Crystallization of ApoA1 and ApoE4 Nanolipoprotein Particles and Initial XFEL-Based Structural Studies

 , ,
, ,  , , , , and
, , , , and 
Abstract
:1. Introduction
2. Materials and Methods
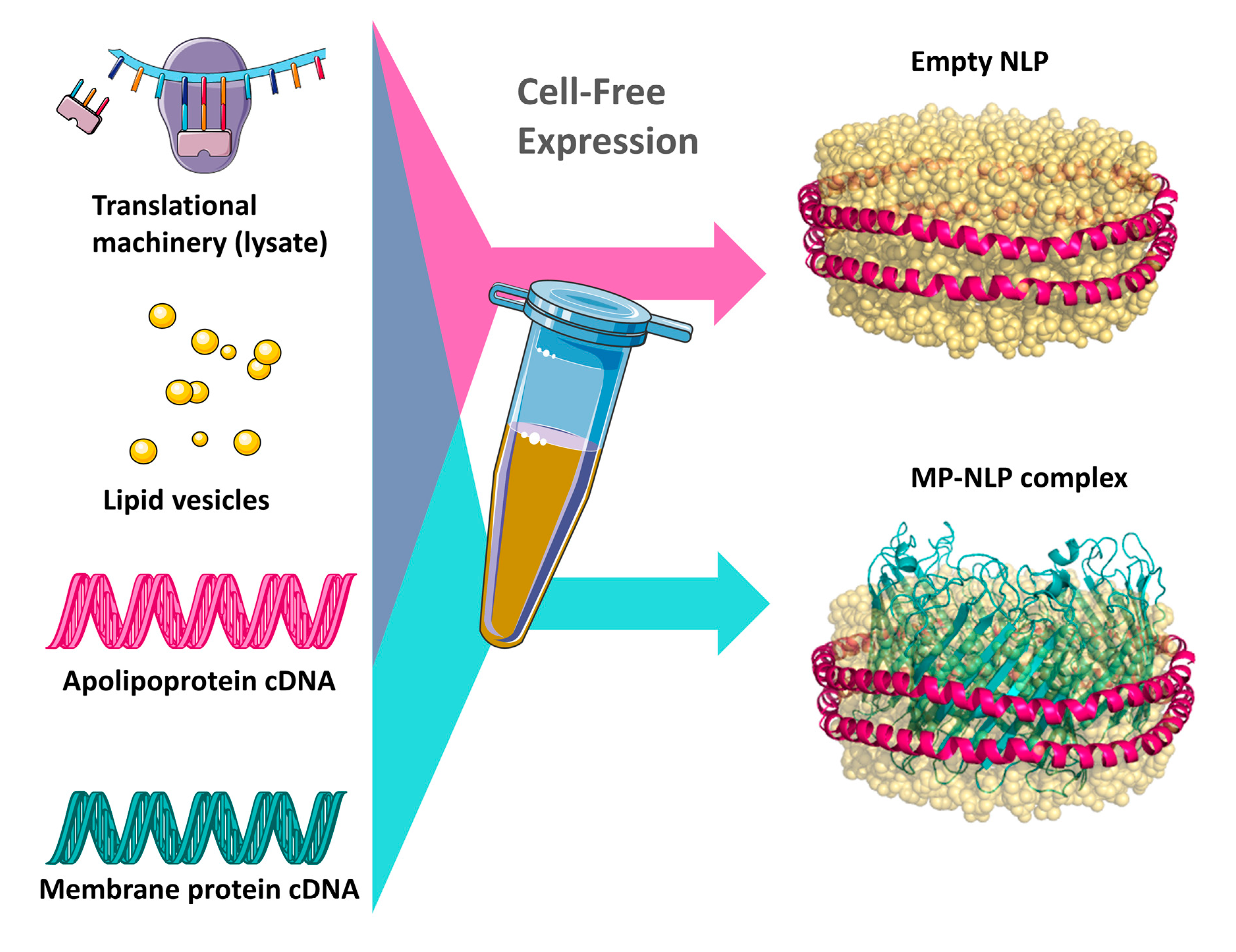
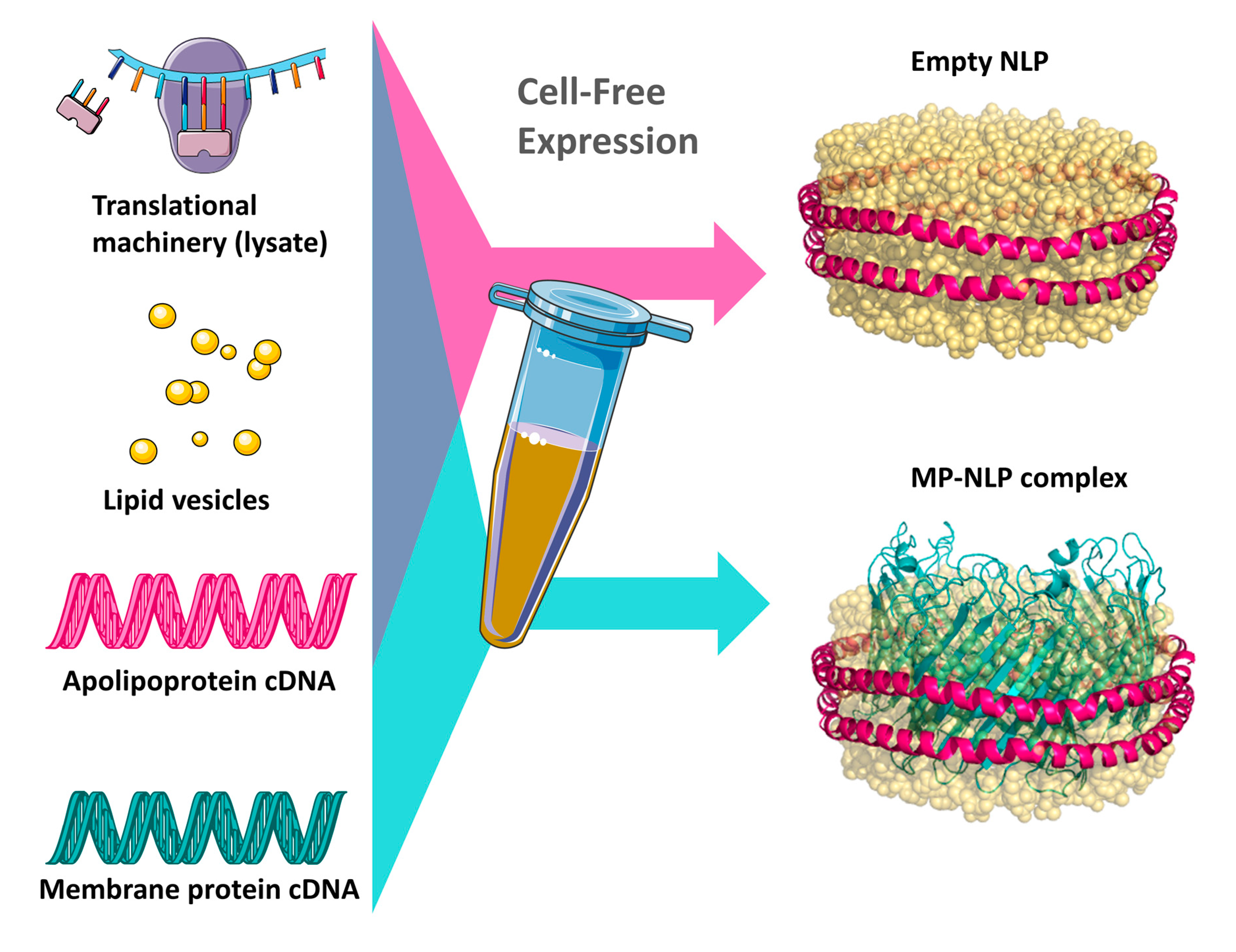
2.1. Cell-Free Expression, Preparation, and Characterization of ApoA1 and ApoE4 Nanolipoprotein Particles
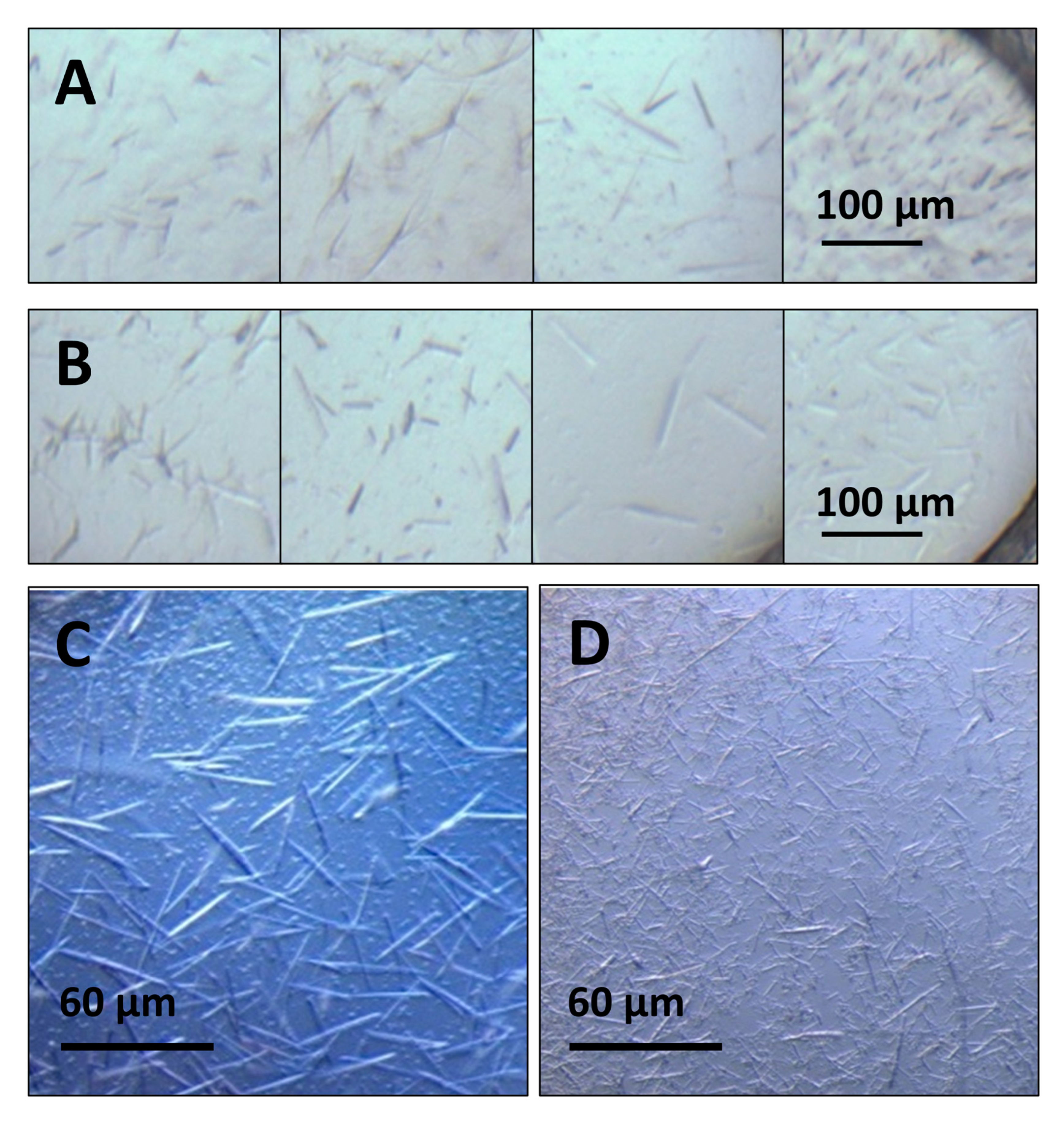
2.2. High-Throughput Crystallization Screening
2.3. Preparation of NLP Crystals for SFX Experiments
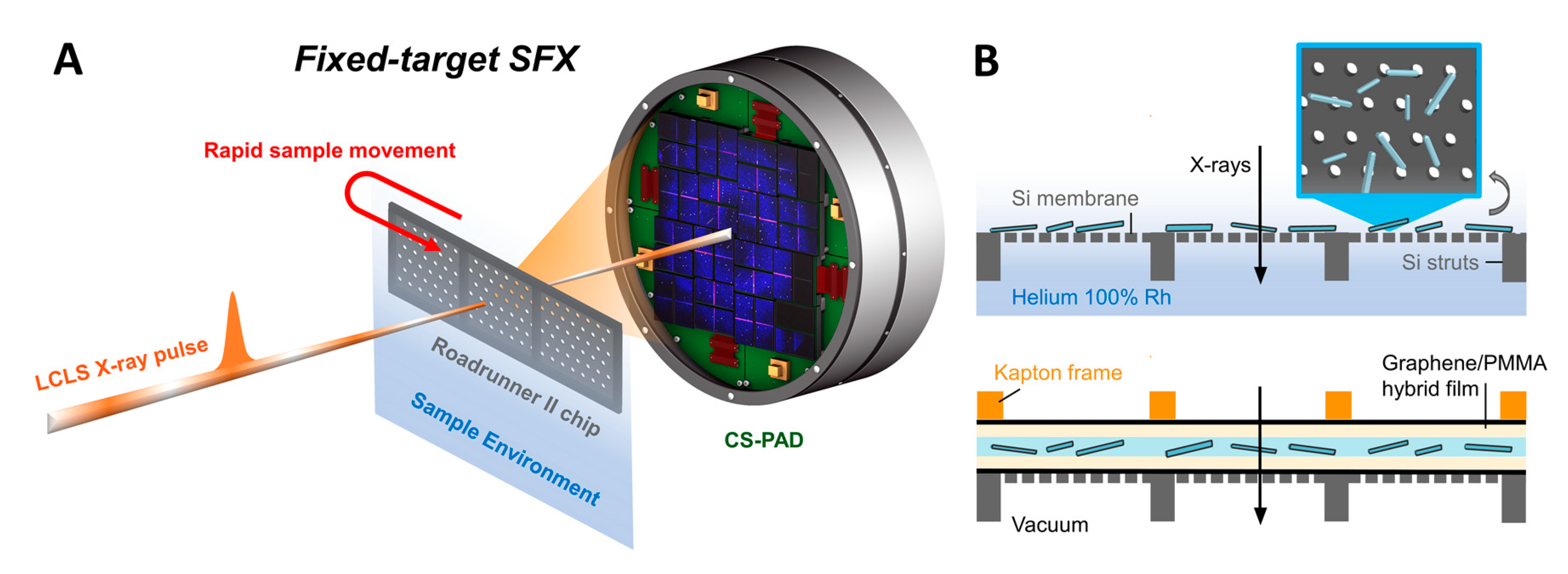
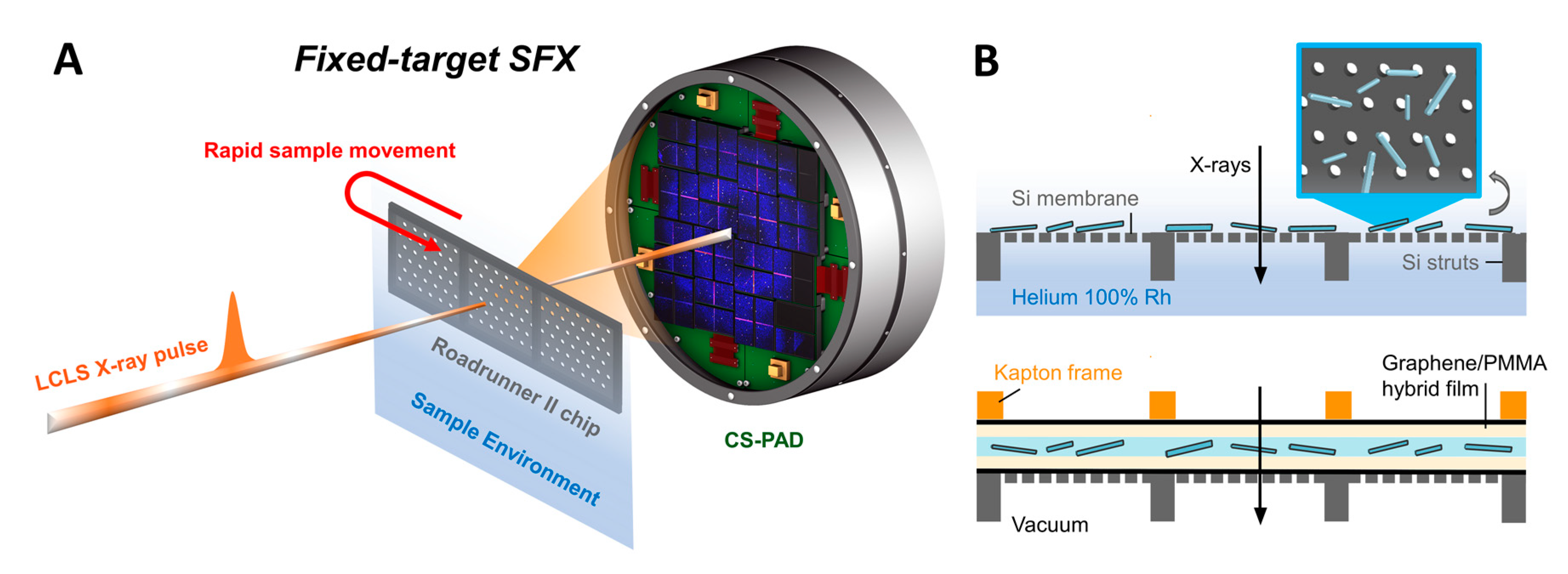
2.4. Fixed Target Preparation and Sample Deposition
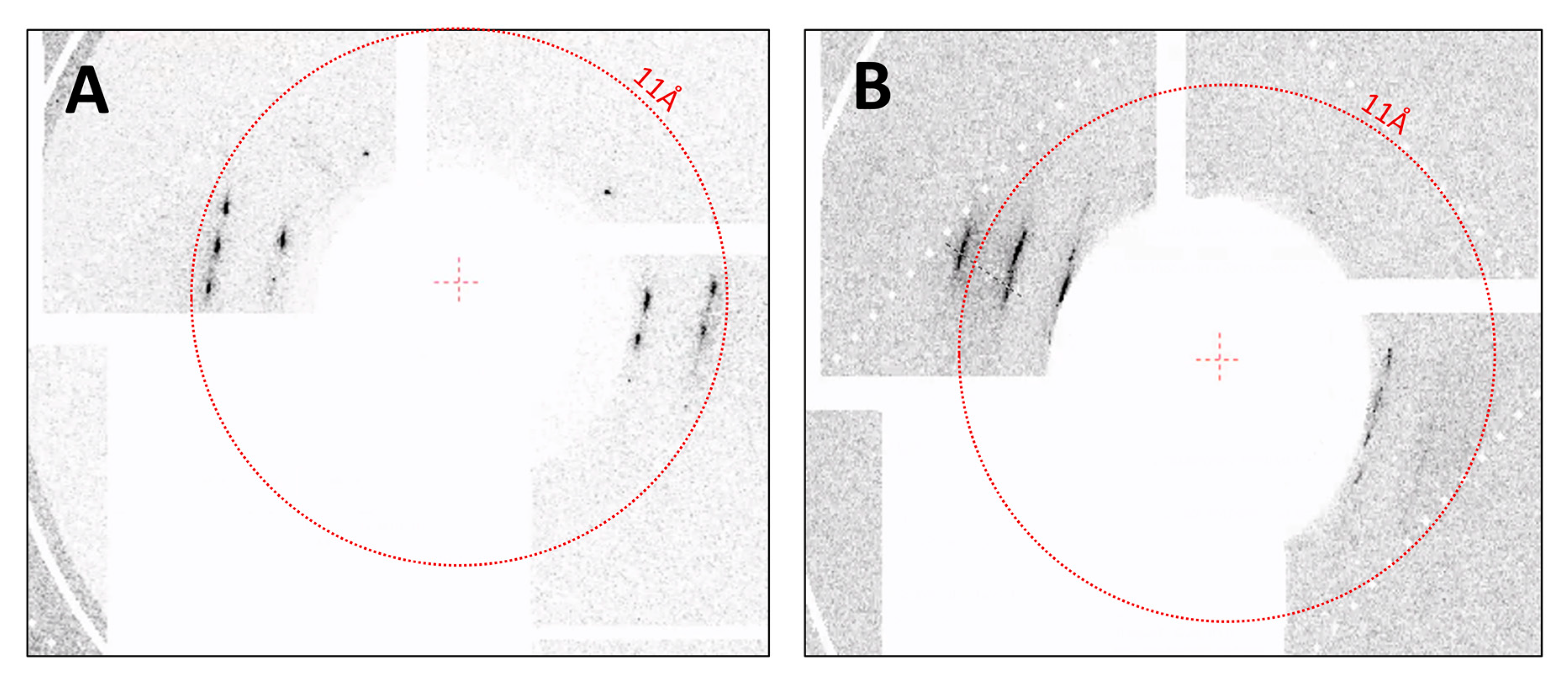
2.5. Sfx Experiments
3. Results and Discussion
3.1. Cell-Free Expression
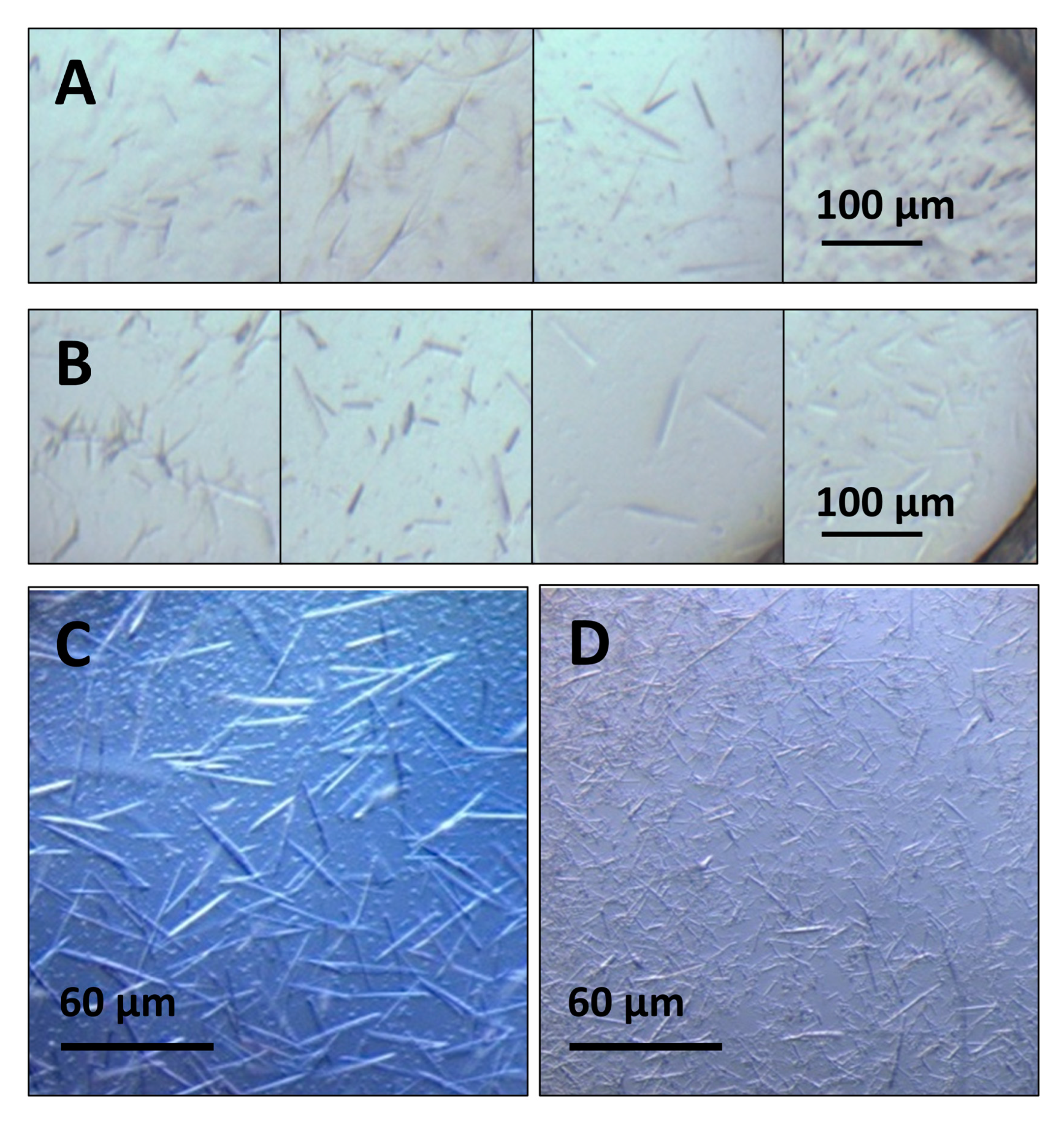
3.2. Crystallization Optimization
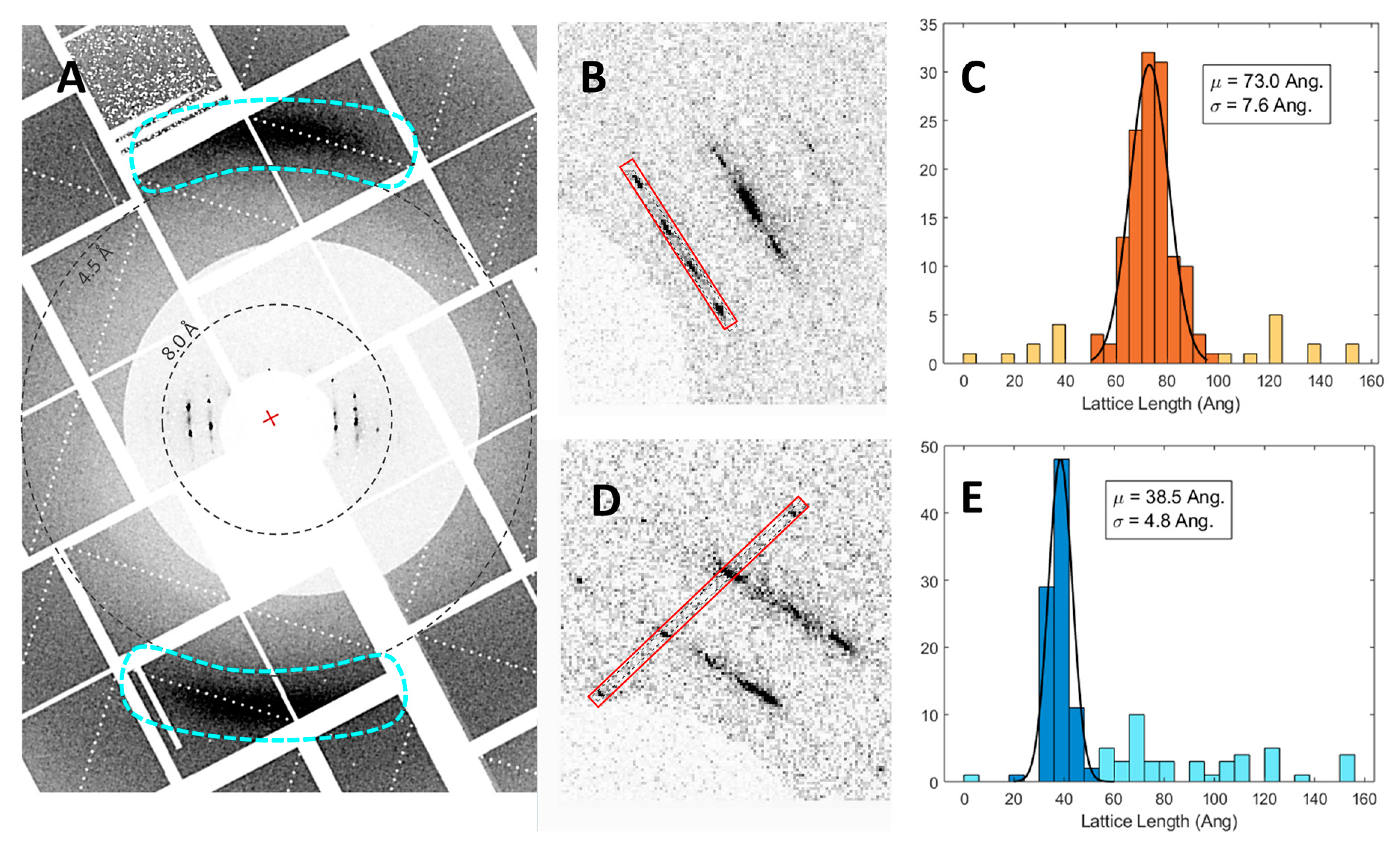
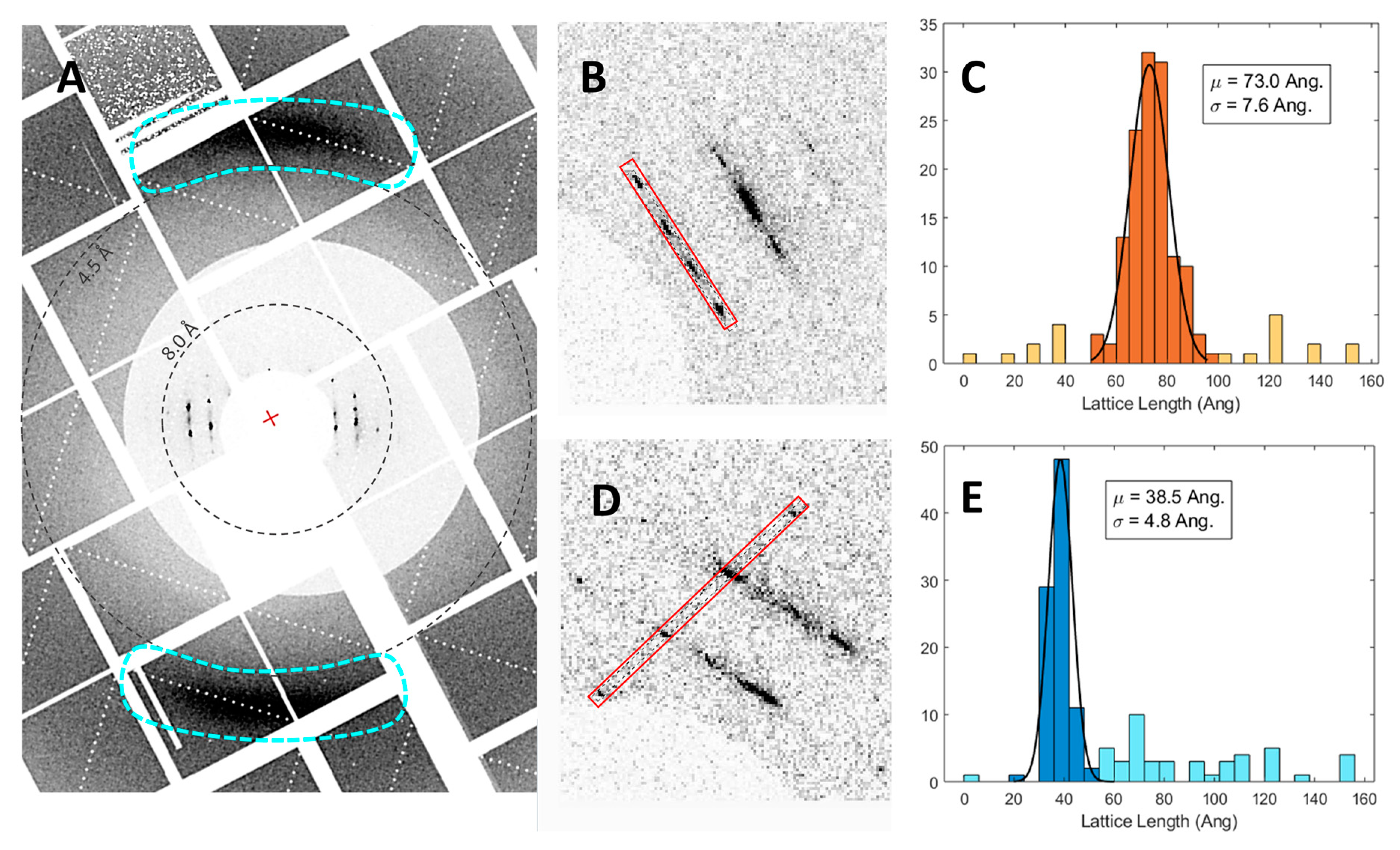
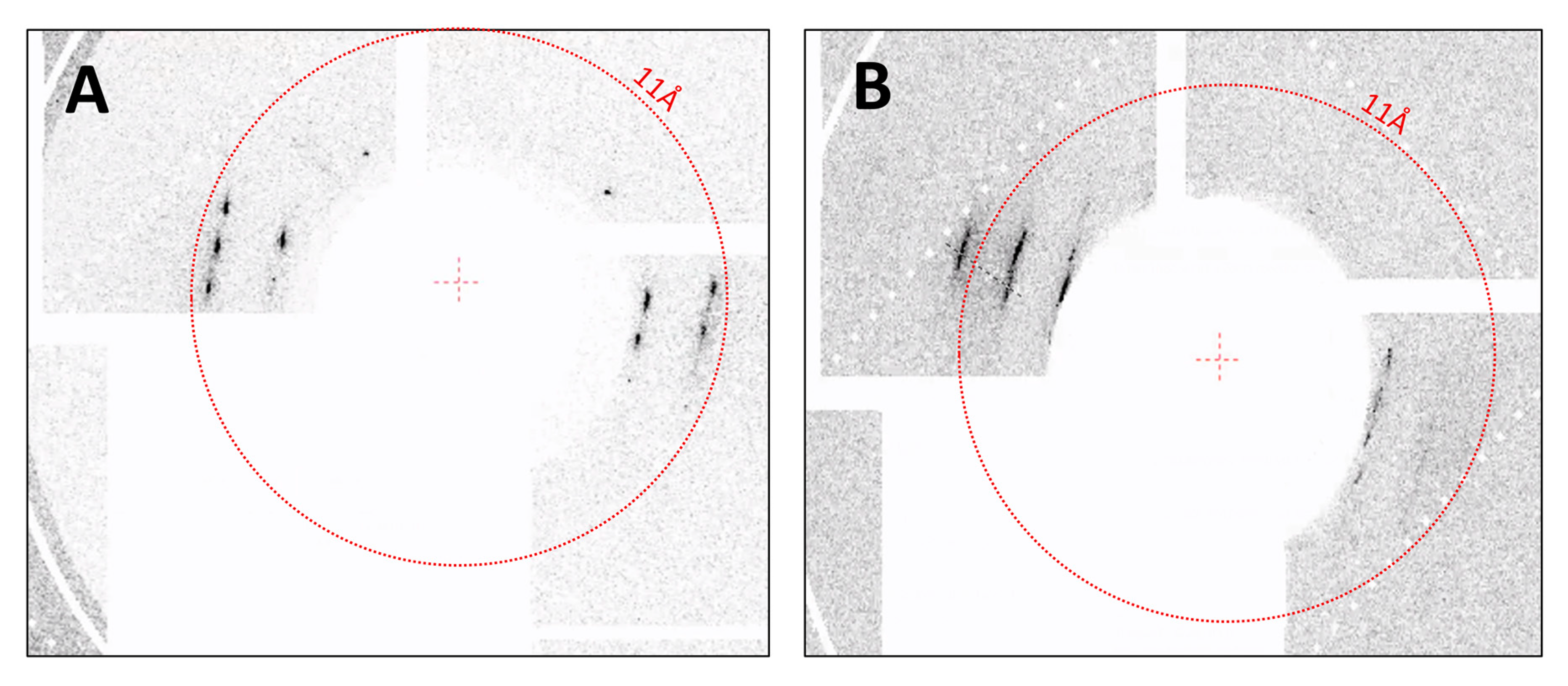
3.3. SFX Experiments
4. Summary
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Feher, V.A.; Randall, A.; Baldi, P.; Bush, R.M.; de la Maza, L.M.; Amaro, R.E. A 3-dimensional trimeric beta-barrel model for Chlamydia MOMP contains conserved and novel elements of Gram-negative bacterial porins. PLoS ONE 2013, 8, e68934. [Google Scholar] [CrossRef]
- Autzen, H.E.; Myasnikov, A.G.; Campbell, M.G.; Asarnow, D.; Julius, D.; Cheng, Y. Structure of the human TRPM4 ion channel in a lipid nanodisc. Science 2018, 359, 228–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roh, S.-H.; Stam, N.J.; Hryc, C.F.; Couoh-Cardel, S.; Pintilie, G.; Chiu, W.; Wilkens, S. The 3.5-Å CryoEM structure of nanodisc-reconstituted yeast vacuolar ATPase Vo proton channel. Mol. Cell 2018, 69, 993–1004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Machen, A.J.; Akkaladevi, N.; Trecazzi, C.; O’Neil, P.T.; Mukherjee, S.; Qi, Y.; Dillard, R.; Im, W.; Gogol, E.P.; White, T.A.; et al. Asymmetric Cryo-EM Structure of anthrax toxin protective antigen pore with lethal factor n-terminal domain. Toxins 2017, 9, 298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Y.; Cao, E.; Julius, D.; Cheng, Y. TRPV1 structures in nanodiscs reveal mechanisms of ligand and lipid action. Nature 2016, 534, 347. [Google Scholar] [CrossRef] [Green Version]
- Kedrov, A.; Sustarsic, M.; de Keyzer, J.; Caumanns, J.J.; Wu, Z.C.; Driessen, A.J.M. Elucidating the native architecture of the YidC: Ribosome complex. J. Mol. Biol. 2013, 425, 4112–4124. [Google Scholar] [CrossRef] [Green Version]
- Akkaladevi, N.; Hinton-Chollet, L.; Katayama, H.; Mitchell, J.; Szerszen, L.; Mukherjee, S.; Gogol, E.P.; Pentelute, B.L.; Collier, R.J.; Fisher, M.T. Assembly of anthrax toxin pore: Lethal-factor complexes into lipid nanodiscs. Protein Sci. Publ. Protein Soc. 2013, 22, 492–501. [Google Scholar] [CrossRef]
- Gogol, E.P.; Akkaladevi, N.; Szerszen, L.; Mukherjee, S.; Chollet-Hinton, L.; Katayama, H.; Pentelute, B.L.; Collier, R.J.; Fisher, M.T. Three dimensional structure of the anthrax toxin translocon-lethal factor complex by cryo-electron microscopy. Protein Sci. Publ. Protein Soc. 2013, 22, 586–594. [Google Scholar] [CrossRef] [Green Version]
- Frauenfeld, J.; Gumbart, J.; Sluis, E.O.; Funes, S.; Gartmann, M.; Beatrix, B.; Mielke, T.; Berninghausen, O.; Becker, T.; Schulten, K.; et al. Cryo-EM structure of the ribosome-SecYE complex in the membrane environment. Nat. Struct. Mol. Biol. 2011, 18, 614–621. [Google Scholar] [CrossRef] [Green Version]
- Midtgaard, S.R.; Pedersen, M.C.; Arleth, L. Small-angle X-ray scattering of the cholesterol incorporation into human ApoA1-POPC discoidal particles. Biophys. J. 2015, 109, 308–318. [Google Scholar] [CrossRef] [Green Version]
- Bayburt, T.H.; Grinkova, Y.V.; Sligar, S.G. Assembly of single bacteriorhodopsin trimers in bilayer nanodiscs. Arch. Biochem. Biophys. 2006, 450, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Raschle, T.; Hiller, S.; Yu, T.-Y.; Rice, A.J.; Walz, T.; Wagner, G. Structural and functional characterization of the integral membrane protein VDAC-1 in lipid bilayer Nanodiscs. J. Am. Chem. Soc. 2009, 131, 17777–17779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kijac, A.Z.; Li, Y.; Sligar, S.G.; Rienstra, C.M. Magic-angle spinning solid-state NMR spectroscopy of nanodisc-embedded human CYP3A4. Biochemistry 2007, 46, 13696–13703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katayama, H.; Wang, J.; Tama, F.; Chollet, L.; Gogol, E.P.; Collier, R.J.; Fisher, M.T. Three-dimensional structure of the anthrax toxin pore inserted into lipid nanodiscs and lipid vesicles. Proc. Natl. Acad. Sci. USA 2010, 107, 3453–3457. [Google Scholar] [CrossRef] [Green Version]
- Bayburt, T.H.; Sligar, S.G. Membrane protein assembly into Nanodiscs. FEBS Lett. 2010, 584, 1721–1727. [Google Scholar] [CrossRef] [Green Version]
- Glomset, J.A. The plasma lecithins:cholesterol acyltransferase reaction. J. Lipid Res. 1968, 9, 155–167. [Google Scholar]
- Glomset, J.A.; Janssen, E.T.; Kennedy, R.; Dobbins, J. Role of plasma lecithin:cholesterol acyltransferase in the metabolism of high density lipoproteins. J. Lipid Res. 1966, 7, 638–648. [Google Scholar]
- Fielding, C.J.; Shore, V.G.; Fielding, P.E. A protein cofactor of lecithin:Cholesterol acyltransferase. Biochem. Biophys. Res. Commun. 1972, 46, 1493–1498. [Google Scholar] [CrossRef]
- Jonas, A.; Sweeny, S.A.; Herbert, P.N. Discoidal complexes of A and C apolipoproteins with lipids and their reactions with lecithin: Cholesterol acyltransferase. J. Biol. Chem. 1984, 259, 6369–6375. [Google Scholar]
- Jonas, A. Lecithin-cholesterol acyltransferase in the metabolism of high-density lipoproteins. Biochim. Biophys. Acta (BBA)-Lipids Lipid Metab. 1991, 1084, 205–220. [Google Scholar] [CrossRef]
- Rajaram, O.V.; Barter, P.J. Reactivity of human lipoproteins with purified lecithin: Cholesterol acyltransferase during incubations in vitro. Biochim. Biophys. Acta 1985, 835, 41–49. [Google Scholar] [CrossRef]
- Remaley, A.T.; Amar, M.; Sviridov, D. HDL-replacement therapy: Mechanism of action, types of agents and potential clinical indications. Expert Rev. Cardiovasc. Ther. 2008, 6, 1203–1215. [Google Scholar] [CrossRef] [Green Version]
- Sorci-Thomas, M.G.; Bhat, S.; Thomas, M.J. Activation of lecithin: Cholesterol acyltransferase by HDL ApoA-I central helices. Clin. Lipidol. 2009, 4, 113–124. [Google Scholar] [CrossRef] [Green Version]
- Chromy, B.A.; Arroyo, E.; Blanchette, C.D.; Bench, G.; Benner, H.; Cappuccio, J.A.; Coleman, M.A.; Henderson, P.T.; Hinz, A.K.; Kuhn, E.A.; et al. Different apolipoproteins impact nanolipoprotein particle formation. J. Am. Chem. Soc. 2007, 129, 14348–14354. [Google Scholar] [CrossRef] [PubMed]
- Marcink, T.C.; Simoncic, J.A.; An, B.; Knapinska, A.M.; Fulcher, Y.G.; Akkaladevi, N.; Fields, G.B.; Van Doren, S.R. MT1-MMP binds membranes by opposite tips of its β propeller to position it for pericellular proteolysis. Structure 2019, 27, 281–292. [Google Scholar] [CrossRef] [Green Version]
- Bibow, S.; Polyhach, Y.; Eichmann, C.; Chi, C.N.; Kowal, J.; Albiez, S.; McLeod, R.A.; Stahlberg, H.; Jeschke, G.; Güntert, P.; et al. Solution structure of discoidal high-density lipoprotein particles with a shortened apolipoprotein A-I. Nat. Struct. Mol. Biol. 2017, 24, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Mackey, R.H.; Greenland, P.; Goff, D.C.; Lloyd-Jones, D.; Sibley, C.T.; Mora, S. High-density lipoprotein cholesterol and particle concentrations, carotid atherosclerosis, and coronary events: MESA (Multi-ethnic study of atherosclerosis). J. Am. Coll. Cardiol. 2012, 60, 508–516. [Google Scholar] [CrossRef] [Green Version]
- Davidson, W.S. HDL-C vs HDL-P: How changing one letter could make a difference in understanding the role of high-density lipoprotein in disease. Clin. Chem. 2014, 60, e1–e3. [Google Scholar] [CrossRef] [Green Version]
- He, W.; Felderman, M.; Evans, A.C.; Geng, J.; Homan, D.; Bourguet, F.; Fischer, N.O.; Li, Y.; Lam, K.S.; Noy, A.; et al. Cell-free production of a functional oligomeric form of a Chlamydia major outer-membrane protein (MOMP) for vaccine development. J. Biol. Chem. 2017, 292, 15121–15132. [Google Scholar] [CrossRef] [Green Version]
- Pourmousa, M.; Song, H.D.; He, Y.; Heinecke, J.W.; Segrest, J.P.; Pastor, R.W. Tertiary structure of apolipoprotein A-I in nascent high-density lipoproteins. Proc. Natl. Acad. Sci. USA 2018, 115, 5163–5168. [Google Scholar] [CrossRef] [Green Version]
- Cleveland, T.E.; He, W.; Evans, A.C.; Fischer, N.O.; Lau, E.Y.; Coleman, M.A.; Butler, P. Small-angle X-ray and neutron scattering demonstrates that cell-free expression produces properly formed disc-shaped nanolipoprotein particles. Protein Sci. 2018, 27, 780–789. [Google Scholar]
- Peters-Libeu, C.A.; Newhouse, Y.; Hatters, D.M.; Weisgraber, K.H. model of biologically active apolipoprotein e bound to dipalmitoylphosphatidylcholine. J. Biol. Chem. 2006, 281, 1073–1079. [Google Scholar]
- Newhouse, Y.; Peters-Libeu, C.; Weisgraber, K.H. Crystallization and preliminary X-ray diffraction analysis of apolipoprotein E-containing lipoprotein particles. Acta Cryst. Sect. F Struct. Biol. Cryst. Commun. 2005, 61, 981–984. [Google Scholar]
- Neutze, R.; Wouts, R.; Spoel, D.V.D.; Weckert, E.; Hajdu, J. Potential for biomolecular imaging with femtosecond X-ray pulses. Nature 2000, 406, 752. [Google Scholar] [PubMed]
- Chapman, H.N.; Fromme, P.; Barty, A.; White, T.A.; Kirian, R.A.; Aquila, A.; Hunter, M.S.; Schulz, J.; DePonte, D.P.; Weierstall, U.; et al. Femtosecond X-ray protein nanocrystallography. Nature 2011, 470, 73–77. [Google Scholar] [PubMed]
- Boutet, S.; Lomb, L.; Williams, G.J.; Barends, T.R.M.; Aquila, A.; Doak, R.B.; Weierstall, U.; DePonte, D.P.; Steinbrener, J.; Shoeman, R.L.; et al. High-resolution protein structure determination by serial femtosecond crystallography. Science 2012, 337, 362–364. [Google Scholar]
- Seibert, M.M.; Ekeberg, T.; Maia, F.R.N.C.; Svenda, M.; Andreasson, J.; Jönsson, O.; Odić, D.; Iwan, B.; Rocker, A.; Westphal, D.; et al. Single mimivirus particles intercepted and imaged with an X-ray laser. Nature 2011, 470, 78–81. [Google Scholar]
- Sun, Z.; Fan, J.; Li, H.; Jiang, H. Current status of single particle imaging with X-ray lasers. Appl. Sci. 2018, 8, 132. [Google Scholar]
- Pedrini, B.; Tsai, C.-J.; Capitani, G.; Padeste, C.; Hunter, M.S.; Zatsepin, N.A.; Barty, A.; Benner, W.H.; Boutet, S.; Feld, G.K.; et al. 7 Å resolution in protein two-dimensional-crystal X-ray diffraction at Linac Coherent Light Source. Phil. Trans. R. Soc. B 2014, 369, 20130500. [Google Scholar]
- Frank, M.; Carlson, D.B.; Hunter, M.S.; Williams, G.J.; Messerschmidt, M.; Zatsepin, N.A.; Barty, A.; Benner, W.H.; Chu, K.; Graf, A.T.; et al. Femtosecond X-ray diffraction from two-dimensional protein crystals. IUCrJ. 2014, 1, 95–100. [Google Scholar]
- Casadei, C.M.; Tsai, C.J.; Barty, A.; Hunter, M.S.; Zatsepin, N.A.; Padeste, C.; Capitani, G.; Benner, W.H.; Boutet, S.; Hau-Riege, S.P.; et al. Resolution extension by image summing in serial femtosecond crystallography of two-dimensional membrane-protein crystals. Int. Union Crystalogr. J. 2018, 5, 103–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seuring, C.; Ayyer, K.; Filippaki, E.; Barthelmess, M.; Longchamp, J.-N.; Ringler, P.; Pardini, T.; Wojtas, D.H.; Coleman, M.A.; Dörner, K. Femtosecond X-ray coherent diffraction of aligned amyloid fibrils on low background graphene. Nat. Commun. 2018, 9, 1836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirian, R.A.; Wang, X.; Weierstall, U.; Schmidt, K.E.; Spence, J.C.H.; Hunter, M.; Fromme, P.; White, T.; Chapman, H.N.; Holton, J. Femtosecond protein nanocrystallography—data analysis methods. Opt. Express 2010, 18, 5713–5723. [Google Scholar] [CrossRef] [PubMed]
- Chavas, L.M.G.; Gumprecht, L.; Chapman, H.N. Possibilities for serial femtosecond crystallography sample delivery at future light sources. Struct. Dynam. 2015, 2, 041709. [Google Scholar]
- White, T.A.; Mariani, V.; Brehm, W.; Yefanov, O.; Barty, A.; Beyerlein, K.R.; Chervinskii, F.; Galli, L.; Gati, C.; Nakane, T.; et al. Recent developments in CrystFEL. J. Appl. Crystallogr. 2016, 49, 680–689. [Google Scholar] [CrossRef] [Green Version]
- Roedig, P.; Ginn, H.M.; Pakendorf, T.; Sutton, G.; Harlos, K.; Walter, T.S.; Meyer, J.; Fischer, P.; Duman, R.; Vartiainen, I.; et al. High-speed fixed-target serial virus crystallography. Nat. Methods, 2017; submitted for publication. [Google Scholar] [CrossRef]
- Roedig, P.; Vartiainen, I.; Duman, R.; Panneerselvam, S.; Stübe, N.; Lorbeer, O.; Warmer, M.; Sutton, G.; Stuart, D.I.; Weckert, E.; et al. A micro-patterned silicon chip as sample holder for macromolecular crystallography experiments with minimal background scattering. Sci. Rep. 2015, 5, 10451. [Google Scholar] [CrossRef]
- Shelby, M.L.; Gilbile, D.; Grant, T.D.; Seuring, C.; Segelke, B.W.; He, W.; Evans, A.C.; Pakendorf, T.; Fischer, P.; Hunter, M.S.; et al. A fixed-target platform for serial femtosecond crystallography in a hydrated environment. Int Union Crystalogr. J. 2020, 7, 30–41. [Google Scholar] [CrossRef] [Green Version]
- Cappuccio, J.A.; Blanchette, C.D.; Sulchek, T.A.; Arroyo, E.S.; Kralj, J.M.; Hinz, A.K.; Kuhn, E.A.; Chromy, B.A.; Segelke, B.W.; Rothschild, K.J.; et al. Cell-free Co-expression of functional membrane proteins and apolipoprotein, forming soluble nanolipoprotein particles. Mol. Cell. Proteom. 2008, 7, 2246–2253. [Google Scholar] [CrossRef] [Green Version]
- Klammt, C.; Schwarz, D.; Lohr, F.; Schneider, B.; Dotsch, V.; Bernhard, F. Cell-free expression as an emerging technique for the large scale production of integral membrane protein. FEBS J. 2006, 273, 4141–4153. [Google Scholar] [CrossRef]
- Koglin, A.; Klammt, C.; Trbovic, N.; Schwarz, D.; Schneider, B.; Schafer, B.; Lohr, F.; Bernhard, F.; Dotsch, V. Combination of cell-free expression and NMR spectroscopy as a new approach for structural investigation of membrane proteins. Magn. Reason. Chem. 2006, 44, S17–S23. [Google Scholar] [CrossRef]
- Lieske, J.; Cerv, M.; Kreida, S.; Komadina, D.; Fischer, J.; Barthelmess, M.; Fischer, P.; Pakendorf, T.; Yefanov, O.; Mariani, V.; et al. On-chip crystallization for serial crystallography experiments and on-chip ligand-binding studies. IUCrJ 2019, 6, 714–728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schropp, A.; Hoppe, R.; Meier, V.; Patommel, J.; Seiboth, F.; Lee, H.J.; Nagler, B.; Galtier, E.C.; Arnold, B.; Zastrau, U.; et al. Full spatial characterization of a nanofocused X-ray free-electron laser beam by ptychographic imaging. Sci. Rep. 2013, 3, 1633. [Google Scholar] [CrossRef] [Green Version]
- Blaj, G.; Caragiulo, P.; Carini, G.; Carron, S.; Dragone, A.; Freytag, D.; Haller, G.; Hart, P.; Hasi, J.; Herbst, R.; et al. X-ray detectors at the Linac Coherent Light Source. J. Synchrotron Radiat. 2015, 22, 577–583. [Google Scholar] [CrossRef] [PubMed]
- Roedig, P.; Duman, R.; Sanchez-Weatherby, J.; Vartiainen, I.; Burkhardt, A.; Warmer, M.; David, C.; Wagner, A.; Meents, A. Room-temperature macromolecular crystallography using a micro-patterned silicon chip with minimal background scattering. J. Appl. Crystallogr. 2016, 49, 968–975. [Google Scholar] [CrossRef] [Green Version]
- Tolstikova, A.; Levantino, M.; Yefanov, O.; Hennicke, V.; Fischer, P.; Meyer, J.; Mozzanica, A.; Redford, S.; Crosas, E.; Opara, N.L.; et al. 1 kHz fixed-target serial crystallography using a multilayer monochromator and an integrating pixel detector. IUCrJ 2019, 6, 927–937. [Google Scholar] [CrossRef] [PubMed]
- Mariani, V.; Morgan, A.; Yoon, C.H.; Lane, T.J.; White, T.A.; O’Grady, C.; Kuhn, M.; Aplin, S.; Koglin, J.; Barty, A.; et al. OnDA: Online data analysis and feedback for serial X-ray imaging. J. Appl. Crystallogr. 2016, 49, 1073–1080. [Google Scholar] [CrossRef] [Green Version]
- Barty, A.; Kirian, R.A.; Maia, F.R.N.C.; Hantke, M.; Yoon, C.H.; White, T.A.; Chapman, H. Cheetah: Software for high-throughput reduction and analysis of serial femtosecond X-ray diffraction data. J. Appl. Crystallogr. 2014, 47, 1118–1131. [Google Scholar] [CrossRef] [Green Version]
- Arvai, A. ADXV-A Program to Display X-ray Diffraction Images; Scripps Research Institute: La Jolla, CA, USA, 2012. [Google Scholar]
- Cappuccio, J.; Hinz, A.; Kuhn, E.; Fletcher, J.; Arroyo, E.; Henderson, P.; Blanchette, C.; Walsworth, V.; Corzett, M.; Law, R.; et al. Cell-free expression for nanolipoprotein particles: Building a high-throughput membrane protein solubility platform. In High Throughput Protein Expression and Purification; Doyle, S., Ed.; Humana Press: Totowa, NJ, USA, 2009; pp. 273–295. [Google Scholar]
- He, W.; Luo, J.; Bourguet, F.; Xing, L.; Yi, S.K.; Gao, T.; Blanchette, C.; Henderson, P.T.; Kuhn, E.; Malfatti, M.; et al. Controlling the diameter, monodispersity, and solubility of ApoA1 nanolipoprotein particles using telodendrimer chemistry. Protein Sci. 2013, 22, 1078–1086. [Google Scholar] [CrossRef] [Green Version]
- Blanchette, C.D.; Cappuccio, J.A.; Kuhn, E.A.; Segelke, B.W.; Benner, W.H.; Chromy, B.A.; Coleman, M.A.; Bench, G.; Hoeprich, P.D.; Sulchek, T.A. Atomic force microscopy differentiates discrete size distributions between membrane protein containing and empty nanolipoprotein particles. Biochim. Biophys. Acta (BBA) Biomembr. 2009, 1788, 724–731. [Google Scholar] [CrossRef] [Green Version]
- Blanchette, C.; Segelke, B.; Fischer, N.; Corzett, M.; Kuhn, E.; Cappuccio, J.; Benner, W.H.; Coleman, M.; Chromy, B.; Bench, G.; et al. Characterization and purification of polydisperse reconstituted lipoproteins and nanolipoprotein particles. Int. J. Mol. Sci. 2009, 10, 2958–2971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blanchette, C.D.; Law, R.; Benner, W.H.; Pesavento, J.B.; Cappuccio, J.A.; Walsworth, V.; Kuhn, E.A.; Corzett, M.; Chromy, B.A.; Segelke, B.W.; et al. Quantifying size distributions of nanolipoprotein particles with single-particle analysis and molecular dynamic simulations. J. Lipid Res. 2008, 49, 1420–1430. [Google Scholar] [CrossRef] [Green Version]
- Luft, J.R.; Collins, R.J.; Fehrman, N.A.; Lauricella, A.M.; Veatch, C.K.; DeTitta, G.T. A deliberate approach to screening for initial crystallization conditions of biological macromolecules. J. Struct. Biol. 2003, 142, 170–179. [Google Scholar] [CrossRef]
- Koszelak-Rosenblum, M.; Krol, A.; Mozumdar, N.; Wunsch, K.; Ferin, A.; Cook, E.; Veatch, C.K.; Nagel, R.; Luft, J.R.; DeTitta, G.T.; et al. Determination and application of empirically derived detergent phase boundaries to effectively crystallize membrane proteins. Protein Sci. 2009, 18, 1828–1839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alzate, O.; Osorio, C.; DeKroon, R.M.; Corcimaru, A.; Gunawardena, H.P. Differentially charged isoforms of apolipoprotein E from human blood are potential biomarkers of Alzheimer’s disease. Alzheimers Res. Ther. 2014, 6, 43. [Google Scholar] [CrossRef] [Green Version]
- McIntosh, T.J.; Simon, S.A. Area per molecule and distribution of water in fully hydrated dilauroylphosphatidylethanolamine bilayers. Biochemistry 1986, 25, 4948–4952. [Google Scholar] [CrossRef]
- McIntosh, T.J.; Simon, S.A. Hydration force and bilayer deformation: A reevaluation. Biochemistry 1986, 25, 4058–4066. [Google Scholar] [CrossRef]
- Zhang, L.; Song, J.; Cavigiolio, G.; Ishida, B.Y.; Zhang, S.; Kane, J.P.; Weisgraber, K.H.; Oda, M.N.; Rye, K.-A.; Pownall, H.J.; et al. Morphology and structure of lipoproteins revealed by an optimized negative-staining protocol of electron microscopy. J. Lipid Res. 2011, 52, 175–184. [Google Scholar] [CrossRef] [Green Version]
- Spence, J.C.H. XFELs for structure and dynamics in biology. IUCrJ 2017, 4, 322–339. [Google Scholar] [CrossRef]
- Dandey, V.P.; Budell, W.C.; Wei, H.; Bobe, D.; Maruthi, K.; Kopylov, M.; Eng, E.T.; Kahn, P.A.; Hinshaw, J.E.; Kundu, N.; et al. Time-resolved cryo-EM using Spotiton. Nat. Methods 2020, 17, 897–900. [Google Scholar] [CrossRef]
- Mäeots, M.-E.; Lee, B.; Nans, A.; Jeong, S.-G.; Esfahani, M.M.N.; Ding, S.; Smith, D.J.; Lee, C.-S.; Lee, S.S.; Peter, M.; et al. Modular microfluidics enables kinetic insight from time-resolved cryo-EM. Nat. Commun. 2020, 11, 3465. [Google Scholar] [CrossRef] [PubMed]
- Kaledhonkar, S.; Fu, Z.; Caban, K.; Li, W.; Chen, B.; Sun, M.; Gonzalez, R.L.; Frank, J. Late steps in bacterial translation initiation visualized using time-resolved cryo-EM. Nature 2019, 570, 400–404. [Google Scholar] [CrossRef] [PubMed]
- Frank, J. Time-resolved cryo-electron microscopy: Recent progress. J. Struct. Biol. 2017, 200, 303–306. [Google Scholar] [CrossRef] [PubMed]
- Fischer, N.; Konevega, A.L.; Wintermeyer, W.; Rodnina, M.V.; Stark, H. Ribosome dynamics and tRNA movement by time-resolved electron cryomicroscopy. Nature 2010, 466, 329–333. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cocktail Number | Buffer Concentration and pH | PEG Precipitant Concentration | Salt additive Concentration | Length Range, µm (Average) | Estimated Density, #crystals/µL | |
|---|---|---|---|---|---|---|
| ApoE4 NLPs | M1328 | 0.1M Tris pH 8.5 | 22.6% w/v PEG 4000 | 0.2M Lithium Sulfate monohydrate | 50–140 (85) | 73 |
| M318 | 0.1M Sodium Citrate pH 5.6 | 16% w/v PEG 4000 | 0.1M Ammonium Phosphate-dibasic | 50–150 (70) | 17.5 | |
| M548 | 0.2M Tris pH 8.5 | 35% w/v PEG 4000 | 0.2M Lithium Sulfate monohydrate | 25–50 (42) | 8 | |
| M750 | 0.1M Tris pH 8.5 | 40% w/v PEG 1000 | 0.2M Lithium Sulfate monohydrate | |||
| M752 | 0.1M Tris pH 8.5 | 37.5% w/v PEG 1000 | 0.3M Lithium Sulfate monohydrate | |||
| S1022 | 0.1M Sodium citrate pH 5.5 | 20% w/v PEG 1000 | 0.1M Lithium sulfate monohydrate | 40–130 (52) | 74 | |
| S1438 | --- | 22.6% w/v PEG 4000 | 0.2M Sodium Citrate tribasic dihydrate | |||
| ApoA1 NLPs | S714 | 0.1M Tris pH 8.0 | 20% w/v PEG 1000 | 0.1M Magnesium chloride-hexahydrate | ||
| S789 | 0.1M CAPS, pH 10 | 20% w/v PEG 1000 | 0.1M Lithium chloride | |||
| S816 | 0.1M CAPS pH 10 | 20% w/v PEG 1000 | 0.1M Sodium bromide | 80–200 (120) | 84 | |
| S816-2 | 0.1M CAPS pH 10 | 25% w/v PEG 1000 | 0.1M Sodium bromide | 20–140 (58) | 229 | |
| S816-3 | 0.1M CAPS pH 10 | 30% w/v PEG 1000 | 0.1M Sodium bromide | 30–100 (62) | 198 | |
| S816-4 | 0.1M CAPS pH 10 | 35% w/v PEG 1000 | 0.1M Sodium bromide | 20–160 (75) | 92 | |
| S831 | 0.1M CAPS pH 10 | 20% w/v PEG 1000 | 0.1M Lithium sulfate-monohydrate |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shelby, M.L.; Gilbile, D.; Grant, T.D.; Bauer, W.J.; Segelke, B.; He, W.; Evans, A.C.; Crespo, N.; Fischer, P.; Pakendorf, T.; et al. Crystallization of ApoA1 and ApoE4 Nanolipoprotein Particles and Initial XFEL-Based Structural Studies. Crystals 2020, 10, 886. https://doi.org/10.3390/cryst10100886
Shelby ML, Gilbile D, Grant TD, Bauer WJ, Segelke B, He W, Evans AC, Crespo N, Fischer P, Pakendorf T, et al. Crystallization of ApoA1 and ApoE4 Nanolipoprotein Particles and Initial XFEL-Based Structural Studies. Crystals. 2020; 10(10):886. https://doi.org/10.3390/cryst10100886
Chicago/Turabian StyleShelby, Megan L., Deepshika Gilbile, Thomas D. Grant, William J. Bauer, Brent Segelke, Wei He, Angela C. Evans, Natalia Crespo, Pontus Fischer, Tim Pakendorf, and et al. 2020. "Crystallization of ApoA1 and ApoE4 Nanolipoprotein Particles and Initial XFEL-Based Structural Studies" Crystals 10, no. 10: 886. https://doi.org/10.3390/cryst10100886
APA StyleShelby, M. L., Gilbile, D., Grant, T. D., Bauer, W. J., Segelke, B., He, W., Evans, A. C., Crespo, N., Fischer, P., Pakendorf, T., Hennicke, V., Hunter, M. S., Batyuk, A., Barthelmess, M., Meents, A., Kuhl, T. L., Frank, M., & Coleman, M. A. (2020). Crystallization of ApoA1 and ApoE4 Nanolipoprotein Particles and Initial XFEL-Based Structural Studies. Crystals, 10(10), 886. https://doi.org/10.3390/cryst10100886