Modulation of Human Mesenchymal Stem Cells by Electrical Stimulation Using an Enzymatic Biofuel Cell

 , , ,
, , ,
Abstract
1. Introduction
2. Results
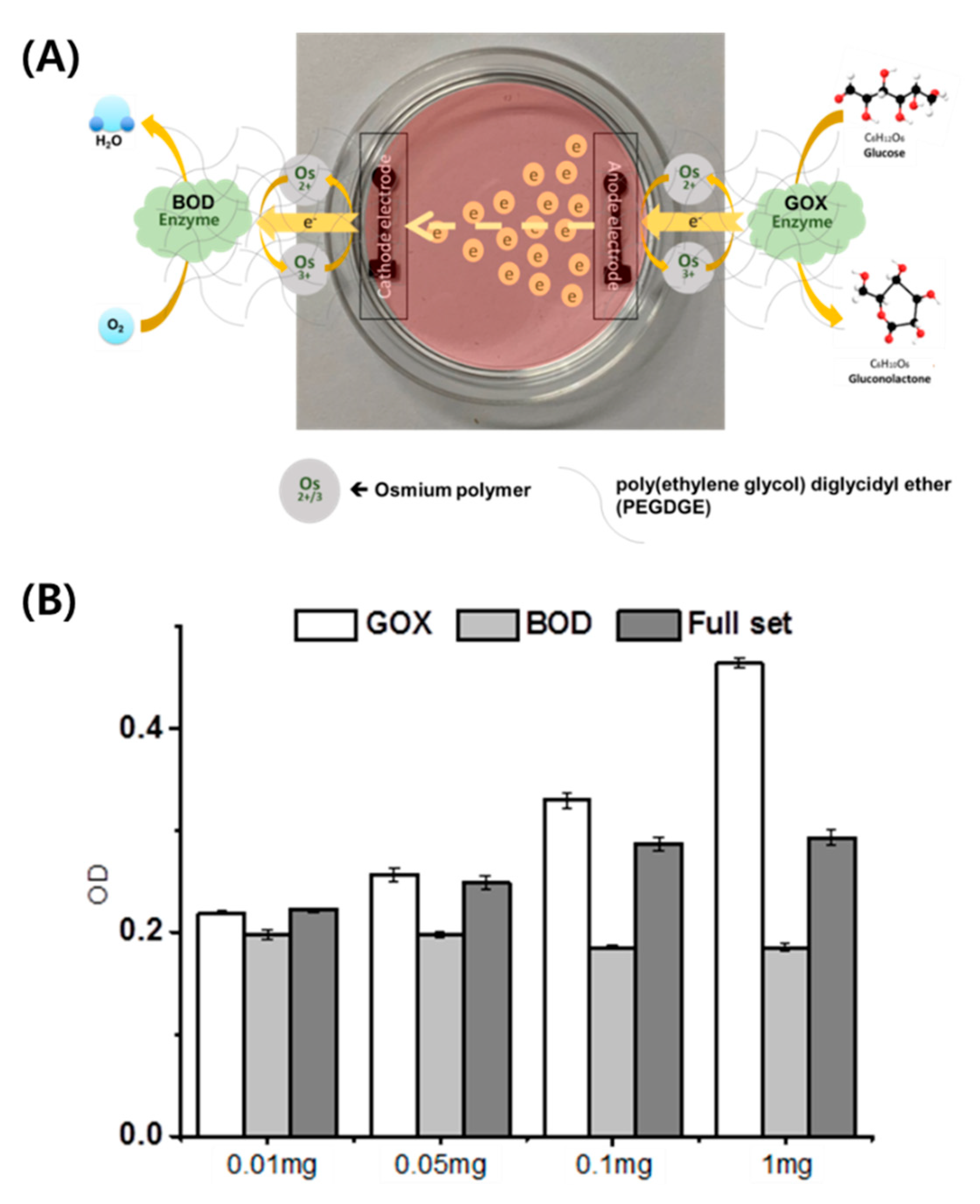
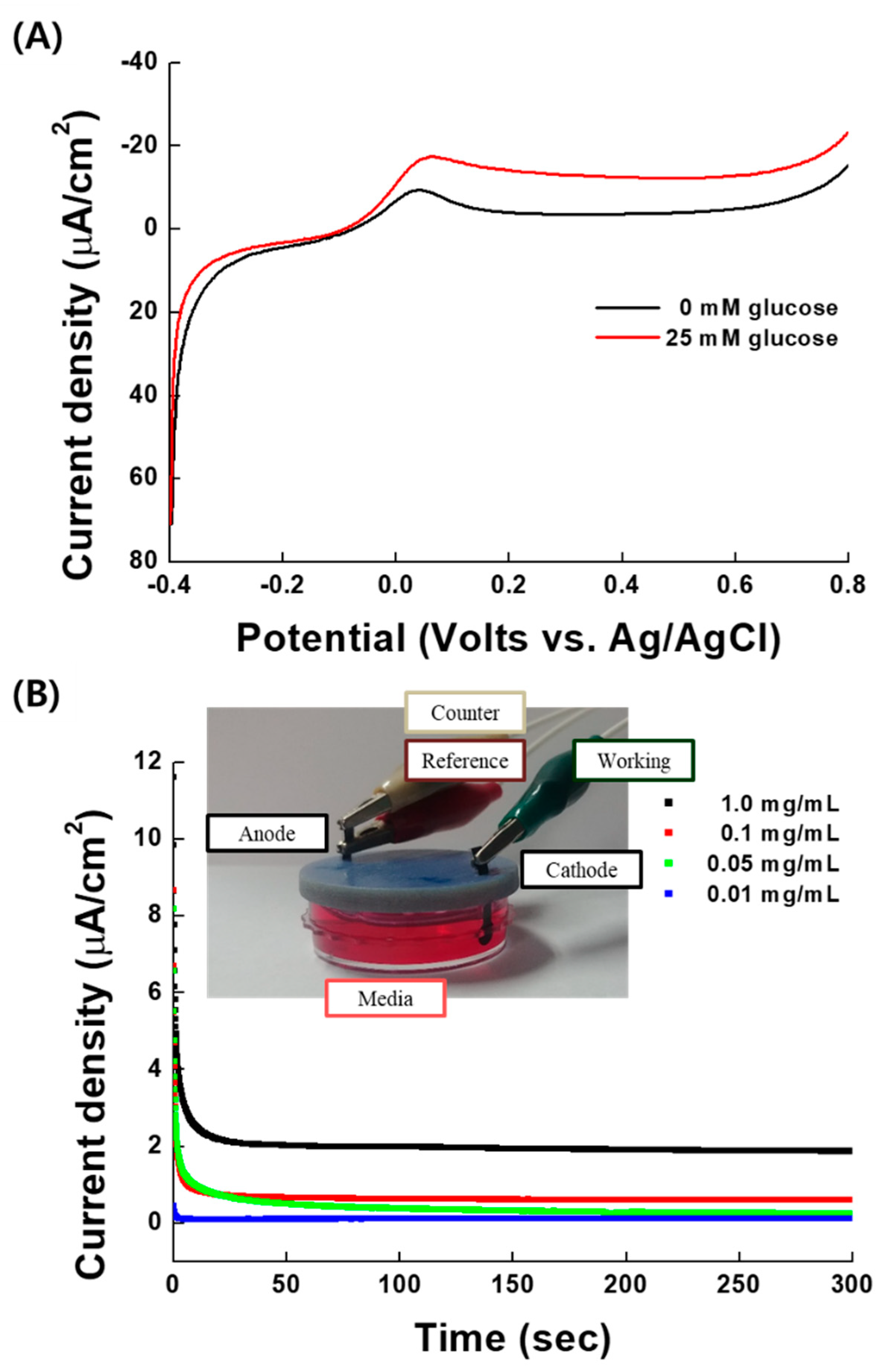
2.1. Characterization of EBFCs Used for Cellular Studies
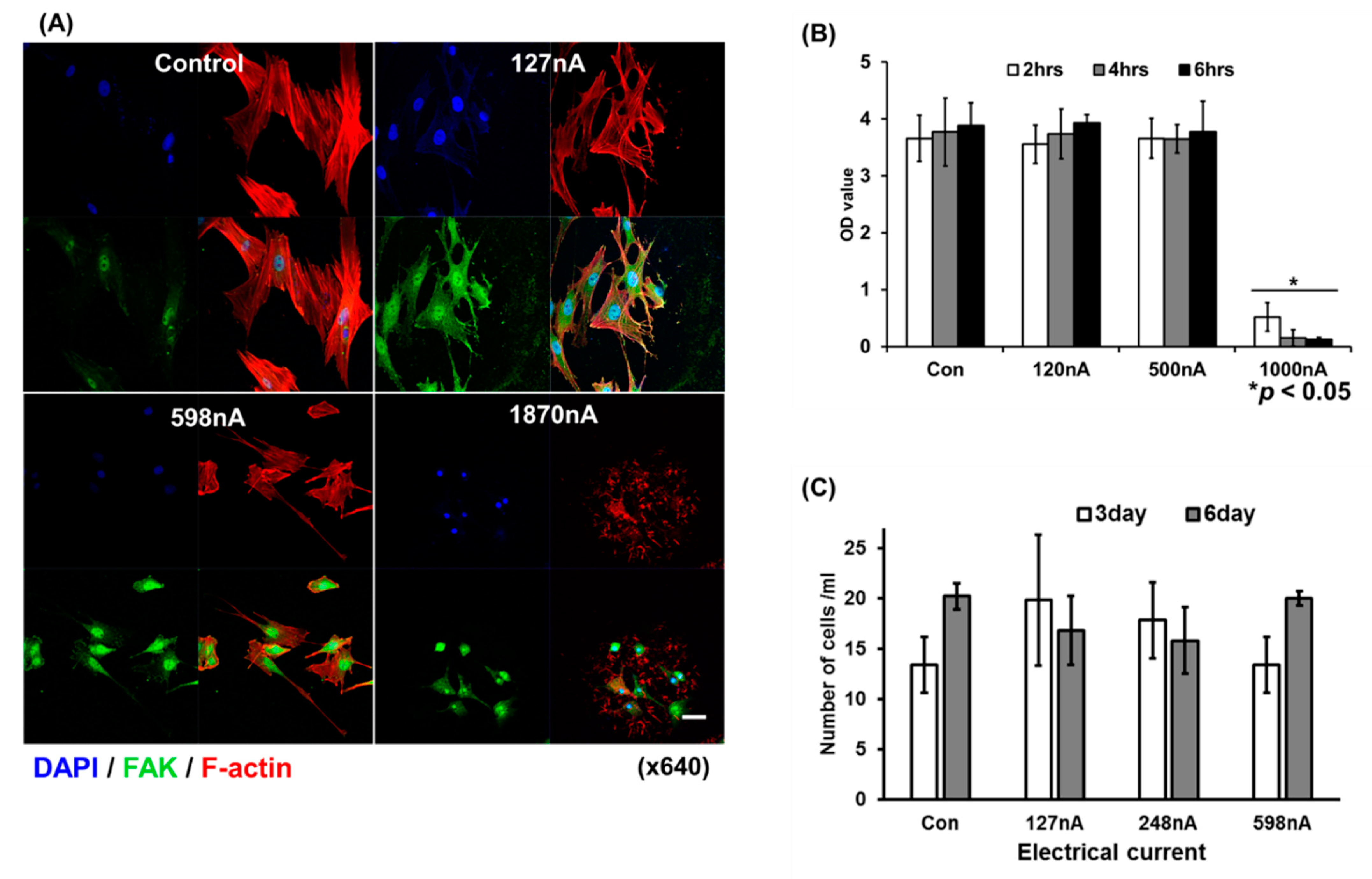
2.2. EBFC Electrical Current for Determining Cell Viability
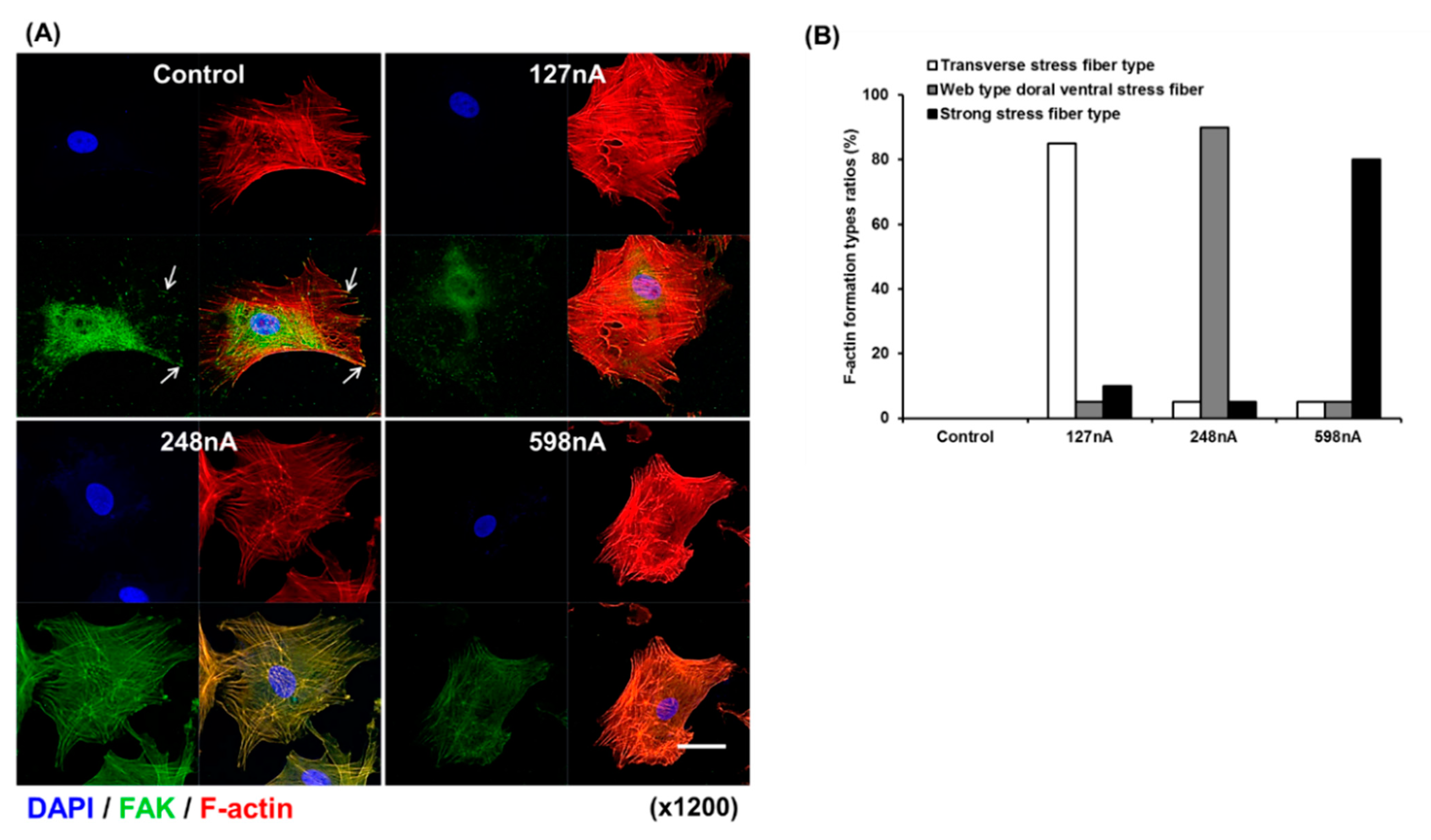
2.3. Morphological Analysis of Electrically Stimulated hAD-MSCs
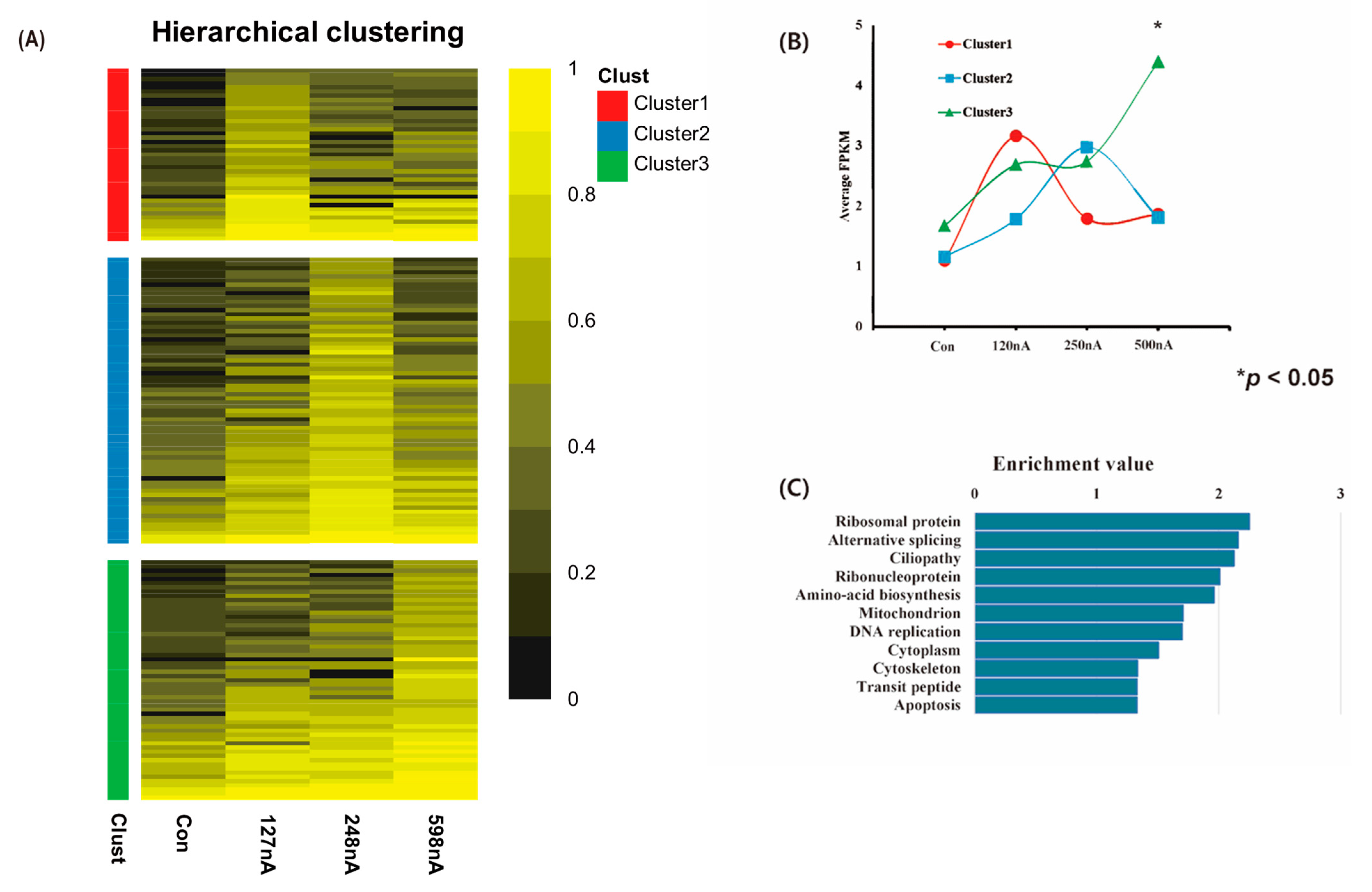
2.4. Transcriptomic Analysis of Electrically Stimulated hAD-MSCs
2.5. Transcriptome Visualization of Electrically Stimulated hAD-MSCs by Reduce Visualize Gene Ontology (REVIGO) Program
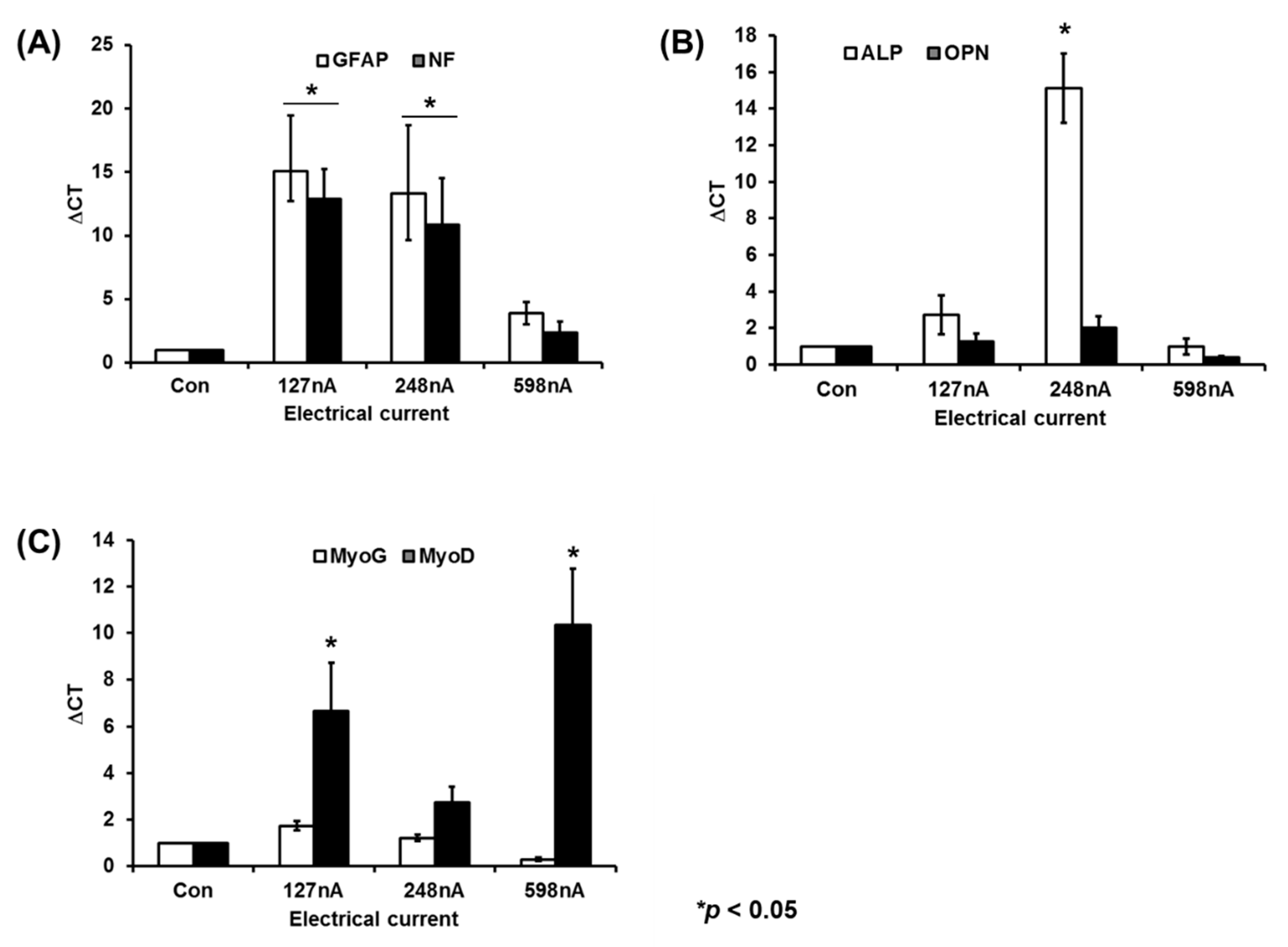
2.6. Early-Phase Differentiation-Related Gene Expression of Electrically Stimulated hAD-MSCs
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Preparation of Enzyme Electrodes
4.3. EBFC Set-Up for 2D Cell Culture
4.4. Measurements of Electrical Currents
4.5. Human Adipose-Derived Mesenchymal Stem Cell Preparation and Maintenance
4.6. Cell Survival, Proliferation and Cell Morphology under Electrical Stimulation
4.7. mRNA-Seq Analysis
4.8. Gene Ontology Analysis of Transcriptome Data
4.9. Reverse Transcription-qPCR Analysis
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Hickey, D.P.; Reid, R.C.; Milton, R.D.; Minteer, S.D. A self-powered amperometric lactate biosensor based on lactate oxidase immobilized in dimethylferrocene-modified LPEI. Biosens. Bioelectron. 2016, 77, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Jeon, W.Y.; Lee, J.H.; Dashnyam, K.; Choi, Y.B.; Kim, T.H.; Lee, H.H.; Kim, H.W.; Kim, H.H. Performance of a glucose-reactive enzyme-based biofuel cell system for biomedical applications. Sci. Rep. 2019, 9, 10872. [Google Scholar] [CrossRef]
- Kim, J.; Jeerapan, I.; Sempionatto, J.R.; Barfidokht, A.; Mishra, R.K.; Campbell, A.S.; Hubble, L.J.; Wang, J. Wearable Bioelectronics: Enzyme-Based Body-Worn Electronic Devices. Acc. Chem. Res. 2018, 51, 2820–2828. [Google Scholar] [CrossRef] [PubMed]
- Burdick, J.A.; Vunjak-Novakovic, G. Engineered microenvironments for controlled stem cell differentiation. Tissue Eng. Part A 2009, 15, 205–219. [Google Scholar] [CrossRef] [PubMed]
- Kondrashova, M.N. Control of mitochondria respiration during potentiating effects on the cell. Biofizika 1970, 15, 312–323. [Google Scholar] [PubMed]
- Levin, M. Bioelectric mechanisms in regeneration: Unique aspects and future perspectives. Semin. Cell Dev. Biol. 2009, 20, 543–556. [Google Scholar] [CrossRef] [PubMed]
- Levin, M.; Stevenson, C.G. Regulation of cell behavior and tissue patterning by bioelectrical signals: Challenges and opportunities for biomedical engineering. Annu. Rev. Biomed. Eng. 2012, 14, 295–323. [Google Scholar] [CrossRef] [PubMed]
- Peckham, P.H.; Knutson, J.S. Functional electrical stimulation for neuromuscular applications. Annu. Rev. Biomed. Eng. 2005, 7, 327–360. [Google Scholar] [CrossRef]
- Song, B.; Gu, Y.; Pu, J.; Reid, B.; Zhao, Z.; Zhao, M. Application of direct current electric fields to cells and tissues in vitro and modulation of wound electric field in vivo. Nat. Protoc. 2007, 2, 1479–1489. [Google Scholar] [CrossRef]
- Ud-Din, S.; Bayat, A. Electrical Stimulation and Cutaneous Wound Healing: A Review of Clinical Evidence. Healthcare 2014, 2, 445–467. [Google Scholar] [CrossRef] [PubMed]
- Chang, K.A.; Kim, J.W.; Kim, J.A.; Lee, S.E.; Kim, S.; Suh, W.H.; Kim, H.S.; Kwon, S.; Kim, S.J.; Suh, Y.H. Biphasic electrical currents stimulation promotes both proliferation and differentiation of fetal neural stem cells. PLoS ONE 2011, 6, e18738. [Google Scholar] [CrossRef]
- Du, J.; Zhen, G.; Chen, H.; Zhang, S.; Qing, L.; Yang, X.; Lee, G.; Mao, H.Q.; Jia, X. Optimal electrical stimulation boosts stem cell therapy in nerve regeneration. Biomaterials 2018, 181, 347–359. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Hu, B.; Li, Z.; Li, J.; Gao, Y.; Wang, Z.; Hao, J. Synergistic Effects of Electrical Stimulation and Aligned Nanofibrous Microenvironment on Growth Behavior of Mesenchymal Stem Cells. ACS Appl. Mater. Interfaces 2018, 10, 18543–18550. [Google Scholar] [CrossRef] [PubMed]
- Kwon, H.J.; Lee, G.S.; Chun, H. Electrical stimulation drives chondrogenesis of mesenchymal stem cells in the absence of exogenous growth factors. Sci. Rep. 2016, 6, 39302. [Google Scholar] [CrossRef] [PubMed]
- Mobini, S.; Leppik, L.; Barker, J.H. Direct current electrical stimulation chamber for treating cells in vitro. Biotechniques 2016, 60, 95–98. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.U.; Blasiak, A.; Agrawal, D.R.; Loong, D.T.B.; Thakor, N.V.; All, A.H.; Ho, J.S.; Yang, I.H. Subcellular electrical stimulation of neurons enhances the myelination of axons by oligodendrocytes. PLoS ONE 2017, 12, e0179642. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Grumbles, R.M.; Thomas, C.K. Electrical stimulation of embryonic neurons for 1 h improves axon regeneration and the number of reinnervated muscles that function. J. Neuropathol. Exp. Neurol. 2013, 72, 697–707. [Google Scholar] [CrossRef]
- Malyshevskaya, O.; Shiraishi, Y.; Kimura, F.; Yamamoto, N. Role of electrical activity in horizontal axon growth in the developing cortex: A time-lapse study using optogenetic stimulation. PLoS ONE 2013, 8, e82954. [Google Scholar] [CrossRef]
- Zhu, R.; Sun, Z.; Li, C.; Ramakrishna, S.; Chiu, K.; He, L. Electrical stimulation affects neural stem cell fate and function in vitro. Exp. Neurol. 2019, 319, 112963. [Google Scholar] [CrossRef]
- Jackson, A.; Zimmermann, J.B. Neural interfaces for the brain and spinal cord--restoring motor function. Nat. Rev. Neurol. 2012, 8, 690–699. [Google Scholar] [CrossRef]
- Jaermann, T.; Suter, F.; Osterwalder, D.; Luechinger, R. Measurement and analysis of electromagnetic fields of pulsed magnetic field therapy systems for private use. J. Radiol. Prot. 2011, 31, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Yahiro, A.T.; Lee, S.M.; Kimble, D.O. Bioelectrochemistry. I. Enzyme Utilizing Bio-Fuel Cell Studies. Biochim. Biophys. Acta 1964, 88, 375–383. [Google Scholar] [PubMed]
- Barton, S.C.; Gallaway, J.; Atanassov, P. Enzymatic biofuel cells for implantable and microscale devices. Chem. Rev. 2004, 104, 4867–4886. [Google Scholar] [CrossRef] [PubMed]
- Heller, A. Miniature biofuel cells. Phys. Chem. Chem. Phys. 2004, 6, 209. [Google Scholar] [CrossRef]
- Mano, N.; Mao, F.; Heller, A. Characteristics of a miniature compartment-less glucose-O2 biofuel cell and its operation in a living plant. J. Am. Chem. Soc. 2003, 125, 6588–6594. [Google Scholar] [CrossRef]
- Rapoport, B.I.; Kedzierski, J.T.; Sarpeshkar, R. A glucose fuel cell for implantable brain-machine interfaces. PLoS ONE 2012, 7, e38436. [Google Scholar] [CrossRef]
- Halamkova, L.; Halamek, J.; Bocharova, V.; Szczupak, A.; Alfonta, L.; Katz, E. Implanted biofuel cell operating in a living snail. J. Am. Chem. Soc. 2012, 134, 5040–5043. [Google Scholar] [CrossRef]
- Rasmussen, M.; Ritzmann, R.E.; Lee, I.; Pollack, A.J.; Scherson, D. An implantable biofuel cell for a live insect. J. Am. Chem. Soc. 2012, 134, 1458–1460. [Google Scholar] [CrossRef]
- Wen, D.; Eychmuller, A. Enzymatic Biofuel Cells on Porous Nanostructures. Small 2016, 12, 4649–4661. [Google Scholar] [CrossRef]
- Zebda, A.; Gondran, C.; Le Goff, A.; Holzinger, M.; Cinquin, P.; Cosnier, S. Mediatorless high-power glucose biofuel cells based on compressed carbon nanotube-enzyme electrodes. Nat. Commun. 2011, 2, 370. [Google Scholar] [CrossRef]
- Justin, G.A.; Zhang, Y.; Cui, X.T.; Bradberry, C.W.; Sun, M.; Sclabassi, R.J. A metabolic biofuel cell: Conversion of human leukocyte metabolic activity to electrical currents. J. Biol. Eng. 2011, 5, 5. [Google Scholar] [CrossRef][Green Version]
- Lee, K.D. Applications of mesenchymal stem cells: An updated review. Chang. Gung Med. J. 2008, 31, 228–236. [Google Scholar] [PubMed]
- Sigurjonsson, O.E.; Guethmundsson, K.O.; Guethmundsson, S. Mesenchymal stem cells. A review. Laeknabladid 2001, 87, 627–632. [Google Scholar] [PubMed]
- Guilak, F.; Cohen, D.M.; Estes, B.T.; Gimble, J.M.; Liedtke, W.; Chen, C.S. Control of stem cell fate by physical interactions with the extracellular matrix. Cell Stem Cell 2009, 5, 17–26. [Google Scholar] [CrossRef]
- Park, J.; Kim, P.; Helen, W.; Engler, A.J.; Levchenko, A.; Kim, D.H. Control of stem cell fate and function by engineering physical microenvironments. Integr. Biol. 2012, 4, 1008–1018. [Google Scholar]
- Zhang, B.; Fan, L.; Zhong, H.; Liu, Y.; Chen, S. Graphene nanoelectrodes: Fabrication and size-dependent electrochemistry. J. Am. Chem. Soc. 2013, 135, 10073–10080. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Jiao, Y. Next-generation sequencing applied to flower development: RNA-seq. Methods Mol. Biol. 2014, 1110, 401–411. [Google Scholar] [CrossRef] [PubMed]
- Banks, M.A.; Persily, G.L. Campus perspective on the National Institutes of Health public access policy: University of California, San Francisco, library experience. J. Med. Libr. Assoc. 2010, 98, 256–259. [Google Scholar] [CrossRef]
- Zhang, J.; Jiao, J. Molecular Biomarkers for Embryonic and Adult Neural Stem Cell and Neurogenesis. Biomed. Res. Int. 2015, 2015, 727542. [Google Scholar] [CrossRef]
- Hill, C.B.; Cassin, A.; Keeble-Gagnere, G.; Doblin, M.S.; Bacic, A.; Roessner, U. De novo transcriptome assembly and analysis of differentially expressed genes of two barley genotypes reveal root-zone-specific responses to salt exposure. Sci. Rep. 2016, 6, 31558. [Google Scholar] [CrossRef]
- Uzer, G.; Fuchs, R.K.; Rubin, J.; Thompson, W.R. Concise Review: Plasma and Nuclear Membranes Convey Mechanical Information to Regulate Mesenchymal Stem Cell Lineage. Stem Cells 2016, 34, 1455–1463. [Google Scholar] [CrossRef] [PubMed]
- Kleinschmidt, E.G.; Schlaepfer, D.D. Focal adhesion kinase signaling in unexpected places. Curr. Opin. Cell Biol. 2017, 45, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Visavadiya, N.P.; Keasey, M.P.; Razskazovskiy, V.; Banerjee, K.; Jia, C.; Lovins, C.; Wright, G.L.; Hagg, T. Integrin-FAK signaling rapidly and potently promotes mitochondrial function through STAT3. Cell Commun. Signal. 2016, 14, 32. [Google Scholar] [CrossRef] [PubMed]
- Schlaepfer, D.D.; Hauck, C.R.; Sieg, D.J. Signaling through focal adhesion kinase. Prog. Biophys. Mol. Biol. 1999, 71, 435–478. [Google Scholar] [CrossRef]
- Moujaber, O.; Stochaj, U. The Cytoskeleton as Regulator of Cell Signaling Pathways. Trends Biochem. Sci. 2020, 45, 96–107. [Google Scholar] [CrossRef] [PubMed]
- Wozniak, M.A.; Modzelewska, K.; Kwong, L.; Keely, P.J. Focal adhesion regulation of cell behavior. Biochim. Biophys. Acta 2004, 1692, 103–119. [Google Scholar] [CrossRef] [PubMed]
- Alenghat, F.J.; Ingber, D.E. Mechanotransduction: All signals point to cytoskeleton, matrix, and integrins. Sci. STKE 2002, 2002, pe6. [Google Scholar] [CrossRef]
- Lee, F.Y.; Zhen, Y.Y.; Yuen, C.M.; Fan, R.; Chen, Y.T.; Sheu, J.J.; Chen, Y.L.; Wang, C.J.; Sun, C.K.; Yip, H.K. The mTOR-FAK mechanotransduction signaling axis for focal adhesion maturation and cell proliferation. Am. J. Transl. Res. 2017, 9, 1603–1617. [Google Scholar]
- Leucht, P.; Kim, J.B.; Currey, J.A.; Brunski, J.; Helms, J.A. FAK-Mediated mechanotransduction in skeletal regeneration. PLoS ONE 2007, 2, e390. [Google Scholar] [CrossRef]
- Shafrir, Y.; Forgacs, G. Mechanotransduction through the cytoskeleton. Am. J. Physiol. Cell Physiol. 2002, 282, C479–C486. [Google Scholar] [CrossRef]
- Shao, Y.; Mann, J.M.; Chen, W.; Fu, J. Global architecture of the F-actin cytoskeleton regulates cell shape-dependent endothelial mechanotransduction. Integr. Biol. 2014, 6, 300–311. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.; Chen, Q.; Zheng, Y.; Nan, L.; Liao, N.; Mo, S. Potential Role of Integrin alpha(5)beta(1)/Focal Adhesion Kinase (FAK) and Actin Cytoskeleton in the Mechanotransduction and Response of Human Gingival Fibroblasts Cultured on a 3-Dimension Lactide-Co-Glycolide (3D PLGA) Scaffold. Med. Sci. Monit. 2020, 26, e921626. [Google Scholar] [CrossRef] [PubMed]
- Engler, A.J.; Sen, S.; Sweeney, H.L.; Discher, D.E. Matrix elasticity directs stem cell lineage specification. Cell 2006, 126, 677–689. [Google Scholar] [CrossRef] [PubMed]
- Zemel, A.; Rehfeldt, F.; Brown, A.E.; Discher, D.E.; Safran, S.A. Cell shape, spreading symmetry and the polarization of stress-fibers in cells. J. Phys. Condens. Matter. 2010, 22, 194110. [Google Scholar] [CrossRef] [PubMed]
- Denis-Donini, S.; Caprini, A.; Frassoni, C.; Grilli, M. Members of the NF-kappaB family expressed in zones of active neurogenesis in the postnatal and adult mouse brain. Brain Res. Dev. Brain Res. 2005, 154, 81–89. [Google Scholar] [CrossRef]
- Furst, D.O.; Osborn, M.; Weber, K. Myogenesis in the mouse embryo: Differential onset of expression of myogenic proteins and the involvement of titin in myofibril assembly. J. Cell Biol. 1989, 109, 517–527. [Google Scholar] [CrossRef]
- Rutkovskiy, A.; Stenslokken, K.O.; Vaage, I.J. Osteoblast Differentiation at a Glance. Med. Sci. Monit. Basic Res. 2016, 22, 95–106. [Google Scholar] [CrossRef]
- Zhou, X.; Liao, W.J.; Liao, J.M.; Liao, P.; Lu, H. Ribosomal proteins: Functions beyond the ribosome. J. Mol. Cell Biol. 2015, 7, 92–104. [Google Scholar] [CrossRef]
- Atanasova, M.; Whitty, A. Understanding cytokine and growth factor receptor activation mechanisms. Crit. Rev. Biochem. Mol. Biol. 2012, 47, 502–530. [Google Scholar] [CrossRef]
- Sanders, J.M.; Wampole, M.E.; Thakur, M.L.; Wickstrom, E. Molecular determinants of epidermal growth factor binding: A molecular dynamics study. PLoS ONE 2013, 8, e54136. [Google Scholar] [CrossRef]
- Bentzinger, C.F.; Wang, Y.X.; Rudnicki, M.A. Building muscle: Molecular regulation of myogenesis. Cold Spring Harb. Perspect. Biol. 2012, 4. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, Y.; Kato, K.; Miyake, T.; Nagamine, K.; Ofuji, T.; Yoshino, S.; Nishizawa, M. Organic transdermal iontophoresis patch with built-in biofuel cell. Adv. Healthc. Mater. 2015, 4, 506–510. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.Y.; Yun, J.H.; Park, Y.B.; Hyeon, J.S.; Jang, Y.; Choi, Y.B.; Kim, H.H.; Kang, T.M.; Ovalle, R.; Baughman, R.H.; et al. Two-Ply Carbon Nanotube Fiber-Typed Enzymatic Biofuel Cell Implanted in Mice. IEEE Trans. Nanobiosci. 2020, 19, 333–338. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Condition | Anode Electrodes GOx (Cathode Electrode; BOD Fixed to 0.1 mg/mL) | |||
|---|---|---|---|---|
| Enzyme concentration | 0.01 mg/mL | 0.05 mg/mL | 0.1 mg/mL | 1.0 mg/mL |
| IT current (nA/cm2) of the full system | 127 ± 9 | 248 ± 15 | 598 ± 75 | 1870 ± 305 |
| Forward Primer | Reverse Primer | |
|---|---|---|
| GFAP | 5′-GGCGCTCAATGCTGGCTTCA-3′ | 5′-TCTGCCTCCAGCCTCAGGTT-3′ |
| NF | 5′-TCAAGGCTAAGACCCTGGAG-3 | 5-AGGCCATCTTGACATTGAGG-3 |
| ALP | 5′-CCTCGTTGACACCTGGAAGAG-3′ | 5′-TTCCGTGCGGTTCCAGA-3′ |
| OPN | 5-ATCGATGTCCTTACTG-3 | 5-GGGATACTGTTCATCAGAAA-3 |
| MyoG | 5-GGATATGTCTGTTGCCTTC-3′ | 5-TGGGTGTTAGCCTTATGT-3 |
| MyoD | 5-GGAGTGGCAGAAAGTTAAG-3 | 5-ACGGGTCATCATAGAAGTC-3 |
| beta-actin | 5′-GCACCACACCTTCTACAATG-3′ | 5′-TGCTTGCTGATCCACATCTG-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeon, W.-Y.; Mun, S.; Ng, W.B.; Kang, K.; Han, K.; Hwang, S.; Kim, H.-H.; Lee, J.H. Modulation of Human Mesenchymal Stem Cells by Electrical Stimulation Using an Enzymatic Biofuel Cell. Catalysts 2021, 11, 62. https://doi.org/10.3390/catal11010062
Jeon W-Y, Mun S, Ng WB, Kang K, Han K, Hwang S, Kim H-H, Lee JH. Modulation of Human Mesenchymal Stem Cells by Electrical Stimulation Using an Enzymatic Biofuel Cell. Catalysts. 2021; 11(1):62. https://doi.org/10.3390/catal11010062
Chicago/Turabian StyleJeon, Won-Yong, Seyoung Mun, Wei Beng Ng, Keunsoo Kang, Kyudong Han, Sohyun Hwang, Hyug-Han Kim, and Jae Ho Lee. 2021. "Modulation of Human Mesenchymal Stem Cells by Electrical Stimulation Using an Enzymatic Biofuel Cell" Catalysts 11, no. 1: 62. https://doi.org/10.3390/catal11010062
APA StyleJeon, W.-Y., Mun, S., Ng, W. B., Kang, K., Han, K., Hwang, S., Kim, H.-H., & Lee, J. H. (2021). Modulation of Human Mesenchymal Stem Cells by Electrical Stimulation Using an Enzymatic Biofuel Cell. Catalysts, 11(1), 62. https://doi.org/10.3390/catal11010062