Construction of a Vitreoscilla Hemoglobin Promoter-Based Tunable Expression System for Corynebacterium glutamicum

 ,
,  ,
,  and
and
Abstract
1. Introduction
2. Results and Discussion
2.1. Construction of Pvgb Promoter-Based Tunable Expression Systems for C. glutamicum and Evaluation of Green Fluorescent Protein in the Recombinant Strains
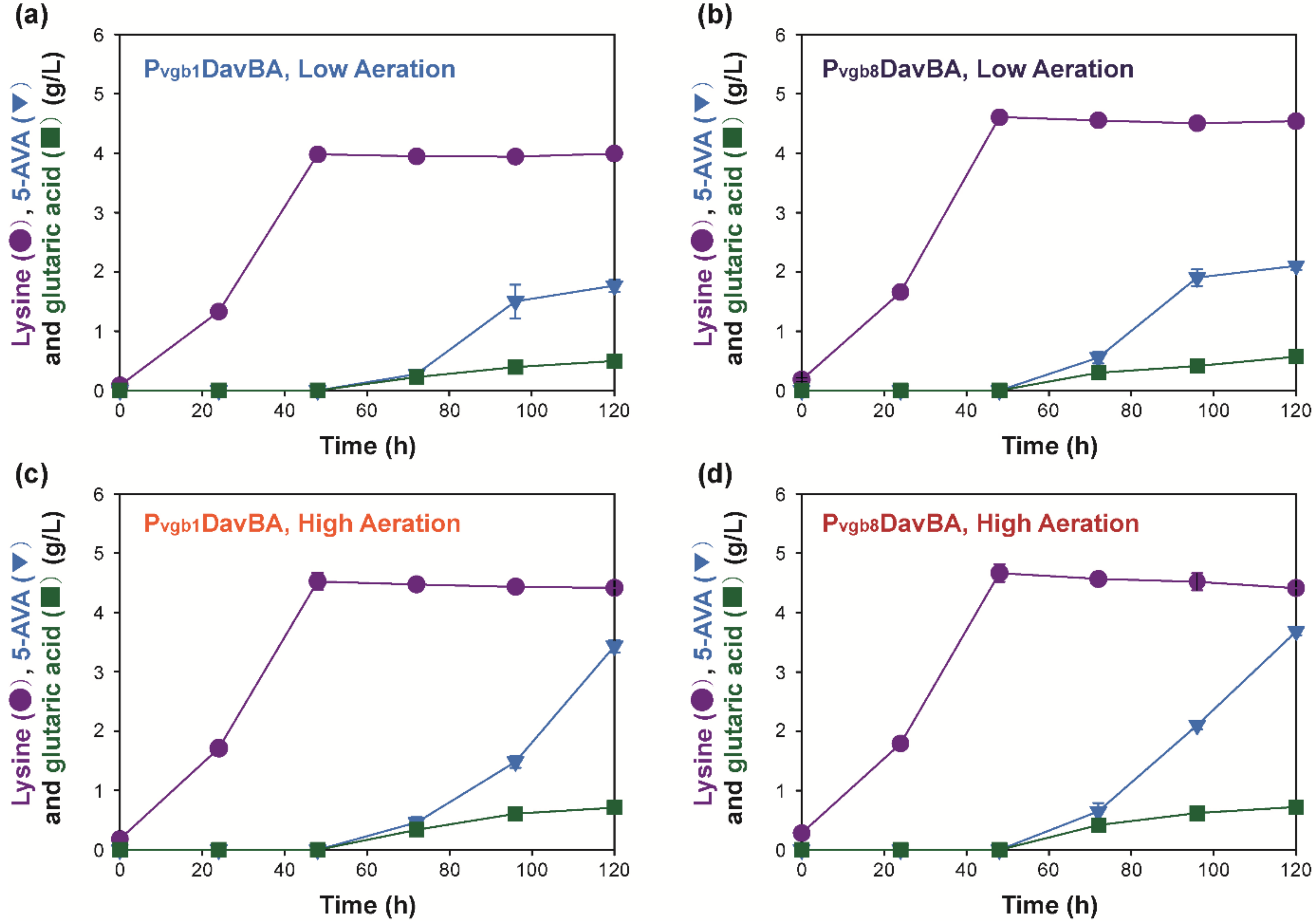
2.2. 5-AVA Production Using Recombinant C. glutamicum with Tunable Pvgb Promoter-Based Expression Systems
2.3. Gamma-Aminobutyric Acid Production Using Recombinant C. glutamicum with Tunable Pvgb Promoter-Based Expression Systems
3. Materials and Methods
3.1. Bacterial Strains and Plasmids
3.2. Plasmid Construction
3.3. Culture Conditions
3.4. Analysis
3.5. Fluorescence-Activated cell Sorting Analysis (FACS) for Measuring GFP Expression by Recombinant Strains
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Becker, J.; Wittmann, C. Advanced Biotechnology: Metabolically Engineered Cells for the Bio-Based Production of Chemicals and Fuels, Materials, and Health-Care Products. Angew. Chem. Int. Ed. 2015, 54, 3328–3350. [Google Scholar] [CrossRef] [PubMed]
- Oh, Y.H.; Eom, I.Y.; Joo, J.C.; Yu, J.H.; Song, B.G.; Lee, S.H.; Hong, S.H.; Park, S.J. Recent advances in development of biomass pretreatment technologies used in biorefinery for the production of bio-based fuels, chemicals and polymers. Korean J. Chem. Eng. 2015, 32, 1945–1959. [Google Scholar] [CrossRef]
- Lee, J.W.; Kim, H.U.; Choi, S.; Yi, J.H.; Lee, S.Y. Microbial production of building block chemicals and polymers. Curr. Opin. Biotechnol. 2011, 22, 758–767. [Google Scholar] [CrossRef] [PubMed]
- Baritugo, K.; Kim, H.T.; David, Y.; Choi, J.H.; Choi, J.; Kim, T.W.; Park, C.; Hong, S.H.; Na, J.G.; Jeong, K.J.; et al. Recent advances in metabolic engineering of Corynebacterium glutamicum strains as potential platform microorganisms for biorefinery. Biofuel Bioprod. Biorefin. 2018, 12, 899–925. [Google Scholar] [CrossRef]
- Baritugo, K.; Kim, H.T.; David, Y.; Choi, J.; Hong, S.H.; Jeong, K.J.; Joo, J.C.; Park, S.J. Metabolic engineering of Corynebacterium glutamicum for fermentative production of chemicals in biorefinery. Appl. Microbiol. Biotechnol. 2018, 102, 3915–3937. [Google Scholar] [CrossRef] [PubMed]
- Joo, J.C.; Oh, Y.H.; Yu, J.H.; Hyun, S.M.; Khang, T.U.; Kang, K.H.; Song, B.K.; Park, K.; Oh, M.K.; Lee, S.Y.; et al. Production of 5-aminovaleric acid in recombinant Corynebacterium glutamicum strains from a Miscanthus hydrolysate solution prepared by a newly developed Miscanthus hydrolysis process. Bioresour. Technol. 2017, 244, 1692–1700. [Google Scholar] [CrossRef] [PubMed]
- David, Y.; Joo, J.C.; Yang, J.E.; Oh, Y.H.; Lee, S.Y.; Park, S.J. Biosynthesis of 2-hydroxyacid-containing polyhydroxyalkanoates by employing butyryl-CoA transferases in metabolically engineered Escherichia coli. Biotechnol. J. 2017, 12. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.Y.; Parks, S.J.; Kim, W.J.; Yang, J.E.; Lee, H.; Shin, J.; Lee, S.Y. One-step fermentative production of poly(lactate-co-glycolate) from carbohydrates in Escherichia coli. Nat. Biotechnol. 2016, 34, 435–440. [Google Scholar] [CrossRef] [PubMed]
- Kind, S.; Neubauer, S.; Becker, J.; Yamamoto, M.; Völkert, M.; Abendroth, G.; Zelder, O.; Wittmann, C. From zero to hero—Production of bio-based nylon from renewable re-sources using engineered Corynebacterium glutamicum. Metab. Eng. 2014, 25, 113–123. [Google Scholar] [CrossRef] [PubMed]
- Chae, C.G.; Kim, Y.J.; Lee, S.J.; Oh, Y.H.; Yang, J.E.; Joo, J.C.; Kang, K.H.; Jang, Y.A.; Lee, H.; Park, A.R.; et al. Biosynthesis of poly(2-hydroxybutyrate-co-lactate) in metabolically engineered Escherichia coli. Biotechnol. Bioprocess Eng. 2016, 21, 169–174. [Google Scholar] [CrossRef]
- David, Y.; Baylon, M.G.; Sudheer, P.D.V.N.; Baritugo, K.; Chae, C.G.; Kim, Y.J.; Kim, T.W.; Kim, M.; Na, J.G.; Park, S.J. Screening of microorganisms able to degrade low-rank coal in aerobic conditions: Potential coal bio-solubilization mediators from coal to biochemical. Biotechnol. Bioprocess Eng. 2017, 22, 178–185. [Google Scholar] [CrossRef]
- Baylon, G.; David, Y.; Pamidimarri, S.; Baritugo, K.; Chae, C.G.; Kim, Y.J.; Wan, T.W.; Kim, M.S.; Na, J.G.; Park, S.J. Bio-solubilization of the untreated low rank coal by alkali-producing bacteria isolated from soil. Korean J. Chem. Eng. 2016, 34, 105–109. [Google Scholar] [CrossRef]
- Sudheer, P.D.V.N.; David, Y.; Chae, C.; Kim, Y.J.; Baylon, M.G.; Baritugo, K.; Kim, T.W.; Kim, M.; Na, J.G.; Park, S.J. Advances in the biological treatment of coal for synthetic natural gas and chemicals. Korean J. Chem. Eng. 2016, 10, 2788–2801. [Google Scholar] [CrossRef]
- Yang, J.E.; Park, S.J.; Kim, W.J.; Kim, H.J.; Kim, B.; Lee, H.; Shin, J.; Lee, S.Y. One-step fermentative production of aromatic polyesters from glucose by metabolically engineered Escherichia coli strains. Nat. Commun. 2018, 9, 79. [Google Scholar] [CrossRef] [PubMed]
- Joo, J.C.; Khusnutdinova, A.N.; Flick, R.; Kim, T.H.; Bornscheuer, U.T.; Yakunin, A.F.; Mahadevan, R. Alkene hydrogenation activity of enoate reductases for an environmentally benign biosynthesis of adipic acid. Chem. Sci. 2017, 8, 1406–1413. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Lama, S.; Kim, J.R.; Park, S.H. Production of 1,3-Propanediol from Glucose by Recombinant Escherichia coli BL21(DE3). Biotechnol. Bioprocess Eng. 2017, 23, 250–258. [Google Scholar] [CrossRef]
- Li, J.; Feng, R.; Wen, Z.; Zhang, A. Overexpression of ARO10 in pdc5Δmutant resulted in higher isobutanol titers in Saccharomyces cerevisiae. Biotechnol. Bioprocess Eng. 2017, 22, 382–389. [Google Scholar] [CrossRef]
- Zong, H.; Liu, X.; Chen, W.; Zhuge, B.; Sun, J. Construction of glycerol synthesis pathway in Klebsiella pneumoniae for bioconversion of glucose into 1,3-propanediol. Biotechnol. Bioprocess Eng. 2017, 22, 549–555. [Google Scholar] [CrossRef]
- Kim, H.S.; Oh, Y.H.; Jang, Y.; Kang, K.H.; David, Y.; Yu, J.H.; Song, B.K.; Choi, J.; Chang, Y.K.; Joo, J.C.; et al. Recombinant Ralstonia eutropha engineered to utilize xylose and its use for the production of poly(3-hydroxybutyrate) from sunflower stalk hydrolysate solution. Microb. Cell Factories 2016, 15, 95. [Google Scholar] [CrossRef] [PubMed]
- Park, S.J.; Jang, Y.A.; Noh, W.; Oh, Y.H.; Lee, H.; David, Y.; Baylon, M.G.; Shin, J.; Yang, J.E.; Choi, S.Y.; et al. Metabolic engineering of Ralstonia eutropha for the production of polyhydroxyalkanoates from sucrose. Biotechnol. Bioeng. 2015, 112, 638–643. [Google Scholar] [CrossRef] [PubMed]
- Pang, X.; Zhuang, X.; Tang, Z.; Chen, X. Polylactic acid (PLA): Research, development and industrialization. Biotechnol. J. 2010, 5, 1125–1136. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Guo, B.H. Poly (butylene succinate) and its copolymers: Research, development and industrialization. Biotechnol. J. 2010, 5, 1149–1163. [Google Scholar] [CrossRef] [PubMed]
- Kind, S.; Wittmann, C. Bio-based production of the platform chemical 1, 5-diaminopentane. Appl. Microbiol. Biotechnol. 2011, 91, 1287–1296. [Google Scholar] [CrossRef] [PubMed]
- Park, S.J.; Kim, E.Y.; Noh, W.; Park, H.M.; Oh, Y.H.; Lee, S.H.; Song, B.K.; Jegal, J.; Lee, S.Y. Metabolic engineering of Escherichia coli for the production of 5-aminovalerate and glutarate as C5 platform chemicals. Metab. Eng. 2013, 16, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.H.; Park, S.H.; Oh, Y.H.; Choi, J.W.; Lee, M.H.; Cho, J.S.; Jeong, K.J.; Joo, J.C.; Yu, J.; Park, S.J.; et al. Metabolic engineering of Corynebacterium glutamicum for enhanced production of 5-aminovaleric acid. Microb. Cell Factories 2016, 15, 174. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.W.; Yim, S.S.; Lee, S.H.; Kang, T.J.; Park, S.J.; Jeong, K.J. Enhanced production of gamma-aminobutyrate (GABA) in recombinant Corynebacterium glutamicum by expressing glutamate decarboxylase active in expanded pH range. Microb. Cell Factories 2015, 14, 21. [Google Scholar] [CrossRef] [PubMed]
- Park, S.J.; Kim, E.Y.; Won, N.; Oh, Y.H.; Kim, H.Y.; Song, B.K.; Cho, K.M.; Hong, S.H.; Lee, S.H.; Jegal, J. Synthesis of nylon 4 from gamma-aminobutyrate (GABA) produced by recombinant Escherichia coli. Bioprocess Biosyst. Eng. 2013, 36, 885–892. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.T.; Baritugo, K.A.; Oh, Y.H.; Hyun, S.M.; Khang, T.U.; Kang, K.H.; Jung, S.H.; Song, B.K.; Park, K.; Kim, I.K.; et al. Metabolic engineering of Corynebacterium glutamicum for the high-level production of cadaverine that can be used for synthesis of biopolyamide 510. ACS Sustain. Chem. Eng. 2018, 6, 5296–5305. [Google Scholar] [CrossRef]
- Oh, Y.H.; Choi, J.W.; Kim, E.Y.; Song, B.K.; Jeong, K.J.; Park, K.; Kim, I.; Woo, H.M.; Lee, S.H.; Park, S.J. Construction of Synthetic Promoter-Based Expression Cassettes for the production of Cadaverine in Recombinant Corynebacterium glutamicum. Appl. Biochem. Biotech. 2015, 176, 2065–2075. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.T.; Kang, T.U.; Baritugo, K.; Hyun, S.M.; Kang, K.H.; Jung, S.H.; Song, B.K.; Park, K.; Oh, M.; Kim, G.B.; et al. Metabolic engineering of Corynebacterium glutamicum for the production of glutaric acid, a C5 dicarboxylic acid platform chemical. Metab. Eng. 2018. [Google Scholar] [CrossRef] [PubMed]
- Baritugo, K.; Kim, H.T.; David, Y.; Khang, T.U.; Hyun, S.M.; Kang, K.H.; Yu, J.H.; Choi, J.H.; Song, J.J.; Joo, J.C.; et al. Enhanced production of gamma-aminobutyrate (GABA) in recombinant Corynebacterium glutamicum strains from empty fruit bunch biosugar solution. Microb. Cell Factories 2018, 17, 129. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Wang, J.; Geng, Y.; Li, Y.; Wang, Q.; Liang, Q.; Qi, Q. A strategy of gene overexpression based on tandem repetitive promoters in Escherichia coli. Microb. Cell Factories 2012, 11, 19. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Wang, H.; Chen, J.; Chen, G.Q. Effects of cascaded vgb promoters on poly(hydroxybutyrate) (PHB) synthesis by recombinant Escherichia coli grown micro-aerobically. Appl. Microbiol. Biotechnol. 2014, 98, 10013–10021. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Zhang, J.; Wei, X.X.; Ouyang, S.P.; Wu, Q.; Chen, G.Q. Microbial production of L-glutamate and L-glutamine by recombinant Corynebacterium glutamicum harboring Vitreoscilla hemoglobin gene vgb. Appl. Microbiol. Biotechnol. 2008, 77, 1297–1304. [Google Scholar] [CrossRef] [PubMed]
- Ko, K.; Lee, B.; Cheong, D.; Han, Y.; Choi, J.H.; Song, J.J. Bacterial Cell Surface Display of a Multifunctional Cellulolytic Enzyme Screened from a Bovine Rumen Metagenomic Resource. J. Microbiol. Biotechnol. 2015, 25, 1835–1841. [Google Scholar] [CrossRef] [PubMed]
- Sambrook, J.; Russell, D.W. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2001; Volume 3. [Google Scholar]
- Cabezudo, M.D.; Hermosín, I.; Chicón, R.M. Free amino acid composition and botanical origin of honey. Food Chem. 2003, 83, 263–268. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains and Plasmids | Relevant Characteristics | Reference |
|---|---|---|
| Strains | ||
| E. coli XL1-Blue | recA1 endA1 gyrA96 thi-1 hsdR17 supE44 relA1 lac [FA1proAB lacIqZΔM15 Tn10 (TetR)] | Stratagene |
| C. glutamicum KCTC 1447 | L-Glutamate producer | KCTC |
| C. glutamicum KCTC 1852 | L-Glutamate producer | KCTC |
| C. glutamicum KCTC 1857 | L-Lysine producer | KCTC |
| C. glutamicum ATCC 13032 | Wild type | ATCC |
| EGFPV1 | C. glutamicum KCTC 1857 with pCES208V:eGFP | This study |
| EGFPV4 | C. glutamicum KCTC 1857 with pCES208V4:eGFP | This study |
| EGFPV8 | C. glutamicum KCTC 1857 with pCES208V8:eGFP | This study |
| 5AVA1 | C. glutamicum KCTC 1857 with pCES208V:DavBA | This study |
| 5AVA8 | C. glutamicum KCTC 1857 with pCES208V8:DavBA | This study |
| 5AVA1His | C. glutamicum KCTC 1857 with pCES208V:DavBHisDavA | This study |
| 5AVA8His | C. glutamicum KCTC 1857 with pCES208V8:DavBHisDavA | This study |
| H30_AVA | C. glutamicum KCTC 1857 with pCES208H30:DavBA | 30 |
| H30_AVAHis | C. glutamicum KCTC 1857 with pCES208H30:DavBHisA | 30 |
| H36GM13032 | C. glutamicum ATCC 13032 with pCES208H36:GadBmut | This study |
| H36GM1447 | C. glutamicum KCTC 1447 with pCES208H36:GadBmut | This study |
| H36GM1852 | C. glutamicum KTC 1852 with pCES208H36:GadBmut | This study |
| H30GM13032 | C. glutamicum ATCC 13032 with pCES208H30:GadBmut | This study |
| H30GM1447 | C. glutamicum KCTC 1447 with pCES208H30:GadBmut | This study |
| H30GM1852 | C. glutamicum KTC 1852 with pCES208H30:GadBmut | This study |
| V1GD133032 | C. glutamicum ATCC 13032 with pCES208V:GadBmut | This study |
| V1GD1447 | C. glutamicum KCTC 1447 with pCES208V:GadBmut | This study |
| V1GD1852 | C. glutamicum KTC 1852 with pCES208V:GadBmut | This study |
| V8GD133032 | C. glutamicum ATCC 13032 with pCES208V8:GadBmut | This study |
| V8GD1447 | C. glutamicum KCTC 1447 with pCES208V8:GadBmut | This study |
| V8GD1852 | C. glutamicum KTC 1852 with pCES208V8:GadBmut | This study |
| Plasmids | ||
| pCES208V:eGFP | pCES208 derivative; Pvgb promoter, eGFP, KmR | This study |
| pCES208V4:eGFP | pCES208 derivative; Pvgb4 promoter, eGFP, KmR | This study |
| pCES208V8:eGFP | pCES208 derivative; Pvgb8 promoter, eGFP, KmR | This study |
| pCES208V:DavBA | pCES208 derivative; Pvgb promoter, Pseudomonas putida KT2440 davBA genes, KmR | This study |
| pCES208V8:DavBA | pCES208 derivative; Pvgb8 promoter, Pseudomonas putida KT2440 davBA genes, KmR | This study |
| pCES208V:DavBHisA | pCES208 derivative; Pvgb promoter, Pseudomonas putida KT2440 davBHis and davA genes, KmR | This study |
| pCES208V8:DavBHisA | pCES208 derivative; Pvgb8 promoter, Pseudomonas putida KT2440 davBHis and davA genes, KmR | This study |
| pCES208H30:DavBA | pCES208 derivative; PH30 promoter, Pseudomonas putida KT2440 davBA genes, KmR | 30 |
| pCES208H30:DavBHisA | pCES208 derivative; PH30 promoter, Pseudomonas putida KT2440 davBHis and davA genes, KmR | 30 |
| pCES208V:GadBmut | pCES208 derivative; Pvgb promoter, E. coli mutated gadB (Glu89Gln/Δ452-466) gene, KmR | This study |
| pCES208V8:GadBmut | pCES208 derivative; Pvgb8 promoter, E. coli mutated gadB (Glu89Gln/Δ452-466) gene, KmR | This study |
| pHGmut / pCES208H36:GadBmut | pCES208 derivative; PH36 promoter, E. coli mutated gadB (Glu89Gln/Δ452-466) gene, KmR | 26 |
| pCES208H30:GadBmut | pCES208 derivative; PH30 promoter, E. coli mutated gadB (Glu89Gln/Δ452-466) gene, KmR | This study |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baritugo, K.-A.; Kim, H.T.; Rhie, M.N.; Jo, S.Y.; Khang, T.U.; Kang, K.H.; Song, B.K.; Lee, B.; Song, J.J.; Choi, J.H.; et al. Construction of a Vitreoscilla Hemoglobin Promoter-Based Tunable Expression System for Corynebacterium glutamicum. Catalysts 2018, 8, 561. https://doi.org/10.3390/catal8110561
Baritugo K-A, Kim HT, Rhie MN, Jo SY, Khang TU, Kang KH, Song BK, Lee B, Song JJ, Choi JH, et al. Construction of a Vitreoscilla Hemoglobin Promoter-Based Tunable Expression System for Corynebacterium glutamicum. Catalysts. 2018; 8(11):561. https://doi.org/10.3390/catal8110561
Chicago/Turabian StyleBaritugo, Kei-Anne, Hee Taek Kim, Mi Na Rhie, Seo Young Jo, Tae Uk Khang, Kyoung Hee Kang, Bong Keun Song, Binna Lee, Jae Jun Song, Jong Hyun Choi, and et al. 2018. "Construction of a Vitreoscilla Hemoglobin Promoter-Based Tunable Expression System for Corynebacterium glutamicum" Catalysts 8, no. 11: 561. https://doi.org/10.3390/catal8110561
APA StyleBaritugo, K.-A., Kim, H. T., Rhie, M. N., Jo, S. Y., Khang, T. U., Kang, K. H., Song, B. K., Lee, B., Song, J. J., Choi, J. H., Lee, D.-H., Joo, J. C., & Park, S. J. (2018). Construction of a Vitreoscilla Hemoglobin Promoter-Based Tunable Expression System for Corynebacterium glutamicum. Catalysts, 8(11), 561. https://doi.org/10.3390/catal8110561