Use of Lactobacillus rhamnosus (ATCC 53103) as Whole-Cell Biocatalyst for the Regio- and Stereoselective Hydration of Oleic, Linoleic, and Linolenic Acid



Abstract
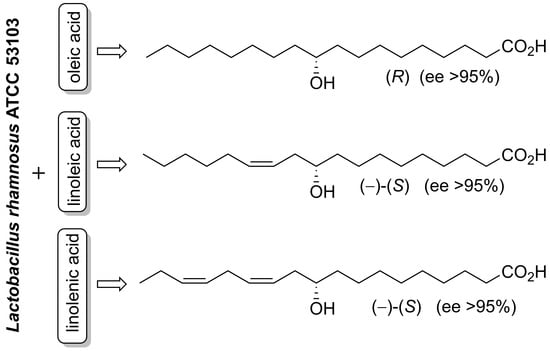
{kind=link}
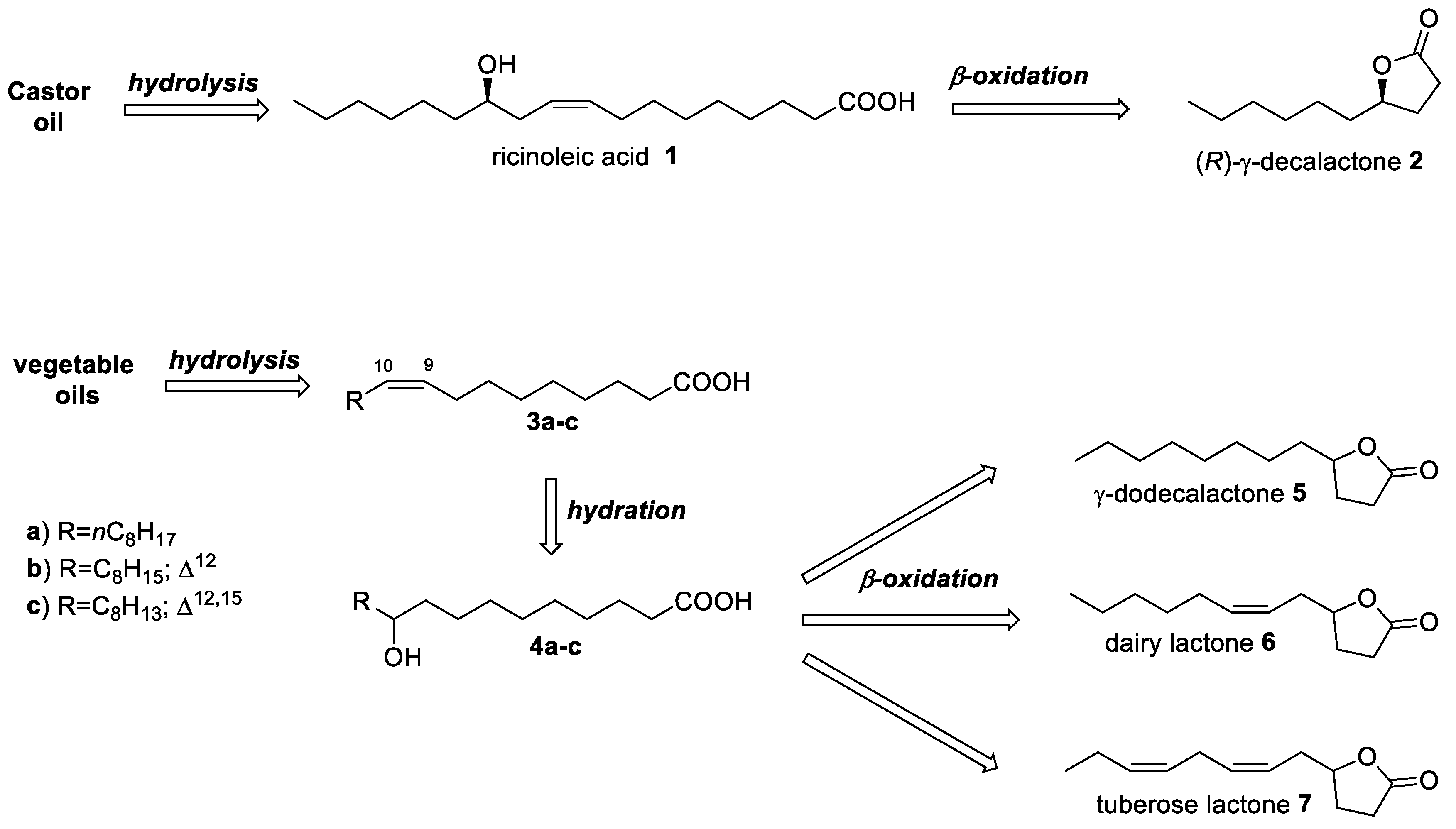{kind=link}
{kind=link}
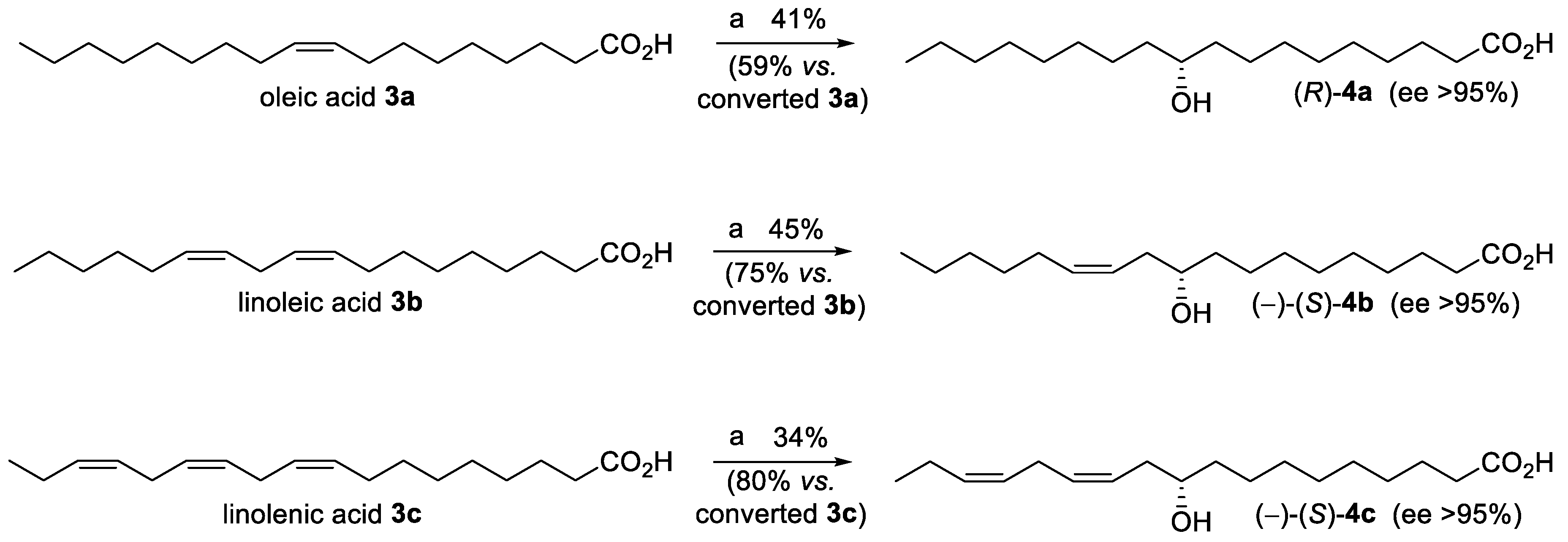{kind=link}
{kind=link}
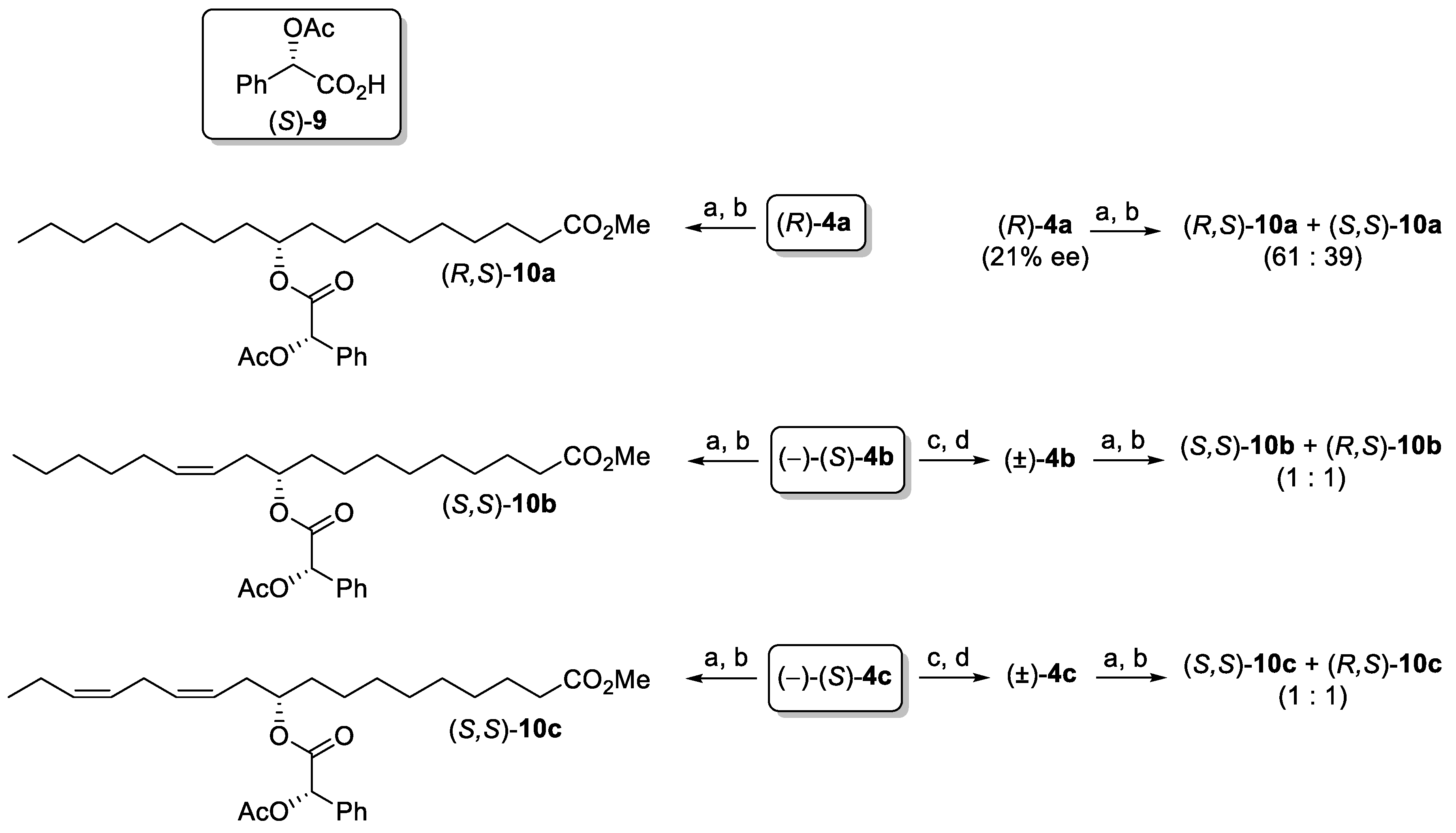{kind=link}
{kind=link}
1. Introduction
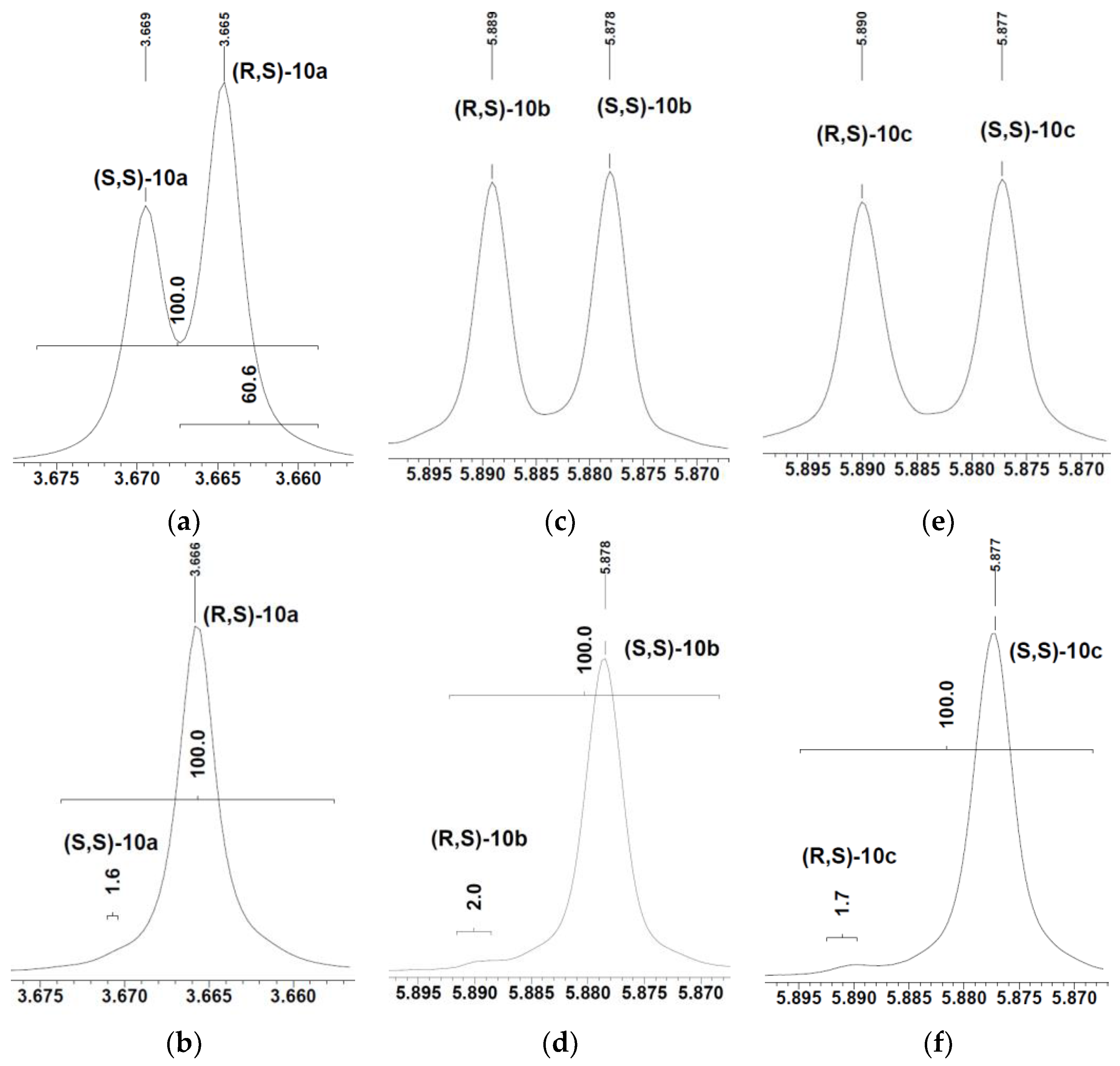
2. Results and Discussion
3. Materials and Methods
3.1. Materials and General Methods
3.2. Analytical Methods and Characterization of the Products Deriving from the Biotransformation Experiments
3.3. Biotransformation Experiments
3.3.1. Representative Procedure for Flask-Scale Biotransformations
3.3.2. Representative Procedure for Preparative Biotransformations
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Kim, K.-R.; Oh, D.-K. Production of hydroxy fatty acids by microbial fatty acid-hydroxylation enzymes. Biotechnol. Adv. 2013, 31, 1473–1485. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Zhang, X. Production of long-chain hydroxy fatty acids by microbial conversion. Appl. Microbiol. Biotechnol. 2013, 97, 3323–3331. [Google Scholar] [CrossRef] [PubMed]
- Lu, W.; Ness, J.E.; Xie, W.; Zhang, X.; Minshull, J.; Gross, R.A. Biosynthesis of monomers for plastics from renewable oils. J. Am. Chem. Soc. 2010, 132, 15451–15455. [Google Scholar] [CrossRef] [PubMed]
- Serra, S.; Fuganti, C.; Brenna, E. Biocatalytic preparation of natural flavours and fragrances. Trends Biotechnol. 2005, 23, 193–198. [Google Scholar] [CrossRef] [PubMed]
- Braga, A.; Belo, I. Biotechnological production of γ-decalactone, a peach like aroma, by Yarrowia lipolytica. World J. Microbiol. Biotechnol. 2016, 32, 169. [Google Scholar] [CrossRef] [PubMed]
- Waché, Y.; Aguedo, M.; Nicaud, J.-M.; Belin, J.-M. Catabolism of hydroxyacids and biotechnological production of lactones by Yarrowia lipolytica. Appl. Microbiol. Biotechnol. 2003, 61, 393–404. [Google Scholar] [CrossRef] [PubMed]
- Kim, A.Y. Application of biotechnology to the production of natural flavor and fragrance chemicals. In Natural Flavors and Fragrances; American Chemical Society: Washington, DC, USA, 2005; Volume 908, pp. 60–75. ISBN 0-8412-3904-5. [Google Scholar]
- Maurer, B.; Hauser, A. Identification and synthesis of new γ-lactones from tuberose absolute (Polianthes tuberosa). Helv. Chim. Acta 1982, 65, 462–476. [Google Scholar] [CrossRef]
- Wallen, L.L.; Benedict, R.G.; Jackson, R.W. The microbiological production of 10-hydroxystearic acid from oleic acid. Arch. Biochem. Biophys. 1962, 99, 249–253. [Google Scholar] [CrossRef]
- Davis, E.N.; Wallen, L.L.; Goodwin, J.C.; Rohwedder, W.K.; Rhodes, R.A. Microbial hydration of cis-9-alkenoic acids. Lipids 1969, 4, 356–362. [Google Scholar] [CrossRef] [PubMed]
- Seo, C.W.; Yamada, Y.; Takada, N.; Okada, H. Hydration of squalene and oleic acid by Corynebacterium sp. S-401. Agric. Biol. Chem. 1981, 45, 2025–2030. [Google Scholar] [CrossRef]
- Koritala, S.; Hou, C.T.; Hesseltine, C.W.; Bagby, M.O. Microbial conversion of oleic acid to 10-hydroxystearic acid. Appl. Microbiol. Biotechnol. 1989, 32, 299–304. [Google Scholar] [CrossRef]
- El-Sharkawy, S.H.; Yang, W.; Dostal, L.; Rosazza, J.P. Microbial oxidation of oleic acid. Appl. Environ. Microbiol. 1992, 58, 2116–2122. [Google Scholar] [PubMed]
- Koritala, S.; Bagby, M.O. Microbial conversion of linoleic and linolenic acids to unsaturated hydroxy fatty acids. J. Am. Oil Chem. Soc. 1992, 69, 575–578. [Google Scholar] [CrossRef]
- Kaneshiro, T.; Huang, J.-K.; Weisleder, D.; Bagby, M.O. 10(R)-Hydroxystearic acid production by a novel microbe, NRRL B-14797, isolated from compost. J. Ind. Microbiol. 1994, 13, 351–355. [Google Scholar] [CrossRef]
- Hou, C.T. Conversion of linoleic acid to 10-hydroxy-12(Z)-octadecenoic acid by Flavobacterium sp. (NRRL B-14859). J. Am. Oil Chem. Soc. 1994, 71, 975–978. [Google Scholar] [CrossRef]
- Gocho, S.; Tabogami, N.; Inagaki, M.; Kawabata, C.; Komai, T. Biotransformation of oleic acid to optically active γ-dodecalactone. Biosci. Biotechnol. Biochem. 1995, 59, 1571–1572. [Google Scholar] [CrossRef]
- Hudson, J.A.; MacKenzie, C.A.M.; Joblin, K.N. Conversion of oleic acid to 10-hydroxystearic acid by two species of ruminal bacteria. Appl. Microbiol. Biotechnol. 1995, 44, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Hou, C.T. Is strain DS5 hydratase a C-10 positional specific enzyme? Identification of bioconversion products from α- and γ-linolenic acids by Flavobacterium sp. DS5. J. Ind. Microbiol. 1995, 14, 31–34. [Google Scholar] [CrossRef]
- Morvan, B.; Joblin, K.N. Hydration of oleic acid by Enterococcus gallinarum, Pediococcus acidilactici and Lactobacillus sp. isolated from the rumen. Anaerobe 1999, 5, 605–611. [Google Scholar] [CrossRef]
- Kishimoto, N.; Yamamoto, I.; Toraishi, K.; Yoshioka, S.; Saito, K.; Masuda, H.; Fujita, T. Two distinct pathways for the formation of hydroxy FA from linoleic acid by lactic acid bacteria. Lipids 2003, 38, 1269–1274. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.H.; Park, M.S.; Chung, C.H.; Kim, C.T.; Kim, Y.S.; Kyung, K.H. Conversion of unsaturated food fatty acids into hydroxy fatty acids by lactic acid bacteria. J. Microbiol. Biotechnol. 2003, 13, 360–365. [Google Scholar]
- Kim, B.-N.; Yeom, S.-J.; Oh, D.-K. Conversion of oleic acid to 10-hydroxystearic acid by whole cells of Stenotrophomonas nitritireducens. Biotechnol. Lett. 2011, 33, 993–997. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, M.; Kishino, S.; Tanabe, K.; Hirata, A.; Park, S.-B.; Shimizu, S.; Ogawa, J. Hydroxy fatty acid production by Pediococcus sp. Eur. J. Lipid Sci. Technol. 2013, 115, 386–393. [Google Scholar] [CrossRef]
- Serra, S.; De Simeis, D. New insights on the baker’s yeast-mediated hydration of oleic acid: The bacterial contaminants of yeast are responsible for the stereoselective formation of (R)-10-hydroxystearic acid. J. Appl. Microbiol. 2018, 124, 719–729. [Google Scholar] [CrossRef] [PubMed]
- Bevers, L.E.; Pinkse, M.W.H.; Verhaert, P.D.E.M.; Hagen, W.R. Oleate hydratase catalyzes the hydration of a nonactivated carbon-carbon bond. J. Bacteriol. 2009, 191, 5010–5012. [Google Scholar] [CrossRef] [PubMed]
- Joo, Y.-C.; Jeong, K.-W.; Yeom, S.-J.; Kim, Y.-S.; Kim, Y.; Oh, D.-K. Biochemical characterization and fad-binding analysis of oleate hydratase from Macrococcus caseolyticus. Biochimie 2012, 94, 907–915. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Chen, H.; Song, Y.; Chen, Y.Q.; Zhang, H.; Chen, W. Myosin-cross-reactive antigens from four different lactic acid bacteria are fatty acid hydratases. Biotechnol. Lett. 2013, 35, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Jo, Y.-S.; An, J.-U.; Oh, D.-K. γ-dodecelactone production from safflower oil via 10-hydroxy-12(Z)-octadecenoic acid intermediate by whole cells of Candida boidinii and Stenotrophomonas nitritireducens. J. Agric. Food Chem. 2014, 62, 6736–6745. [Google Scholar] [CrossRef] [PubMed]
- Oh, H.-J.; Kim, S.-U.; Song, J.-W.; Lee, J.-H.; Kang, W.-R.; Jo, Y.-S.; Kim, K.-R.; Bornscheuer, U.T.; Oh, D.-K.; Park, J.-B. Biotransformation of linoleic acid into hydroxy fatty acids and carboxylic acids using a linoleate double bond hydratase as key enzyme. Adv. Synth. Catal. 2015, 357, 408–416. [Google Scholar] [CrossRef]
- Takeuchi, M.; Kishino, S.; Hirata, A.; Park, S.-B.; Kitamura, N.; Ogawa, J. Characterization of the linoleic acid Δ9 hydratase catalyzing the first step of polyunsaturated fatty acid saturation metabolism in Lactobacillus plantarum AKU 1009a. J. Biosci. Bioeng. 2015, 119, 636–641. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.-R.; Oh, H.-J.; Park, C.-S.; Hong, S.-H.; Park, J.-Y.; Oh, D.-K. Unveiling of novel regio-selective fatty acid double bond hydratases from Lactobacillus acidophilus involved in the selective oxyfunctionalization of mono- and di-hydroxy fatty acids. Biotechnol. Bioeng. 2015, 112, 2206–2213. [Google Scholar] [CrossRef] [PubMed]
- Hiseni, A.; Arends, I.W.C.E.; Otten, L.G. New cofactor-independent hydration biocatalysts: Structural, biochemical, and biocatalytic characteristics of carotenoid and oleate hydratases. ChemCatChem 2015, 7, 29–37. [Google Scholar] [CrossRef]
- Chen, Y.Y.; Liang, N.Y.; Curtis, J.M.; Gänzle, M.G. Characterization of linoleate 10-hydratase of Lactobacillus plantarum and novel antifungal metabolites. Front. Microbiol. 2016, 7, 1561. [Google Scholar] [CrossRef] [PubMed]
- Kang, W.-R.; Seo, M.-J.; An, J.-U.; Shin, K.-C.; Oh, D.-K. Production of δ-decalactone from linoleic acid via 13-hydroxy-9(Z)-octadecenoic acid intermediate by one-pot reaction using linoleate 13-hydratase and whole Yarrowia lipolytica cells. Biotechnol. Lett. 2016, 38, 817–823. [Google Scholar] [CrossRef] [PubMed]
- Demming, R.M.; Otte, K.B.; Nestl, B.M.; Hauer, B. Optimized reaction conditions enable the hydration of non-natural substrates by the oleate hydratase from Elizabethkingia meningoseptica. ChemCatChem 2017, 9, 758–766. [Google Scholar] [CrossRef]
- Lorenzen, J.; Driller, R.; Waldow, A.; Qoura, F.; Loll, B.; Bruck, T. Rhodococcus erythropolis oleate hydratase: A new member in the oleate hydratase family tree biochemical and structural studies. ChemCatchem 2018, 10, 407–414. [Google Scholar] [CrossRef]
- Bourdichon, F.; Casaregola, S.; Farrokh, C.; Frisvad, J.C.; Gerds, M.L.; Hammes, W.P.; Harnett, J.; Huys, G.; Laulund, S.; Ouwehand, A.; et al. Food fermentations: Microorganisms with technological beneficial use. Int. J. Food Microbiol. 2012, 154, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Gorbach, S.H.; Goldin, B.R. Lactobacillus Strains and Methods of Selection. U.S. Patent 4839281, 13 June 1989. [Google Scholar]
- Ku, S.; You, H.J.; Park, M.S.; Ji, G.E. Whole-cell biocatalysis for producing ginsenoside Rd from Rb1 using Lactobacillus rhamnosus GG. J. Microbiol. Biotechnol. 2016, 26, 1206–1215. [Google Scholar] [CrossRef] [PubMed]
- Sturme, M.H.J.; Francke, C.; Siezen, R.J.; de Vos, W.M.; Kleerebezem, M. Making sense of quorum sensing in lactobacilli: A special focus on Lactobacillus plantarum WCSF1. Microbiology 2007, 153, 3939–3947. [Google Scholar] [CrossRef] [PubMed]
- Murata, A.; Kai, K.; Tsutsui, K.; Takeuchi, J.; Todoroki, Y.; Furihata, K.; Yokoyama, M.; Baldermann, S.; Watanabe, N. Enantio-selective reduction of the flowering related compound KODA and its analogues in Pharbitis nil cv. Violet. Tetrahedron 2012, 68, 5583–5589. [Google Scholar] [CrossRef][Green Version]
- Yang, W.; Dostal, L.; Rosazza, J.P.N. Stereospecificity of microbial hydrations of oleic acid to 10-hydroxystearic acid. Appl. Environ. Microbiol. 1993, 59, 281–284. [Google Scholar] [PubMed]
- Mancuso, A.J.; Huang, S.-L.; Swern, D. Oxidation of long-chain and related alcohols to carbonyls by dimethyl sulfoxide “activated” by oxalyl chloride. J. Org. Chem. 1978, 43, 2480–2482. [Google Scholar] [CrossRef]
- Ebbers, E.J.; Ariaans, G.J.A.; Bruggink, A.; Zwanenburg, B. Controlled racemization and asymmetric transformation of α-substituted carboxylic acids in the melt. Tetrahedron Asymmetry 1999, 10, 3701–3718. [Google Scholar] [CrossRef]






© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Serra, S.; De Simeis, D. Use of Lactobacillus rhamnosus (ATCC 53103) as Whole-Cell Biocatalyst for the Regio- and Stereoselective Hydration of Oleic, Linoleic, and Linolenic Acid. Catalysts 2018, 8, 109. https://doi.org/10.3390/catal8030109
Serra S, De Simeis D. Use of Lactobacillus rhamnosus (ATCC 53103) as Whole-Cell Biocatalyst for the Regio- and Stereoselective Hydration of Oleic, Linoleic, and Linolenic Acid. Catalysts. 2018; 8(3):109. https://doi.org/10.3390/catal8030109
Chicago/Turabian StyleSerra, Stefano, and Davide De Simeis. 2018. "Use of Lactobacillus rhamnosus (ATCC 53103) as Whole-Cell Biocatalyst for the Regio- and Stereoselective Hydration of Oleic, Linoleic, and Linolenic Acid" Catalysts 8, no. 3: 109. https://doi.org/10.3390/catal8030109
APA StyleSerra, S., & De Simeis, D. (2018). Use of Lactobacillus rhamnosus (ATCC 53103) as Whole-Cell Biocatalyst for the Regio- and Stereoselective Hydration of Oleic, Linoleic, and Linolenic Acid. Catalysts, 8(3), 109. https://doi.org/10.3390/catal8030109