Whole-Cells of Yarrowia lipolytica Applied in “One Pot” Indolizine Biosynthesis

 ,
,  ,
,  ,
, 

Abstract
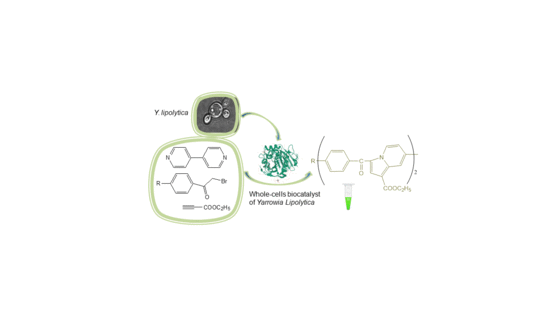
1. Introduction
2. Results and Discussion
2.1. Evaluation of the Toxicity of Reactants on the Yeasts’ Physiological Activity
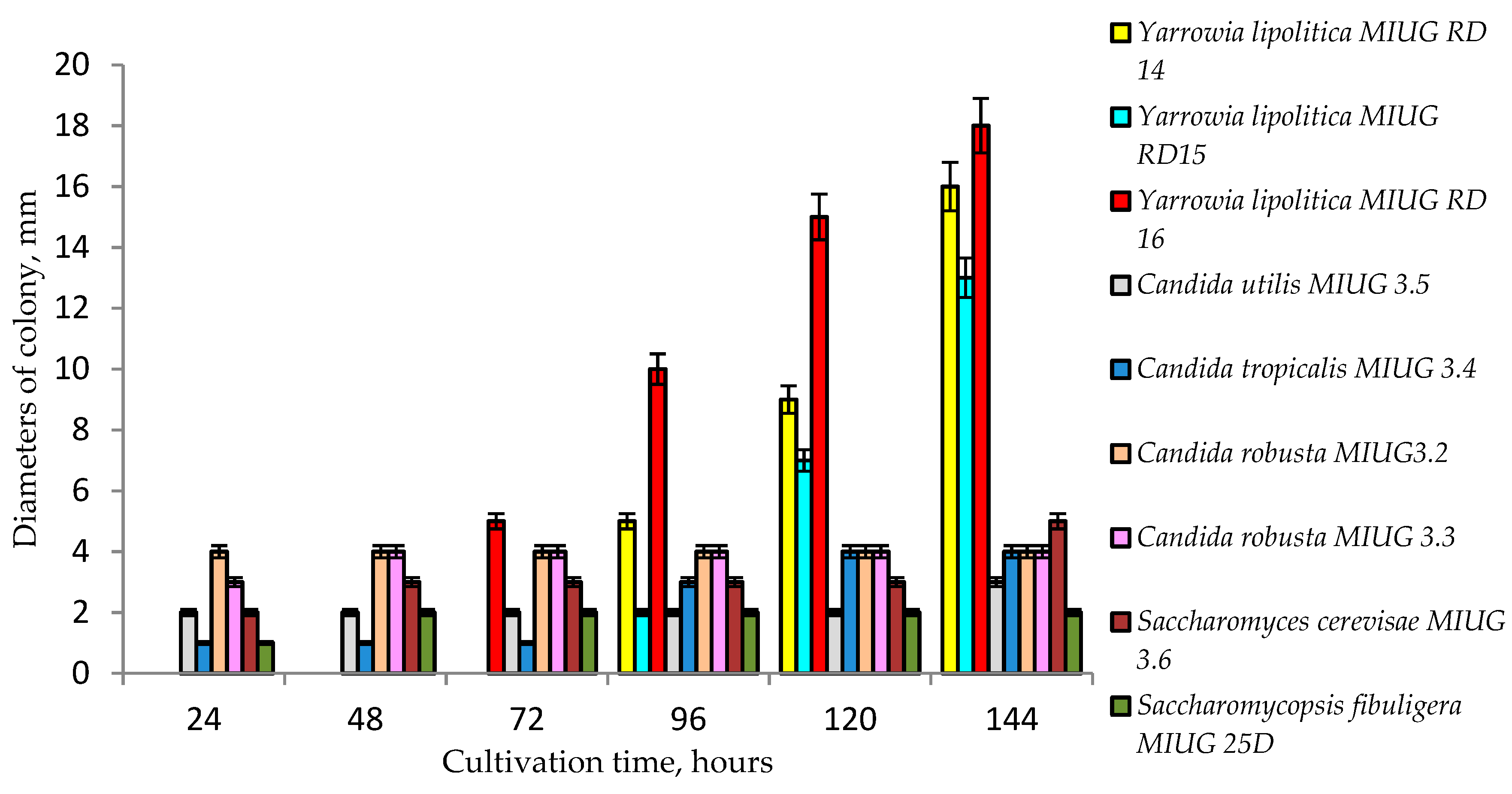
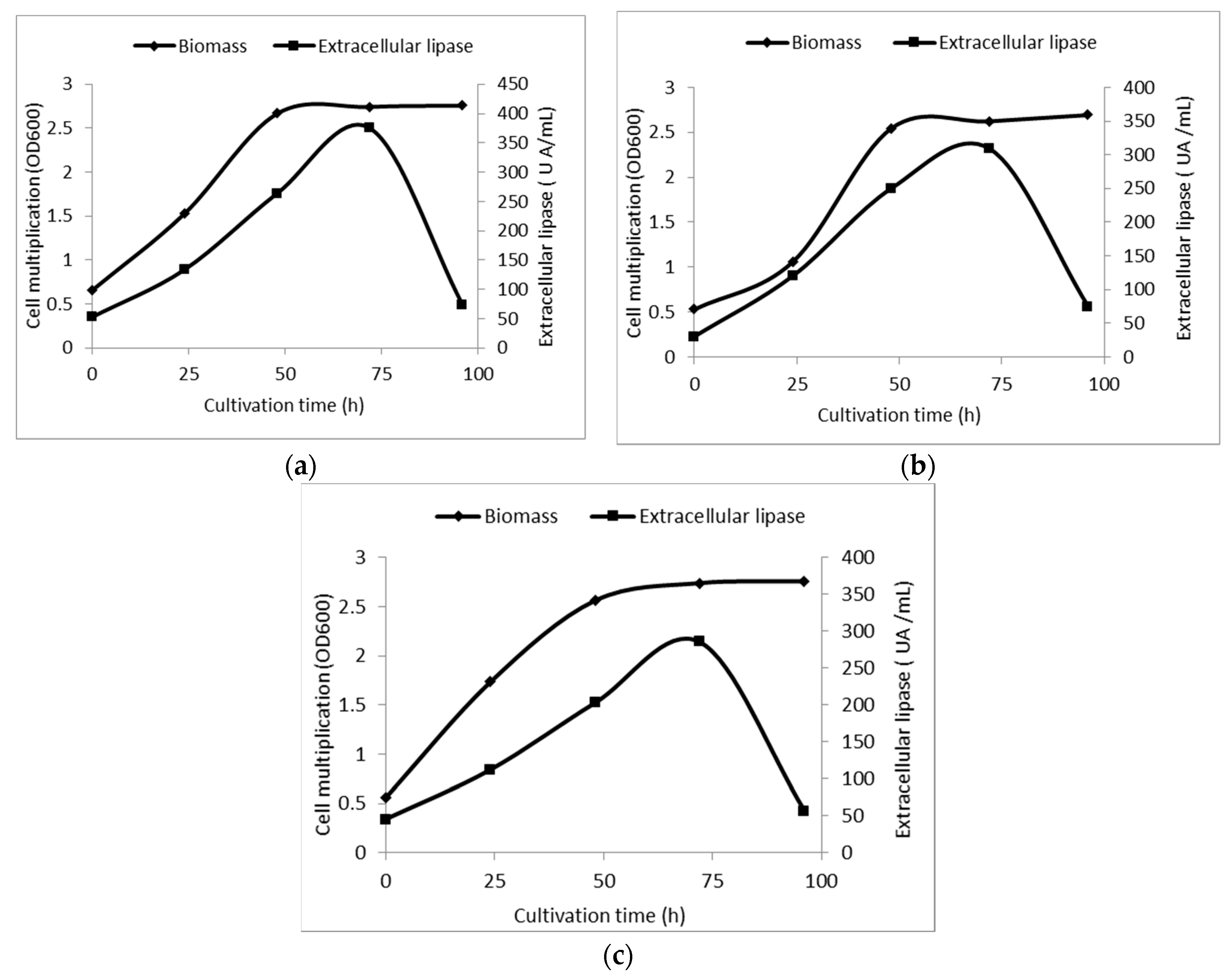
2.2. Growth and Bioconversion Potential of Yarrowia lipolytica in Aerobe Submerged Cultivation Conditions in the Presence of Chemicals
2.3. Indolizine Synthesis by Bioconversion
2.4. Evaluation of the Toxicity of Indolizines on the Germination of Triticum estivum L.
3. Materials and Methods
3.1. Microorganisms and Chemicals
3.2. Cultivation Conditions
3.3. Evaluation of the Toxicity of the Reactants on the Microorganisms’ Physiological Functionality
3.4. Yeast Multiplication by Submerged Cultivation in the Presence of Chemicals
3.5. Lipase Assay
3.6. Preparation of Crude Biocatalysts for Cycloaddition Reactions
3.7. Indolizine Bioformation Procedure
3.8. Separation and Purification of the Indolizines Bioconversion Products
3.9. Evaluation of the Toxicity of Indolizines on the Germination of Triticum azestivum L.
3.10. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zhao, Q.; Ansorge-Schumacher, M.B.; Haag, R.; Wu, C. Living whole-cell catalysis in compartmentalized emulsion. Bioresour. Technol. 2020, 295, 122221. [Google Scholar] [CrossRef]
- Feng, G.; Wu, H.; Li, X.; Lai, F.; Zhao, G.; Yang, M.; Liu, H. A new, efficient and highly-regioselective approach to synthesis of 6-O-propionyl-d-glucose by using whole-cell biocatalysts. Biochem. Eng. J. 2015, 95, 56–61. [Google Scholar] [CrossRef]
- Anteneh, Y.S.; Franco, C.M.M. Whole cell actinobacteria as biocatalysts. Front. Microbiol. 2019, 10, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Garzón-Posse, F.; Becerra-Figueroa, L.; Hernández-Arias, J.; Gamba-Sánchez, D. Whole cells as biocatalysts in organic transformations. Molecules 2018, 23, 1265. [Google Scholar] [CrossRef]
- Dinica, R.M.; Furdui, B.; Ghinea, I.O.; Bahrim, G.; Bonte, S.; Demeunynck, M. Novel one-pot green synthesis of indolizines biocatalysed by Candida antarctica lipases. Mar. Drugs 2013, 11, 431–439. [Google Scholar] [CrossRef]
- Matsuda, T.; Yamanaka, R.; Nakamura, K. Recent progress in biocatalysis for asymmetric oxidation and reduction. Tetrahedron Asymmetry 2009, 20, 513–557. [Google Scholar] [CrossRef]
- Kudalkar, G.P.; Tiwari, V.K.; Lee, J.D.; Berkowitz, D.B. A Hammett Study of Clostridium acetobutylicum Alcohol Dehydrogenase (CaADH): An enzyme with remarkable substrate promiscuity and utility for organic synthesis. Synlett 2020, 31, 237–247. [Google Scholar] [CrossRef]
- Antonopoulou, I.; Dilokpimol, A.; Iancu, L.; Mäkelä, M.R.; Varriale, S.; Cerullo, G.; Hüttner, S.; Uthoff, S.; Jütten, P.; Piechot, A.; et al. The synthetic potential of fungal feruloyl esterases: A correlation with current classification systems and predicted structural properties. Catalysts 2018, 8, 242. [Google Scholar] [CrossRef]
- Yin, D.H.; Liu, W.; Wang, Z.X.; Huang, X.; Zhang, J.; Huang, D.C. Enzyme-catalyzed direct three-component aza-Diels–Alder reaction using lipase from Candida sp. 99–125. Chin. Chem. Lett. 2017, 28, 153–158. [Google Scholar] [CrossRef]
- Carballeira, J.D.; Quezada, M.A.; Hoyos, P.; Simeó, Y.; Hernaiz, M.J.; Alcantara, A.R.; Sinisterra, J.V. Microbial cells as catalysts for stereoselective red-ox reactions. Biotechnol. Adv. 2009, 27, 686–714. [Google Scholar] [CrossRef]
- Ratnayake, N.D.; Theisen, C.; Walter, T.; Walker, K.D. Whole-cell biocatalytic production of variously substituted β-aryl- and β-heteroaryl-β-amino acids. J. Biotechnol. 2016, 217, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Birolli, W.G.; Ferreira, I.M.; Alvarenga, N.; De Santos, D.A.; De Matos, I.L.; Comasseto, J.V.; Porto, A.L.M. Biocatalysis and biotransformation in Brazil: An overview. Biotechnol. Adv. 2015, 33, 481–510. [Google Scholar] [CrossRef]
- Wachtmeister, J.; Rother, D. Recent advances in whole cell biocatalysis techniques bridging from investigative to industrial scale. Curr. Opin. Biotechnol. 2016, 42, 169–177. [Google Scholar] [CrossRef] [PubMed]
- Rywińska, A.; Juszczyk, P.; Wojtatowicz, M.; Robak, M.; Lazar, Z.; Tomaszewska, L.; Rymowicz, W. Glycerol as a promising substrate for Yarrowia lipolytica biotechnological applications. Biomass Bioenergy 2013, 48, 148–166. [Google Scholar] [CrossRef]
- Zieniuk, B.; Fabiszewska, A. Yarrowia lipolytica: A beneficious yeast in biotechnology as a rare opportunistic fungal pathogen: A minireview. World J. Microbiol. Biotechnol. 2019, 35, 1–8. [Google Scholar] [CrossRef]
- Fickers, P.; Benetti, P.H.; Waché, Y.; Marty, A.; Mauersberger, S.; Smit, M.S.; Nicaud, J.M. Hydrophobic substrate utilisation by the yeast Yarrowia lipolytica, and its potential applications. FEMS Yeast Res. 2005, 5, 527–543. [Google Scholar] [CrossRef]
- Gonçalves, F.A.G.; Colen, G.; Takahashi, J.A. Yarrowia lipolytica and its multiple applications in the biotechnological industry. Sci. World J. 2014, 2014, 1–14. [Google Scholar] [CrossRef]
- Casas-Godoy, L.; Marty, A.; Sandoval, G.; Ferreira-Dias, S. Optimization of medium chain length fatty acid incorporation into olive oil catalyzed by immobilized Lip2 from Yarrowia lipolytica. Biochem. Eng. J. 2013, 77, 20–27. [Google Scholar] [CrossRef]
- Facin, B.R.; Melchiors, M.S.; Valério, A.; Oliveira, J.V.; De Oliveira, D. Driving immobilized lipases as biocatalysts: 10 years state of the art and future prospects. Ind. Eng. Chem. Res. 2019, 58, 5358–5378. [Google Scholar] [CrossRef]
- Arvin-Berod, M.; Desroches-Castan, A.; Bonte, S.; Brugière, S.; Couté, Y.; Guyon, L.; Feige, J.J.; Baussanne, I.; Demeunynck, M. Indolizine-based scaffolds as efficient and versatile tools: Application to the synthesis of biotin-tagged antiangiogenic drugs. ACS Omega 2017, 2, 9221–9230. [Google Scholar] [CrossRef]
- Sharma, V.; Kumar, V. Indolizine: A biologically active moiety. Med. Chem. Res. 2014, 23, 3593–3606. [Google Scholar] [CrossRef]
- Narayanaswamy, K.; Id, V.; Chandrashekharappa, S.; Pillay, M.; Abdallah, H.H.; Mahomoodally, F.M.; Bhandary, S.; Chopra, D.; Attimarad, M.; Aldhubiab, B.E.; et al. Computational, crystallographic studies, cytotoxicity and anti-tubercular activity of substituted 7-methoxy-indolizine analogues. PLoS ONE 2019, 14, e0217270. [Google Scholar] [CrossRef]
- Venugopala, K.N.; Tratrat, C.; Chandrashekharappa, S. Anti-tubercular potency and computationally assessed drug-likeness and toxicology of diversely substituted indolizines. Indian J. Pharm. Educ. 2019, 53. [Google Scholar] [CrossRef]
- Uppar, V.; Chandrashekharappa, S.; Basarikatti, A.I.; Banuprakash, G.; Mahendra, K.; Chougala, M.; Mudnakudu-nagaraju, K.K.; Ningegowda, R.; Padmashali, B. Synthesis, antibacterial, and antioxidant studies of 7-amino-3-(4-fluorobenzoyl)indolizine-1-carboxylate derivatives. J. Appl. Pharm. Sci. 2020, 10, 77–85. [Google Scholar]
- Narajji, C.; Karvekar, M.D.; Das, A.K. Synthesis and antioxidant activity of 3,3'-diselanediylbis (N,N-disubstituted indolizine-1-carboxamide) and derivatives. S. Afr. J. Chem. 2008, 61, 53–55. [Google Scholar]
- Moon, S.H.; Jung, Y.; Kim, S.H.; Kim, I. Synthesis, characterization and biological evaluation of anti-cancer indolizine derivatives via inhibiting β-catenin activity and activating p53. Bioorganic Med. Chem. Lett. 2016, 26, 110–113. [Google Scholar] [CrossRef]
- Belal, A.; Gouda, A.M.; Ahmed, A.S.; Abdel Gawad, N.M. Synthesis of novel indolizine, diazepinoindolizine and Pyrimidoindolizine derivatives as potent and selective anticancer agents. Res. Chem. Intermed. 2015, 41, 9687–9701. [Google Scholar] [CrossRef]
- Ghinet, A.; Abuhaie, C.M.; Gautret, P.; Rigo, B.; Dubois, J.; Farce, A.; Belei, D.; Bîcu, E. Studies on indolizines. Evaluation of their biological properties as microtubule-interacting agents and as melanoma targeting compounds. Eur. J. Med. Chem. 2015, 89, 115–127. [Google Scholar] [CrossRef]
- Weide, T.; Arve, L.; Prinz, H.; Waldmann, H.; Kessler, H. 3-Substituted indolizine-1-carbonitrile derivatives as phosphatase inhibitors. Bioorganic Med. Chem. Lett. 2006, 16, 59–63. [Google Scholar] [CrossRef]
- Sharma, V.; Kumar, V. Pharmacophore mapping studies on indolizine derivatives as 15-LOX inhibitors. Bull. Fac. Pharm. Cairo Univ. 2015, 53, 63–68. [Google Scholar] [CrossRef]
- Huang, W.; Zuo, T.; Luo, X.; Jin, H.; Liu, Z.; Yang, Z.; Yu, X.; Zhang, L.; Zhang, L. Indolizine derivatives as HIV-1 VIF-elonginc interaction inhibitors. Chem. Biol. Drug Des. 2013, 81, 730–741. [Google Scholar] [CrossRef]
- De Souza, C.R.; Gonçalves, A.C.; Amaral, M.F.Z.J.; Dos Santos, A.A.; Clososki, G.C. Recent synthetic developments and reactivity of aromatic indolizines. Targets Heterocycl. Syst. 2016, 20, 365–392. [Google Scholar]
- Sadowski, B.; Klajn, J.; Gryko, D.T. Recent advances in the synthesis of indolizines and their π-expanded analogues. Org. Biomol. Chem. 2016, 14, 7804–7828. [Google Scholar] [CrossRef]
- Ghinea, I.O.; Dinica, R.M. Breakthroughs in indole and indolizine chemistry-new synthetic pathways, new applications. In Scope of Selective Heterocycles from Organic and Pharmaceutical Perspective; Chapter 5; Ravi, V., Ed.; IntechOpen: London, UK, 2016; pp. 115–142. [Google Scholar] [CrossRef]
- Kapoor, M.; Gupta, M.N. Lipase promiscuity and its biochemical applications. Process. Biochem. 2012, 47, 555–569. [Google Scholar] [CrossRef]
- Majumder, A.B.; Ramesh, N.G.; Gupta, M.N. A lipase catalyzed condensation reaction with a tricyclic diketone: Yet another example of biocatalytic promiscuity. Tetrahedron Lett. 2009, 50, 5190–5193. [Google Scholar] [CrossRef]
- Reetz, M.T.; Mondière, R.; Carballeira, J.D. Enzyme promiscuity: First protein-catalyzed Morita-Baylis-Hillman reaction. Tetrahedron Lett. 2007, 48, 1679–1681. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, C.Y.; Wang, H.R.; Zhang, H.; Su, Y.L.; Ji, T.F.; Wang, L. Lipase-catalyzed Knoevenagel condensation between α,β unsaturated aldehydes and active methylene compounds. Chin. Chem. Lett. 2014, 25, 802–804. [Google Scholar] [CrossRef]
- Yang, Y.; Zhang, J.; Wu, D.; Xing, Z.; Zhou, Y.; Shi, W.; Li, Q. Chemoenzymatic synthesis of polymeric materials using lipases as catalysts: A review. Biotechnol. Adv. 2014, 32, 642–651. [Google Scholar] [CrossRef]
- Parfene, G.; Horincar, V.B.; Bahrim, G.; Vannini, L.; Gottardi, D.; Maria Elisabetta, G. Lipolytic activity of lipases from different strains of Yarrowia lipolytica in hydrolysed vegetable fats at low temperature and water activity. Rom. Biotechnol. Lett. 2011, 16, 46–52. [Google Scholar]
- Destain, J.; Roblain, D.; Thonart, P. Improvement of lipase production from Yarrowia lipolytica. Biotechnol. Lett. 1997, 19, 105–108. [Google Scholar] [CrossRef]
- Pignede, G.; Wang, H.J.; Fudalej, F.; Seman, M.; Gaillardin, C.; Nicaud, J.M. Autocloning and amplification of LIP2 in Yarrowia lipolytica. Appl. Environ. Microbiol. 2000, 66, 3283–3289. [Google Scholar] [CrossRef] [PubMed]
- Fickers, P.; Fudalej, F.; Nicaud, J.M.; Destain, J.; Thonart, P. Selection of new over-producing derivatives for the improvement of extracellular lipase production by the non-conventional yeast Yarrowia lipolytica. J. Biotechnol. 2005, 115, 379–386. [Google Scholar] [CrossRef] [PubMed]
- Białecka-Florjańczyk, E.; Krzyczkowska, J.; Stolarzewicz, I.; Kapturowska, A. Synthesis of 2-phenylethyl acetate in the presence of Yarrowia lipolytica KKP 379 biomass. J. Mol. Catal. B Enzym. 2012, 74, 241–245. [Google Scholar] [CrossRef]
- Napora, K.; Wrodnigg, T.M.; Kosmus, P.; Thonhofer, M.; Robins, K.; Winkler, M. Yarrowia lipolytica dehydrogenase/reductase: An enzyme tolerant for lipophilic compounds and carbohydrate substrates. Bioorganic Med. Chem. Lett. 2013, 23, 3393–3395. [Google Scholar] [CrossRef] [PubMed]
- Druta, I.; Dinica, R.M.; Bacu, E.; Humelnicu, I. Synthesis of 7, 7’-bisindolizines by the reaction of 4,4’-bipyridinium-ylides with activated alkynes. Tetrahedron 1998, 54, 10811–10818. [Google Scholar] [CrossRef]
- Mazumder, P.; Khwairakpam, M.; Kalamdhad, A.S. Bio-inherent attributes of water hyacinth procured from contaminated water body–effect of its compost on seed germination and radicle growth. J. Environ. Manag. 2020, 257, 109990. [Google Scholar] [CrossRef]
- Li, R.; He, J.; Xie, H.; Wang, W.; Bose, S.K.; Sun, Y.; Hu, J.; Yin, H. Effects of chitosan nanoparticles on seed germination and seedling growth of wheat (Triticum aestivum L.). Int. J. Biol. Macromol. 2019, 126, 91–100. [Google Scholar] [CrossRef]
- Cudalbeanu, M.; Furdui, B.; Cârâc, G.; Barbu, V.; Iancu, A.V.; Campello, M.P.C.; Leitão, J.H.; Sousa, S.A.; Dinica, R.M. Antifungal, Antitumoral and Antioxidant Potential of the Danube Delta Nymphaea alba Extracts. Antibiotion 2019, 9, 7. [Google Scholar] [CrossRef]
- Tabacaru, A.; Botezatu-Dediu, A.-V.; Horincar, G.; Furdui, B.; Dinica, R.M. Green Accelerated Synthesis, Antimicrobial Activity. Molecules 2019, 24, 2424. [Google Scholar] [CrossRef] [PubMed]
- OECD. OECD test guideline 208: Terrestrial plant test—Seedling emergence and seedling growth test. Guidel. Test Chem. Terr. Plant Test Seedl. Emerg. Seedl. Growth Test 2006, 227, 1–21. [Google Scholar]
- Luo, Y.; Liang, J.; Zeng, G.; Chen, M.; Mo, D.; Li, G.; Zhang, D. Seed germination test for toxicity evaluation of compost: Its roles, problems and prospects. Waste Manag. 2018, 71, 109–114. [Google Scholar] [CrossRef] [PubMed]
- Tiquia, S.M.; Tam, N.F.Y.; Hodgkiss, I.J. Effects of composting on phytotoxicity of spent pig-manure sawdust litter. Environ. Pollut. 1996, 93, 249–256. [Google Scholar] [CrossRef]
- Barth, G.; Gaillardin, C. Yarrowia lipolytica. In Nonconventional Yeasts in Biotechnology: A Handbook; Springer: Berlin/Heidelberg, Germany, 1996; pp. 313–388. ISBN 978-3-642-79856-6. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Physiological Parameters | Concentration of the Synthetic Indolizines | Control | |||
|---|---|---|---|---|---|
| 4a 10−3 M | 4a 10−4 M | 4c 10−3 M | 4c 10−4 M | ||
| SG% | 80.33 ± 4.16 | 86.67 ± 4.73 | 81.00 ± 2.01 | 81.33 ± 6.11 | 80.00 ± 6.32 |
| RRG% | 110.26 ± 3.11 | 108.7 ± 2.75 | 101.7 ± 2.98 | 102.91 ± 2.48 | 100.00 ± 0.00 |
| RSG% | 104.68 ± 4.2 | 113.19 ± 5.3 | 105.48 ± 3.8 | 108.33 ± 5.1 | 100.00 ± 0.00 |
| GI% | 88.57 ± 3.6 | 94.21 ± 3.7 | 82.38 ± 2.49 | 83.69 ± 4.2 | 80.00 ± 6.2 |
| VI% | 85.77 ± 0.87 | 79.13 ± 0.48 | 89.06 ± 0.73 | 93.74 ± 0.96 | 85.80 ± 0.52 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Botezatu, A.V.; Horincar, G.; Ghinea, I.O.; Furdui, B.; Bahrim, G.-E.; Barbu, V.; Balanescu, F.; Favier, L.; Dinica, R.-M. Whole-Cells of Yarrowia lipolytica Applied in “One Pot” Indolizine Biosynthesis. Catalysts 2020, 10, 629. https://doi.org/10.3390/catal10060629
Botezatu AV, Horincar G, Ghinea IO, Furdui B, Bahrim G-E, Barbu V, Balanescu F, Favier L, Dinica R-M. Whole-Cells of Yarrowia lipolytica Applied in “One Pot” Indolizine Biosynthesis. Catalysts. 2020; 10(6):629. https://doi.org/10.3390/catal10060629
Chicago/Turabian StyleBotezatu (Dediu), Andreea Veronica, Georgiana Horincar, Ioana Otilia Ghinea, Bianca Furdui, Gabriela-Elena Bahrim, Vasilica Barbu, Fanica Balanescu, Lidia Favier, and Rodica-Mihaela Dinica. 2020. "Whole-Cells of Yarrowia lipolytica Applied in “One Pot” Indolizine Biosynthesis" Catalysts 10, no. 6: 629. https://doi.org/10.3390/catal10060629
APA StyleBotezatu, A. V., Horincar, G., Ghinea, I. O., Furdui, B., Bahrim, G.-E., Barbu, V., Balanescu, F., Favier, L., & Dinica, R.-M. (2020). Whole-Cells of Yarrowia lipolytica Applied in “One Pot” Indolizine Biosynthesis. Catalysts, 10(6), 629. https://doi.org/10.3390/catal10060629