Design and Construction of an Effective Expression System with Aldehyde Tag for Site-Specific Enzyme Immobilization




Abstract

1. Introduction
2. Results and Discussion
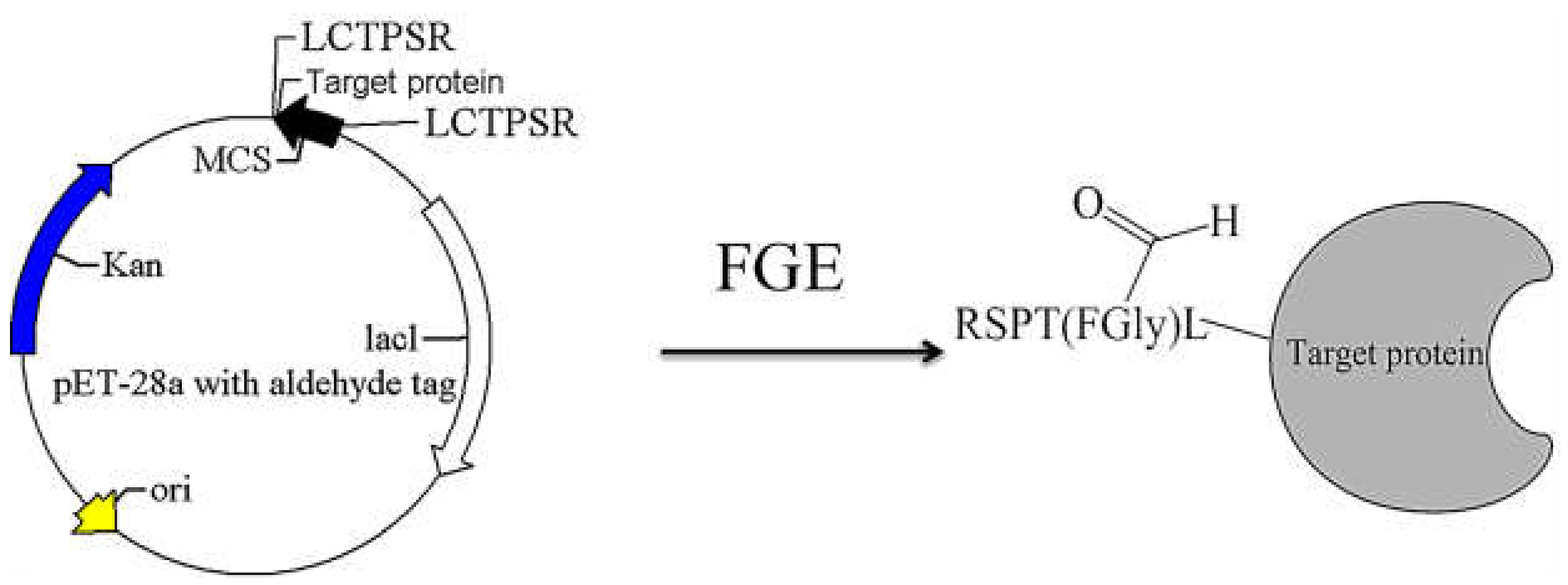
2.1. Construction of Recombinant Plasmid with the Aldehyde Tag
2.2. Expression and Activity Assay of the Recombinant Proteins
2.3. Identification of Aldehyde Tag
2.4. Immobilization Efficiency and Activity Recovery Rate
2.5. Reutilization of Immobilized Enzyme
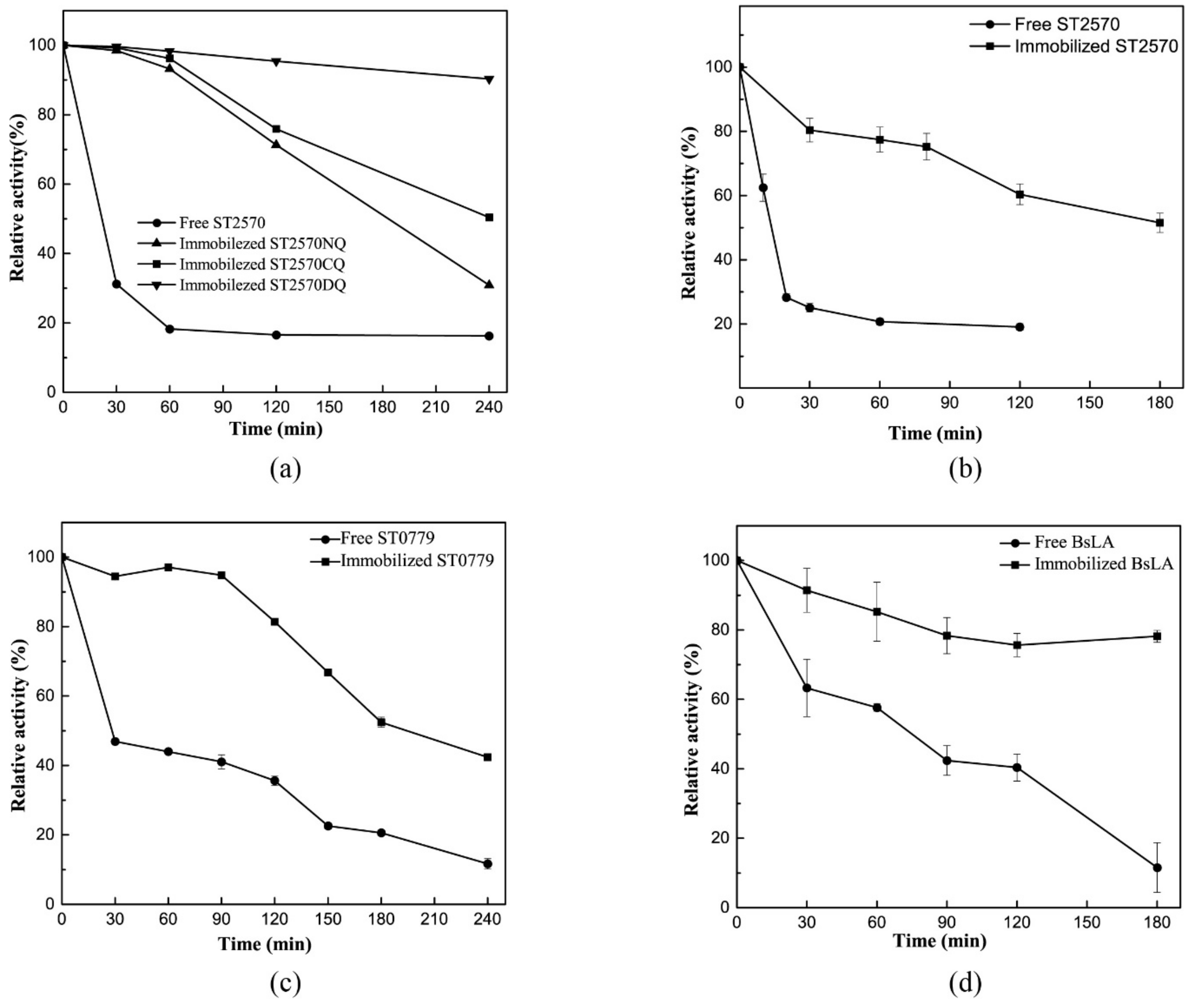
2.6. The Thermostability of Immobilized Enzyme
3. Materials and Methods
3.1. Materials
3.2. Construction and Transformation of the Recombinant Plasmid with the Aldehyde Tag
3.3. Expression and Purification of the Recombinant Proteins in E. coli
3.4. Examination of the Genetically Encoded Aldehyde Tag Using Reporter Labeling
3.5. Enzyme Assay
3.6. Covalent Immobilization
3.7. The Reutilization and Thermostability of Immobilized Enzyme
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Bilal, M.; Asgher, M.; Parra-Saldivar, R. Immobilized ligninolytic enzymes: An innovative and environmental responsive technology to tackle dye-based industrial pollutants—A review. Sci Total Environ. 2017, 576, 646–659. [Google Scholar] [CrossRef] [PubMed]
- Bilal, M.; Iqbal, H.M.N.; Shuqi, G. State-of-the-art protein engineering approaches using biological macromolecules: A review from immobilization to implementation view point. Int. J. Biol. Macromol. 2018, 108, 893–901. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, J.C.S.; Barbosa, O.; Ortiz, C. Importance of the Support Properties for Immobilization or Purification of Enzymes. Chemcatchem 2015, 7, 2413–2432. [Google Scholar] [CrossRef]
- Garcia-Galan, C.; Berenguer-Murcia, A.; Fernandez-Lafuente, R. Potential of Different Enzyme Immobilization Strategies to Improve Enzyme Performance. Adv. Synth. Catal. 2011, 353, 2885–2904. [Google Scholar] [CrossRef]
- Sheldon, R.A. Enzyme immobilization: The quest for optimum performance. Adv. Synth. Catal. 2007, 349, 1289–1307. [Google Scholar] [CrossRef]
- Sheldon, R.A.; Van Pelt, S. Enzyme immobilisation in biocatalysis: Why, what and how. Chem. Soc. Rev. 2013, 42, 6223–6235. [Google Scholar] [CrossRef] [PubMed]
- Cowan, D.A.; Fernandez-Lafuente, R. Enhancing the functional properties of thermophilic enzymes by chemical modification and immobilization, Enzyme Microb. Technol. 2011, 49, 326–346. [Google Scholar]
- Sheldon, R.A. Characteristic features and biotechnological applications of cross-linked enzyme aggregates (CLEAs). Appl. Microbiol. Biotechnol. 2011, 92, 467–477. [Google Scholar] [CrossRef]
- Cabana, H.; Jones, J.P.; Agathos, S.N. Utilization of cross-linked laccase aggregates in a perfusion basket reactor for the continuous elimination of endocrine-disrupting chemicals. Biotechnol. Bioeng. 2009, 102, 1582–1592. [Google Scholar] [CrossRef]
- Subramanian, A.; Woodward, J.; Doktycz, M.J.; Kennel, S.J. Comparison of techniques for enzyme immobilization on silicon supports. Enzyme Microb. Technol. 1999, 24, 26–34. [Google Scholar] [CrossRef]
- Brady, D.; Jordaan, J. Advances in enzyme immobilisation. Biotechnol. Lett. 2009, 31, 1639–1650. [Google Scholar] [CrossRef] [PubMed]
- Ispas, C.; Sokolov, I.; Andreescu, S. Enzyme-functionalized mesoporous silica for bioanalytical applications. Anal. Bioanal. Chem. 2009, 393, 543–554. [Google Scholar] [CrossRef] [PubMed]
- Ovsejevi, K.; Manta, C.; Batista-Viera, F. Reversible covalent immobilization of enzymes via disulfide bonds. Methods Mol. Biol. 2013, 1051, 89–116. [Google Scholar] [PubMed]
- Rusmini, F.; Zhong, Z.; Feijen, J. Protein immobilization strategies for protein biochips. Biomacromolecules 2007, 8, 1775–1789. [Google Scholar] [CrossRef]
- Nilsson, J.; Ståhl, S.; Lundeberg, J. Affinity fusion strategies for detection, purification, and immobilization of recombinant proteins. Protein Expres. Purif. 1997, 11, 1–16. [Google Scholar] [CrossRef]
- Zakeri, B.; Fierer, J.O.; Celik, E. Peptide tag forming a rapid covalent bond to a protein, through engineering a bacterial adhesin. Proc. Nati. Acad. Sci. USA 2012, 109, 690–697. [Google Scholar] [CrossRef]
- Lesaicherre, M.L.; Uttamchandani, M.; Chen, G.Y.J. Developing site-specific immobilization strategies of peptides in a microarray. Bioorg. Med. Chem. Lett. 2002, 12, 2079–2083. [Google Scholar] [CrossRef]
- Hernandez, K.; Fernandez-Lafuente, R. Control of protein immobilization: Coupling immobilization and site-directed mutagenesis to improve biocatalyst or biosensor performance. Enzyme Microb. Tech. 2011, 48, 107–122. [Google Scholar] [CrossRef]
- Tiefenbrunn, T.K.; Dawson, P.E. Chemoselective ligation techniques: Modern applications of time-honored chemistry. Peptide Sci. 2010, 94, 95–106. [Google Scholar] [CrossRef]
- Scheck, R.A.; Dedeo, M.T.; Iavarone, A.T.; Francis, M.B. Optimization of a biomimetic transamination reaction. J. Am. Chem. Soc. 2008, 130, 11762–11770. [Google Scholar] [CrossRef]
- Geoghegan, K.F.; Stroh, J.G. Site-directed conjugation of nonpeptide groups to peptides and proteins via periodate oxidation of a 2-amino alcohol. Application to modification at N-terminal serine. Bioconjugate Chem. 1992, 3, 138–146. [Google Scholar] [CrossRef] [PubMed]
- Rush, J.S.; Bertozzi, C.R. New Aldehyde Tag Sequences Identified by Screening Formylglycine Generating Enzymes in Vitro and in Vivo. J. Am. Chem. Soc. 2008, 130, 12240–12241. [Google Scholar] [CrossRef] [PubMed]
- Peng, Q.; Zang, B.; Zhao, W.; Li, D.; Ren, J.; Ji, F.; Jia, L. Efficient continuous-flow aldehyde tag conversion using immobilized formylglycine generating enzyme. Catal. Sci. Technol. 2020, 10. [Google Scholar] [CrossRef]
- Wang, A.; Du, F.; Wang, F.; Shen, Y.; Gao, W.; Zhang, P. Convenient one-step purification and immobilization of lipase using a genetically encoded aldehyde tag. Biochem. Eng. J. 2013, 73, 86–92. [Google Scholar] [CrossRef]
- Carrico, I.S.; Carlson, B.L.; Bertozzi, C.R. Introducing genetically encoded aldehydes into proteins. Nat. Chem. Biol. 2007, 3, 321–322. [Google Scholar] [CrossRef]
- Wu, P.; Shui, W.; Carlson, B.L.; Hu, N.; Rabuka, D.; Lee, J.; Bertozzi, C.R. Site-specific chemical modification of recombinant proteins produced in mammalian cells by using the genetically encoded aldehyde tag. Proc. Natl. Acad. Sci. USA 2009, 106, 3000–3005. [Google Scholar] [CrossRef]
- Rabuka, D.; Rush, J.S.; deHart, G.W.; Wu, P.; Bertozzi, C.R. Site-specific chemical protein conjugation using genetically encoded aldehyde tags. Nat. Protoc. 2012, 7, 1052–1067. [Google Scholar] [CrossRef]
- Jian, H.; Wang, Y.; Bai, Y.; Li, R.; Gao, R. Site-specific, covalent immobilization of dehalogenase ST2570 catalyzed by formylglycine-generating enzymes and its application in batch and semi-continuous flow reactors. Molecules 2016, 21, 895. [Google Scholar] [CrossRef]
- Lyu, J.; Li, Z.; Men, J. Covalent immobilization of Bacillus subtilis lipase A on Fe3O4 nanoparticles by aldehyde tag: An ideal immobilization with minimal chemical modification. Process Biochem. 2019, 81, 63–69. [Google Scholar] [CrossRef]
- Guimaraes, C.P.; Witte, M.D.; Theile, C.S. Site-specific C-terminal and internal loop labeling of proteins using sortase-mediated reactions. Nat. Protoc. 2013, 8, 1787–1799. [Google Scholar] [CrossRef]
- Theile, C.S.; Witte, M.D.; Blom, A.E.M. Site-specific N-terminal labeling of proteins using sortase-mediated reactions. Nat. Protoc. 2013, 8, 1800–1807. [Google Scholar] [CrossRef] [PubMed]
- Rye, C.A.; Isupov, M.N.; Lebedev, A.A. Biochemical and structural studies of a L-haloacid dehalogenase from the thermophilic archaeon Sulfolobus tokodaii. Extremophiles 2009, 13, 179–190. [Google Scholar] [CrossRef] [PubMed]
- Rye, C.A.; Isupov, M.N.; Lebedev, A.A. An order-disorder twin crystal of l-2-haloacid dehalogenase from Sulfolobus tokodaii. Acta Crystallogr. D 2007, 63, 926–930. [Google Scholar] [CrossRef] [PubMed]
- Van Pouderoyen, G.; Eggert, T.; Jaeger, K.E. The crystal structure of Bacillus subtilis lipase: A minimal alpha/beta hydrolase fold enzyme. J. Mol. Biol. 2001, 309, 215–226. [Google Scholar] [CrossRef] [PubMed]
- Bartlam, M.; Wang, G.; Yang, H.; Gao, R.; Zhao, X.; Xie, G. Crystal structure of an acylpeptide hydrolase/esterase from Aeropyrum pernix k1. Structure (Cambridge) 2004, 12, 1481–1488. [Google Scholar] [CrossRef] [PubMed]
- Rao, L.; Bi, Y.F.; Xie, G.Q. Study on a specific site tyr444 on a hyperthermophilic enzyme APE1547. Chem. Res. Chin. Univ. 2009, 25, 353–356. [Google Scholar]
- Holyavka, M.G.; Kondratyev, M.S.; Lukin, A.N.; Agapov, B.L.; Artyukhov, V.G. Immobilization of inulinase on KU-2 ion-exchange resin matrix. Int. J. Biol. Macromol. 2019, 138, 681–692. [Google Scholar] [CrossRef]
- Ahmad, R.; Sardar, M. Enzyme immobilization: An overview on nanoparticles as immobilization matrix, Biochem. Anal. Biochem. 2015, 4, 1–8. [Google Scholar]
- Armenia, I.; Grazú Bonavia, M.V.; De Matteis, L.; Ivanchenko, P.; Martra, G.; Gornati, R.; de la Fuente, J.M.; Bernardini, G. Enzyme activation by alternating magnetic field: Importance of the bioconjugation methodology. J. Colloid. Interface. Sci. 2019, 537, 615–628. [Google Scholar] [CrossRef]
- Lu, A.H.; Salabas, E.L.; Schueth, F. Magnetic nanoparticles: Synthesis, protection, functionalization, and application. Angew. Chem. Internat. Ed. 2007, 46, 1222–1244. [Google Scholar] [CrossRef]
- Wu, W.; Wu, Z.; Yu, T.; Jiang, C.; Kim, W.S. Recent progress on magnetic iron oxide nanoparticles: Synthesis, surface functional strategies and biomedical applications, Sci. Technol. Adv. Mater. 2015, 16, 023501. [Google Scholar] [CrossRef] [PubMed]
- Rong, L.; Fei, Z.; Cao, S.G. Expression and Characterization of a Thermostable Acyl-peptide Releasing Enzyme ST0779 from Sulfolobus tokodaii. Chem. Res. Chin. Univ. 2012, 28, 851–855. [Google Scholar]
- Mohamad, N.R.; Marzuki, N.H.C.; Buang, N.A.; Huyop, F.; Wahab, R.A. An overview of technologies for immobilization of enzymes and surface analysis techniques for immobilized enzymes. Biotechnol. Biotec. Eq. 2015, 29, 205–220. [Google Scholar] [CrossRef] [PubMed]
- Sheldon, R.A. Cross-linked enzyme aggregates (CLEAs): Stable and recyclable biocatalysts. Biochem. Soc. 2007, 35, 1583–1587. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.; Anming, W.; Lifeng, H.; Haifeng, L.; Zhenming, C.; Qiuyan, W.; Xiaopu, Y. Recent advance in the support and technology used in enzyme immobilization. Afr. J. Biotechnol. 2009, 8, 4724–4733. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Immobilized Enzyme | Activity Recovery Rate (%) | Immobilization Efficiency (%) |
|---|---|---|
| BsLACQ | 68.05 | 93.72 |
| BsLANQ | 22.74 | 89.94 |
| BsLADQ | 51.33 | 51.33 |
| Immobilized Enzyme | Activity (U/g) | Activity Recovery Rate (%) | Immobilization Efficiency (%) |
|---|---|---|---|
| ST0779CQ | 21.7 | 86.7 | 84.6 |
| ST0779NQ | 16.8 | 38.5 | 82.3 |
| ST0779DQ | 11.1 | 72.9 | 67.7 |
| Immobilized Enzyme | Activity (U/g) | Activity Recovery Rate (%) | Immobilization Efficiency (%) |
|---|---|---|---|
| ST2570CQ | 187.2 | 40.7 | 32.7 |
| ST2570NQ | 105.6 | 25.6 | 13.5 |
| ST2570DQ | 168.9 | 59.9 | 58.9 |
| Name | Primer Sequence (5’ to 3’) |
|---|---|
| 28aDH up | AGATATCATATGGGCAGCAGCCAT |
| 28aDH down | CTGCCCATATGATATCTCCTTCTTAAAG |
| 28aNQ up | CAGCCTTTGTACTCCTTCTCGCAGCAGCGGCCTGG |
| 28aNQ down | TGCTGCGAGAAGGAGTACAAAGGCTGCTGCCCATGG |
| 28aCQ up | CGAGCACTATGTACACCAAGTCGGTAATCCGGCTGCTAAC |
| 28aCQ down | CGGATTACCGACTTGGTGTACATAGTGCTCGAGTGCGGCC |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, F.; Li, R.; Jian, H.; Huang, Z.; Wang, Y.; Guo, Z.; Gao, R. Design and Construction of an Effective Expression System with Aldehyde Tag for Site-Specific Enzyme Immobilization. Catalysts 2020, 10, 410. https://doi.org/10.3390/catal10040410
Wang F, Li R, Jian H, Huang Z, Wang Y, Guo Z, Gao R. Design and Construction of an Effective Expression System with Aldehyde Tag for Site-Specific Enzyme Immobilization. Catalysts. 2020; 10(4):410. https://doi.org/10.3390/catal10040410
Chicago/Turabian StyleWang, Fang, Rong Li, Hui Jian, Zihao Huang, Yingwu Wang, Zheng Guo, and Renjun Gao. 2020. "Design and Construction of an Effective Expression System with Aldehyde Tag for Site-Specific Enzyme Immobilization" Catalysts 10, no. 4: 410. https://doi.org/10.3390/catal10040410
APA StyleWang, F., Li, R., Jian, H., Huang, Z., Wang, Y., Guo, Z., & Gao, R. (2020). Design and Construction of an Effective Expression System with Aldehyde Tag for Site-Specific Enzyme Immobilization. Catalysts, 10(4), 410. https://doi.org/10.3390/catal10040410