
Rhizobium’s Reductase for Chromium Detoxification, Heavy Metal Resistance, and Artificial Neural Network-Based Predictive Modeling



Abstract
1. Introduction
2. Results
2.1. Screening of Potential Rhizobacteria
Heavy Metal Tolerance Capability of Strain OS-1
2.2. Optimum pH and Temperature for Heavy Metals Accumulation
2.3. The 16s rRNA Gene Sequencing and Characterization of Strain OS-1
2.4. Rhizobium OS-1 Strain Phylogenetic Tree Construction
2.5. Chromium (VI) Reduction Assessment
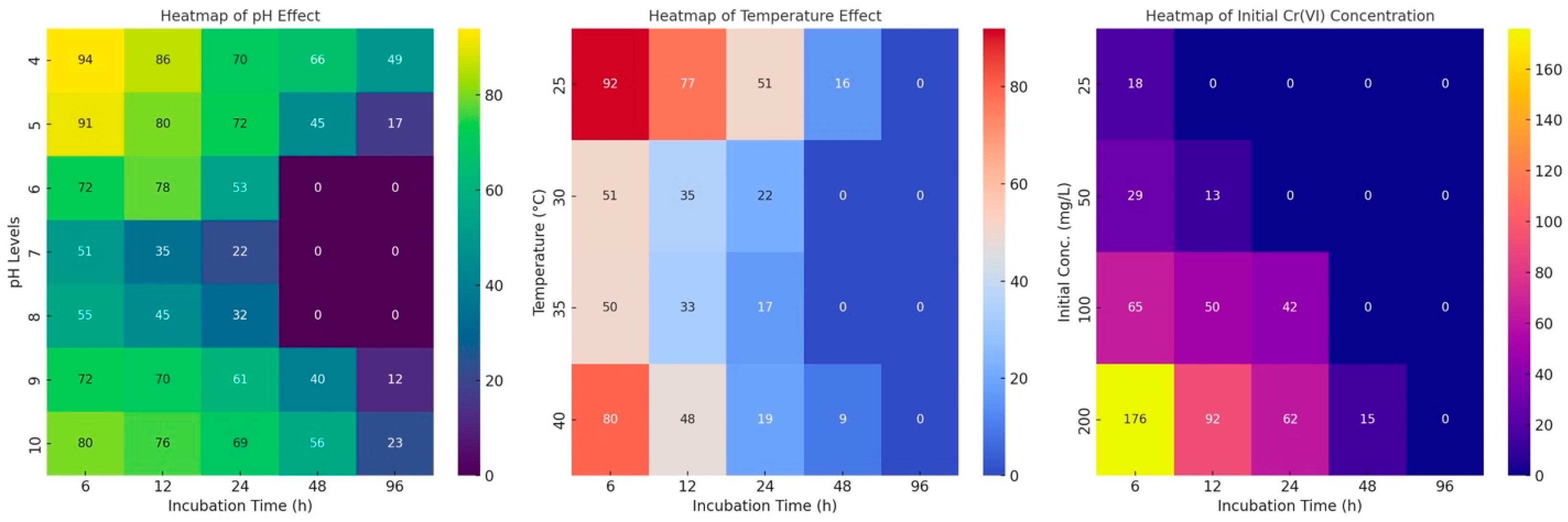
2.5.1. Chromium Initial Concentration, pH, and Temperature Effect on Cr(VI) Reduction
2.5.2. Optimum pH and Temperature for Heavy Metal Accumulation
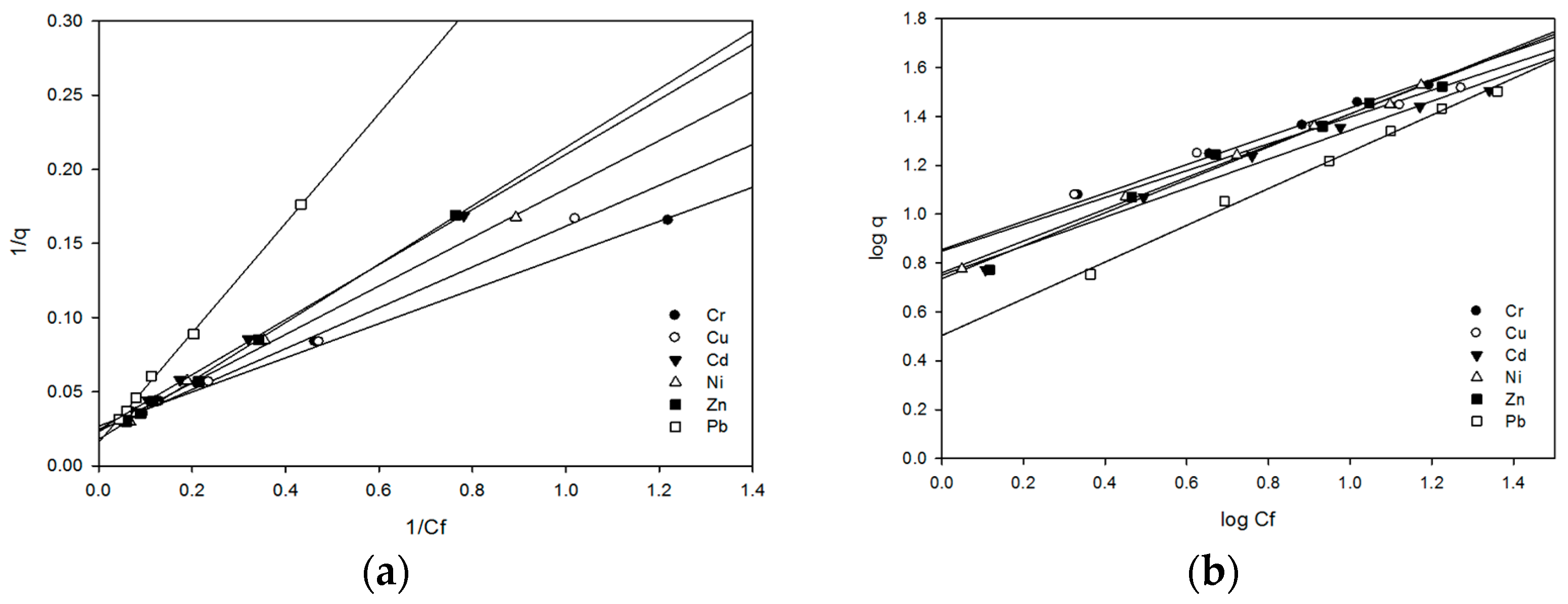
2.5.3. Biosorption Profile of Bacterial Isolate OS-1
2.6. Chromium Reduction Results at Variable Parameters
2.6.1. Effect of pH on Chromium Reduction
2.6.2. Effect of Temperature on Chromium Reduction
2.6.3. Effect of Contact Time on Chromium (VI) Reduction
2.6.4. At Variable Initial Cr(VI) Concentration
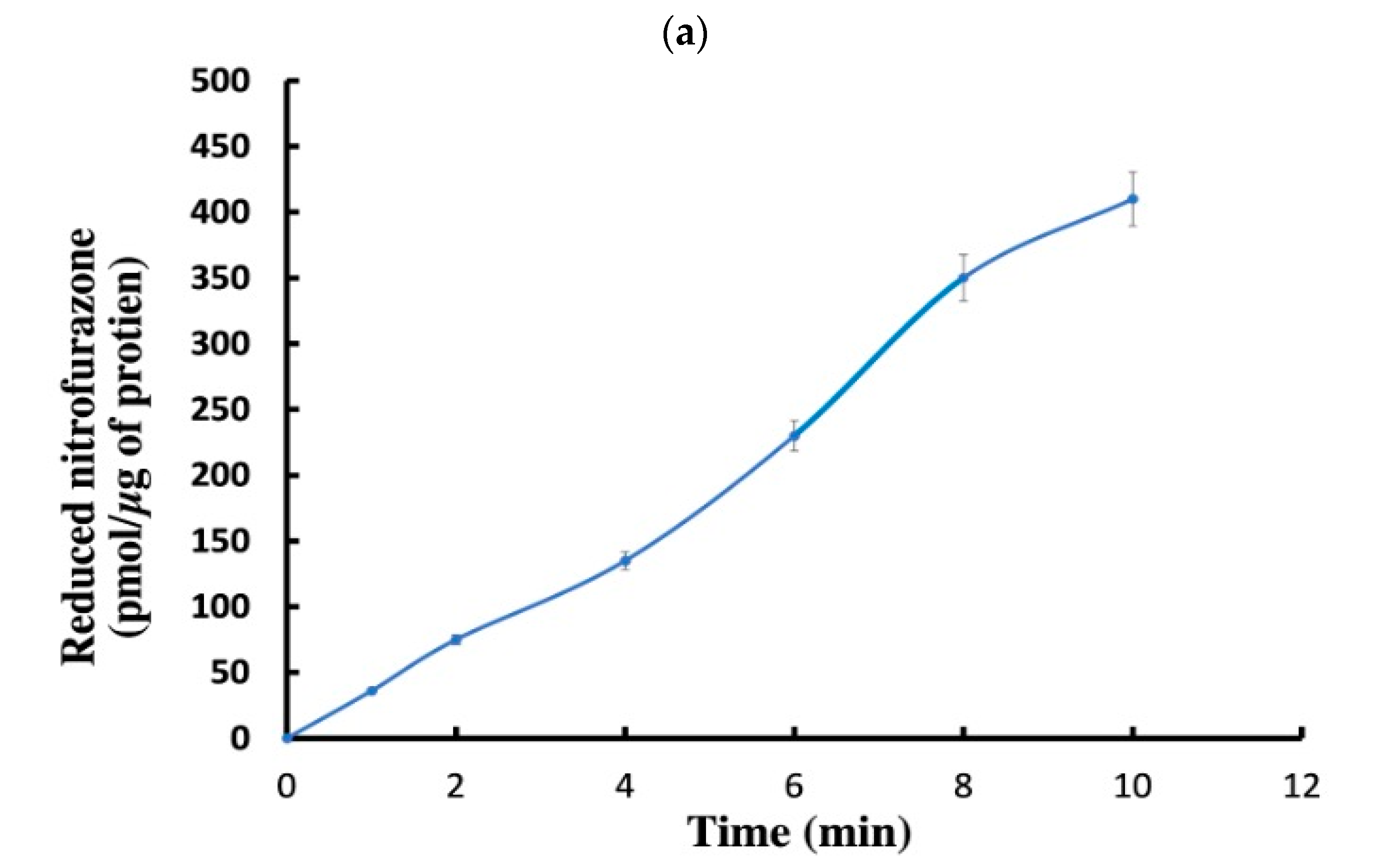
2.6.5. Analysis of Nitroreductase and Chromate Reductase Activity
Nitroreductase Activity Assay
The Period for TNT Biotransformation
Comparison of Chromate Reductase Activity
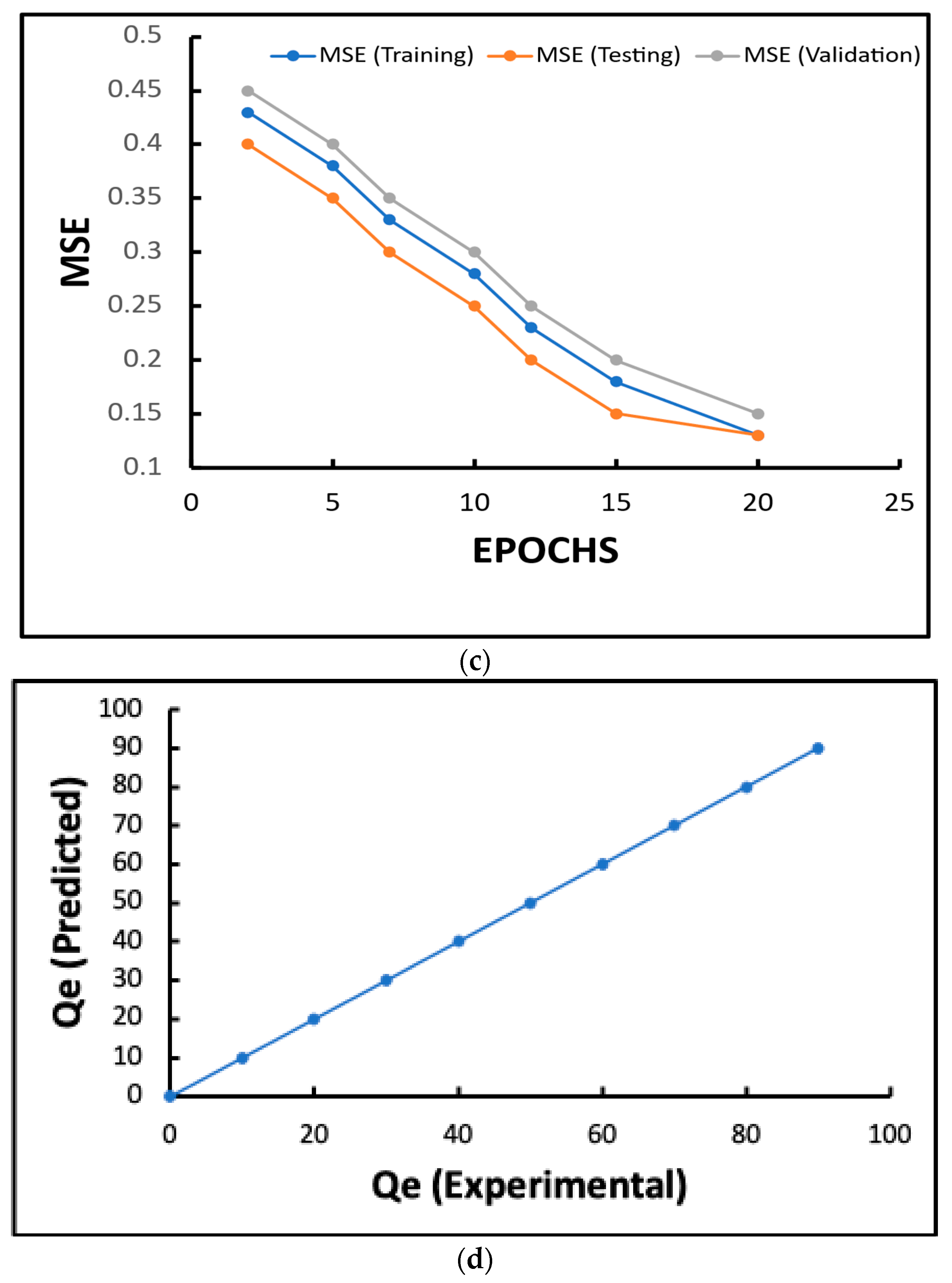
2.7. Detailed Results of Artificial Neural Network (ANN) Prediction
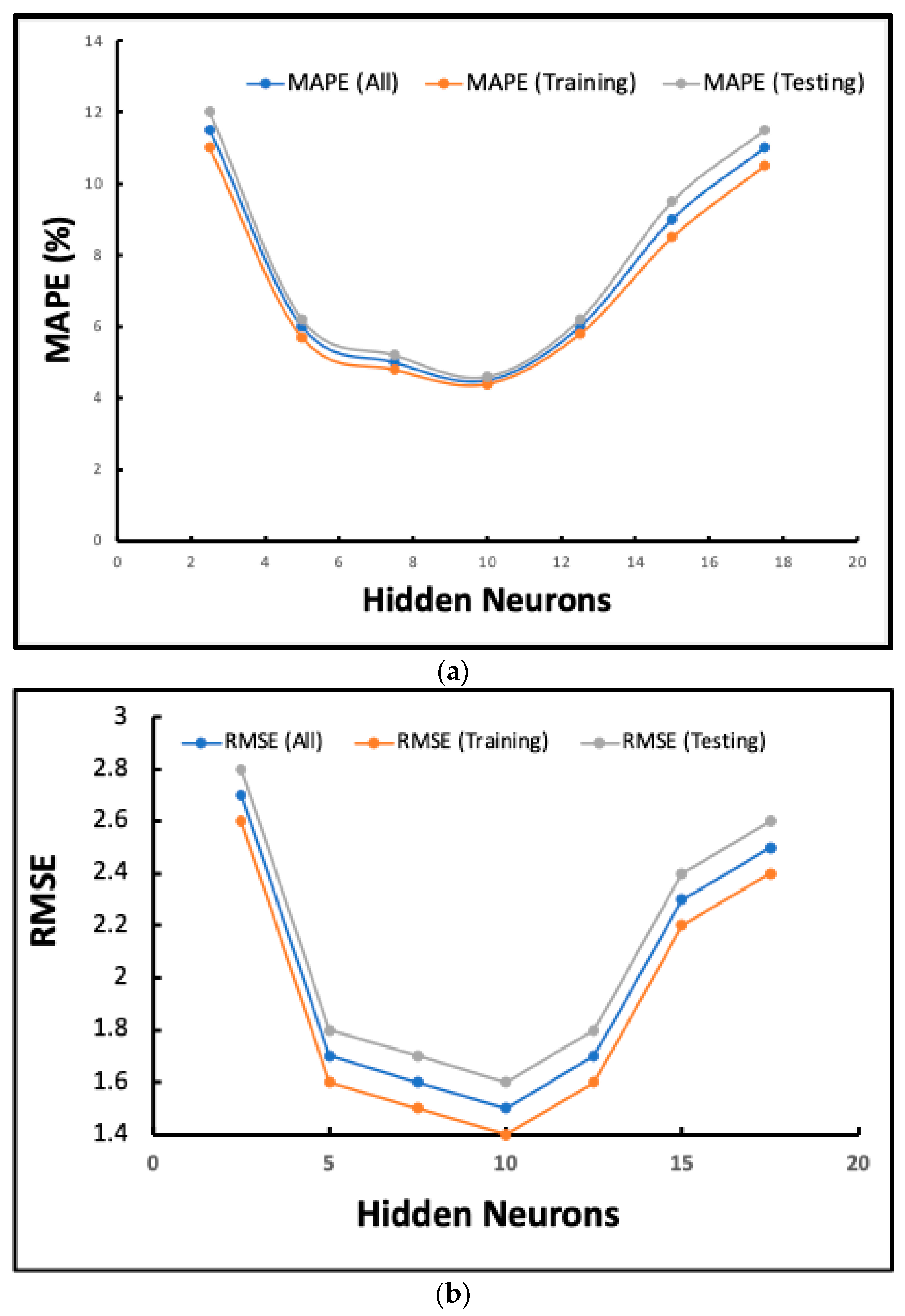
2.7.1. Prediction Accuracy
2.7.2. MAPE and Error Analysis
2.7.3. The Predictions of Incubation Time and Initial Concentration
2.7.4. Robustness of Model
Observed Trends
Key Insights
3. Discussion
4. Materials and Methods
4.1. Sample Collection
4.2. Screening of Metal-Tolerant N-Fixing and P-Solubilizing Bacteria
Protocol for Biochemical Testing of Bacterial Isolate
4.3. Sequencing of the 16S rRNA Gene and Assessment of Phylogenetic Tree
4.4. Bioremediation Studies
4.4.1. Chromium Reduction
4.4.2. Chromium Concentration and pH Effect on Reduction
4.4.3. Temperature Effect on Reduction
4.4.4. Biosorption Study
Development of Stock Solution and Bacterial Biosorbent
Biosorption by Rhizobium Biomass
Isotherm Freundlich and Langmuir
Metal Ions Separation Factor (Sf) and Surface Coverage (Ø)
4.4.5. Analysis of Nitroreductase and Chromate Reductase Activity
4.5. A Neural Network Model for Predicting Chromium Reduction
4.5.1. Model Sensitivity Analysis
4.5.2. Statistical Analysis
5. Conclusions
Recommendation for Future Research
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sharma, N.; Sodhi, K.K.; Kumar, M.; Singh, D.K. Heavy metal pollution: Insights into chromium eco-toxicity and recent advancement in its remediation. Environ. Nanotechnol. Monit. Manag. 2021, 15, 100388. [Google Scholar] [CrossRef]
- Kolopajlo, L. A review of sustainable leather tanning. Green Chem. Process. Dev. Sci. Math Eng. Technol. 2024, 11, 67. [Google Scholar]
- Xia, S.; Song, Z.; Jeyakumar, P.; Shaheen, S.M.; Rinklebe, J.; Ok, Y.S.; Bolan, N.; Wang, H. A critical review on bioremediation technologies for Cr(VI)-contaminated soils and wastewater. Crit. Rev. Environ. Sci. Technol. 2019, 49, 1027–1078. [Google Scholar] [CrossRef]
- Staszak, K.; Kruszelnicka, I.; Ginter-Kramarczyk, D.; Góra, W.; Baraniak, M.; Lota, G.; Regel-Rosocka, M. Advances in the removal of Cr(III) from spent industrial effluents—A review. Materials 2022, 16, 378. [Google Scholar] [CrossRef] [PubMed]
- Oves, M.; Khan, M.S.; Zaidi, A. Chromium reducing and plant growth promoting novel strain Pseudomonas aeruginosa OSG41 enhance chickpea growth in chromium amended soils. Eur. J. Soil Biol. 2013, 56, 72–83. [Google Scholar] [CrossRef]
- Karthik, C.; Elangovan, N.; Kumar, T.S.; Govindharaju, S.; Barathi, S.; Oves, M.; Arulselvi, P.I. Characterization of multifarious plant growth promoting traits of rhizobacterial strain AR6 under Chromium (VI) stress. Microbiol. Res. 2017, 204, 65–71. [Google Scholar] [CrossRef]
- Jaiswal, S. Bioremediation of Chromium contamination by Aspergillus and Rhizopus. Res. J. Sci. Technol. 2024, 16, 270–273. [Google Scholar] [CrossRef]
- Valentine, A.J.; Benedito, V.A.; Kang, Y. Legume nitrogen fixation and soil abiotic stress: From physiology to genomics and beyond. In Nitrogen Metabolism in Plants in the Post-Genomic Era; Foyer, C., Zhang, H., Eds.; Wiley-Blackwell Publishing Ltd.: West Sussex, UK, 2010; Volume 42. [Google Scholar]
- Liu, Y.; He, G.; He, T.; Saleem, M. Signaling and detoxification strategies in plant-microbes symbiosis under heavy metal stress: A mechanistic understanding. Microorganisms 2022, 11, 69. [Google Scholar] [CrossRef] [PubMed]
- Upadhyay, S.K.; Rajput, V.D.; Kumari, A.; Espinosa-Saiz, D.; Menendez, E.; Minkina, T.; Dwivedi, P.; Mandzhieva, S. Plant growth-promoting rhizobacteria: A potential bio-asset for restoration of degraded soil and crop productivity with sustainable emerging techniques. Environ. Geochem. Health 2023, 45, 9321–9344. [Google Scholar] [CrossRef]
- Palai, J.B.; Malik, G.C.; Maitra, S.; Banerjee, M. Role of Rhizobium on growth and development of groundnut: A review. Int. J. Agric. Environ. Biotechnol. 2021, 14, 63–73. [Google Scholar] [CrossRef]
- Fahde, S.; Boughribil, S.; Sijilmassi, B.; Amri, A. Rhizobia: A promising source of plant growth-promoting molecules and their non-legume interactions: Examining applications and mechanisms. Agriculture 2023, 13, 1279. [Google Scholar] [CrossRef]
- Ahirwar, N.K.; Singh, R.; Chaurasia, S.; Chandra, R.; Ramana, S. Effective role of beneficial microbes in achieving the sustainable agriculture and eco-friendly environment development goals: A review. Front. Microbiol 2020, 5, 111–123. [Google Scholar] [CrossRef]
- Bergey, D.H. Bergey’s Manual of Determinative Bacteriology; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 1994. [Google Scholar]
- Sahgal, M.; Jaggi, V. Rhizobia: Culture Collections, Identification, and Methods of Preservation. In Microbial Resource Conservation: Conventional to Modern Approaches; Springer International Publishing: Cham, Switzerland, 2018; pp. 175–197. [Google Scholar]
- Alves, L.C.; De Souza, J.A.M.; de Mello Varani, A.; de Macedo Lemos, E.G. The Family Rhizobiaceae; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 419–437. [Google Scholar]
- Abdian, P.L.; Zorreguieta, A. Extracellular factors involved in biofilm matrix formation by Rhizobium leguminosarum. In The Perfect Slime–Microbial Extracellular Polymeric Substances (EPS); IWA Publishing Group: London, UK, 2016; pp. 227–247. [Google Scholar]
- O’Brian, M.R. Heme synthesis in the rhizobium-legume symbiosis: A palette for bacterial and eukaryotic pigments. J. Bacteriol. 1996, 178, 2471–2478. [Google Scholar] [CrossRef] [PubMed]
- Kuypers, M.M.; Marchant, H.K.; Kartal, B. The microbial nitrogen-cycling network. Nat. Rev. Microbiol. 2018, 16, 263–276. [Google Scholar] [CrossRef]
- Ransy, C.; Vaz, C.; Lombès, A.; Bouillaud, F. Use of H2O2 to cause oxidative stress, the catalase issue. Int. J. Mol. Sci. 2020, 21, 9149. [Google Scholar] [CrossRef]
- Alleman, A.B.; Peters, J.W. Mechanisms for generating low potential electrons across the metabolic diversity of nitrogen-fixing bacteria. Appl. Environ. Microbiol. 2023, 89, e00378-23. [Google Scholar] [CrossRef]
- Johnson, J.S.; Spakowicz, D.J.; Hong, B.Y.; Petersen, L.M.; Demkowicz, P.; Chen, L.; Leopold, S.R.; Hanson, B.M.; Agresta, H.O.; Gerstein, M.; et al. Evaluation of 16S rRNA gene sequencing for species and strain-level microbiome analysis. Nat. Commun. 2019, 10, 5029. [Google Scholar] [CrossRef]
- Mitra, S.; Stärk, M.; Huson, D.H. Analysis of 16S rRNA environmental sequences using MEGAN. In BMC Genomics; BioMed Central: New York, NY, USA, 2011; Volume 12, pp. 1–7. [Google Scholar]
- Faniyan, O.; Akpe, V.; Cock, I.E. Analyzing bacterial species from different environments using direct 16S rRNA gene sequencing methods. Pharmacogn. Commun. 2023, 13, 24–33. [Google Scholar] [CrossRef]
- Keklik, G. Understanding evolutionary relationships and analysis methods through mega software. Int. J. New Horiz. Sci. 2023, 1, 83–90. [Google Scholar]
- Egamberdieva, D.; Shurigin, V.; Gopalakrishnan, S.; Sharma, R. Microbial strategies for the improvement of legume production in hostile environments. In Legumes Under Environmental Stress: Yield, Improvement and Adaptations; John Wiley & Sons: Hoboken, NJ, USA, 2015; pp. 133–144. [Google Scholar]
- Rosier, A.; Medeiros, F.H.; Bais, H.P. Defining plant growth promoting rhizobacteria molecular and biochemical networks in beneficial plant-microbe interactions. Plant Soil 2018, 428, 35–55. [Google Scholar] [CrossRef]
- Szewczuk-Karpisz, K.; Tomczyk, A.; Grygorczuk-Płaneta, K.; Naveed, S. Rhizobium leguminosarum bv. trifolii exopolysaccharide and sunflower husk biochar as factors affecting immobilization of both tetracycline and Cd2+ ions on soil solid phase. J. Soils Sediments 2022, 22, 2620–2639. [Google Scholar] [CrossRef]
- Jobby, R.; Jha, P.; Gupta, A.; Gupte, A.; Desai, N. Biotransformation of chromium by root nodule bacteria Sinorhizobium sp. SAR1. PLoS ONE 2019, 14, e0219387. [Google Scholar] [CrossRef]
- Luka, Y.; Highina, B.K.; Zubairu, A.; Adeleke, A.J.; Hamadou, M.; Musti, Y.A.; Abubakar, A.M.; Yunus, M.U. Biosorption as technique for remediation of heavy metals from wastewater using microbial biosorbent. Biol. Sci. 2024, 4, 564–574. [Google Scholar] [CrossRef]
- Micheal, H.S.R.; Thyagarajan, D.; Govindaraj, M.; Saravanakumar, V.K.; Mohammed, N.B.; Murugasamy Maheswari, K. Biosorption of halophilic fungal melanized membrane–PUR/melanin polymer for heavy metal detoxification with electrospinning technology. Environ. Technol. 2024, 45, 5865–5877. [Google Scholar] [CrossRef]
- Qin, H.; Wang, Z.; Sha, W.; Song, S.; Qin, F.; Zhang, W. Role of plant-growth-promoting rhizobacteria in plant machinery for soil heavy metal detoxification. Microorganisms 2024, 12, 700. [Google Scholar] [CrossRef]
- Xie, S. Biosorption of heavy metal ions from contaminated wastewater: An eco-friendly approach. Green Chem. Lett. Rev. 2024, 17, 2357213. [Google Scholar] [CrossRef]
- Alfadaly, R.A.; Elsayed, A.; Hassan, R.Y.; Noureldeen, A.; Darwish, H.; Gebreil, A.S. Microbial sensing and removal of heavy metals: Bioelectrochemical detection and removal of chromium (VI) and cadmium (II). Molecules 2021, 26, 2549. [Google Scholar] [CrossRef] [PubMed]
- Vasilica, S.T.A.N.; Gament, E.; Cornea, C.P.; Voaideş, C.; Mirela, D.U.Ş.A.; Plopeanu, G. Effects of heavy metal from polluted soils on the Rhizobium diversity. Not. Bot. Horti Agrobot. Cluj-Napoca 2011, 39, 88–95. [Google Scholar] [CrossRef]
- Fagorzi, C.; Checcucci, A.; DiCenzo, G.C.; Debiec-Andrzejewska, K.; Dziewit, L.; Pini, F.; Mengoni, A. Harnessing rhizobia to improve heavy-metal phytoremediation by legumes. Genes 2018, 9, 542. [Google Scholar] [CrossRef]
- Joshi, S.; Gangola, S.; Bhandari, G.; Bhandari, N.S.; Nainwal, D.; Rani, A.; Malik, S.; Slama, P. Rhizospheric bacteria: The key to sustainable heavy metal detoxification strategies. Front. Microbiol. 2023, 14, 1229828. [Google Scholar] [CrossRef] [PubMed]
- Khosravi, R.; Moussavi, G.; Ghaneian, M.T.; Ehrampoush, M.H.; Barikbin, B.; Ebrahimi, A.A.; Sharifzadeh, G. Chromium adsorption from aqueous solution using novel green nanocomposite: Adsorbent characterization, isotherm, kinetic and thermodynamic investigation. J. Mol. Liq. 2018, 256, 163–174. [Google Scholar] [CrossRef]
- Chidambaram, R. Isotherm modelling, kinetic study and optimization of batch parameters using response surface methodology for effective removal of Cr(VI) using fungal biomass. PLoS ONE 2015, 10, e0116884. [Google Scholar]
- Basnet, P.; Gyawali, D.; Ghimire, K.N.; Paudyal, H. An assessment of the lignocellulose-based biosorbents in removing Cr(VI) from contaminated water: A critical review. Results Chem. 2022, 4, 100406. [Google Scholar] [CrossRef]
- Yan, G.; Gao, Y.; Xue, K.; Qi, Y.; Fan, Y.; Tian, X.; Wang, J.; Zhao, R.; Zhang, P.; Liu, Y.; et al. Toxicity mechanisms and remediation strategies for chromium exposure in the environment. Front. Environ. Sci. 2023, 11, 1131204. [Google Scholar] [CrossRef]
- Serrano-González, M.Y.; Chandra, R.; Castillo-Zacarias, C.; Robledo-Padilla, F.; Rostro-Alanis, M.D.J.; Parra-Saldivar, R. Biotransformation and degradation of 2, 4, 6-trinitrotoluene by microbial metabolism and their interaction. Def. Technol. 2018, 14, 151–164. [Google Scholar] [CrossRef]
- Ryberg, D.; Alexander, J. Inhibitory action of hexavalent chromium (Cr(VI)) on the mitochondrial respiration and a possible coupling to the reduction of Cr(VI). Biochem. Pharmacol. 1984, 33, 2461–2466. [Google Scholar] [CrossRef]
- Rahman, Z.; Thomas, L.; Chetri, S.P.; Bodhankar, S.; Kumar, V.; Naidu, R. A comprehensive review on chromium (Cr) contamination and Cr(VI)-resistant extremophiles in diverse extreme environments. Environ. Sci. Pollut. Res. 2023, 30, 59163–59193. [Google Scholar] [CrossRef]
- Firincă, C.; Zamfir, L.G.; Constantin, M.; Răut, I.; Capră, L.; Popa, D.; Jinga, M.-L.; Baroi, A.M.; Fierăscu, R.C.; Corneli, N.O.; et al. Microbial removal of heavy metals from contaminated environments using metal-resistant indigenous strains. J. Xenobiot. 2023, 14, 51–78. [Google Scholar] [CrossRef]
- Alotaibi, B.S.; Khan, M.; Shamim, S. Unraveling the underlying heavy metal detoxification mechanisms of Bacillus species. Microorganisms 2021, 9, 1628. [Google Scholar] [CrossRef] [PubMed]
- Maamoun, I.; Rushdi, M.A.; Falyouna, O.; Eljamal, R.; Eljamal, O. Insights into machine-learning modeling for Cr(VI) removal from contaminated water using nano-nickel hydroxide. Sep. Purif. Technol. 2023, 308, 122863. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, Y.; He, B.; Li, J.; Yang, T.; Sun, H.; Shao, Q.; Xu, C. Deep learning algorithms in predicting Cr(VI) removal performance of S-ZVI: Models building and optimal parameters prediction. Sep. Purif. Technol. 2024, 330, 125487. [Google Scholar] [CrossRef]
- Huang, Y.; Tang, J.; Zhang, B.; Long, Z.E.; Ni, H.; Fu, X.; Zou, L. Influencing factors and mechanism of Cr(VI) reduction by facultative anaerobic Exiguobacterium sp. PY14. Front. Microbiol. 2023, 14, 1242410. [Google Scholar] [CrossRef] [PubMed]
- Gu, B.; Chen, J. Enhanced microbial reduction of Cr(VI) and U(VI) by different natural organic matter fractions. Geochim. Cosmochim. Acta 2003, 67, 3575–3582. [Google Scholar] [CrossRef]
- Chen, G.; Bai, Y.; Zeng, R.J.; Qin, L. Effects of different metabolic pathways and environmental parameters on Cr isotope fractionation during Cr(VI) reduction by extremely thermophilic bacteria. Geochim. Cosmochim. Acta 2019, 256, 135–146. [Google Scholar] [CrossRef]
- Padhiari, B.M.; Ray, A.; Champati, B.B.; Jena, S.; Sahoo, A.; Kuanar, A.; Halder, T.; Ghosh, B.; Naik, P.K.; Patnaik, J.; et al. Artificial neural network (ANN) model for prediction and optimization of bacoside A content in Bacopa monnieri: A statistical approach and experimental validation. Plant Biosyst.-Int. J. Deal. All Asp. Plant Biol. 2022, 156, 1346–1357. [Google Scholar] [CrossRef]
- Tariq, R.; Abatal, M.; Bassam, A. Computational intelligence for empirical modeling and optimization of methylene blue adsorption phenomena using available local zeolites and clay of Morocco. J. Clean. Prod. 2022, 370, 133517. [Google Scholar] [CrossRef]
- Mijwel, A.A.S.; Ahmed, A.N.; Afan, H.A.; Alayan, H.M.; Sherif, M.; Elshafie, A. Artificial intelligence models for methylene blue removal using functionalized carbon nanotubes. Sci. Rep. 2023, 13, 18260. [Google Scholar] [CrossRef]
- Marimuthu, S.; Rajendran, K. Artificial neural network modeling and statistical optimization of medium components to enhance production of exopolysaccharide by Bacillus sp. EPS003. Prep. Biochem. Biotechnol. 2023, 53, 136–147. [Google Scholar] [CrossRef] [PubMed]
- Yin, C.; Rosendahl, L.; Luo, Z. Methods to improve prediction performance of ANN models. Simul. Model. Pract. Theory 2003, 11, 211–222. [Google Scholar] [CrossRef]
- Garrity, G.M.; Holt, J.G. The road map to the manual. In Bergey’s Manual® of Systematic Bacteriology; Springer: New York, NY, USA, 2001; pp. 119–166. [Google Scholar]
- Poole, P.; Allaway, D. Carbon and nitrogen metabolism in Rhizobium. Adv. Microb. Physiol. 2000, 43, 117–163. [Google Scholar] [PubMed]
- Khodaverdiloo, H.; Samadi, A. Batch equilibrium study on sorption, desorption, and immobilisation of cadmium in some semi-arid zone soils as affected by soil properties. Soil Res. 2011, 49, 444–454. [Google Scholar] [CrossRef]
- Murphy, O.P.; Vashishtha, M.; Palanisamy, P.; Kumar, K.V. A review on the adsorption isotherms and design calculations for the optimization of adsorbent mass and contact time. ACS Omega 2023, 8, 17407–17430. [Google Scholar] [CrossRef]
- Musah, M.; Azeh, Y.; Mathew, J.T.; Umar, M.T.; Abdulhamid, Z.; Muhammad, A.I. Adsorption kinetics and isotherm models: A review. CaJoST 2022, 4, 20–26. [Google Scholar] [CrossRef]
- Ashraf, M.A.; Mahmood, K.; Wajid, A.; Maah, M.J.; Yusoff, I. Study of low cost biosorbent for biosorption of heavy metals. In Proceedings of the International Conference on Food Engineering and Biotechnology, Bangkok, Thailand, 7–9 May 2011; IPCBEE: Singapore, 2011; Volume 9, pp. 60–68. [Google Scholar]
- Maind, S.D.; Sharma, A.S.; Poojari, A.C.; Jadav, J.N.; Bhalerao, S.A. Utility of Plant Based Waste Materials as Potential Biosorbent for Sequestering Chromium (VI) from Aqueous Solutions. Int. J. Extensive Res. 2015, 9, 12–15. [Google Scholar]
- Oves, M.; Khan, M.S.; Qari, H.A. Ensifer adhaerens for heavy metal bioaccumulation, biosorption, and phosphate solubilization under metal stress condition. J. Taiwan Inst. Chem. Eng. 2017, 80, 540–552. [Google Scholar] [CrossRef]
- Liu, T.; Wang, Y.; Feng, L.; Tian, X.; Cui, J.; Yu, Z.; Wang, C.; Zhang, B.; James, T.D.; Ma, X. 2D strategy for the construction of an enzyme-activated NIR fluorophore suitable for the visual sensing and profiling of homologous nitroreductases from various bacterial species. ACS Sens. 2021, 6, 3348–3356. [Google Scholar] [CrossRef] [PubMed]
- Sanjay, M.S.; Sudarsanam, D.; Raj, G.A.; Baskar, K. Isolation and identification of chromium reducing bacteria from tannery effluent. J. King Saud Univ.-Sci. 2020, 32, 265–271. [Google Scholar] [CrossRef]
- Witek-Krowiak, A.; Chojnacka, K.; Podstawczyk, D.; Dawiec, A.; Bubała, K. Application of response surface methodology and artificial neural network methods in modelling and optimization of biosorption process. Bioresour. Technol. 2014, 160, 150–160. [Google Scholar] [CrossRef]
- Massaoudi, A.; Echouchene, F.; Ben Ayed, M.; Berguiga, A.; Harchay, A.; Al-Ghamdi, S.; Belmabrouk, H. Machine learning models for modeling the biosorption of Fe(III) ions by activated carbon from olive stone. Neural Comput. Appl. 2024, 36, 13357–13372. [Google Scholar] [CrossRef]
- Kardam, A. Simulation and optimization of artificial neural network modeling for prediction of sorption efficiency of nanocellulose fibers for removal of Cd(II) ions from aqueous system. Walailak J. Sci. Technol. 2013, 11, 497–508. [Google Scholar]
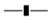 ) and the removal of both (
) and the removal of both ( ) heavy metals and Cr(VI) at variable pH; (c) the cellular growth (
) heavy metals and Cr(VI) at variable pH; (c) the cellular growth ( ) and removal of (
) and removal of ( ) heavy metals by Rhizobium sp. OS1 grown in NB medium with 100 mg/L at different temperatures for 24 h at pH 7.
) and the removal of both () heavy metals and Cr(VI) at variable pH; (c) the cellular growth () and removal of () heavy metals by Rhizobium sp. OS1 grown in NB medium with 100 mg/L at different temperatures for 24 h at pH 7.
) heavy metals by Rhizobium sp. OS1 grown in NB medium with 100 mg/L at different temperatures for 24 h at pH 7.
) and the removal of both () heavy metals and Cr(VI) at variable pH; (c) the cellular growth () and removal of () heavy metals by Rhizobium sp. OS1 grown in NB medium with 100 mg/L at different temperatures for 24 h at pH 7.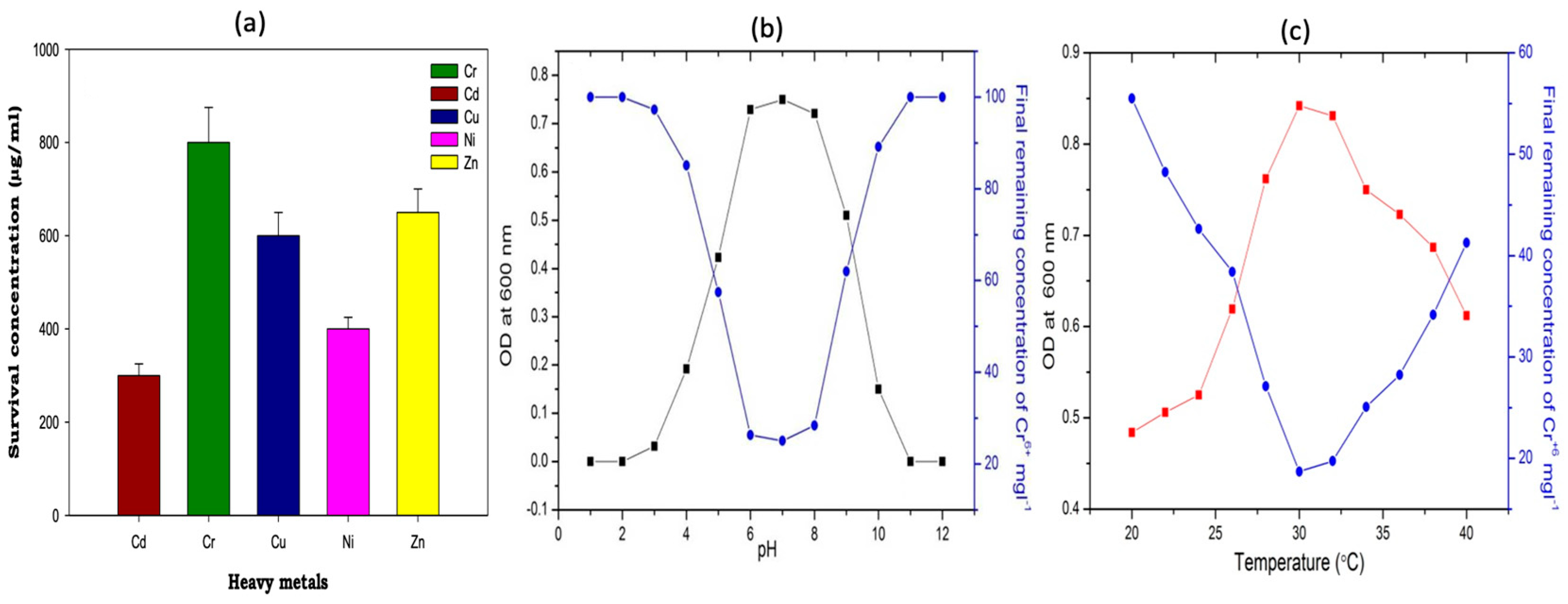

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Rhizobia Groups |
|---|---|
| Gram reaction | G-ve |
| Cell size and shape | Short rods |
| Colony morphology | Transparent, circular, and mucoid |
| Pigment color | pink |
| Nitrate | + |
| Methyl® | − |
| Catalase | + |
| Citrate | + |
| Voges Proskauer | − |
| Indole | − |
| Gelatin | − |
| Starch | + |
| Glucose | + |
| Sucrose | + |
| Mannitol | + |
| Presumptive identification | Rhizobium sp. |
| Final identification | HE663761.1 |
| (A) | ||||||||
|---|---|---|---|---|---|---|---|---|
| Metal Concentration (mg/L) | Percent Removal | |||||||
| Cr(VI) | Cu(II) | Cd(II) | Ni(II) | Zn(II) | Pb(II) | |||
| 25 | 96.70 | 96.10 | 94.90 | 95.50 | 94.80 | 90.80 | ||
| 50 | 95.68 | 95.76 | 93.76 | 94.36 | 94.16 | 90.16 | ||
| 75 | 93.96 | 94.36 | 92.33 | 92.96 | 93.76 | 88.16 | ||
| 100 | 92.35 | 91.84 | 90.54 | 91.84 | 91.44 | 87.44 | ||
| 125 | 91.64 | 89.41 | 88.15 | 89.99 | 91.08 | 86.6 | ||
| 150 | 89.58 | 87.52 | 85.37 | 90.05 | 88.78 | 84.72 | ||
| (B) | ||||||||
| Metal | Adsorption Isotherm | |||||||
| Langmuir | Freundlich | |||||||
| Qmax | b | r2 | Sf | Ø | k | 1/n | r2 | |
| Cr(VI) | 37.17 | 0.234 | 0.995 | 0.041 | 0.959 | 7.166 | 0.580 | 0.994 |
| Cu(II) | 41.66 | 0.174 | 0.996 | 0.054 | 0.945 | 7.059 | 0.549 | 0.960 |
| Cd(II) | 41.15 | 0.131 | 0.999 | 0.071 | 0.929 | 5.634 | 0.593 | 0.983 |
| Ni(II) | 43.10 | 0.142 | 0.996 | 0.065 | 0.934 | 5.757 | 0.651 | 0.996 |
| Zn(II) | 55.86 | 0.090 | 0.998 | 0.099 | 0.901 | 5.445 | 0.674 | 0.981 |
| Pb(II) | 62.89 | 0.043 | 0.998 | 0.122 | 0.977 | 3.187 | 0.752 | 0.994 |
| Metric | Training Dataset | Testing Dataset |
|---|---|---|
| MAPE (%) | Lowest (at 10 hidden neurons) | Lowest (at 10 hidden neurons) |
| RMSE | Lowest (at 10 hidden neurons) | Lowest (at 10 hidden neurons) |
| MSE | Lowest (at 10 hidden neurons) | Lowest (at 10 hidden neurons) |
| R2 | High (close to 1, at optimal neurons) | High (close to 1, at optimal neurons) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oves, M.; Al-Shaeri, M.A.; Qari, H.A.; Khan, M.S. Rhizobium’s Reductase for Chromium Detoxification, Heavy Metal Resistance, and Artificial Neural Network-Based Predictive Modeling. Catalysts 2025, 15, 726. https://doi.org/10.3390/catal15080726
Oves M, Al-Shaeri MA, Qari HA, Khan MS. Rhizobium’s Reductase for Chromium Detoxification, Heavy Metal Resistance, and Artificial Neural Network-Based Predictive Modeling. Catalysts. 2025; 15(8):726. https://doi.org/10.3390/catal15080726
Chicago/Turabian StyleOves, Mohammad, Majed Ahmed Al-Shaeri, Huda A. Qari, and Mohd Shahnawaz Khan. 2025. "Rhizobium’s Reductase for Chromium Detoxification, Heavy Metal Resistance, and Artificial Neural Network-Based Predictive Modeling" Catalysts 15, no. 8: 726. https://doi.org/10.3390/catal15080726
APA StyleOves, M., Al-Shaeri, M. A., Qari, H. A., & Khan, M. S. (2025). Rhizobium’s Reductase for Chromium Detoxification, Heavy Metal Resistance, and Artificial Neural Network-Based Predictive Modeling. Catalysts, 15(8), 726. https://doi.org/10.3390/catal15080726