Abstract
The whitefish industry generates a huge amount of rest raw material, which is currently wasted or underutilized in the production of low-value products such as animal feed. While fish muscle is the primary product of use for human consumption, rest raw material has great potential as a source of protein and bioactive peptides for the production of food ingredients and nutraceuticals. Enzymatic hydrolysis is a biotechnological processing method that can be used to extract protein from fish rest raw material into a protein hydrolysate. This study aimed at investigating the functionality of ultrafiltration as an industrial processing method and its effect on the bioactivity of protein hydrolysates. Protein hydrolysates were produced by enzymatic hydrolysis of saithe (Pollachius virens) head and backbone caught at two separate occasions to investigate the effect of seasonal variations. Ultrafiltration effectively concentrated larger peptides (>4 kDa) and smaller peptides (<4 kDa) in separate fractions, with a protein yield of 31% in the fraction <4 kDa. The unfiltered hydrolysate was found to have a higher antioxidative activity compared to the <4 kDa fraction in ABTS, FRAP, and ORAC assays. These results indicate that ultrafiltration does not effectively increase bioactivity by concentrating small peptides and that bioactivity is dependent on several properties, including interaction with larger peptides.
1. Introduction
It is well documented that fish is an excellent source of several health-beneficial nutritional components, such as polyunsaturated fatty acids and protein with a well-balanced amino acid composition []. The consumption of pure muscle in the form of filet or processed fish products is the most common way of including fish protein as part of the human diet. In addition, niche products consisting of heads, roe, and liver are consumed in some countries, but fish rest raw material (RRM) is predominantly an underutilized resource. RRM can be defined as the parts of the fish that are not the primary product of use, which in most cases is the filet; RRM includes heads, backbones, and viscera []. Fish heads and backbones have a high protein content of 15–20% [,,,]. However, RRM needs to be processed in order to be suitable as food or food ingredients for human consumption due to the nature of its composition and appearance. Biotechnological processing by enzymatic hydrolysis can be used to extract proteins from the RRM by the hydrolytic activity of endogenous or commercial enzymes. The enzymatic activity results in solubilization of the proteins, which then are extracted as the main product of enzymatic hydrolysis: the fish protein hydrolysate (FPH). Hence, the processing of fish RRM enables a better utilization of fish as a food resource, which is beneficial from both sustainability and a bioeconomical perspectives [,]. In addition to being an excellent source of amino acids for human metabolism, fish RRM can be a source of bioactive peptides that are inactive as part of the native protein but can be released through a hydrolytic process []. Bioactive peptides are small molecules, ranging from three to 20 amino acids, with inherent health-beneficial properties beyond that of normal nutrition [,]. Several studies have identified bioactive peptides in whitefish [,]. Cod protein hydrolysates have been found to possess both antioxidative and blood pressure reducing activities in vitro [,,,,], while in vivo animal studies have been less conclusive [].
Reactive oxygen species (ROS) are generated continuously and unavoidably through cellular respiration in our body and during lipid oxidation in foods and are therefore a cause of great concern both regarding human health and food stability. Oxidative stress, defined as an unbalance between ROS and endogenous antioxidants, has been linked to several adverse health effects and human diseases, and lipid oxidation in food can cause a reduction of shelf-life and nutritional value as well as unwanted changes to odor and texture [,]. Antioxidants are molecules capable of reducing oxidative stress and lipid oxidation in food by inhibiting oxidation []. These molecules are naturally occurring in living tissue, and both natural and synthetic antioxidants are used as additives in various foods. However, commonly used synthetic antioxidants, such as butylated hydroxyanisole (BHA) and butylated hydroxytoluene (BHT), have received negative attention due to the toxicity of these in high concentrations [,]. Thus, there is a strong interest in finding natural alternatives of antioxidants for food purposes but also for the use as nutraceuticals []. For any substance to act as an antioxidant, it must be able to neutralize ROS or other prooxidative components, thereby preventing them from causing oxidative damage in food systems or human tissue. This can be achieved by several mechanisms, including the scavenging of ROS and metal chelation []. In turn, ROS can be neutralized by hydrogen donation or electron transfer, which are abilities measured by the oxygen radical absorbance capacity (ORAC) and 2.2′-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid) diammonium salt (ABTS)/ferric-reducing antioxidant power (FRAP) assays, respectively. A combination of assays is preferred to analyze antioxidative activity, as the individual assays are limited by their specific conditions and the mechanisms they assess [,].
The Norwegian whitefish industry generates approximately 300,000 tons of RRM each year []. Improving the utilization of RRM is fundamental for making fisheries more sustainable to meet the UN Sustainable Development Goals (SDGs). Sustainable fisheries are crucial to improve the utilization of available food resources (SDG 12), reduce hunger and malnutrition by making available more healthy food (SDG 2), and prevent climate change (SDG 13) []. This study aimed at investigating the possibility of extracting and concentrating peptides from saithe (P. virens) RRM by enzymatic hydrolysis and membrane ultrafiltration (UF). UF is a processing and refinement method that can be used to concentrate low molecular weight peptides of a hydrolysate, which could potentially increase the bioactivity of this fraction [,,]. Head and backbone from saithe caught at two separate occasions, to account for seasonal variation in spawning, were processed by enzymatic hydrolysis in bioreactors to extract proteins. The resulting saithe protein hydrolysates (SPH) were further fractionated using UF with a 4 kDa molecular weight cut-off (MWCO) membrane to concentrate small peptides. The entire processing was conducted in pilot scale to simulate an industrial process. As this study is part of a larger experimental design aiming to increase the utilization of saithe RRM, all experiments were conducted with a continuous focus on the adaptability of the process to industrial processing lines. The potential for implementing enzymatic hydrolysis in the Norwegian whitefish industry was discussed by Hjellnes et al. []. UF applied in an industrial setting would entail extra cost and logistical challenges, which means that the value of the processing product must be high to justify the cost–benefit balance. To the authors’ knowledge, few studies have investigated the effect of membrane filtration on the bioactivity of whitefish hydrolysates [,,,], and most studies have focused on cod. Therefore, an investigation of intraspecies similarities between cod and saithe, as well as the potential effect of spawning on bioactivity, was of interest in this study.
2. Results and Discussion
2.1. Membrane Filtration and Protein Flow
Prior to filtration with 4 kDa molecular weight cut-off membrane (MWCO), 5 grams of SPH was dissolved in 500 mL of distilled water and filtrated through a ceramic membrane with 150 kDa MWCO in order to remove high molecular weight peptides and unwanted compounds such as lipids. Such compounds could otherwise cause clogging and the formation of a dynamic cake on membranes with a lower MWCO, reducing the filtration efficiency []. The initial filtration distributed the protein content of SPH in a retentate (R150) and a permeate (P150), containing peptides larger and smaller than 150 kDa, respectively. Ultrafiltration (UF) was subsequently performed on P150 using PESH membranes with a MWCO of 4 kDa, creating a new retentate (R4, >4 kDa) and permeate (P4, <4 kDa).
UF of the eight SPH from enzymatic hydrolysis of saithe heads (H) and backbone (B), in two bioreactors (I, II), from October (HOI, HOII, BOI, BOII) and January (HJI, HJII, BJI, BJII) resulted in a mass distribution presented in Figure 1. Filtration over 150 kDa MWCO ceramic membranes resulted in 1.42 ± 0.08 g SPH in R150 and 2.78 ± 0.05 g SPH in P150, which equals a protein yield of 56% in P150. The subsequent UF over 4 kDa cut-off PESH membranes resulted in 1.08 ± 0.08 g SPH in R4 and 1.56 ± 0.16 g in P4, which is equivalent to a protein yield of 31% of the initial 5 g of SPH in P4. The total protein loss from both filtrations was 19%.
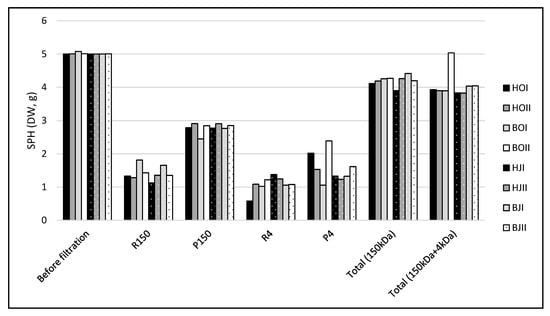
Figure 1.
Mass distribution (n = 1) of saithe protein hydrolysates (SPH) before filtration and after filtration with 150 kDa cut-off ceramic membrane (retentate: R150, permeate: P150), and subsequently 4 kDa cut-off PESH membranes (retentate: R4, permeate: P4). Total mass of R150 and P150 (Total (150kDa)) and total mass of R150, R4, and P4 (Total (150 kDa + 4 kDa) is included. SPH were obtained from the enzymatic hydrolysis of heads (H) and backbones (B), in two bioreactors (I, II), from saithe caught in October 2019 (HOI, HOII, BOI, BOII) and January 2020 (HJI, HJII, BJI, BJII).
Based on an initial protein content of 16.3% and 19.2% in saithe head and backbone, respectively [], enzymatic hydrolysis followed by UF over 150 kDa and 4 kDa MWCO membranes would give a processing protein flow, as presented in Figure 2.
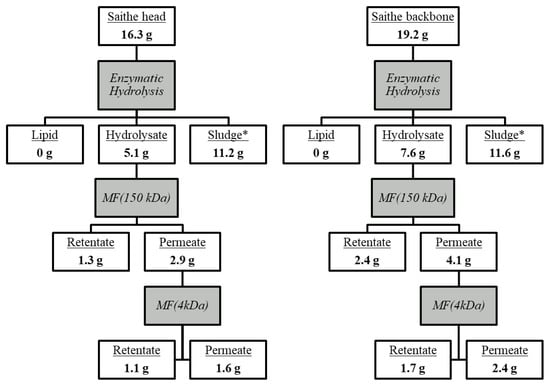
Figure 2.
Protein flow (g) from raw material to product after processing of 100 g minced saithe heads (left) and backbones (right) by enzymatic hydrolysis [] and filtration with a 150 kDa cut-off ceramic membrane followed by a 4 kDa cut-off PESH membrane. *Calculated value.
The processing of 100 g minced saithe heads and backbone yielded 5.1 g and 7.6 g protein in the hydrolysate, while 1.6 g and 2.4 g would end up in the P4, respectively. In an industrial processing line, where the goal is to obtain a pure protein fraction from fish RRM and isolating peptides with high bioactivity, SPH and P4 would be the main products of interest. This means that a large fraction of RRM protein would end up in the less regarded processing products. For the processing to be beneficial from an economic point of view, P4 would have to generate high incomes as specialized products for human consumption. However, for the processing to be sustainable, it would also be very important to find areas of application for the sludge, R150, and R4. The sludge fraction could be used for gelatin extraction and animal feed production [,], while the functional characteristics of the larger peptides in R150 and R4 could make them suitable as food ingredients with water-binding, emulsifying, and gelling properties []. It is also worth noting that it is not common practice to report protein yield of UF processing, which in the authors’ opinion is a highly relevant factor in evaluating the potential for industrialization.
2.2. Amino Acid Composition
The amino acid composition of RRM and SPH from enzymatic hydrolysis of heads (HOI, HOII) and backbones (BOI, BOII) from saithe captured in October 2019 and heads (HJI, HJII) and backbones (BJI, BJII) from saithe captured in January 2020 are presented in Table 1. Glu (7.73 ± 1.75–17.13 ± 1.06 g/100 g protein), Gly/Arg (5.03 ± 0.59–17.04 ± 0.26 g/100 g protein), and Asp (5.34 ± 0.59–11.57 ± 0.73 g/100 g protein) were found to be the dominating amino acids in both RRM and SPH. Fish protein is known to have a high content of the essential amino acids Lys and Leu [], which was also confirmed by this study, where the Lys and Leu contents were found to be 5.03 ± 0.54–9.07 ± 0.59 g/100 g protein and 3.44 ± 0.38 g/100 g protein, respectively. These results are in correspondence with Farvin et al. [], who found Gly, Glu, Lys, and Ala to be the predominant amino acids in commercial cod protein hydrolysates; Jensen et al. [], who found Glu, Asp, Ala, Leu, and Lys to be the dominating amino acids of cod muscle; Girgih et al. [], who found Glu/Gln, Asn/Asp, Arg, Lys, and Leu to be the dominating amino acids in cod backbone hydrolysate >1 kDa; and Remme and Austnes [], who found Asp, Glu, and Gly to be the dominating amino acids in cod head hydrolysates.

Table 1.
Total amino acid composition (g/100 g protein) in RRM and SPH obtained from enzymatic hydrolysis of heads (H) and backbones (B) in two bioreactors (I, II) from saithe caught in October 2019 (HOI, HOII, BOI, BOII) and January 2020 (HJI, HJII, BJI, BJII) ( ± SEM, n = 3). Essential [] and hydrophobic [] amino acids are indicated by e and h, respectively. * Calculated value.
Spawning did not seem to have a significant effect on the amino acid composition of RRM nor SPH. However, backbone RRM was found to have a significantly higher content of Asp (F(1,10) = 14.03, p < 0.05), Glu (F(1,10) = 6.06, p < 0.05), Val (F(1,10) = 9.04, p < 0.05), Ile (F(1,10) = 25.84, p < 0.05), Leu (F(1,10) = 26.14, p < 0.05), and Lys (F(1,10) = 13.67, p < 0.05), and a significantly lower content of His (F(1,10) = 7.28, p < 0.05) and Gly/Arg (F(1,10) = 15.37, p < 0.05), compared to head RRM. Most amino acids were also found to be prevalent in higher amounts in RRM compared to SPH. The largest difference between head RRM and backbone RRM was found for Ile and Leu, which is interesting from a nutritional perspective considering that both of there are essential hydrophobic amino acids [,]. A product higher in essential amino acids might be more attractive both as a food ingredient and a nutraceutical.
However, the difference in amino acid composition between head RRM and backbone RRM was not reflected in their respective SPH. The only detectable trend was SPH from backbone having a significantly lower content of Gly/Arg (F(1,22) = 24.09, p < 0.05) compared to SPH from head. For the remaining amino acids, equal or higher amounts were found in SPH from head compared to SPH from backbone. However, the protein yield in SPH from backbone after enzymatic hydrolysis was found to be higher than in SPH from head []. This might indicate that hydrophobic amino acids, of which Ile, Leu, and Val were detected in higher amounts in backbone RRM, are harder to extract in the water-soluble phase during enzymatic hydrolysis. It is also evident from Table 1 that there are variations between the individual hydrolysates, indicating that it can be difficult to achieve a standardized processing outcome, which again might mask potential differences between SPH from head and backbone.
Although the amount of Trp, Cys, and Pro could not be detected due to the analytical method used in this study, several of the other essential and conditionally essential amino acids were prevalent in RRM and SPH. This is especially relevant when considering the nutritional quality of a protein product. The protein efficiency ratio (PER) test is a method used to evaluate the quality of a protein source based on the weight gain of rats when fed a diet consisting of 10% of the evaluated protein []. However, calculations on relative quantities of specific amino acids, yielding a PER value, can be used as an estimator for protein quality. The calculated PER values for RRM (1.93–3.04) and SPH (1.31–1.53) are presented in Table 1. These results correspond to those of Šližytė et al. [], who found PER values of 2.99 for cod muscle and 1.60–2.27 for hydrolysates from hydrolysis of cod backbone and viscera under similar conditions using Flavorzyme and Neutrase. PER values were found to be significantly (T(9) = 3.345, p = 0.05) higher for RRM compared to SPH, indicating that several essential amino acids end up in the sludge. This further substantiates the importance of investigating the possibility of using the sludge fraction as a source of nutrition as well. The sludge was found to be the fraction where the majority of the RRM dry matter ended after enzymatic hydrolysis []. Thus, finding areas of application is important in order to minimize the amount of processing waste while simultaneously maximizing the value addition of the rest raw material.
The amount of His was found to be 0.42 ± 0.14–1.07 ± 0.09 g/100 g protein in SPH. In addition to being an essential amino acid, His has been associated with antioxidative properties due to the ability of its imidazole to form complexes with several metal ions and scavenge ROS [,]. The metal ion-chelating abilities of His is, among other physiological functions, important for the binding of iron in hemoglobin and myoglobin []. Trp and Gly, but also Cys, Ser (2.30 ± 0.53–4.98 ± 0.24g/100 g protein in SPH), Lys (5.03 ± 0.54–6.57 ± 0.13 g/100 g protein in SPH), and Pro, have been shown to potentially inhibit oxidation []. Despite a relatively low His content, SPH are high in Gly/Arg (5.03 ± 0.59–14.27 ± 0.53 g/100 g protein), Ser, and Lys and might thus be a source of bioactive peptides with antioxidative properties. Based on the higher content of Gly/Arg, SPH from head is likely to have better antioxidative properties compared to SPH from backbone. However, the antioxidative activity in peptides is not solely dependent on the specific amino acid present but also the sequence of their arrangement and the overall peptide composition [,]. The overall hydrophobicity is also important for bioactive peptides to be able to exert their antioxidative activity by interacting with lipid systems in both our body and in foods []. Several hydrophobic amino acids were present in SPH, including Val (2.23 ± 0.25–3.63 ± 0.15 g/100 g protein) and Leu (3.44 ± 0.38–4.90 ± 0.18 g/100 g protein). However, Leu was found to be one of the most prevalent free amino acids in SPH, which might negatively affect its contribution to antioxidative activity and reduce bioavailability []. In general, the amount of free amino acids in SPH was low, ranging from 42.78 to 69.50 mg/g. This was expected, considering that both enzymes used for the enzymatic hydrolysis were endopeptidases, the low DH (14.8–18.1%) of SPH, and that head and backbone are RRM fractions with low endogenous enzyme activity [].
2.3. Molecular Weight Distribution
The molecular weight distribution of SPH and P4 from saithe head and backbone are presented in Figure 3, Figure 4, Figure 5 and Figure 6.
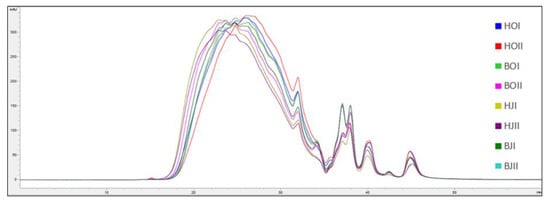
Figure 3.
Chromatograms (mAU/min) from analysis of molecular weight distribution ( ± SEM, n = 2) of saithe protein hydrolysates obtained from enzymatic hydrolysis of heads (H) and backbones (B), in two bioreactors (I, II), from saithe caught in October 2019 (HOI (blue), HOII (red), BOI (green), BOII (pink)) and January 2020 (HJI (beige), HJII (purple), BJI (dark green), BJII (turquoise)).
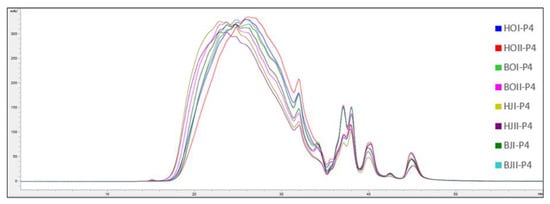
Figure 4.
Chromatograms (mAU/min) from analysis of molecular weight distribution ( ± SEM, n = 2) of permeates (P4) after ultrafiltration (<4 kDa) of saithe protein hydrolysates obtained from enzymatic hydrolysis of heads (H) and backbones (B), in two bioreactors (I, II), from saithe caught in October 2019 (HOI-P4 (blue), HOII-P4 (red), BOI-P4 (green), BOII-P4 (pink)) and January 2020 (HJI-P4 (beige), HJII-P4 (purple), BJI-P4 (dark green), BJII-P4 (turquoise)).
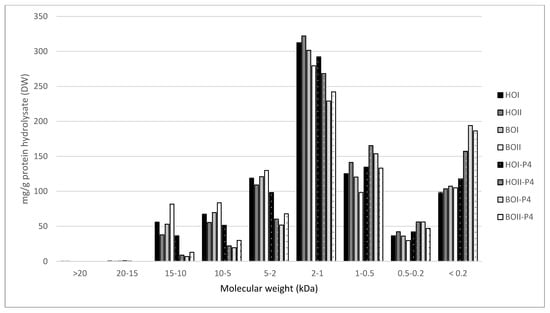
Figure 5.
Molecular weight distribution ( ± SEM, n = 2) of saithe protein hydrolysates (SPH) (HOI, HOII, BOII, BOII) and their corresponding permeates (HOI-P4, HOII-P4, BOI-P4, BOII-P4) after ultrafiltration (<4 kDa). SPH were obtained from enzymatic hydrolysis of heads (H) and backbones (B), in two bioreactors (I, II), from saithe caught in October 2019 (HOI, HOII, BOI, BOII).
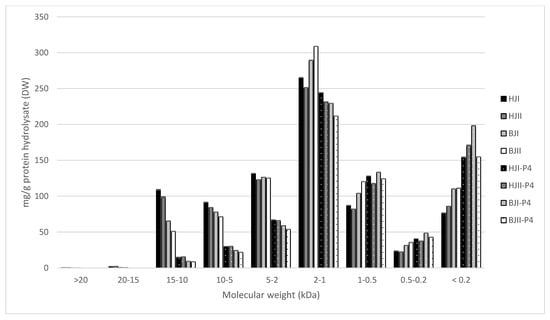
Figure 6.
Molecular weight distribution ( ± SEM, n = 2) of saithe protein hydrolysates (SPH) (HJI, HJII, BJII, BJII) and their corresponding permeates (HJI-P4, HJII-P4, BJI-P4, BJII-P4) after ultrafiltration (<4 kDa). SPH were obtained from enzymatic hydrolysis of heads (H) and backbones (B), in two bioreactors (I, II), from saithe caught in January 2020 (HJI, HJII, BJI, BJII).
Spawning did not affect the molecular weight distribution of SPH and P4, and no difference was observed between SPH and P4 produced from head RRM compared to backbone RRM. SPH were found to have a significantly higher content of peptides in the size range 15–10 kDa, 10–5 kDa, 5–2 kDa, and 2–1 kDa, while P4 were found to have a significantly higher content of peptides in the size range 1–0.5 kDa, 0.5–0.2 kDa, and <0.2 kDa. The largest difference was observed for the size ranges 10–5 kDa (T(30) = 12.58, p < 0.05) and 5–2 kDa (T(30) = 14.71, p <0.05), where the SPH average was found to be 75.08 mg/g and 123.04 mg/g, respectively. P4 were found to have on average 28.69 mg/g peptides in the size range 10–5 kDa and 65.44 mg/g peptides in the size range 5–2 kDa.
These results indicate that peptides >5 kDa to a large degree have been concentrated in R4 after UF, and that the majority of peptides in P4 have a molecular weight <4 kDa. However, the MWCO at 4 kDa is not clear cut, as is evident from the presence of peptides with molecular size 5–15 kDa in P4. This lack of a sharp separation of peptides during UF has also been demonstrated in other studies [,]. Chabeaud et al. [,] suggested potential fouling, lowered flux and insufficient membrane cleaning to be reasons for the poor separation of peptides. In their studies on UF of fish protein hydrolysates, they found that peptides <0.5 kDa were better separated at lower concentrations of saithe protein hydrolysate (30 g/L vs. 150 g/L) and lower pressure (1 bar vs. 5 bar). Thus, it is likely that parameters of UF can be adjusted in order to improve size-based separation of peptides.
Of the total peptide content of SPH, 23.8–35.6% were found to have a molecular size <1 kDa. Combined with a relatively low DH and the low content of free amino acids, these results indicate that SPH contains considerable amounts of larger peptides []. A less extensive hydrolysis might be beneficial for obtaining SPH with desired functional characteristics such as water binding, emulsification, and foaming [,,]. SPH containing larger peptides might also be less bitter, considering that bitterness is mainly associated with low molecular weight peptides and hydrophobicity [,]. Sensory properties will in turn be decisive for consumer acceptance. Thus, size-based separation of peptides in SPH by UF has the potential to generate fractions with improved functional and sensory properties (R4 and R150) as well as fractions with increased bioactivity (P4).
2.4. Antioxidative Activity
The results from analysis of antioxidative activity in SPH, P4, R4 and R150 generated from enzymatic hydrolysis of saithe head (H) and backbone (B), and subsequent UF, is presented in Table 2. Three different in vitro assays were used to evaluate the antioxidative activity of SPH and UF fractions: ABTS, FRAP, and ORAC. These assays measure the ability of bioactive peptides to neutralize ROS by electron transfer, reduce Fe3+, and neutralize ROS by hydrogen transfer, respectively. A combination of assays is preferred, as each assay measures a specific type of antioxidative activity within the assay-specific conditions and not the total antioxidative activity [].
Table 2.
Antioxidative activity ( ± SEM, n = 12) of saithe protein hydrolysates (SPH), permeates (P4), and retentates (R4, R150) after membrane filtration with a 150 kDa cut-off ceramic membrane and 4 kDa cut-off PESH membrane. SPH were obtained from enzymatic hydrolysis of saithe heads (H) and backbones (B). Results from ABTS are given as µmol/g propyl gallate equivalents, FRAP, and ORAC are given as µmol/g Trolox equivalents. Results of Tukey’s post hoc grouping are indicated (H: h1–3, B: b1–3) for the individual analysis.
Spawning was not found to affect the antioxidative activity of SPH, P4, R4, and R150, and thus, results are presented as average values for heads (H) and backbones (B) from October 2019 and January 2020 to include results of ANOVA and Tukey’s post hoc test (Table 2). SPH and UF fractions from backbone were found to have a significantly higher antioxidant activity compared to head measured by all assays (ABTS (T(93) = 9.23, p < 0.05), FRAP (T(94) = 6.19, p < 0.05), and ORAC (T(70) = 1.40, p < 0.05). This indicates that more antioxidative peptides are released from backbone during enzymatic hydrolysis compared to heads, which is somewhat contradictory to the observed higher prevalence of amino acids with known antioxidative activity in SPH from head (Section 2.2). However, this confirms the assumption that amino acid composition cannot be used as the sole indicator of antioxidative activity.
ABTS antioxidative activity was found to be significantly different between SPH and UF fraction for both head (F(3,43) = 111.75, p < 0.05) and backbone (F(3,44) = 14.93, p < 0.05). A Tukey post hoc test revealed that SPH had a significantly higher ABTS antioxidative activity compared to all other fractions. Both retentates (R4 and R150) from backbone SPH had significantly higher ABTS antioxidative activity compared to the permeate (P4), while no significant difference was observed between retentate R150 and permeate from head SPH. When measured by FRAP assay, a significant differences was observed between SPH and the UF fractions for both head (F(3,43) = 106.76, p < 0.05) and backbone (F(3,44) = 21.49, p < 0.05). Retentate R150 had the significantly highest FRAP antioxidative activity in both head and backbone. Retentate R4 and the permeate were both significantly higher than SPH for head, while the permeate was found to have the significantly lowest activity measured by FRAP for backbone. Regarding ORAC antioxidative activity, significant differences between fractions were observed only for backbone (F(3,44) = 5.88, p < 0.05), where SPH and P4 were found to have a significantly higher ORAC antioxidative activity compared to R4 and R150. This suggests that small peptides are important for ORAC antioxidative activity, but that a mixture of small and larger peptides are equally effective as the concentrated permeate.
No correlation was found between the results obtained from ABTS, FRAP, and ORAC. SPH was found to have the highest antioxidative activity when measured by both ABTS and ORAC but among the lowest when measured by FRAP. FRAP and ABTS assay both evaluate the ability of an antioxidant to reduce an oxidant with a comparable redox potential, and therefore, these results might be more comparable than results from ORAC assay, which measures the ability of an antioxidant to neutralize peroxide radicals by hydrogen donation [,,]. However, while ABTS and ORAC assays are carried out at neutral pH, the FRAP assay is conducted in acidic conditions, which could suppress the antioxidative activity of bioactive peptides due to protonation [,]. This could explain the lower antioxidative activity of SPH found in the FRAP assay compared to the ORAC and ABTS assay. However, the acidic condition of the FRAP assay could be relevant when considering the stability and bioavailability of antioxidative peptides in the human gastrointestinal system. An instability of bioactive peptides in acidic conditions could also explain why R150 showed the highest antioxidative activity in the FRAP assay, as this UF fraction consisted of larger peptides that are possibly more resistant to changes in pH.
Another trend that can be observed from Table 2 is that the antioxidative activity of P4 was found to be equal to or lower than SPH, R4, and R150 in all three assays. These findings contradict what was the expected outcome of SPH processing. UF was applied to SPH with the intention of concentrating low molecular weight peptides, which are associated with higher bioactivity. Under this assumption, the expected result for antioxidative activity for UF fractions would be P4 > R4 > R150. SPH, having a smaller concentration of low molecular weight peptides relative to 4, would also be expected to exhibit lower bioactivity. Several studies on the UF of hydrolysates from various raw materials have confirmed this theory, including that of Farvin et al. [], who found that applying UF to commercial cod hydrolysate significantly increased antioxidative activity. However, both Girgih et al. [] and Picot et al. [] found the unfractionated hydrolysate to have the highest antioxidative activity after UF of cod frame digests and commercial hydrolysates from cod and pollock skin, respectively. These studies both involved lower MWCO and lower molecular weight peptides, which indicates that further separation might not be effective for increasing bioactivity. This was further confirmed in this study by unpublished data on preliminary experiments with UF using 2 kDa MWCO membranes on SPH.
Regardless of the similar trends observed for SPH and UF fractions in this study, the measured antioxidative activity varies considerably among the assays. This inconsistency in the degree of antioxidative activity was also reported by Kristinova et al. [] when comparing the results of Folin–Ciocalteu, DPPH, and ABTS assays, and it can be a result of limitations of the methodology of the assays as discussed above. Therefore, it can be challenging to compare results from various assays both within and among studies [,]. Furthermore, it is important to take into consideration that in vitro methods for assessing antioxidative activity cannot be directly extrapolated to functionality in food or the human body, as they do not consider intermolecular interaction, stability, or bioavailability [].
The use of UF processing in an industrial setting would require an investment in both expensive equipment and expertise. Based on published results, it is also questionable whether such processing is likely to yield a product with increased bioactivity that could target a high-value market, which would be necessary to cover the extra cost. However, further studies are needed to refine and adapt the UF process to fish protein hydrolysates. In the event of an adapted process, where the fractions generated can be utilized based on their specific functional properties, e.g., water binding, emulsification, foaming, etc. for retentates and bioactivity for permeates, it would be possible to optimize the utilization of fish RRM proteins.
3. Materials and Methods
3.1. Chemicals
2,2′-Azino-bis(3-ethylbenzothiazoline-6-sulfonic acid (ABTS), potassium persulfate, ferric chloride, 2,3,5-triphenyltetrazolium chloride (TPTZ), 2,2′-azobis-(isobuttersa ureamidin)-dihydroclorid) (AAPH), propyl gallate, Trolox, acetate, phosphate, hydrochloric acid, methanol, and ethanol (Merck Life Science AS, Oslo, Norway) were used for the experiments and chemical analysis. All chemicals were of analytical grade.
3.2. Enzymatic Hydrolysis
The experimental procedure was conducted as described by Hjellnes, Rustad, and Falch [] following the factorial design illustrated in Figure 7. Saithe were caught in Trondheimsfjorden, Norway, at two separate occasions: pre-spawning saithe in October 2019 (O) and spawning saithe in January 2020 (J). The fish was hand filleted, and the RRM was separated into three fractions: viscera (V), head (H), and backbone (B). Heads and backbones were used for further analysis.
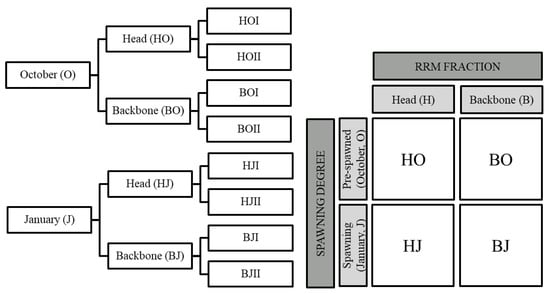
Figure 7.
Schematic overview of the samples generated from (left), and the factorial design used for (right) enzymatic hydrolysis of saithe head and backbones from October 2019 (pre-spawned) and January 2020 (spawning) [].
The enzymatic hydrolysis was conducted on 500 g of minced RRM mixed 1:1 with preheated water (50°) for 60 min at 50 °C, physiological pH (pH = 6.0), and with the addition of 0.1% (1:1) papain (from Carica papaya, 1.5–10 U/mg, EC 3.4.22.2, Merck Life Science AS, Oslo, Norway) and bromelain (from Ananas comosus, 3 U/mg, EC 3.4.22.32, Merck Life Science AS, Oslo, Norway) in bioreactors (Syrris Atlas, Model No. 2101000, Nerliens Meszansky AS, Oslo, Norway) with thermostats (Huber Ministat 125, Nerliens Meszansky AS, Oslo, Norway). A total of four RRM combinations (HO, BO, HJ, BJ) were hydrolyzed in two parallels, as presented in Figure 7. The enzymatic reaction was terminated by heat inactivation (>90 °C, 10 min), and the reaction mixture was transferred to 18 centrifugal tubes (50 mL), centrifuged (10,900× g, 10 min, 20 °C) and frozen (–20 °C). The resulting three fractions, oil, hydrolysate, and sludge, were separated using a scalpel on frozen sample. Saithe protein hydrolysates (SPH) were further freeze dried (Labconco FreeZone 12, Labcono Corporation, Kansas City, MO, USA, −50 °C, <13.3 Pa) and frozen at −40 °C until further processing.
3.3. Membrane Ultrafiltration
Membrane ultrafiltration (UF) was conducted using a pilot-scale system (MMS Membrane Solution Triple System, DUE MILJØ AS, Halden, Norway) measuring 700 × 510 × 510 mm. The system had an 800 mL feed tank and a hold-up volume of 50 mL. A single membrane cell and three serially coupled membranes cells, each with an individual area of 28 cm2, were used for the experiment. A ceramic membrane with a molecular weight cut-off (MWCO) of 150 kDa (Nadir®, MMS Nordic, Silkeborg, Denmark) was used for the single membrane cell, and PESH membranes (Nadir® UH004 p, MMS Nordic, Silkeborg, Denmark), with a thickness of 210–250 µm and a MWCO of 4 kDa, were used for the three serial coupled membranes.
SPH were resuspended in distilled water to a concentration of 1% (10 g/L) and then filtered through the ceramic membrane under a pressure of 500 kPa (Flux: 50 L/m2h) at 21 °C until hold-up volume was reached. The filtration yielded two fractions: a retentate with peptides larger than 150 kDa (R150) and a permeate with peptides smaller than 150 kDa (P150). P150 was subsequently filtrated through the PESH membranes (500 kPa, 21 °C), which generated two new fractions: a retentate with peptides larger than 4 kDa (R4) and a permeate with peptides smaller than 4 kDa (P4). R150, R4, and P4 were freeze dried (Labcono FreeZone 12, −50 °C, <13.3 Pa) and frozen at −40 °C until further analysis. An overview of the processing steps and generated products is presented in Figure 8.
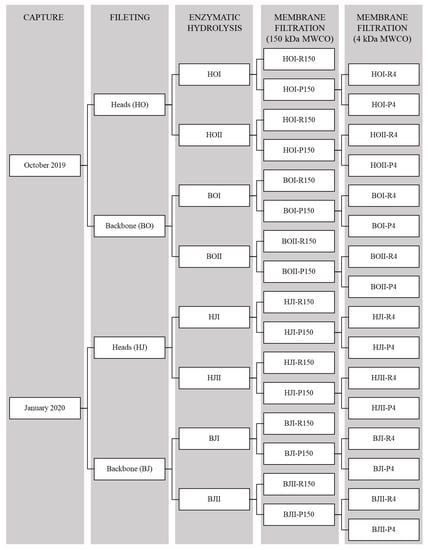
Figure 8.
Overview of the processing steps and products generated from capture, filleting, and enzymatic hydrolysis of saithe rest raw material [] and two membrane filtrations of saithe protein hydrolysates.
3.4. Dry Matter
Dry matter was analyzed gravimetrically according to AOAC []. The samples were analyzed in triplicates.
3.5. Antioxidative Assays
The antioxidative activity of saithe protein hydrolysates was analyzed by 2.2′-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid) diammonium salt (ABTS) radical scavenging activity, ferric-reducing antioxidant power (FRAP) assay, and oxygen radical absorbance capacity (ORAC) assay. ABTS was analyzed according to Re et al. [] and Nenadis et al. [] using propyl gallate as the reference compound []. FRAP was analyzed according to Benzie and Strain [], and ORAC was analyzed according to Dávalos et al. [] with previously described modifications [] using Trolox as the reference compound. Values were reported as µmol/g propyl gallate equivalents and µmol/g Trolox equivalents, respectively. The samples were analyzed in triplicate.
3.6. Amino Acid Composition
Total amino acid composition was analyzed according to Blackburn []. Samples equivalent to 50 mg protein were hydrolyzed with 1 mL 6 M HCl for 22 h at 105 °C. The samples were neutralized (pH = 6.5–7.5) with NaOH and filtered through Whatman GF/C filters (25 cm, GE Healthcare, Boston, MA, USA) using a vacuum pump (Heto MASTER JET, Heto Lab Equipment AS, Allerød, Denmark). Following dilution, the samples were filtered through 0.2 µM syringe filters (VWR International, Oslo, Norway). HPLC (Ultimate 300 dionex, Nova-pak c184 µM 3.9 × 150 mm column, Thermo Fischer Scientific, Waltham, MA, USA) using a fluorescence detector (Rf 200) was performed by Siri Stavrum, NTNU. The samples were analyzed in triplicate.
3.7. Protein Efficiency Ratio (PER) Value
PER values were calculated as described by Šližytė et al. [] using the following Equation (1):
where ΣAA7 = Thr + Val + Met + Ile + Leu + Phe + Lys.
3.8. Molecular Weight Distribution
The molecular weight distribution of SPH and P4 was analyzed by High-Performance Liquid Chromatography (HPLC) by Innolipid AS. Samples were dissolved in distilled water (10 mg/mL) and filtered through a low-protein-binding microfilter (0.22 µm) to remove large peptides and proteins. The analysis was performed on a Superdex column using a wavelength of 214 nm and hydrolyzed albumin as standard. The samples were analyzed in duplicate.
3.9. Statistical Analysis
All statistical analysis was conducted in SPSS software (IBM SPSS Statistics 27, 2020, International Business Machines (IBM), Armonk, NY, USA). Analysis of variance (ANOVA) and t-test were used for the comparison of means for >3 and <3 parallels respectively, assuming normal distribution and equal variance. Tukey’s post hoc test was used in combination with ANOVA to identify significant differences. Significance level was set to p < 0.05. Results are reported as mean values () of [n] number of parallels ± standard error of the mean (SEM).
4. Conclusions
This study aimed at investigating the functionality of ultrafiltration (UF) as an industrial processing method, and its effect on the bioactivity of saithe protein hydrolysates (SPH). An initial filtration was performed on SPH with a 150 kDa MWCO membrane, creating a retentate (R150, >150 kDa) and a permeate (P150, <150 kDa). UF was subsequently performed on P150 using a 4 kDa MWCO membranes, creating a new retentate (R4, >4 kDa) and permeate (P4, <4 kDa).
UF effectively concentrated small peptides in P4. UF processing was expected to increase the antioxidative activity of this peptide fraction, as small peptides have been associated with bioactivity. However, unfiltered SPH was found to have a similar or even higher antioxidative activity compared to P4 measured by ABTS, FRAP, and ORAC assays. These results indicate that concentrating small peptides by UF does not effectively increase bioactivity, and that bioactivity is dependent on several properties, including interaction with larger peptides. No correlation was observed between the results of ABTS, FRAP, and ORAC, and the measured antioxidative activity varied considerably among the assays. This indicates that it can be difficult to compare results among different assays both within and between studies.
The main product of enzymatic hydrolysis is SPH, while P4 can be considered the main product of UF when the goal is to concentrate small peptides. However, during both processing methods, the majority of the protein content ends up in fractions other than the main products. This highlights the importance of finding areas of application for the sludge fraction from enzymatic hydrolysis and the retentates from UF in order to make food production more sustainable by improving the utilization of saithe rest raw material.
Author Contributions
Conceptualization, V.H., T.R., I.-J.J. and E.F.; Data curation, V.H., E.E. and S.M.P.; Formal analysis, V.H., I.-J.J., E.E. and S.M.P.; Funding acquisition, E.F.; Investigation, V.H. and I.-J.J.; Methodology, V.H., T.R., I.-J.J. and E.F.; Project administration, T.R. and E.F.; Resources, V.H., E.E., S.M.P. and E.F.; Software, V.H.; Supervision, V.H., T.R., I.-J.J. and E.F.; Validation, V.H., T.R., I.-J.J. and E.F.; Visualization, V.H.; Writing—original draft, V.H.; Writing—review & editing, V.H., T.R., I.-J.J. and E.F. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Norwegian University of Science and Technology (NTNU) through the project OPTiMAT (Optimal Utilization of Marine Food Resources).
Data Availability Statement
The data presented in this study are available on request from the corresponding author. The data are not publicly available due to ethical restrictions.
Acknowledgments
The authors would like to acknowledge the flexibility and willingness to cooperate of the local fishers who provided the raw material for this study.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Ghaly, A.; Ramakrishnan, V.; Brooks, M.; Budge, S.; Dave, D. Fish Processing Wastes as a Potential Source of Proteins, Amino Acids and Oils: A Critical Review. J. Microb. Biochem. Technol. 2013, 5, 107–129. [Google Scholar] [CrossRef] [Green Version]
- Richardsen, R.; Myhre, M.; Nystøyl, R.; Strandheim, G.; Marthinussen, A. Analyse Marint Restråstoff 2018. Tilgang og Anvendelse av Marint Restråstoff i Norge; 2019:00475; SINTEF Ocean AS: Oslo, Norway, 2019; p. 47. [Google Scholar]
- Hjellnes, V.; Rustad, T.; Falch, E. Enzymatic hydrolysis of pre-spawned and spawning saithe (Pollachius virens) in bioreactors and its potential for implementation in the Norwegian whitefish industry. 2021; submitted for publication. [Google Scholar]
- Jafarpour, A.; Gomes, R.M.; Gregersen, S.; Sloth, J.J.; Jacobsen, C.; Sørensen, A.-D.M. Characterization of cod (Gadus morhua) frame composition and its valorization by enzymatic hydrolysis. J. Food Compos. Anal. 2020, 89, 103469. [Google Scholar] [CrossRef]
- Hou, H.; Li, B.; Zhao, X.; Zhang, Z.; Li, P. Optimization of enzymatic hydrolysis of Alaska pollock frame for preparing protein hydrolysates with low-bitterness. LWT 2011, 44, 421–428. [Google Scholar] [CrossRef]
- Remme, J.F.; Austnes, A. Kvalitetsproteiner fra Torskehoder—HEADS UP II; 2020:00322; SINTEF Ocean AS: Oslo, Norway, 2020; p. 21. [Google Scholar]
- Rustad, T. Utilisation of marine by-products. Elec. J. Env. Agricult. Food Chem. 2003, 2, 458–463. [Google Scholar]
- Hoegh-Guldberg, O. The Ocean as a Solution to Climate Change: Five Opportunities for Action; World Resource Institute: Washington, DC, USA, 2019. [Google Scholar]
- Kim, S.-K.; Mendis, E. Bioactive compounds from marine processing byproducts—A review. Food Res. Int. 2006, 39, 383–393. [Google Scholar] [CrossRef]
- Wijesekara, I.; Kim, S.-K. Angiotensin-I-Converting Enzyme (ACE) Inhibitors from Marine Resources: Prospects in the Pharmaceutical Industry. Mar. Drugs 2010, 8, 1080. [Google Scholar] [CrossRef] [Green Version]
- Guérard, F.; Sellos, D.; Le Gal, Y. Fish and Shellfish Upgrading, Traceability. In Marine Biotechnology; Ulber, R., Le Gal, Y., Eds.; Springer: Berlin/Heidelberg, Germany, 2005; pp. 127–163. [Google Scholar]
- Jensen, I.-J.; Mæhre, H.K. Preclinical and clinical studies on antioxidative, antihypertensive and cardioprotective effect of marine proteins and peptides—A review. Mar. Drugs 2016, 14, 211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dale, H.F.; Madsen, L.; Lied, G.A. Fish–derived proteins and their potential to improve human health. Nutr. Rev. 2019, 77, 572–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Girgih, A.T.; He, R.; Hasan, F.M.; Udenigwe, C.; Gill, T.A.; Aluko, R.E. Evaluation of the in vitro antioxidant properties of a cod (Gadus morhua) protein hydrolysate and peptide fractions. Food Chem. 2014, 173, 652–659. [Google Scholar] [CrossRef] [PubMed]
- Farvin, K.S.; Andersen, L.L.; Otte, J.; Nielsen, H.H.; Jessen, F.; Jacobsen, C. Antioxidant activity of cod (Gadus morhua) protein hydrolysates: Fractionation and characterisation of peptide fractions. Food Chem. 2016, 204, 409–419. [Google Scholar] [CrossRef] [Green Version]
- Picot, L.; Ravallec, R.; Fouchereau-Péron, M.; Vandanjon, L.; Jaouen, P.; Chaplain-Derouiniot, M.; Guérard, F.; Chabeaud, A.; LeGal, Y.; Alvarez, M.; et al. Impact of ultrafiltration and nanofiltration of an industrial fish protein hydrolysate on its bioactive properties. J. Sci. Food Agric. 2010, 90, 1819–1826. [Google Scholar] [CrossRef] [Green Version]
- Ngo, D.-H.; Vo, T.-S.; Ryu, B.; Kim, S.-K. Angiotensin- I- converting enzyme (ACE) inhibitory peptides from Pacific cod skin gelatin using ultrafiltration membranes. Process. Biochem. 2016, 51, 1622–1628. [Google Scholar] [CrossRef] [Green Version]
- Ween, O.; Stangeland, J.K.; Fylling, T.S.; Aas, G.H. Nutritional and functional properties of fishmeal produced from fresh by-products of cod (Gadus morhua L.) and saithe (Pollachius virens). Heliyon 2017, 3, e00343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jensen, I.-J.; Eysturskarð, J.; Madetoja, M.; Eilertsen, K.-E. The potential of cod hydrolyzate to inhibit blood pressure in spontaneously hypertensive rats. Nutr. Res. 2014, 34, 168–173. [Google Scholar] [CrossRef]
- Sarmadi, B.H.; Ismail, A. Antioxidative peptides from food proteins: A review. Peptides 2010, 31, 1949–1956. [Google Scholar] [CrossRef] [PubMed]
- Pihlanto, A. Antioxidative peptides derived from milk proteins. Int. Dairy J. 2006, 16, 1306–1314. [Google Scholar] [CrossRef]
- Gulcin, I. Antioxidants and antioxidant methods: An updated overview. Arch. Toxicol. 2020, 94, 651–715. [Google Scholar] [CrossRef] [Green Version]
- Branen, A.L. Toxicology and biochemistry of butylated hydroxyanisole and butylated hydroxytoluene. J. Am. Oil Chem. Soc. 1975, 52, 59–63. [Google Scholar] [CrossRef]
- Huang, D.; Ou, .A.B.; Prior, R.L. The Chemistry behind Antioxidant Capacity Assays. J. Agric. Food Chem. 2005, 53, 1841–1856. [Google Scholar] [CrossRef]
- FAO. The Future of Food and Agriculture—Trends And Challenges; FAO: Rome, Italy, 2017. [Google Scholar]
- Udenigwe, C.; Aluko, R.E. Food Protein-Derived Bioactive Peptides: Production, Processing, and Potential Health Benefits. J. Food Sci. 2011, 77, R11–R24. [Google Scholar] [CrossRef]
- Kaur, N.; Sharma, P.; Jaimni, S.; Kehinde, B.A.; Kaur, S. Recent developments in purification techniques and industrial applications for whey valorization: A review. Chem. Eng. Commun. 2019, 207, 123–138. [Google Scholar] [CrossRef]
- Gao, R.; Yu, Q.; Shen, Y.; Chu, Q.; Chen, G.; Fen, S.; Yang, M.; Yuan, L.; McClements, D.J.; Sun, Q. Production, bioactive properties, and potential applications of fish protein hydrolysates: Developments and challenges. Trends Food Sci. Technol. 2021, 110, 687–699. [Google Scholar] [CrossRef]
- Hjellnes, V.; Rustad, T.; Falch, E. The value chain of the white fish industry in Norway: History, current status and possibilities for improvement—A review. Reg. Stud. Mar. Sci. 2020, 36, 101293. [Google Scholar] [CrossRef]
- Bacchin, P.; Aimar, P.; Field, R. Critical and sustainable fluxes: Theory, experiments and applications. J. Membr. Sci. 2006, 281, 42–69. [Google Scholar] [CrossRef] [Green Version]
- Araujo, J.; Sica, P.; Costa, C.; Márquez, M.C. Enzymatic Hydrolysis of Fish Waste as an Alternative to Produce High Value-Added Products. Waste Biomass Valorization 2020, 12, 847–855. [Google Scholar] [CrossRef]
- Gildberg, A.; A Arnesen, J.; Carlehög, M. Utilisation of cod backbone by biochemical fractionation. Process. Biochem. 2002, 38, 475–480. [Google Scholar] [CrossRef]
- Karami, Z.; Akbari-Adergani, B. Bioactive food derived peptides: A review on correlation between structure of bioactive peptides and their functional properties. J. Food Sci. Technol. 2019, 56, 535–547. [Google Scholar] [CrossRef]
- Jensen, I.-J.; Larsen, R.; Rustad, T.; Eilertsen, K.-E. Nutritional content and bioactive properties of wild and farmed cod (Gadus morhua L.) subjected to food preparation. J. Food Compos. Anal. 2013, 31, 212–216. [Google Scholar] [CrossRef]
- Coultate, T.P. Food: The Chemistry of its Components; Royal Society of Chemistry: London, UK, 2009. [Google Scholar]
- Branden, C.I.; Tooze, J. The Building Blocks. In Introduction to Protein Structure, 12nd ed.; Garland Science: New York, NY, USA, 2012; pp. 1–12. [Google Scholar]
- Slizyte, R.; Daukšas, E.; Falch, E.; Storrø, I.; Rustad, T. Characteristics of protein fractions generated from hydrolysed cod (Gadus morhua) by-products. Process. Biochem. 2005, 40, 2021–2033. [Google Scholar] [CrossRef]
- HoleČek, M. Histidine in Health and Disease: Metabolism, Physiological Importance, and Use as a Supplement. Nutrients 2020, 12, 848. [Google Scholar] [CrossRef] [Green Version]
- Rajapakse, N.; Mendis, E.; Jung, W.-K.; Je, J.-Y.; Kim, S.-K. Purification of a radical scavenging peptide from fermented mussel sauce and its antioxidant properties. Food Res. Int. 2005, 38, 175–182. [Google Scholar] [CrossRef]
- Marcuse, R. The effect of some amino acids on the oxidation of linoleic acid and its methyl ester. J. Am. Oil Chem. Soc. 1962, 39, 97–103. [Google Scholar] [CrossRef]
- Chen, H.-M.; Muramoto, K.; Yamauchi, F.; Fujimoto, K.; Nokihara, K. Antioxidative Properties of Histidine-Containing Peptides Designed from Peptide Fragments Found in the Digests of a Soybean Protein. J. Agric. Food Chem. 1998, 46, 49–53. [Google Scholar] [CrossRef]
- Siemensma, A.D.; Weijer, W.J.; Bak, H.J. The importance of peptide lengths in hypoallergenic infant formulae. Trends Food Sci. Technol. 1993, 4, 16–21. [Google Scholar] [CrossRef]
- Chabeaud, A.; Vandanjon, L.; Bourseau, P.; Jaouen, P.; Chaplain-Derouiniot, M.; Guerard, F. Performances of ultrafiltration membranes for fractionating a fish protein hydrolysate: Application to the refining of bioactive peptidic fractions. Sep. Purif. Technol. 2009, 66, 463–471. [Google Scholar] [CrossRef]
- Chabeaud, A.; Vandanjon, L.; Bourseau, P.; Jaouen, P.; Guérard, F. Fractionation by ultrafiltration of a saithe protein hydrolysate (Pollachius virens): Effect of material and molecular weight cut-off on the membrane performances. J. Food Eng. 2009, 91, 408–414. [Google Scholar] [CrossRef]
- Damodaran, S.; Parkin, K.L.; Fennema, O.R. Amino acids, peptides, and proteins. In Fennema’s Food Chemistry, 4th ed.; Damodaran, S., Ed.; CRC Press: Boca Raton, FL, USA, 2008; pp. 217–331. [Google Scholar]
- Jeon, Y.-J.; Byun, H.-G.; Kim, S.-K. Improvement of functional properties of cod frame protein hydrolysates using ultrafiltration membranes. Process. Biochem. 1999, 35, 471–478. [Google Scholar] [CrossRef]
- Dávalos, A.; Gómez-Cordovés, A.C.; Bartolomé, B. Extending Applicability of the Oxygen Radical Absorbance Capacity (ORAC−Fluorescein) Assay. J. Agric. Food Chem. 2003, 52, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Saiga, A.; Tanabe, S.; Nishimura, T. Antioxidant activity of peptides obtained from porcine myofibrillar proteins by protease treatment. J. Agric. Food Chem. 2003, 51, 3661–3667. [Google Scholar] [CrossRef]
- Kristinová, V.; Mozuraityte, R.; Storrø, I.; Rustad, T. Antioxidant Activity of Phenolic Acids in Lipid Oxidation Catalyzed by Different Prooxidants. J. Agric. Food Chem. 2009, 57, 10377–10385. [Google Scholar] [CrossRef]
- AOAC. Association of Official Analytical Chemists. Official Methods of Analysis, 15th ed.; AOAC: Arlington, VA, USA; Washington, DC, USA, 1990. [Google Scholar]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free. Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Nenadis, N.; Wang, L.-F.; Tsimidou, M.; Zhang, H.-Y. Estimation of Scavenging Activity of Phenolic Compounds Using the ABTS•+Assay. J. Agric. Food Chem. 2004, 52, 4669–4674. [Google Scholar] [CrossRef] [PubMed]
- Nenadis, N.; Lazaridou, A.O.; Tsimidou, M. Use of Reference Compounds in Antioxidant Activity Assessment. J. Agric. Food Chem. 2007, 55, 5452–5460. [Google Scholar] [CrossRef]
- Benzie, I.; Strain, J. The Ferric Reducing Ability of Plasma (FRAP) as a Measure of “Antioxidant Power”: The FRAP Assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jensen, I.-J.; Abrahamsen, H.; Maehre, H.K.; Elvevoll, E.O. Changes in Antioxidative Capacity of Saithe (Pollachius virens) and Shrimp (Pandalus borealis) duringin VitroDigestion. J. Agric. Food Chem. 2009, 57, 10928–10932. [Google Scholar] [CrossRef]
- Blackburn, S. Amino Acid Determination. Methods and Techniques; Edwars Arnold Ltd.: London, UK, 1968; p. 271. [Google Scholar]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).