Aldoxime Dehydratase Mutants as Improved Biocatalysts for a Sustainable Synthesis of Biorenewables-Based 2-Furonitrile



Abstract
:1. Introduction
2. Results and Discussion
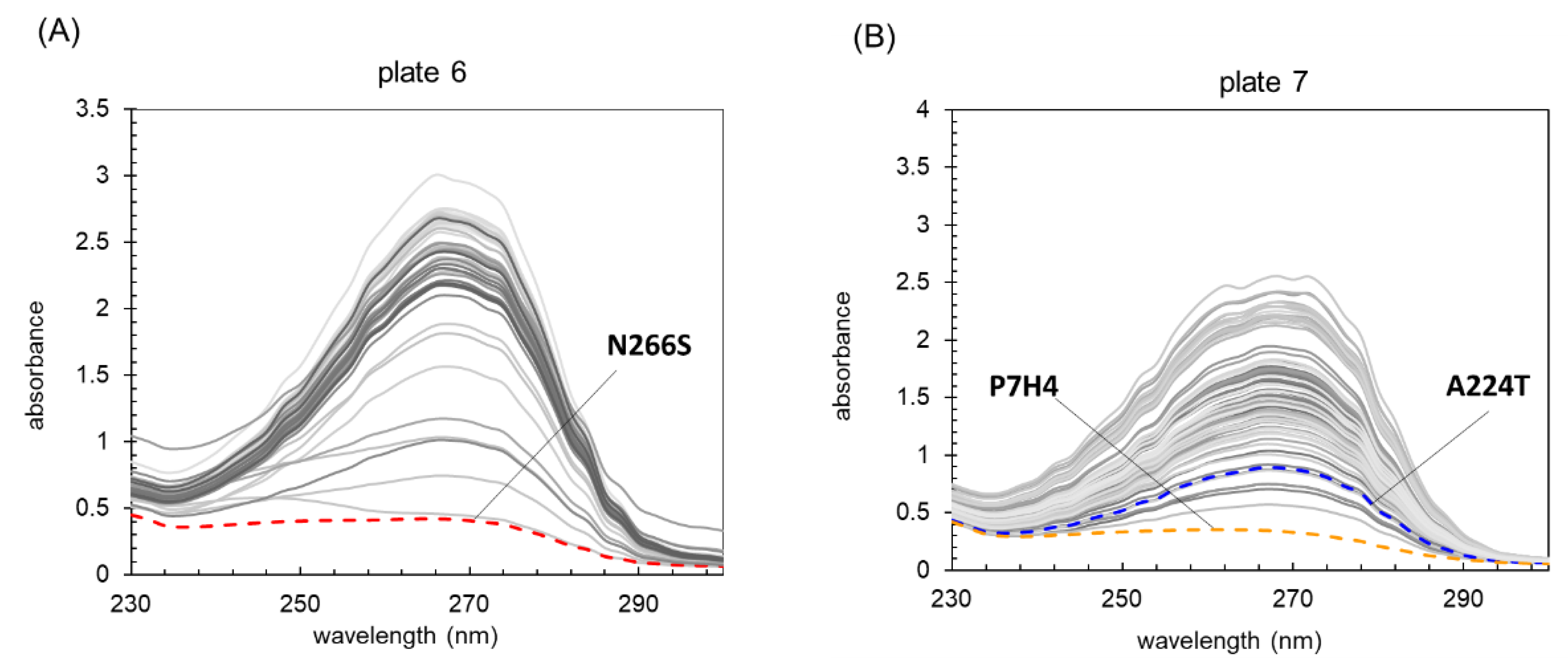
2.1. High Throughput Screening of OxdYH3-3 Mutants by Using a UV Microspectrometer
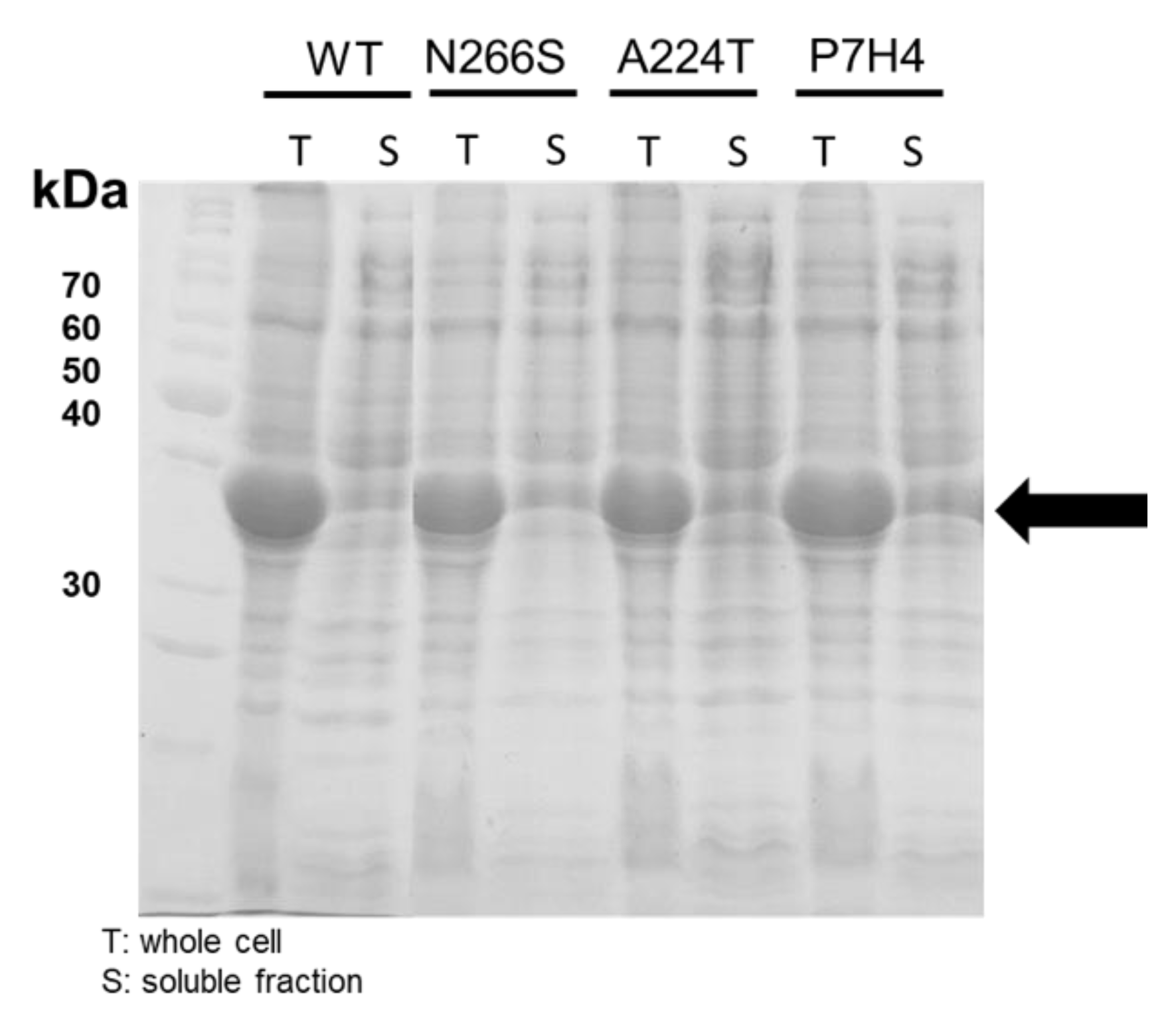
2.2. The Improved Activity Measurement of OxdYH3-3 Mutants
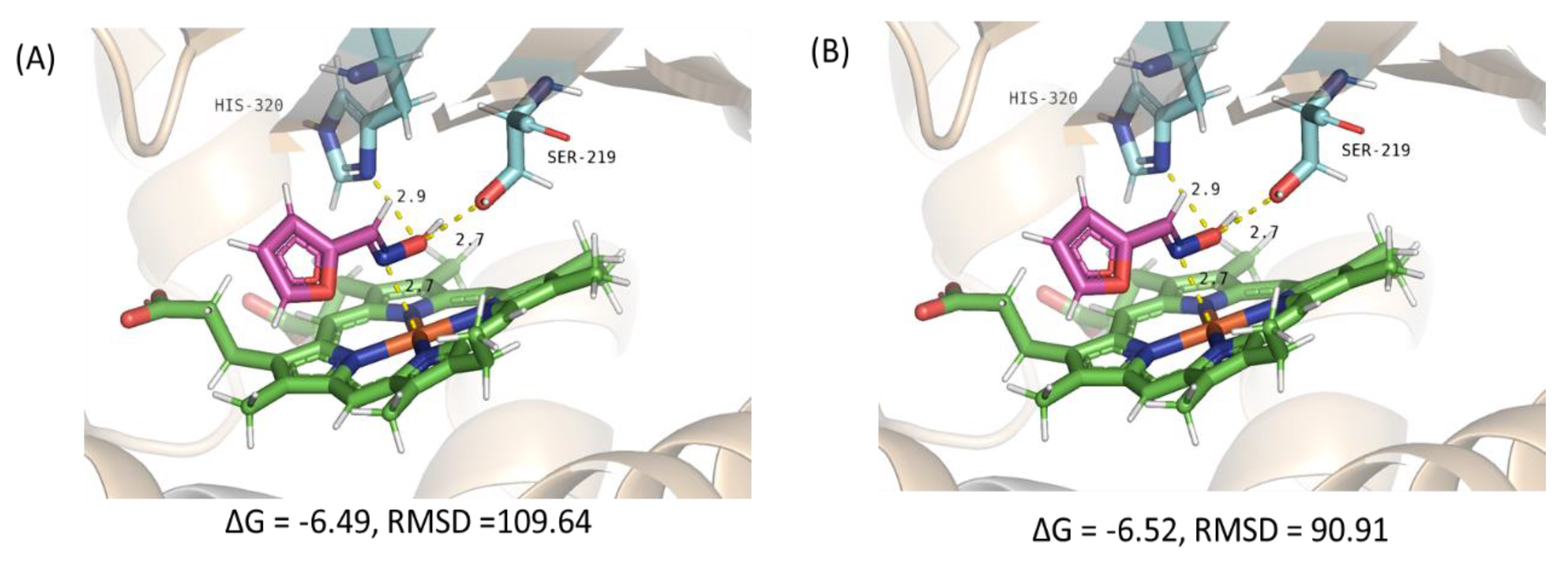
2.3. Homology Modelling of OxdYH WT and N266S
3. Materials and Methods
3.1. Materials and Preparation of OxdYH3-3 Transformants
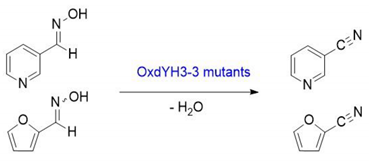
3.2. Synthesis of 2-Furfuryl Aldoxime
3.3. Construction of Mutant Library by Error-Prone PCR
3.4. Screening of OxdYH3-3 WT and Mutants by of UV Spectrum Scan
3.5. Analysis of the DNA Sequence of OxdYH3-3 Mutants
3.6. Protein Purification
3.7. Characterization of Purified OxdYH3-3 Catalyst
3.8. Conversion of Aldoximes to Corresponding Nitriles by Using OxdYH3-3 WT and Mutants
3.9. Determination of Conversion from Aldoximes to Nitrile by GC
3.10. Measurement of the UV Spectrum of Aldoximes and Nitriles After the Reaction
3.11. Homology Modelling
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgements
Conflicts of Interest
References
- Dominguez de Maria, P. Industrial Biorenewables: A Practical Viewpoint; Wiley: Hoboken, NJ, USA, 2016. [Google Scholar]
- Jennings, T.J. Process for Preparing Furonitrile. U.S. Patent 3,260,731, 1964. [Google Scholar]
- Bohre, A.; Dutta, S.; Saha, B.; Abu-Omar, M.M. Upgrading furfurals to drop-in biofuels: An overview. ACS Sustain. Chem. Eng. 2015, 3, 1263–1277. [Google Scholar] [CrossRef]
- Gandini, A.; Belgacem, M.N. Furans in polymer chemistry. Prog. Polym. Sci. 1997, 22, 1203–1379. [Google Scholar] [CrossRef]
- Chatterjee, M.; Ishizaka, T.; Kawanami, H. Reductive amination of furfural to furfurylamine using aqueous ammonia solution and molecular hydrogen: An environmentally friendly approach. Green Chem. 2016, 18, 487–496. [Google Scholar] [CrossRef]
- Nishimura, S.; Mizuhori, K.; Ebitani, K. Reductive amination of furfural toward furfurylamine with aqueous ammonia under hydrogen over Ru-supported catalyst. Res. Chem. Intermed. 2016, 42, 19–30. [Google Scholar] [CrossRef]
- Dunbabin, A.; Subrizi, F.; Ward, J.M.; Sheppard, T.D.; Hailes, H.C. Furfurylamines from biomass: Transaminase catalysed upgrading of furfurals. Green Chem. 2017, 19, 397–404. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.H.; Chang, S. Highly efficient and catalytic conversion of aldoximes to nitriles. Org. Lett. 2001, 3, 4209–4211. [Google Scholar] [CrossRef]
- Bandgar, B.P.; Makone, S.S. Organic reactions in water: Transformation of aldehydes to nitriles using NBS under mild conditions. Synth. Commun. 2006, 36, 1347–1352. [Google Scholar] [CrossRef]
- Narsaiah, A.; Sreenu, D.; Nagaiah, K. Direct oxidative conversion of alcohols, aldehydes and amines into nitriles using hypervalent iodine(III) reagent. Synth. Commun. 2006, 36, 137–140. [Google Scholar] [CrossRef]
- Yadav, L.D.S.; Srivastava, V.P.; Patel, R. Bromodimethylsulfonium bromide (BDMS): A useful reagent for conversion of aldoximes and primary amides to nitriles. Tetrahedron Lett. 2009, 50, 5532–5535. [Google Scholar] [CrossRef]
- Sridhar, M.; Reddy, M.K.K.; Sairam, V.V.; Raveendra, J.; Godala, K.R.; Narsaiah, C.; Ramanaiah, B.C.; Reddy, C.S. Acetohydroxamic acid: A new reagent for efficient synthesis of nitriles directly from aldehydes using Bi(OTf)3 as the catalyst. Tetrahedron Lett. 2012, 53, 3421–3424. [Google Scholar] [CrossRef]
- Asano, Y.; Kato, Y. Z-Phenylacetaldoxime degradation by a novel aldoxime dehydratase from Bacillus sp. strain OxB-1. FEMS Microbiol. Lett. 1998, 158, 185–190. [Google Scholar] [CrossRef]
- Kato, Y.; Nakamura, K.; Sakiyama, H.; Mayhew, S.G.; Asano, Y. Novel heme-containing lyase, phenylacetaldoxime dehydratase from Bacillus sp. strain OxB-1: Purification, characterization, and molecular cloning of the gene. Biochemistry 2000, 39, 800–809. [Google Scholar] [CrossRef] [PubMed]
- Kato, Y.; Ooi, R.; Asano, Y. Distribution of aldoxime dehydratase in microorganisms. Appl. Environ. Microbiol. 2000, 66, 2290–2296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sawai, H.; Sugimoto, H.; Kato, Y.; Asano, Y.; Shiro, Y.; Aono, S. X-ray crystal structure of michaelis complex of aldoxime dehydratase. J. Biol. Chem. 2009, 284, 32089–32096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaguchi, T.; Asano, Y. Draft genome sequence of an aldoxime degrader, Rhodococcus sp. strain YH3-3. Genome Announc. 2016, 4, 406–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Metzner, R.; Okazaki, S.; Asano, Y.; Gröger, H. Cyanide-free enantioselective synthesis of nitriles: Synthetic proof of a biocatalytic concept and mechanistic insights. ChemCatChem 2014, 6, 3105–3109. [Google Scholar] [CrossRef]
- Betke, T.; Rommelmann, P.; Oike, K.; Asano, Y.; Gröger, H. Cyanide-free and broadly applicable enantioselective synthetic platform for chiral nitriles through a biocatalytic approach. Angew. Chem. Int. Ed. 2017, 56, 12361–12366. [Google Scholar] [CrossRef]
- Betke, T.; Maier, M.; Gruber-Wölfler, H.; Gröger, H. Biocatalytic production of adiponitrile and related aliphatic linear α,ω-dinitriles. Nat. Commun. 2018, 9, 5112. [Google Scholar] [CrossRef] [Green Version]
- Hinzmann, A.; Glinski, S.; Worm, M.; Gröger, H. Enzymatic synthesis of aliphatic nitriles at a substrate loading of up to 1.4 kg/L: A biocatalytic record achieved with a heme protein. J. Org. Chem. 2019, 84, 4867–4872. [Google Scholar] [CrossRef]
- Plass, C.; Hinzmann, A.; Terhorst, M.; Brauer, W.; Oike, K.; Yavuzer, H.; Asano, Y.; Vorholt, A.J.; Betke, T.; Gröger, H. Approaching bulk chemical nitriles from alkenes: A hydrogen cyanide-free approach through a combination of hydroformylation and biocatalysis. ACS Catal. 2019, 9, 5198–5203. [Google Scholar] [CrossRef]
- Betke, T.; Higuchi, J.; Rommelmann, P.; Oike, K.; Nomura, T.; Kato, Y.; Asano, Y.; Gröger, H. Biocatalytic synthesis of nitriles through dehydration of aldoximes: The substrate scope of aldoxime dehydratases. ChemBioChem 2018, 19, 768–779. [Google Scholar] [CrossRef] [PubMed]
- Kato, Y.; Ooi, R.; Asano, Y. A new enzymatic method of nitrile synthesis by Rhodococcus sp. strain YH3-3. J. Mol. Catal. B Enzym. 1999, 6, 249–256. [Google Scholar] [CrossRef]
- Choi, J.E.; Shinoda, S.; Inoue, R.; Zheng, D.; Gröger, H.; Asano, Y. Cyanide-free synthesis of an aromatic nitrile from a biorenewable-based aldoxime: Development and application of a recombinant aldoxime dehydratase as a biocatalyst. Biocat. Biotransf. 2019, 37, 414–420. [Google Scholar] [CrossRef]
- Oinuma, K.I.; Hashimoto, Y.; Konishi, K.; Goda, M.; Noguchi, T.; Higashibata, H.; Kobayashi, M. Novel aldoxime dehydratase involved in carbon-nitrogen triple bond synthesis of Pseudomonas chlororaphis B23 sequencing, gene expression, purification, and characterization. J. Bio. Chem. 2003, 278, 29600–29608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, S.X.; Kato, Y.; Komeda, H.; Yoshida, S.; Asano, Y. A gene cluster responsible for alkylaldoxime metabolism coexisting with nitrile hydratase and amidase in Rhodococcus globerulus A-4. Biochemistry 2003, 42, 12056–12066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kato, Y.; Yoshida, S.; Xie, S.X.; Asano, Y. Aldoxime dehydratase co-existing with nitrile hydratase and amidase in the iron-type nitrile hydratase-producer Rhodococcus sp. N-771. J. Biosci. Bioeng. 2004, 97, 250–259. [Google Scholar] [CrossRef]
- Piovesan, D.; Minervini, G.; Tosatto, S.C.E. The RING 2.0 web server for high quality residue interaction networks. Nucleic Acids Res. 2016, 44, 367–374. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- SWISS-MODEL. Available online: http://swissmodel.expasy.org (accessed on 13 February 2019).
- Eswar, N.; Eramian, D.; Webb, B.; Shen, M.Y.; Sali, A. Protein structure modeling with MODELLER. In Structural Proteomics; Kobe, B., Guss, M., Huber, T., Eds.; Methods in Molecular Biology™; Humana Press: Totowa, NJ, USA, 2008; Volume 426, pp. 145–149. [Google Scholar]
- Colovos, C.; Yeates, T.O. Verification of protein structures: Patterns of nonbonded atomic interactions. Protein Sci. 1993, 2, 1511–1519. [Google Scholar] [CrossRef] [Green Version]
- Hanwell, M.D.; Curtis, D.E.; Lonie, D.C.; Vandermeersch, T.; Zurek, E.; Hutchison, G.R. Avogadro: An advanced semantic chemical editor, visualization, and analysis platform. J. Cheminform. 2012, 4, 17. [Google Scholar] [CrossRef] [Green Version]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wallace, A.C.; Laskowski, R.A.; Thornton, J.M. Ligplot: A program to generate schematic diagrams of protein-ligand interactions. Protein Eng. Des. Sel. 1995, 8, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| U·g−1 (µmol·min−1·g−1) | ||||
|---|---|---|---|---|
| OxdYH3-3 | E-3-pyridine aldoxime | 2-furfuryl aldoxime | ||
| 50 mM | 100 mM | 50 mM | 100 mM | |
| WT | 0.62 | 0.52 | 0.15 | 0.29 |
| N266S | 1.85 | 1.73 | 1.24 | 1.16 |
| A224T | 0.68 | 0.52 | 0.37 | 0.28 |
| P7H4 | 0.81 | 0.78 | 0.53 | 0.38 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, J.E.; Shinoda, S.; Asano, Y.; Gröger, H. Aldoxime Dehydratase Mutants as Improved Biocatalysts for a Sustainable Synthesis of Biorenewables-Based 2-Furonitrile. Catalysts 2020, 10, 362. https://doi.org/10.3390/catal10040362
Choi JE, Shinoda S, Asano Y, Gröger H. Aldoxime Dehydratase Mutants as Improved Biocatalysts for a Sustainable Synthesis of Biorenewables-Based 2-Furonitrile. Catalysts. 2020; 10(4):362. https://doi.org/10.3390/catal10040362
Chicago/Turabian StyleChoi, Ji Eun, Suguru Shinoda, Yasuhisa Asano, and Harald Gröger. 2020. "Aldoxime Dehydratase Mutants as Improved Biocatalysts for a Sustainable Synthesis of Biorenewables-Based 2-Furonitrile" Catalysts 10, no. 4: 362. https://doi.org/10.3390/catal10040362
APA StyleChoi, J. E., Shinoda, S., Asano, Y., & Gröger, H. (2020). Aldoxime Dehydratase Mutants as Improved Biocatalysts for a Sustainable Synthesis of Biorenewables-Based 2-Furonitrile. Catalysts, 10(4), 362. https://doi.org/10.3390/catal10040362