
Screening of Cellulolytic Bacteria from Various Ecosystems and Their Cellulases Production under Multi-Stress Conditions

 , ,
, ,  , ,
, , 
Abstract
1. Introduction
2. Results
2.1. Screening for Cellulase-Producing Strains
2.2. Evaluation of the Enzymatic Activity
2.3. Effect of Physicochemical Parameters on Bacterial Growth and Cellulase Production
2.3.1. Effect of pH on Bacterial Growth
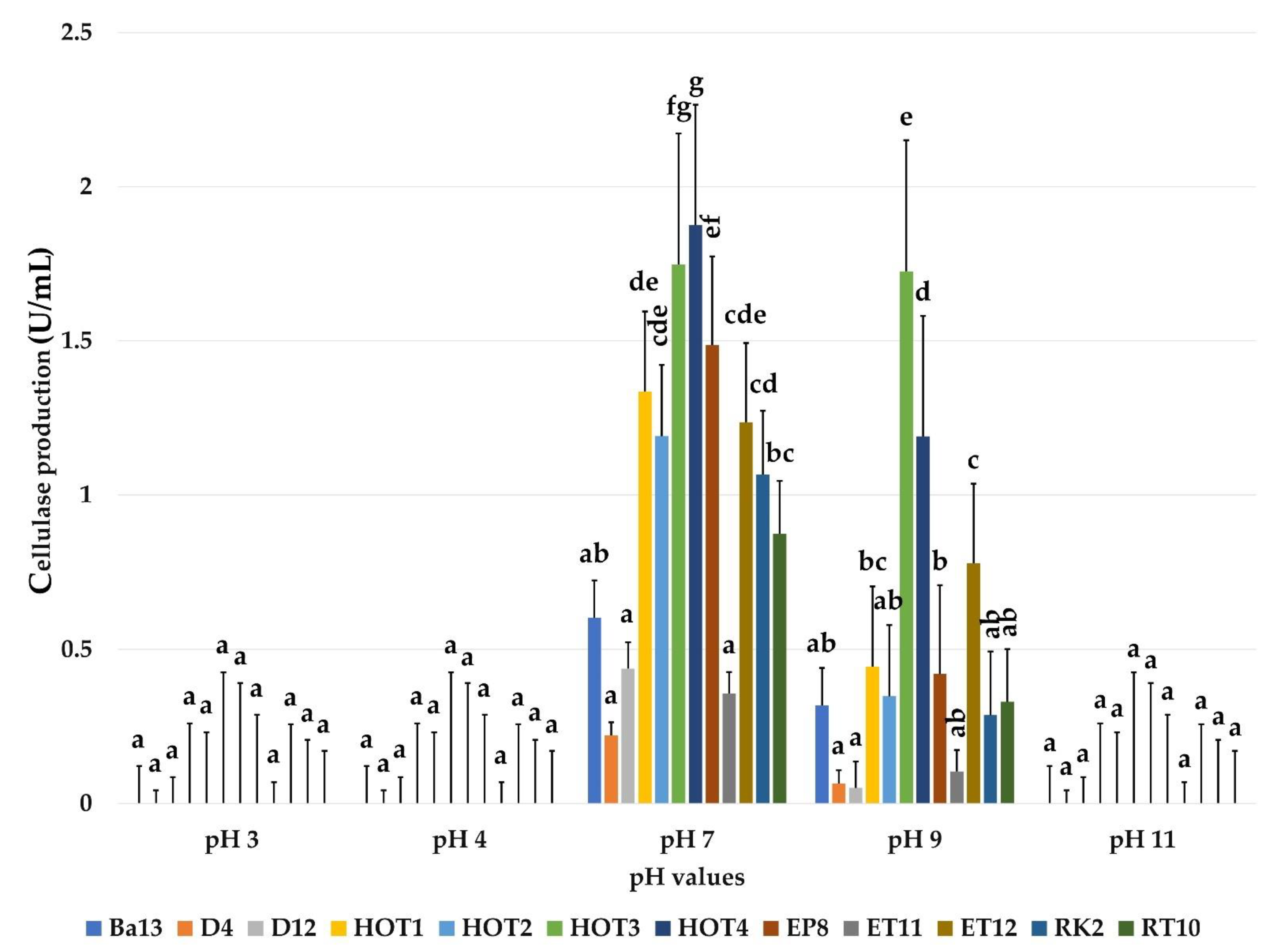
2.3.2. Effect of pH on Cellulase Production
2.3.3. Effect of Temperature on Bacterial Growth
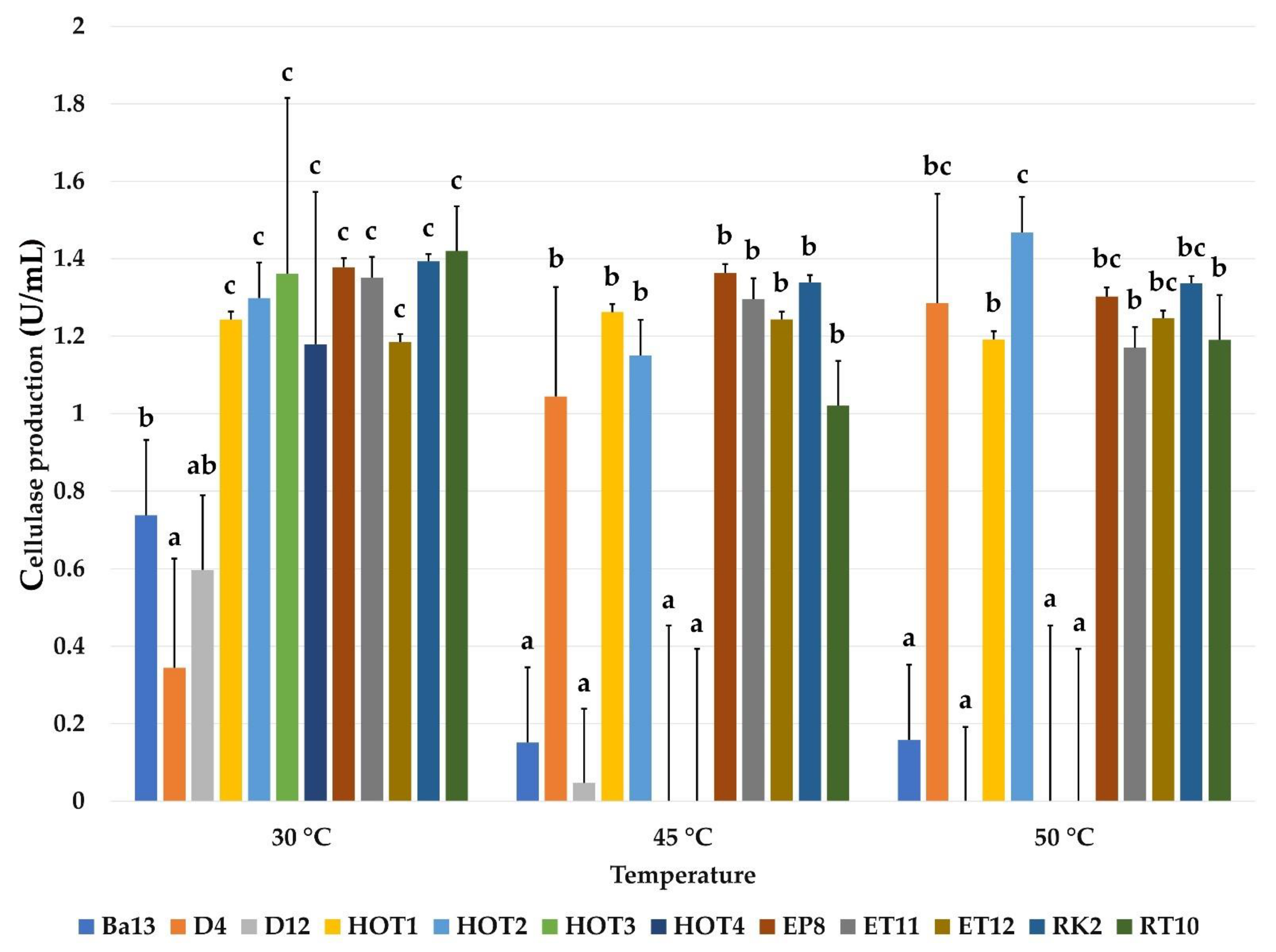
2.3.4. Effect of Temperature on Cellulase Production
2.3.5. Effect of Salinity on Bacterial Growth
2.3.6. Effect of Salinity on Cellulase Production
2.3.7. Effect of Water Activity (aw) on Bacterial Growth
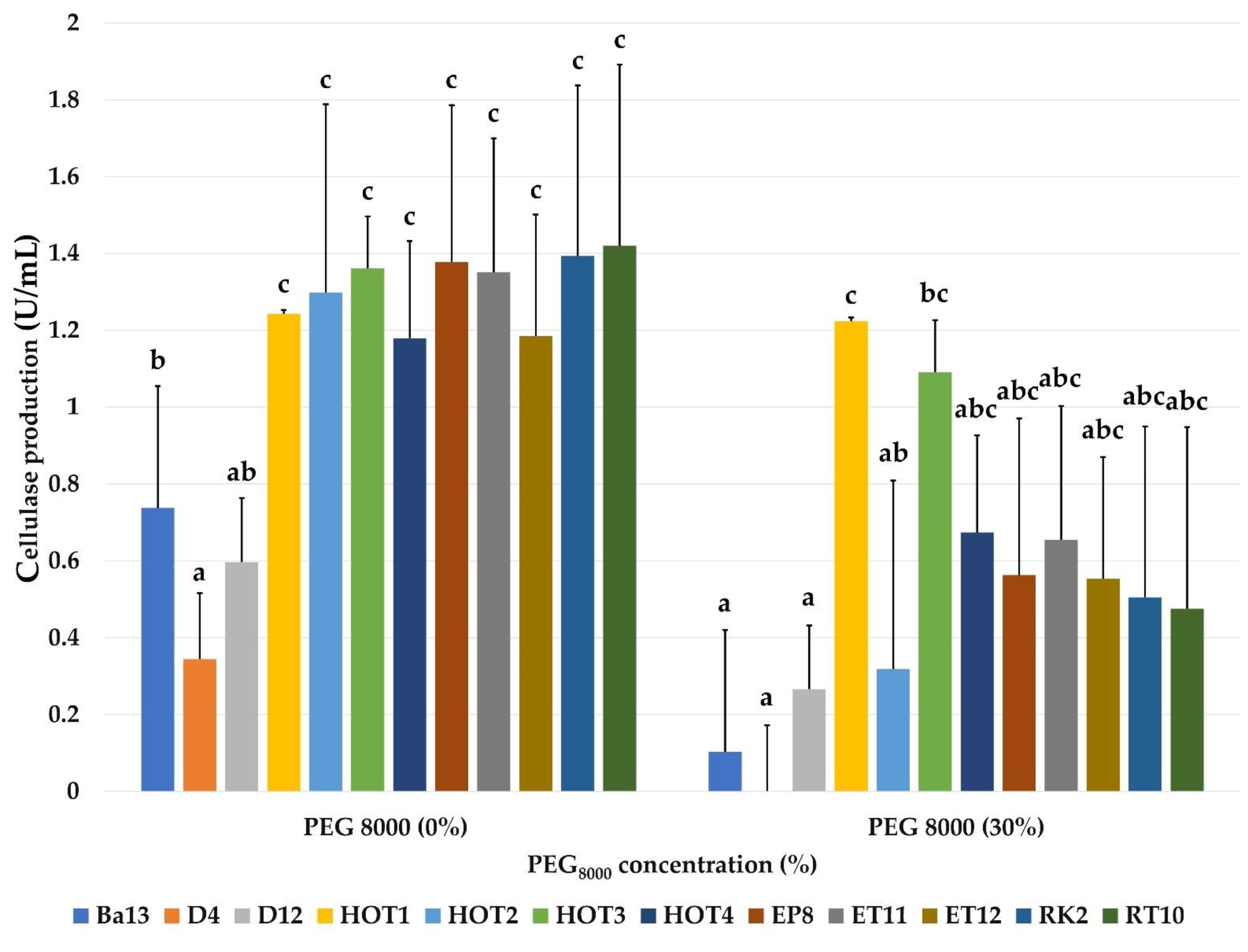
2.3.8. Effect of Water Activity on Cellulase Production
2.4. Molecular Identification of Bacterial Strains and Their Phylogenetic Positions
2.5. Effect of Physico-Chemical Parameters on Cellulase Activity and Stability
2.5.1. Effect of pH on Cellulase Activity and Stability
2.5.2. Effect of Temperature on Cellulase Activity and Stability
2.5.3. Effect of Salinity on Cellulase Activity and Stability
2.5.4. Effect of Water Activity (aw) on Cellulase Activity and Stability
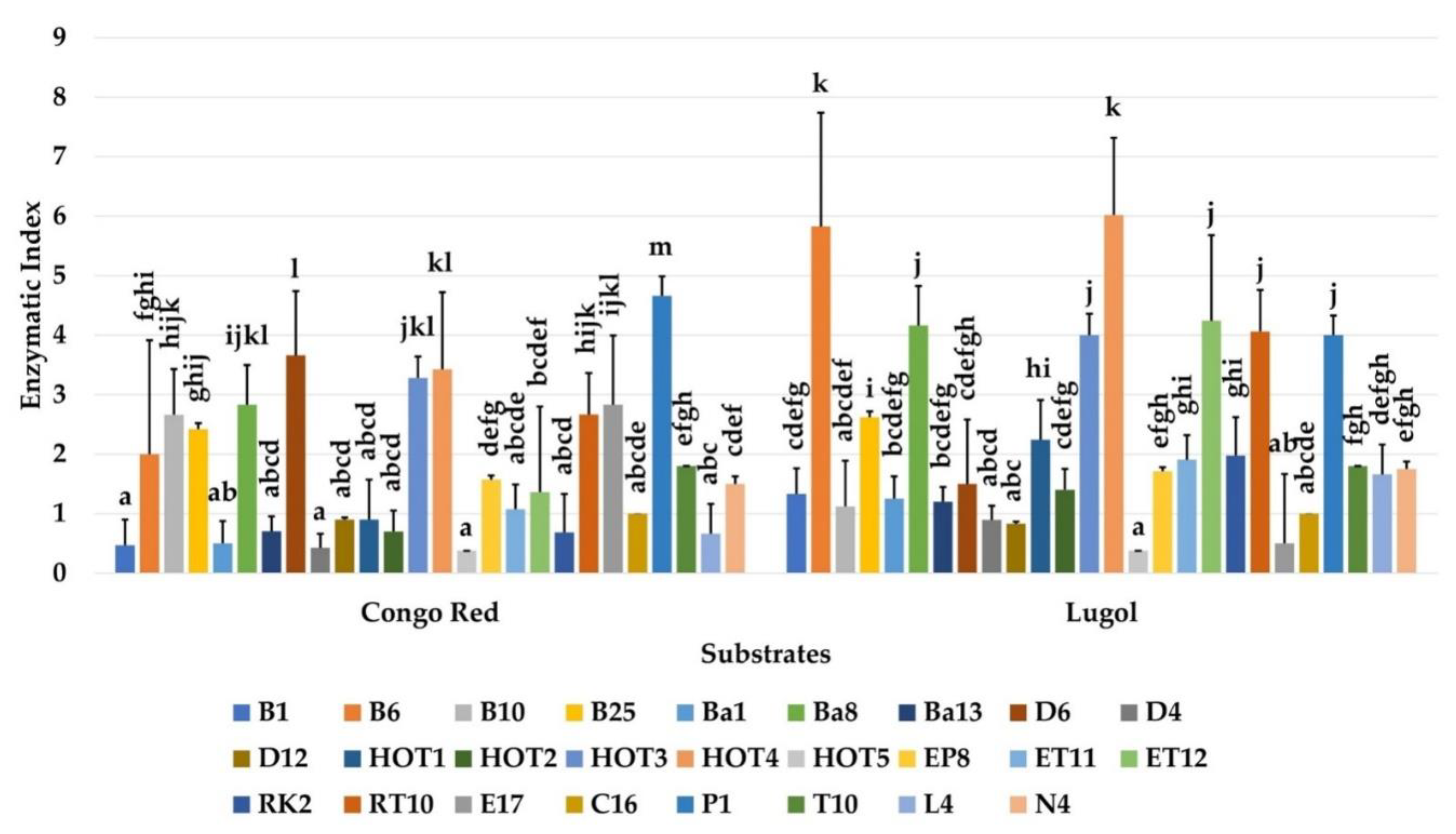
2.6. Hydrolysis of Different Substrates
2.7. Estimation of Total Sugars
2.8. Estimation of Total Proteins
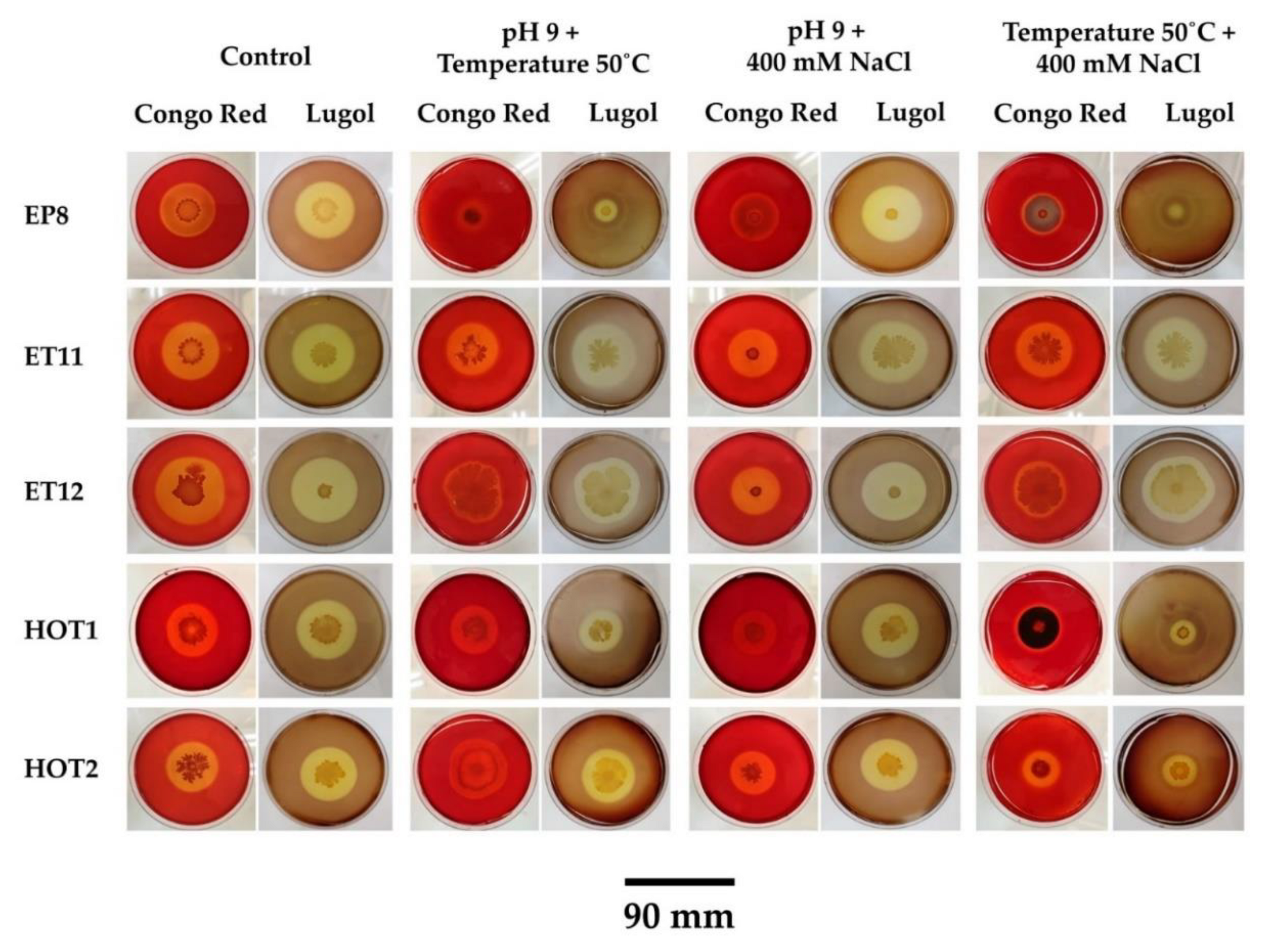
2.9. Effect of Multiple Stress on the Growth and Production of Cellulase on Solid Medium
2.10. Effect of Multiple Stress on the Growth and Production of Cellulase on Liquid Medium
2.11. Effect of Various Cellulosic Substrates on Cellulase Production
3. Discussion
4. Materials and Methods
4.1. Biological Material
4.2. Screening for Cellulase-Producing Isolates
4.3. Cellulase Production on Liquid Medium
4.4. Evaluation of Cellulase Activity
4.5. Effect of Physico-Chemical Parameters on Bacterial Growth and Cellulase Production Capacity
4.5.1. pH
4.5.2. Temperature
4.5.3. Salinity
4.5.4. Water activity (aw)
5. Effect of Physico-Chemical Parameters on Cellulase Activity and Stability
5.1. pH
5.1.1. Temperature
5.1.2. Salinity
5.1.3. Water Activity (aw)
5.2. Molecular Identification of the Selected Isolates
5.3. Hydrolysis of Different Substrates
5.4. Estimation of Total Sugars
5.5. Estimated of Total Proteins
5.6. Effect of Multiple Stress on Growth and Cellulase Production on Solid Medium
5.7. Effect of Multiple Stress on Growth and Cellulase Production on Liquid Medium
5.8. Effect of Various Cellulosic Substrates on the Production of Cellulases
5.9. Statistical Analysis
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Sample Availability
References
- Periyasamy, S.; Karthik, V.; Senthil Kumar, P.S.; Isabel, J.B.; Temesgen, T.; Hunegnaw, B.M.; Melese, B.B.; Mohamed, B.A.; Vo, D.-V.N. Chemical, physical and biological methods to convert lignocellulosic waste into value-added products. A Review. En-Viron. Chem. Letters 2022, 20, 1129–1152. [Google Scholar] [CrossRef]
- Ja’afar, J.A.N.; Shitu, A. Utilization of Lignocellulosic Agro-Waste as an Alternative Carbon Source for Industrial Enzyme Production. In Waste Management, Processing and Valorisation; Embrandiri, A., Kassaw, G.M., Geto, A.K., Wogayehu, B.T., Embrandiri, M., Eds.; Springer: Singapore, 2022; pp. 221–233. [Google Scholar]
- Himanshu; Saini, J.K. Diversity and Biotechnological Importance of Cellulolytic Microorganisms from Biodiversity Hotspots; Academic Press: Pune, India, 2021; pp. 207–230. [Google Scholar] [CrossRef]
- Sahay, S. Deconstruction of lignocelluloses: Potential biological approaches. In Handbook of Biofuels; Academic Press: Cambridge, MA, USA, 2021; pp. 207–232. [Google Scholar] [CrossRef]
- Fatima, A.; Yasir, S.; Khan, M.S.; Manan, S.; Ullah, M.W.; Ul-Islam, M. Plant extract-loaded bacterial cellulose composite membrane for potential biomedical applications. J. Bioresour. Bioprod. 2021, 6, 26–32. [Google Scholar] [CrossRef]
- Xia, Z.; Li, J.; Zhang, J.; Zhang, X.; Zheng, X.; Zhang, J. Processing and valorization of cellulose, lignin and lignocellulose using ionic liquids. J. Bioresour. Bioprod. 2020, 5, 79–95. [Google Scholar] [CrossRef]
- Jayasekara, S.; Ratnayake, R. Microbial cellulases: An overview and applications. In Cellulose; Pascual, A.R., Martín, M.E.E., Eds.; IntechOpen: London, UK, 2019; pp. 1–21. [Google Scholar] [CrossRef]
- Singh, A.; Bajar, S.; Devi, A.; Pant, D. An overview on the recent developments in fungal cellulase production and their industrial applications. Bioresour. Technol. Rep. 2021, 14, 100652. [Google Scholar] [CrossRef]
- Zhang, Y.H.P.; Himmel, M.E.; Mielenz, J.R. Outlook for cellulase improvement: Screening and selection strategies. Biotechnol. Adv. 2006, 24, 452–481. [Google Scholar] [CrossRef] [PubMed]
- Lavanya, D.K.P.K.; Kulkarni, P.K.; Dixit, M.; Raavi, P.K.; Krishna, L.N.V. Sources of cellulose and their applications—A review. Int. J. Drug Dev. Res. 2011, 2, 19–38. [Google Scholar]
- Singh, S.; Jaiswal, D.K.; Sivakumar, N.; Verma, J.P. Developing efficient thermophilic cellulose degrading consortium for glucose production from different agro-residues. Front. Energy Res. 2019, 7, 61. [Google Scholar] [CrossRef]
- Nagendran, S.; Hallen-Adams, H.E.; Paper, J.M.; Aslam, N.; Walton, J.D. Reduced genomic potential for secreted plant cell-wall-degrading enzymes in the ectomycorrhizal fungus Amanita bisporigera, based on the secretome of Trichoderma reesei. Fungal Geneti. Biol. 2009, 46, 427–435. [Google Scholar] [CrossRef]
- Saratale, G.D.; Saratale, R.G.; Oh, S.E. Production and characterization of multiple cellulolytic enzymes by isolated Streptomyces sp. MDS. Biomass Bioenergy 2012, 47, 302–315. [Google Scholar] [CrossRef]
- Sadhu, S.; Maiti, T.K. Cellulase production by bacteria: A review. Br. Microbiol. Res. J. 2013, 3, 235–258. [Google Scholar] [CrossRef]
- Malik, W.A.; Javed, S. Biochemical Characterization of Cellulase from Bacillus subtilis Strain and its Effect on Digestibility and Structural Modifications of Lignocellulose Rich Biomass. Front. Bioeng. Biotechnol. 2021, 9, 800265. [Google Scholar] [CrossRef]
- Chen, G.Q.; Jiang, X.R. Next generation industrial biotechnology based on extremophilic bacteria. Curr. Opin. Biotechnol. 2018, 50, 94–100. [Google Scholar] [CrossRef]
- Sharma, A.; Tewari, R.; Rana, S.S.; Soni, R.; Soni, S.K. Cellulases: Classification, methods of determination and industrial applications. Appl. Biochem. Biotechnol. 2016, 179, 1346–1380. [Google Scholar] [CrossRef]
- Gaur, R.; Tiwari, S. Isolation, production, purification and characterization of an organic-solvent-thermostable alkalophilic cellulase from Bacillus vallismortis RG-07. BMC Biotechnol. 2015, 15, 1–12. [Google Scholar] [CrossRef]
- Nwachukwu, B.C.; Ayangbenro, A.S.; Babalola, O.O. Elucidating the rhizosphere associated bacteria for environmental sustainability. Agriculture 2021, 11, 75. [Google Scholar] [CrossRef]
- Wilson, C.K.; King, G.M. Short-Term Exposure to Thermophilic Temperatures Facilitates CO Uptake by Thermophiles Maintained under Predominantly Mesophilic Conditions. Microorganisms 2022, 10, 656. [Google Scholar] [CrossRef]
- Khelil, O.; Cheba, B. Thermophilic cellulolytic microorganisms from western Algerian sources: Promising isolates for cellulosic biomass recycling. Proc. Technol. 2014, 12, 519–528. [Google Scholar] [CrossRef]
- McNamara, C.J.; Perry, T.D.; Bearce, K.A.; Hernandez-Duque, G.; Mitchell, R. Epilithic and endolithic bacterial communities in limestone from a Maya archaeological site. Microb. Ecol. 2006, 51, 51–64. [Google Scholar] [CrossRef]
- Antonelli, F.; Esposito, A.; Galotta, G.; Davidde Petriaggi, B.; Piazza, S.; Romagnoli, M.; Guerrieri, F. Microbiota in waterlogged archaeological wood: Use of Next-Generation Sequencing to evaluate the risk of biodegradation. Appl. Sci. 2020, 10, 4636. [Google Scholar] [CrossRef]
- Houfani, A.A.; Větrovský, T.; Navarrete, O.U.; Štursová, M.; Tláskal, V.; Beiko, R.G.; Boucherba, N.; Baldrian, P.; Benallaoua, S.; Jorquera, M.A. Cellulase−hemicellulase activities and bacterial community composition of different soils from Algerian ecosystems. Microb. Ecol. 2019, 77, 713–725. [Google Scholar] [CrossRef]
- Kasana, R.C.; Salwan, R.; Dhar, H.; Dutt, S.; Gulati, A. A rapid and easy method for the detection of microbial cellulases on agar plates using Gram’s iodine. Curr. Microbiol. 2008, 57, 503–507. [Google Scholar] [CrossRef] [PubMed]
- Meddeb-Mouelhi, F.; Moisan, J.K.; Beauregard, M. A comparison of plate assay methods for detecting extracellular cellulase and xylanase activity. Enzym. Microb. Technol. 2014, 66, 16–19. [Google Scholar] [CrossRef] [PubMed]
- Maryam, B.; Qadir, A.; Zameer, M.; Ahmad, S.R.; Nelofer, R.; Jamil, N.; Arzoo, S.; Afzaal, R. Production of Cellulases by Bacillus cellulosilyticus Using Lignocellulosic Material. Pol. J. Environ. Stud. 2018, 27, 2659–2667. [Google Scholar] [CrossRef]
- Deka, D.; Bhargavi, P.; Sharma, A.; Goyal, D.; Jawed, M.; Goyal, A. Enhancement of cellulase activity from a new strain of Bacillus subtilis by medium optimization and analysis with various cellulosic substrates. Enzym. Res. 2011, 2011, 151656. [Google Scholar] [CrossRef]
- Mawadza, C.; Hatti-Kaul, R.; Zvauya, R.; Mattiasson, B. Purification and characterization of cellulases produced by two Bacillus strains. J. Biotechnol. 2000, 83, 177–187. [Google Scholar] [CrossRef]
- Nnaemeka, I.C.; Maxwell, O.; Christain, A.O. Optimization and kinetic studies for enzymatic hydrolysis and fermentation of colocynthis vulgaris Shrad seeds shell for bioethanol production. J. Bioresour. Bioprod. 2021, 6, 45–64. [Google Scholar] [CrossRef]
- Sarsan, S.; Merugu, R. Role of bioprocess parameters to improve cellulase production: Part II. In New and Future Developments in Microbial Biotechnology and Bioengineering. From Cellulose to Cellulase: Strategies to Improve Biofuel Production; Srivastava, N., Srivastava, M., Mishra, P.K., Ramteke, P.W., Singh, R.L., Eds.; Elsevier: Pune, India, 2019; pp. 77–97. [Google Scholar]
- Bakare, M.K.; Adewale, I.O.; Ajayi, A.; Shonukan, O.O. Purification and characterization of cellulase from the wild-type and two improved mutants of Pseudomonas fluorescens. Afr. J. Biotechnol. 2005, 4, 898–904. [Google Scholar]
- Shankar, T.; Isaiarasu, L. Cellulase production by Bacillus pumilus EWBCM1 under varying cultural conditions. Middle East J. Sci. Res. 2011, 8, 40–45. [Google Scholar]
- Raju, E.V.N.; Divakar, G. Production of pectinase by using Bacillus circulans isolated from dump yards of vegetable wastes. Int. J. Pharmaceut. Sci. Res. 2013, 4, 2615–2622. [Google Scholar]
- Immanuel, G.; Dhanusha, R.; Prema, P.; Palavesam, A.J.I.J. Effect of different growth parameters on endoglucanase enzyme activity by bacteria isolated from coir retting effluents of estuarine environment. Int. J. Environ. Sci. Technol. 2006, 3, 25–34. [Google Scholar] [CrossRef]
- Annamalai, N.; Thavasi, R.; Vijayalakshmi, S.; Balasubramanian, T. A novel thermostable and halostable carboxymethylcellulase from marine bacterium Bacillus licheniformis AU01. World J. Microbiol. Biotechnol. 2011, 27, 2111–2115. [Google Scholar] [CrossRef]
- Kushner, D.J. Life in high salt and solute concentrations. In Microbial Life in Extreme Environments; Kushner, D.J., Ed.; Academic Press: London, UK, 1978; pp. 317–368. [Google Scholar]
- Vargas, V.A.; Delgado, O.D.; Hatti-Kaul, R.; Mattiasson, B. Bacillus bogoriensis sp. nov., a novel alkaliphilic, halotolerant bacterium isolated from a Kenyan soda lake. Int. J. Sys. Evol. Microbiol. 2005, 55, 899–902. [Google Scholar] [CrossRef][Green Version]
- Yu, H.Y.; Li, X. Alkali-stable cellulase from a halophilic isolate, Gracilibacillus sp. SK1 and its application in lignocellulosic saccharification for ethanol production. Biomass. Bioenergy. 2015, 81, 19–25. [Google Scholar] [CrossRef]
- Gontia-Mishra, I.; Sapre, S.; Sharma, A.; Tiwari, S. Amelioration of drought tolerance in wheat by the interaction of plant growth-promoting rhizobacteria. Plant Biol. 2016, 18, 992–1000. [Google Scholar] [CrossRef]
- Vardharajula, S.; Zulfikar Ali, S.; Grover, M.; Reddy, G.; Bandi, V. Drought-tolerant plant growth promoting Bacillus spp.: Effect on growth, osmolytes, and antioxidant status of maize under drought stress. J. Plant Interact. 2011, 6, 1–14. [Google Scholar] [CrossRef]
- Lee, Y.J.; Kim, B.K.; Lee, B.H.; Jo, K.I.; Lee, N.K.; Chung, C.H.; Lee, Y.C.; Lee, J.W. Purification and characterization of cellulase produced by Bacillus amyoliquefaciens DL-3 utilizing rice hull. Bioresour. Technol. 2008, 99, 378–386. [Google Scholar] [CrossRef]
- Dos Santos, Y.Q.; De Veras, B.O.; De Franca, A.F.J.; Gorlach-Lira, K.; Velasques, J.; Migliolo, L.; Dos Santos, E.A. A new salt-tolerant thermostable cellulase from a marine Bacillus sp. strain. J. Microbiol. Biotechnol. 2018, 28, 1078–1085. [Google Scholar] [CrossRef]
- Samiullah, T.R.; Bakhsh, A.; Rao, A.Q.; Naz, M.; Saleem, M. Isolation, purification and characterization of extracellular β-glucosidase from Bacillus sp. Adv. Environ. Biol. 2009, 3, 269–277. [Google Scholar]
- Sheble Ibrahim, A.S.; El-diwany, A. Isolation and identification of new cellulases thermophilic bacteria from an Egyptian hot spring and some properties of the crude enzyme. Aust. J. Basic Appl. Sci. 2007, 1, 473–478. [Google Scholar]
- Wang, C.Y.; Hsieh, Y.R.; Ng, C.C.; Chan, H.; Lin, H.T.; Tzeng, W.S.; Shyu, Y.T. Purification and characterization of a novel halostable cellulase from Salinivibrio sp. strain NTU-05. Enzyme Microb. Technol. 2009, 44, 373–379. [Google Scholar] [CrossRef]
- Trivedi, N.; Gupta, V.; Kumar, M.; Kumari, P.; Reddy, C.R.K.; Jha, B. An alkali-halotolerant cellulase from Bacillus flexus isolated from green seaweed Ulva lactuca. Carbohydr. Polym. 2011, 83, 891–897. [Google Scholar] [CrossRef]
- Gilbert, H.J.; Hazlewood, G.P. Bacterial cellulases and xylanases. Microbiology 1993, 139, 187–194. [Google Scholar] [CrossRef]
- Li, X.; Lin, W.; Gao, P.; Chen, F. Endoglucanase S, a novel endocellulase exhibiting exoglucanase activity from a newly isolated Streptomyces sp. LX. J. Appl. Microbiol. 1998, 85, 347–356. [Google Scholar] [CrossRef]
- Coughlan, M.P.; Mayer, F. The cellulase decomposing bacteria and their enzyme systems. In The Prokaryotes, 2nd ed.; Balowes, A., Trurer, H., Dworkin, M., Harder, W., Schleifer, K.H., Eds.; Springer: New York, NY, USA, 1992; Volume 1, pp. 460–516. [Google Scholar]
- Hefford, M.A.; Laderoute, K.; Willick, G.E.; Yaguchi, M.; Seligy, V.L. Bipartite organization of the Bacillus subtilis endo-β-1, 4-glucanase revealed by C-terminal mutations. Protein Eng. Des. Sel. 1992, 5, 433–439. [Google Scholar] [CrossRef] [PubMed]
- Wood, T.M.; McCRAE, S.I. The cellulase of Trichoderma koningii, Purification and properties of some endoglucanase components with special reference to their action on cellulose when acting alone and in synergism with the cellobiohydrolase. Biochem. J. 1978, 171, 61–72. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.M.; Revol, J.F.; Gray, D.G. Effect of microcrystallite preparation conditions on the formation of colloid crystals of cellulose. Cellulose 1998, 5, 19–32. [Google Scholar] [CrossRef]
- Roussos, S.; Raimbault, M. Hydrolysis of cellulose by fungi. II. Production of cellulases by Trichoderma harzianium by fermentation in liquid media. Ann. Microbiol. 1982, 133, 465–474. [Google Scholar]
- Palma, C.; Martınez, A.T.; Lema, J.M.; Martınez, M.J. Different fungal manganese-oxidizing peroxidases: A comparison between Bjerkandera sp. and Phanerochaete chrysosporium. J. Biotechnol. 2000, 77, 235–245. [Google Scholar] [CrossRef]
- Ladeira, S.A.; Cruz, E.; Delatorre, A.B.; Barbosa, J.B.; Leal Martins, M.L. Cellulase production by thermophilic Bacillus sp: SMIA-2 and its detergent compatibility. Electron. J. Biotechnol. 2015, 18, 110–115. [Google Scholar] [CrossRef]
- Fastnaught, C.E. Barley fiber. In Handbook of Dietary Fiber; Cho, S.S., Dreher, M.L., Eds.; CRC Press: Boca Raton, FL, USA, 2001; pp. 519–542. [Google Scholar]
- Sullivan, P.; O’Flaherty, J.; Brunton, N.; Gee, V.L.; Arendt, E.; Gallagher, E. Chemical composition and microstructure of milled barley fractions. Eur. Food Res. Technol. 2010, 230, 579–595. [Google Scholar] [CrossRef]
- Mihajlovski, K.; Buntić, A.; Milić, M.; Rajilić-Stojanović, M.; Dimitrijević-Branković, S. From agricultural waste to biofuel: Enzymatic potential of a bacterial isolate Streptomyces fulvissimus CKS7 for bioethanol production. Waste Biomass Valorization 2021, 12, 165–174. [Google Scholar] [CrossRef]
- Vu, V.; Farkas, C.; Riyad, O.; Bujna, E.; Kilin, A.; Sipiczki, G.; Sharma, M.; Usmani, Z.; Gupta, V.K.; Nguyen, Q.D. Enhancement of the enzymatic hydrolysis efficiency of wheat bran using the Bacillus strains and their consortium. Bioresour. Technol. 2022, 343, 126092. [Google Scholar] [CrossRef] [PubMed]
- Kabel, M.A.; Bos, G.; Zeevalking, J.; Voragen, A.G.; Schols, H.A. Effect of pretreatment severity on xylan solubility and enzymatic breakdown of the remaining cellulose from wheat straw. Bioresour. Technol. 2007, 98, 2034–2042. [Google Scholar] [CrossRef] [PubMed]
- Pointner, M.; Kuttner, P.; Obrlik, T.; Jager, A.; Kahr, H. Composition of corncobs as a substrate for fermentation of biofuels. Agron. Res. 2014, 12, 391–396. [Google Scholar]
- Rawat, R.; Tewari, L. Purification and characterization of an acidothermophilic cellulase enzyme produced by Bacillus subtilis strain LFS3. Extremophiles 2012, 16, 637–644. [Google Scholar] [CrossRef]
- Ferbiyanto, A.; Rusmana, I.; Raffiudin, R. Characterization and identification of cellulolytic bacteria from gut of worker Macrotermes gilvus. Hayati J. Biosci. 2015, 22, 197–200. [Google Scholar] [CrossRef]
- Miller, G.L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Hussain, A.A.; Abdel-Salam, M.S.; Abo-Ghalia, H.H.; Hegazy, W.K.; Hafez, S.S. Optimization and molecular identification of novel cellulose degrading bacteria isolated from Egyptian environment. J. Genet. Eng. Biotechnol. 2017, 15, 77–85. [Google Scholar] [CrossRef]
- Kerbab, S.; Silini, A.; Chenari Bouket, A.; Cherif-Silini, H.; Eshelli, M.; El Houda Rabhi, N.; Belbahri, L. Mitigation of NaCl stress in wheat by rhizosphere engineering using salt habitat adapted PGPR halotolerant bacteria. Appl. Sci. 2021, 11, 1034. [Google Scholar] [CrossRef]
- Mefteh, F.B.; Chenari Bouket, A.; Daoud, A.; Luptakova, L.; Alenezi, F.N.; Gharsallah, N.; Belbahri, L. Metagenomic insights and genomic analysis of phosphogypsum and its associated plant endophytic microbiomes reveals valuable actors for waste bioremediation. Microorganisms 2019, 7, 382. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.T.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Lowry, O.C.; Rosebrough, N.; Farr, A.; Randall, R. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Islam, F.; Roy, N. Screening, purification and characterization of cellulase from cellulase producing bacteria in molasses. BMC Res. Notes 2018, 11, 1–6. [Google Scholar] [CrossRef]



















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial Isolates | Sampling Sites | Number of Active Strains (%) |
|---|---|---|
| P1, T10, L4, N4 | Archaeological site (rocks and Roman ruins) of Djemila-Setif | 4/70 (5.71) |
| EP8, ET11, ET12, RK2, RT10 | Rhizosphere of Terfez (Boussaada region) | 5/80 (6.25) |
| B1,B6, B10, B25 | Wheat rhizosphere soil (Setif region) | 4/20 (20) |
| Ba1, Ba8, Ba13 | Wheat rhizosphere soil (Boussaada region) | 3/11 (27) |
| D4, D6, D12 | Barley rhizosphere soil (Djelfa region) | 3/9 (33.3) |
| E17, C16 | The marvelous cave (Jijel region) | 2/200 (1) |
| HOT1, HOT2, HOT3, HOT4, HOT5 | Thermal spring of Hammam Ouled Tebben-Setif | 5/8 (62.5) |
| Sampling Sites | Sampling Site Coordinates | Number of Isolates |
|---|---|---|
| Archaeological site (rocks and Roman ruins) of Djemila-Setif | 36.319096, 5.736135 | 70 |
| Rhizosphere of Terfez (Boussaada region) | 35.217234, 4.273263 | 80 |
| Wheat rhizosphere soil (Setif region) | 36.316769, 5.448367 | 20 |
| Wheat rhizosphere soil (Boussaada region) | 35.348036, 4.408568 | 11 |
| Barley rhizosphere soil (Djelfa region) | 35.062900, 3.535154 | 9 |
| The marvelous cave (Jijel region) | 36.667344, 5.475292 | 200 |
| Thermal spring of Hammam Ouled Tebben-Setif | 35.787571, 5.123604 | 8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Balla, A.; Silini, A.; Cherif-Silini, H.; Bouket, A.C.; Boudechicha, A.; Luptakova, L.; Alenezi, F.N.; Belbahri, L. Screening of Cellulolytic Bacteria from Various Ecosystems and Their Cellulases Production under Multi-Stress Conditions. Catalysts 2022, 12, 769. https://doi.org/10.3390/catal12070769
Balla A, Silini A, Cherif-Silini H, Bouket AC, Boudechicha A, Luptakova L, Alenezi FN, Belbahri L. Screening of Cellulolytic Bacteria from Various Ecosystems and Their Cellulases Production under Multi-Stress Conditions. Catalysts. 2022; 12(7):769. https://doi.org/10.3390/catal12070769
Chicago/Turabian StyleBalla, Amel, Allaoua Silini, Hafsa Cherif-Silini, Ali Chenari Bouket, Amel Boudechicha, Lenka Luptakova, Faizah N. Alenezi, and Lassaad Belbahri. 2022. "Screening of Cellulolytic Bacteria from Various Ecosystems and Their Cellulases Production under Multi-Stress Conditions" Catalysts 12, no. 7: 769. https://doi.org/10.3390/catal12070769
APA StyleBalla, A., Silini, A., Cherif-Silini, H., Bouket, A. C., Boudechicha, A., Luptakova, L., Alenezi, F. N., & Belbahri, L. (2022). Screening of Cellulolytic Bacteria from Various Ecosystems and Their Cellulases Production under Multi-Stress Conditions. Catalysts, 12(7), 769. https://doi.org/10.3390/catal12070769