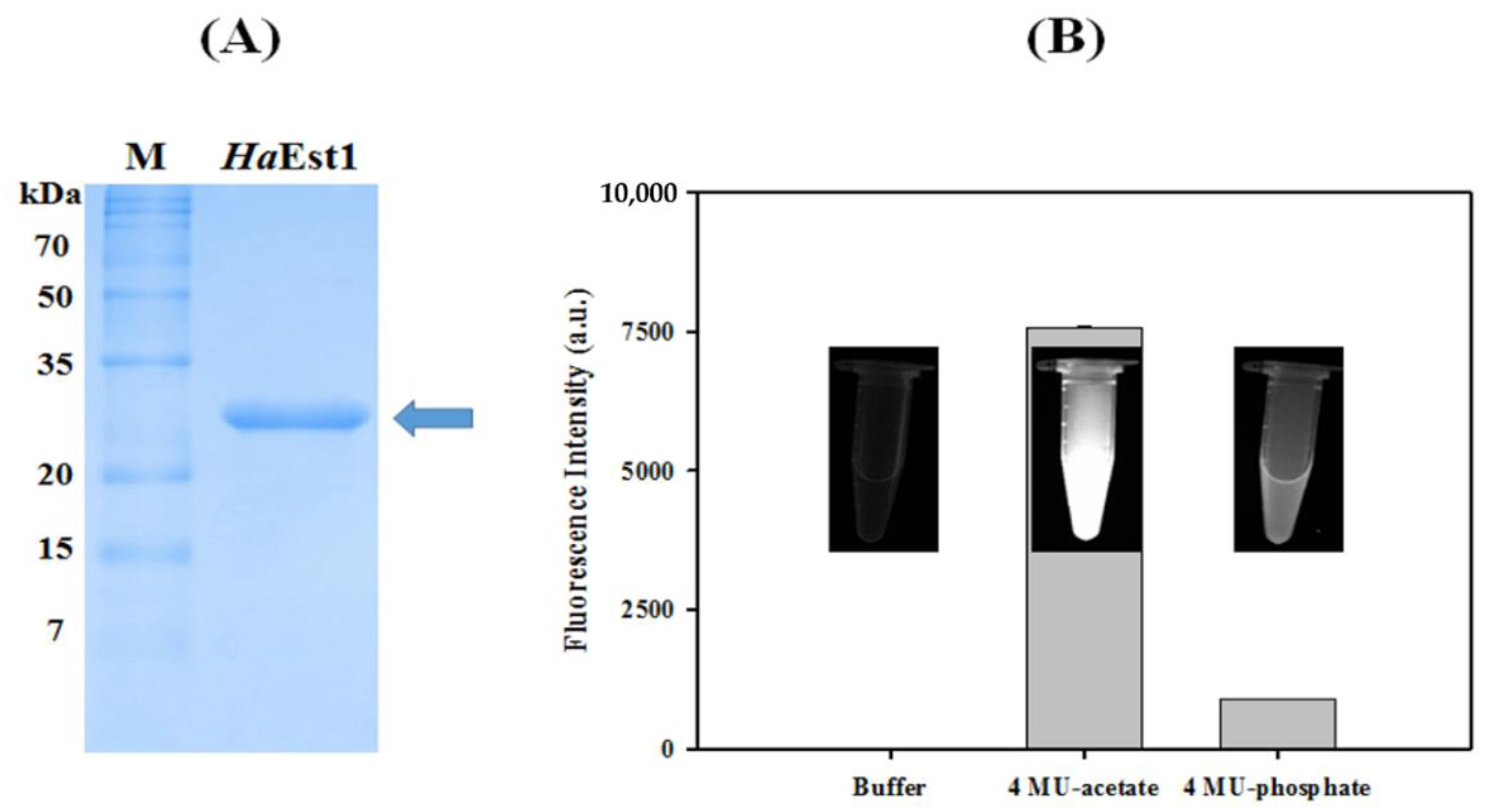
2.3. Activity Measurement of HaEst1
2.4. Crystallization Method
2.5. Data Collection and Processing
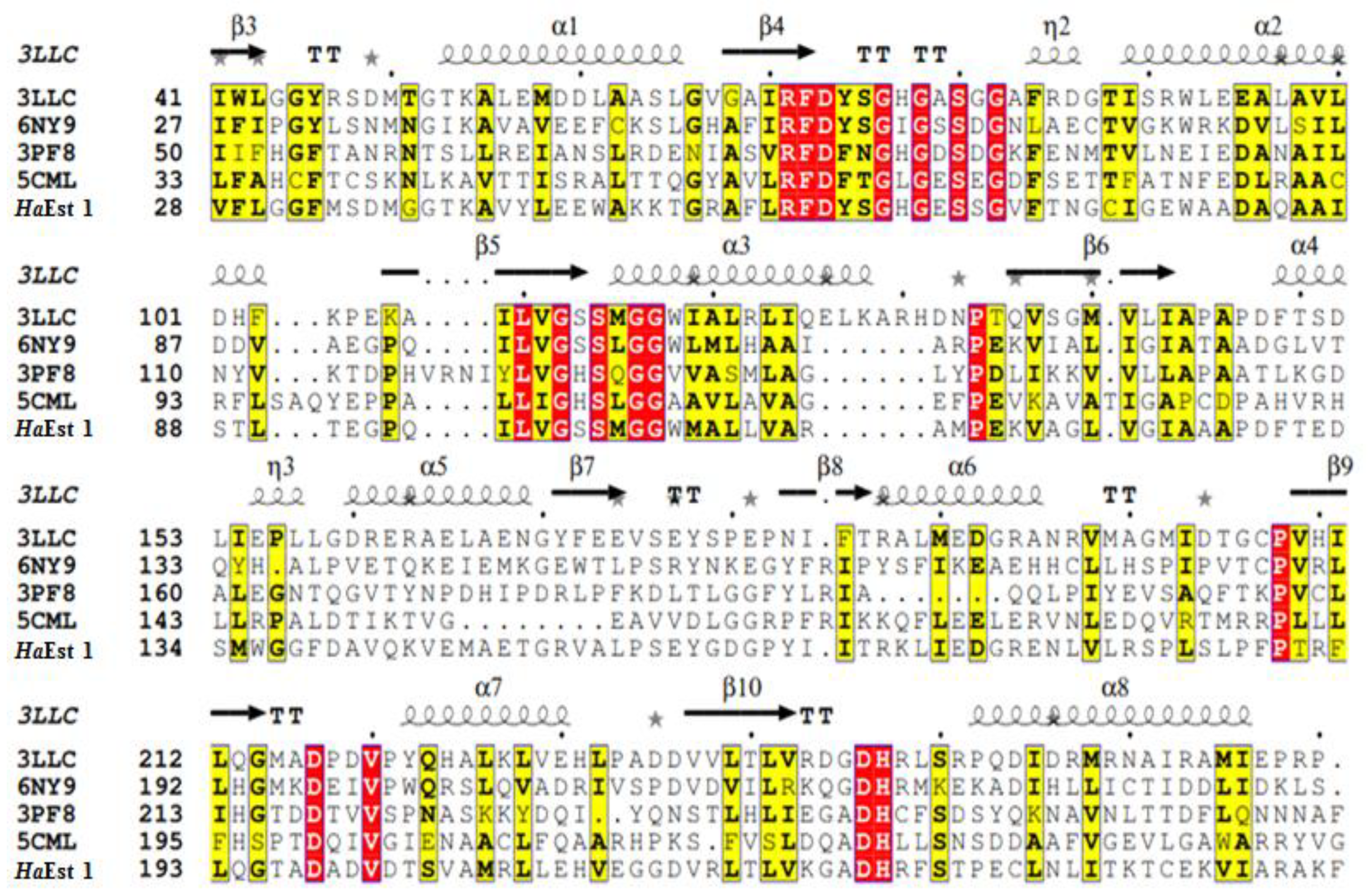
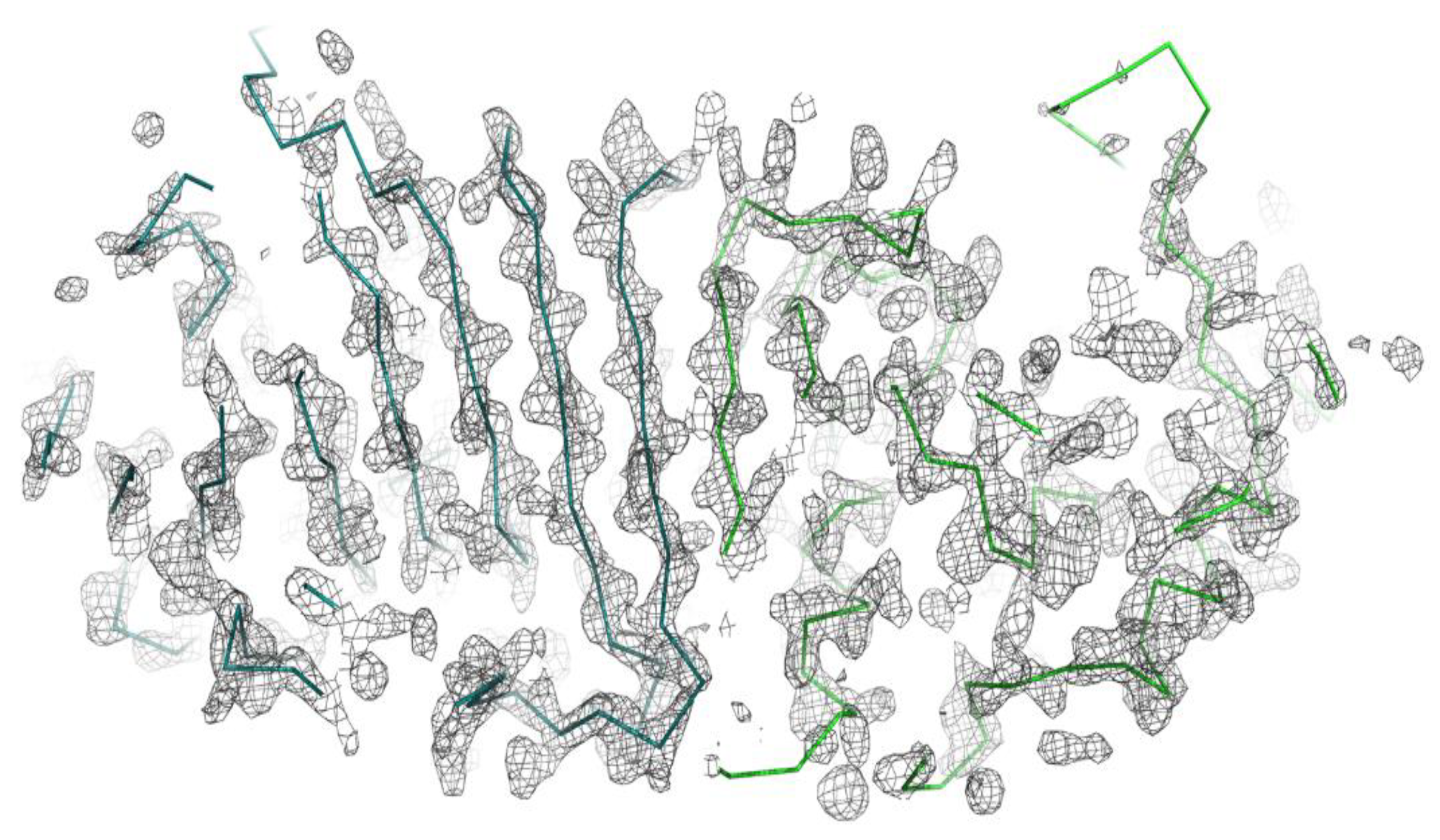
3. Results and Discussion
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Holmquist, M. Alpha/Beta-hydrolase fold enzymes: Structures, functions and mechanisms. Curr. Protein Pept. Sci. 2000, 1, 209–235. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.; Gupta, N.; Rathi, P. Bacterial lipases: An overview of production, purification and biochemical properties. Appl. Microbiol. Biotechnol. 2004, 64, 763–781. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, P.; Bhatt, K.; Huang, Y.; Lin, Z.; Chen, S. Esterase is a powerful tool for the biodegradation of pyrethroid insecticides. Chemosphere 2020, 244, 125507. [Google Scholar] [CrossRef]
- Sarmah, N.; Revathi, D.; Sheelu, G.; Yamuna-Rani, K.Y.; Sridhar, S.; Mehtab, V.; Sumana, C. Recent advances on sources and industrial applications of lipases. Biotechnol. Prog. 2018, 34, 5–28. [Google Scholar] [CrossRef]
- Chen, Y.; Black, D.S.; Reilly, P.J. Carboxylic ester hydrolases: Classification and database derived from their primary, secondary, and tertiary structures. Protein Sci. 2016, 25, 1942–1953. [Google Scholar] [CrossRef]
- Oh, C.; Kim, T.D.; Kim, K.K. Carboxylic ester hydrolases in bacteria: Active site, structure, function and application. Crystals 2019, 9, 597. [Google Scholar] [CrossRef]
- Santiago, M.; Ramírez-Sarmiento, C.A.; Zamora, R.A.; Parra, L.P. Discovery, molecular mechanisms, and industrial applications of cold-active Enzymes. Front. Microbiol. 2016, 7, 1408. [Google Scholar] [CrossRef]
- Siddiqui, K.S. Some like it hot, some like it cold: Temperature dependent biotechnological applications and improvements in extremophilic enzymes. Biotechnol. Adv. 2015, 33, 1912–1922. [Google Scholar] [CrossRef]
- Dumorné, K.; Córdova, D.C.; Astorga-Eló, M.; Renganathan, P. Extremozymes: A potential source for industrial applications. J. Microbiol. Biotechnol. 2017, 27, 649–659. [Google Scholar] [CrossRef]
- Park, S.-H.; Yoo, W.; Lee, C.W.; Jeong, C.S.; Shin, S.C.; Kim, H.-W.; Park, H.; Kim, K.K.; Kim, T.D.; Lee, J.H. Crystal structure and functional characterization of a cold-active acetyl xylan esterase (PbAcE) from psychrophilic soil microbe Paenibacillus sp. PLoS ONE 2018, 13, e0206260. [Google Scholar] [CrossRef] [PubMed]
- Truongvan, N.; Jang, S.H.; Lee, C. Flexibility and stability trade-off in active site of cold-adapted Pseudomonas mandelii esterase EstK. Biochemistry 2016, 55, 3542–3549. [Google Scholar] [CrossRef] [PubMed]
- Esteban-Torres, M.; Mancheño, J.M.; Rivas, B.D.L.; Muñoz, R. Characterization of a cold-active esterase from Lactobacillus plantarum suitable for food fermentations. J. Agric. Food Chem. 2014, 62, 5126–5132. [Google Scholar] [CrossRef]
- Noby, N.; Hussein, A.; Saeed, H.; Embaby, A.M. Recombinant cold -adapted halotolerant, organic solvent-stable esterase (estHIJ) from Bacillus halodurans. Anal. Biochem. 2020, 591, 113554. [Google Scholar] [CrossRef]
- Lee, Y.M.; Baek, K.; Lee, J.; Lee, H.K.; Park, H.; Shin, S.C. Complete genome sequence of Halocynthiibacter arcticus PAMC 20958(T) from an Arctic marine sediment sample. J. Biotechnol. 2016, 224, 12–13. [Google Scholar] [CrossRef]
- Baek, K.; Lee, Y.M.; Shin, S.C.; Hwang, K.; Hwang, C.Y.; Hong, S.G.; Lee, H.K. Halocynthiibacter arcticus sp. nov., isolated from Arctic marine sediment. Int. J. Syst. Evol. Microbiol. 2015, 65, 3861–3865. [Google Scholar] [CrossRef] [PubMed]
- Jeon, S.; Hwang, J.; Yoo, W.; Do, H.; Kim, H.-W.; Kim, K.K.; Lee, J.H.; Kim, T.D. Crystallization and preliminary x-ray diffraction study of a novel bacterial homologue of mammalian hormone-sensitive lipase (HaLip1) from Halocynthiibacter arcticus. Crystals 2020, 10, 963. [Google Scholar] [CrossRef]
- Le, L.T.H.L.; Yoo, W.; Lee, C.; Wang, Y.; Jeon, S.; Kim, K.K.; Lee, J.H.; Kim, T.D. Molecular characterization of a novel cold-active hormone-sensitive lipase (HaHSL) from Halocynthiibacter arcticus. Biomolecules 2019, 9, 704. [Google Scholar] [CrossRef] [PubMed]
- Jang, E.; Ryu, B.H.; Shim, H.W.; Ju, H.; Kim, D.W.; Kim, T.D. Adsorption of microbial esterases on Bacillus subtilis-templated cobalt oxide nanoparticles. Int. J. Biol. Macromol. 2014, 65, 188–192. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Le, L.T.H.L.; Yoo, W.; Lee, C.W.; Kim, K.K.; Lee, J.H.; Kim, T.D. Characterization, immobilization, and mutagenesis of a novel cold-active acetylesterase (EaAcE) from Exiguobacterium antarcticum B7. Int. J. Biol. Macromol. 2019, 136, 1042–1051. [Google Scholar] [CrossRef]
- Lee, C.W.; Yoo, W.; Park, S.H.; Le, L.T.H.L.; Jeong, C.S.; Ryu, B.H.; Shin, S.C.; Kim, H.W.; Park, H.; Kim, K.K.; et al. Structural and functional characterization of a novel cold-active S-formylglutathione hydrolase (SfSFGH) homolog from Shewanella frigidimarina, a psychrophilic bacterium. Microb. Cell Factories 2019, 18, 140. [Google Scholar] [CrossRef]
- Lee, C.W.; Lee, S.; Jeong, C.S.; Hwang, J.; Chang, J.H.; Choi, I.G.; Kim, T.D.; Park, H.; Kim, H.Y.; Lee, J.H. Structural insights into the psychrophilic germinal protease PaGPR and its autoinhibitory loop. J. Microbiol. 2020, 58, 772–779. [Google Scholar] [CrossRef] [PubMed]
- Jensen, M.V.; Horsfall, L.E.; Wardrope, C.; Togneri, P.D.; Marles-Wright, J.; Rosser, S.J. Characterisation of a new family of carboxyl esterases with an OsmC domain. PLoS ONE 2016, 11, e0166128. [Google Scholar] [CrossRef]
- Lai, K.K.; Stogios, P.J.; Vu, C.; Xu, X.; Cui, H.; Molloy, S.; Savchenko, A.; Yakunin, A.; Gonzalez, C.F. An inserted α/β subdomain shapes the catalytic pocket of Lactobacillus johnsonii cinnamoyl esterase. PLoS ONE 2011, 6, e23269. [Google Scholar] [CrossRef]
- Cao, Y.; Qiu, T.; Kathayat, R.S.; Azizi, S.A.; Thorne, A.K.; Ahn, D.; Fukata, Y.; Fukata, M.; Rice, P.A.; Dickinson, B.C. ABHD10 is an S-depalmitoylase affecting redox homeostasis through peroxiredoxin-5. Nat. Chem. Biol. 2019, 15, 1232–1240. [Google Scholar] [CrossRef] [PubMed]
- Matthews, B.W. Solvent content of protein crystals. J. Mol. Biol. 1968, 33, 491–497. [Google Scholar] [CrossRef]
- Vagin, A.; Teplyakov, A. Molecular replacement with MOLREP. Acta Crystallogr. Sect. D Biol. Crystallogr. 2010, 66, 22–25. [Google Scholar] [CrossRef] [PubMed]
- Murshudov, G.N.; Skubák, P.; Lebedev, A.A.; Pannu, N.S.; Steiner, R.A.; Nicholls, R.A.; Winn, M.D.; Long, F.; Vagin, A.A. REFMAC5 for the refinement of macromolecular crystal structures. Acta Crystallogr. Sect. D Biol. Crystallogr. 2011, 67, 355–367. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Data Collection | |
|---|---|
| Synchrotron source | PAL-Korea, beamline 5C |
| Wavelength (Å) | 0.9794 |
| Temperature (K) | 100 |
| Detector | Eiger X 9M |
| Crystal to detector distance (mm) | 400 |
| Rotation range per image (°) | 1 |
| Exposure time per image (s) | 1 |
| Rotation range (°) | 360 |
| Data Processing | |
| Crystal parameters | |
| Space group | P21 |
| No. of molecules in asymmetric unit | 2 |
| Unit-cell parameters (Å, °) | A = 35.69, b = 91.21, c = 79.15, α = 90, β = 96.91, γ = 90 |
| Data Statistics | |
| Resolution range (Å) | 50.00–2.10 (2.14–2.10) |
| Total no. of collected reflections | 186,387 (122) |
| No. of unique reflections | 29,269 (1435) |
| Completeness (%) | 99.0 (99.6) |
| Multiplicity | 6.4 (0.1) |
| ⟨I/σ(I)⟩ | 47.8 (7.67) |
| Rmerge (%) † | 8.1 (36.3) |
| Rmeas (%) ‡ | 8.8 (39.8) |
| CC(1/2) (%) | 99.1 (94.7) |
| Overall B-factor from Wilson plot (Å2) | 39.7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeon, S.; Hwang, J.; Yoo, W.; Chang, J.W.; Do, H.; Kim, H.-W.; Kim, K.K.; Lee, J.H.; Kim, T.D. Purification and Crystallographic Analysis of a Novel Cold-Active Esterase (HaEst1) from Halocynthiibacter arcticus. Crystals 2021, 11, 170. https://doi.org/10.3390/cryst11020170
Jeon S, Hwang J, Yoo W, Chang JW, Do H, Kim H-W, Kim KK, Lee JH, Kim TD. Purification and Crystallographic Analysis of a Novel Cold-Active Esterase (HaEst1) from Halocynthiibacter arcticus. Crystals. 2021; 11(2):170. https://doi.org/10.3390/cryst11020170
Chicago/Turabian StyleJeon, Sangeun, Jisub Hwang, Wanki Yoo, Joo Won Chang, Hackwon Do, Han-Woo Kim, Kyeong Kyu Kim, Jun Hyuck Lee, and T. Doohun Kim. 2021. "Purification and Crystallographic Analysis of a Novel Cold-Active Esterase (HaEst1) from Halocynthiibacter arcticus" Crystals 11, no. 2: 170. https://doi.org/10.3390/cryst11020170
APA StyleJeon, S., Hwang, J., Yoo, W., Chang, J. W., Do, H., Kim, H.-W., Kim, K. K., Lee, J. H., & Kim, T. D. (2021). Purification and Crystallographic Analysis of a Novel Cold-Active Esterase (HaEst1) from Halocynthiibacter arcticus. Crystals, 11(2), 170. https://doi.org/10.3390/cryst11020170