Structural and Functional Analysis of the Only Two Pyridoxal 5′-Phosphate-Dependent Fold Type IV Transaminases in Bacillus altitudinis W3


Abstract
1. Introduction
2. Results and Discussion
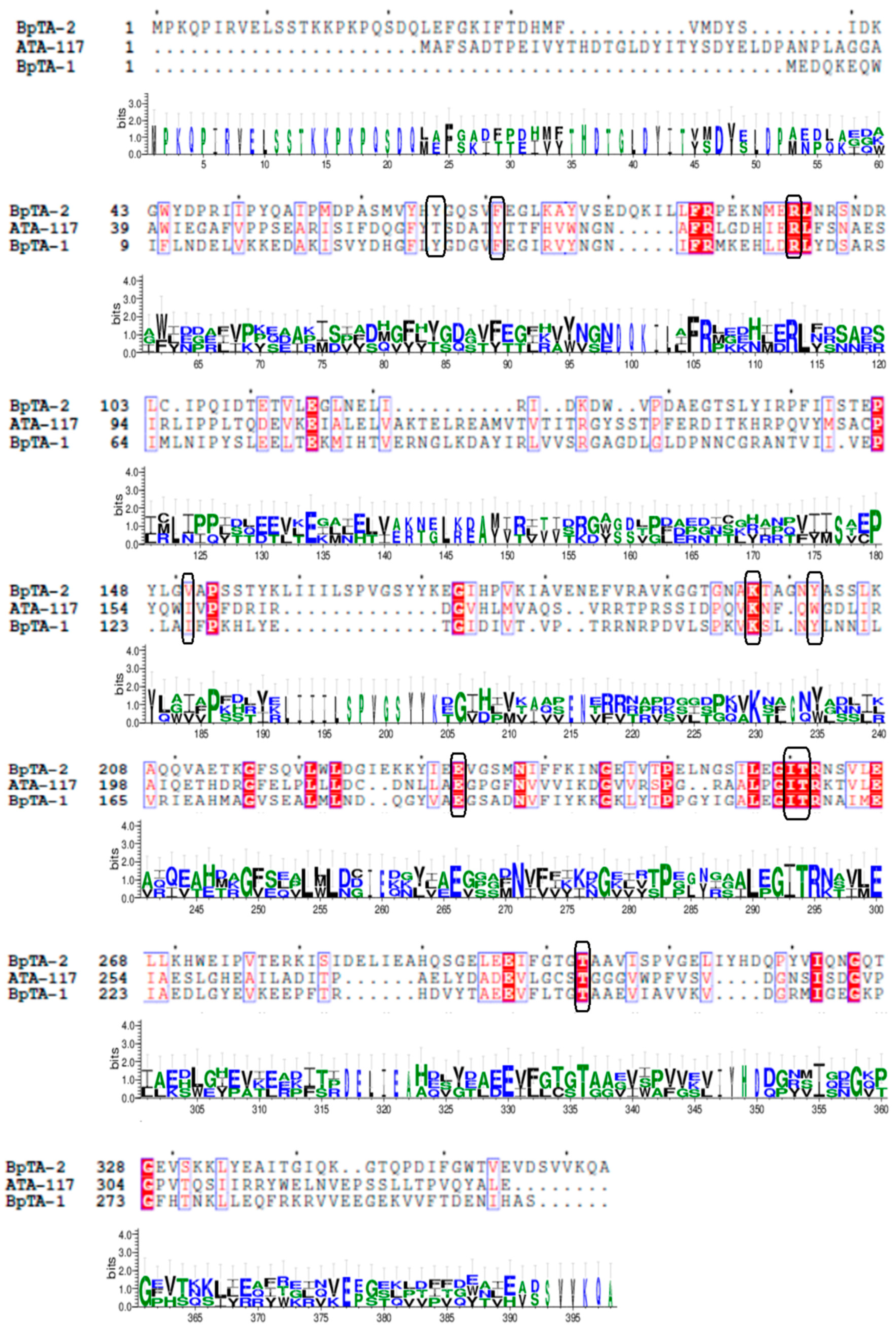
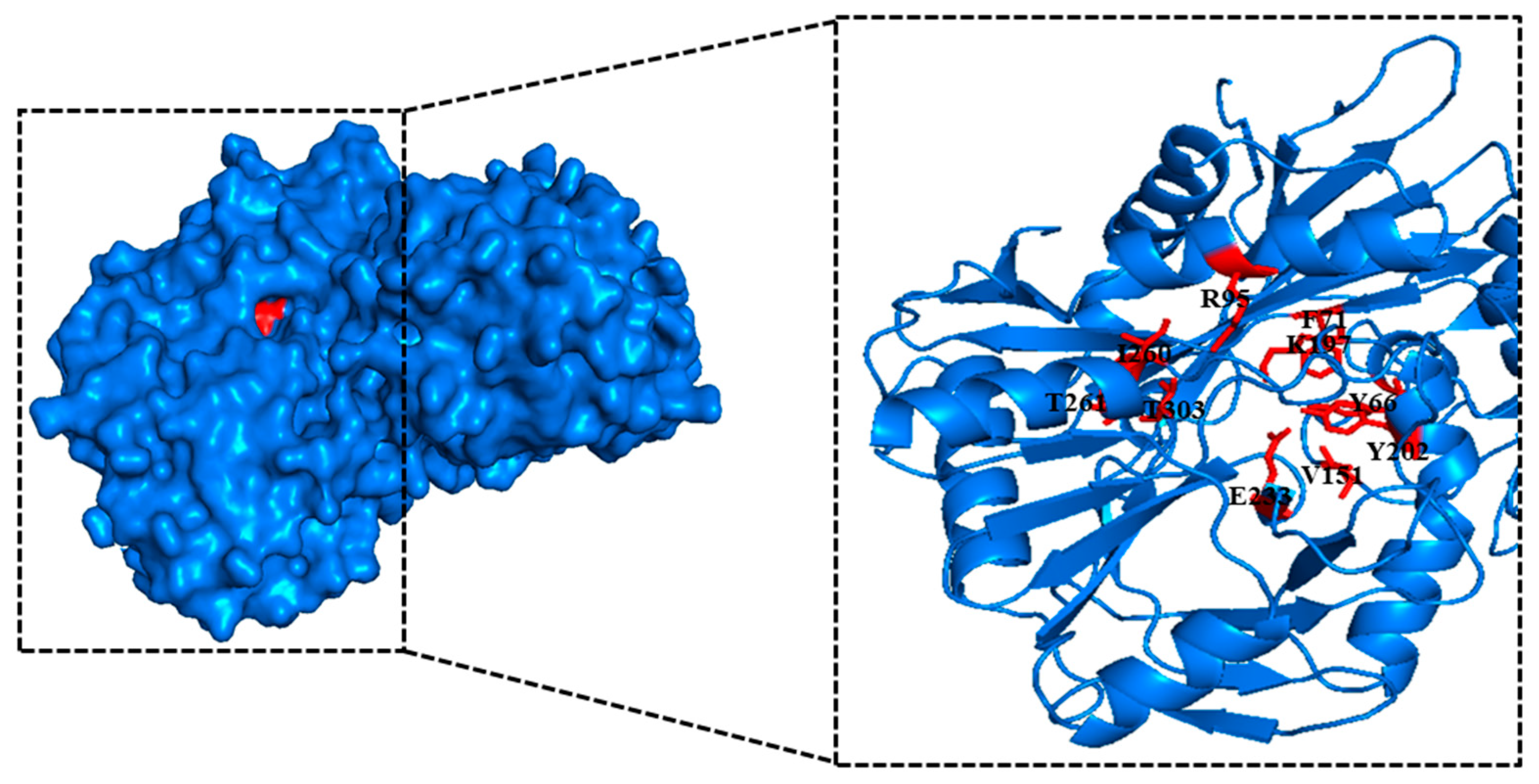
2.1. Bioinformatic Analysis
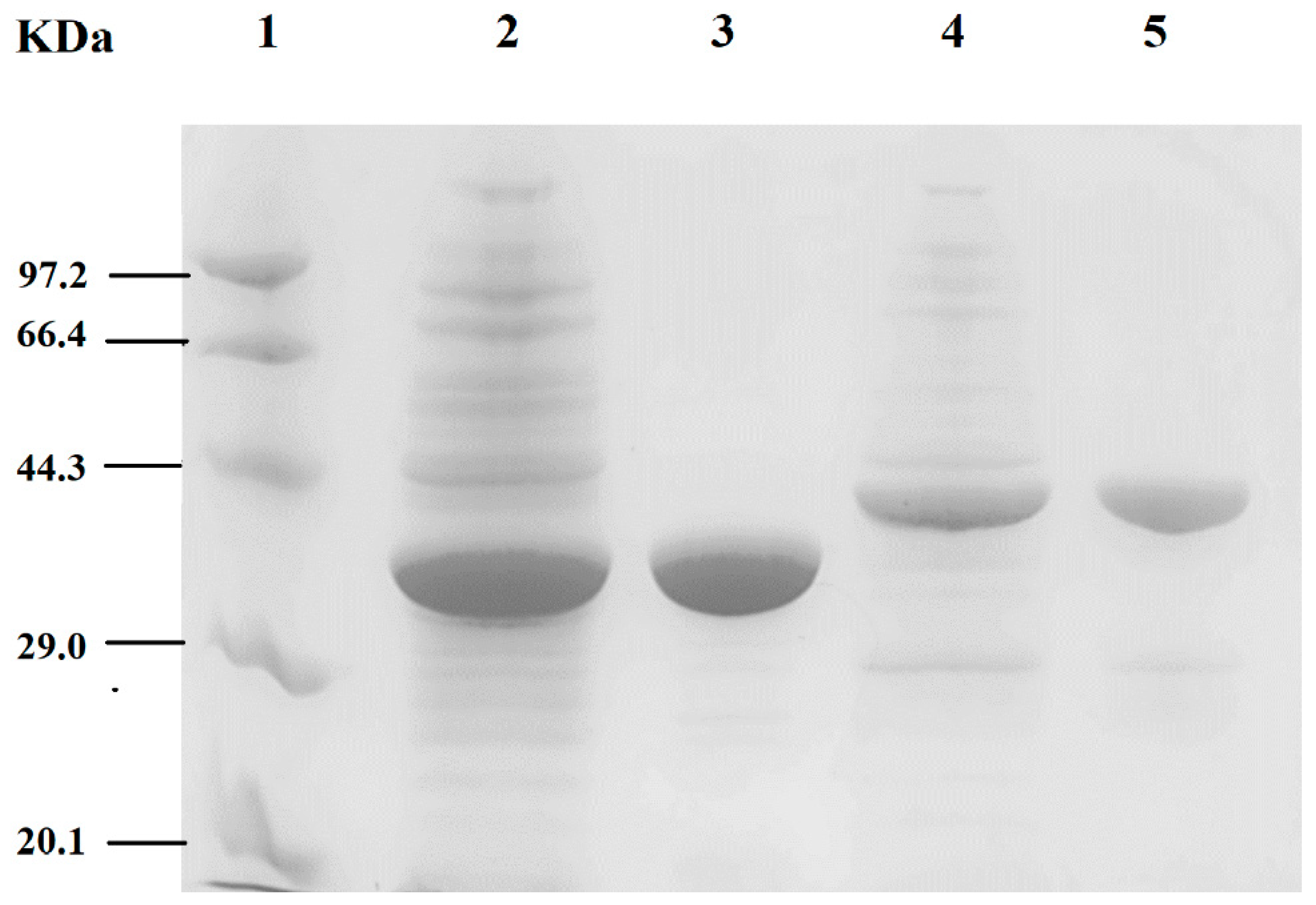
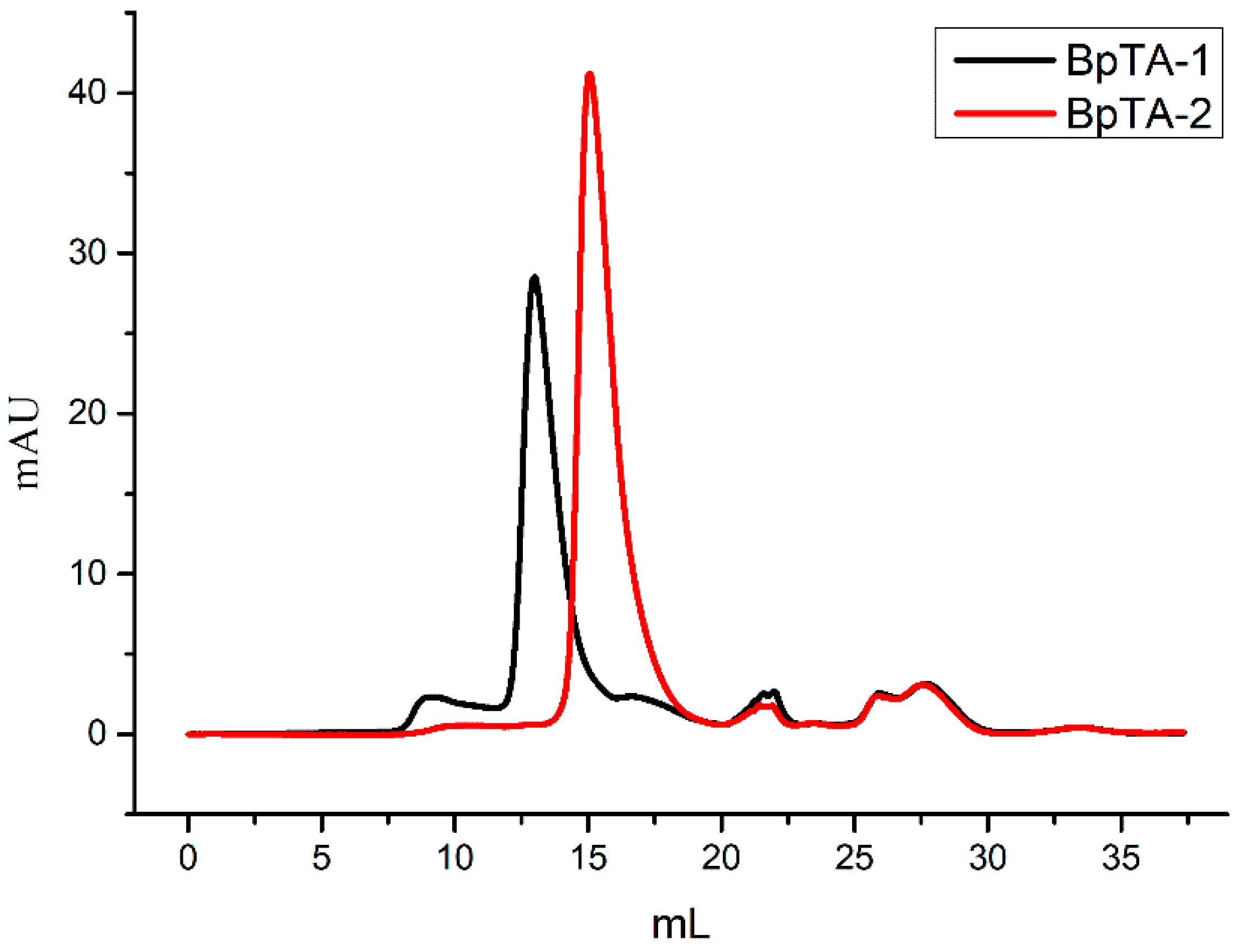
2.2. SDS-PAGE and Gel Filtration Chromatography Analyses
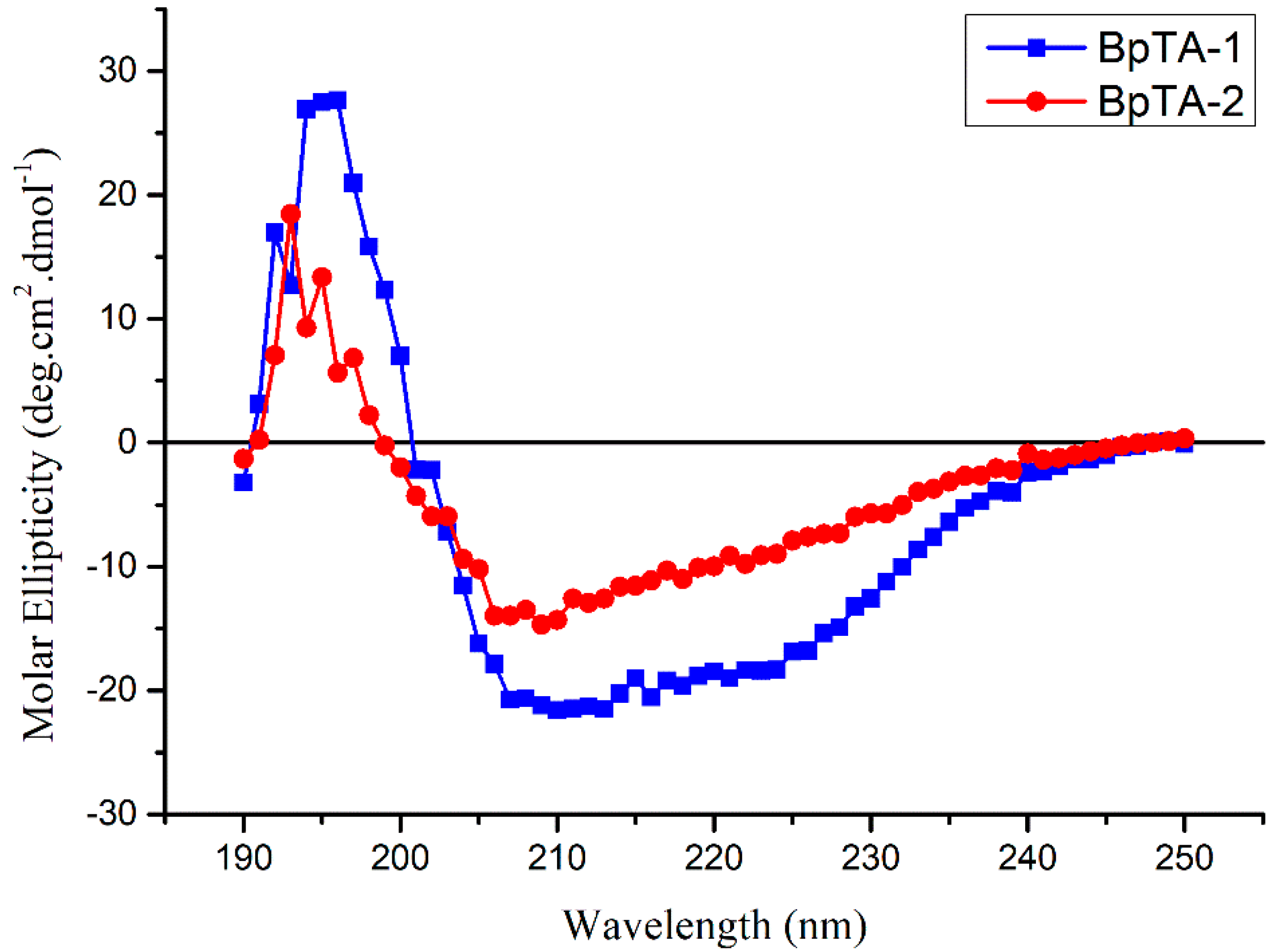
2.3. Secondary Structural Analysis
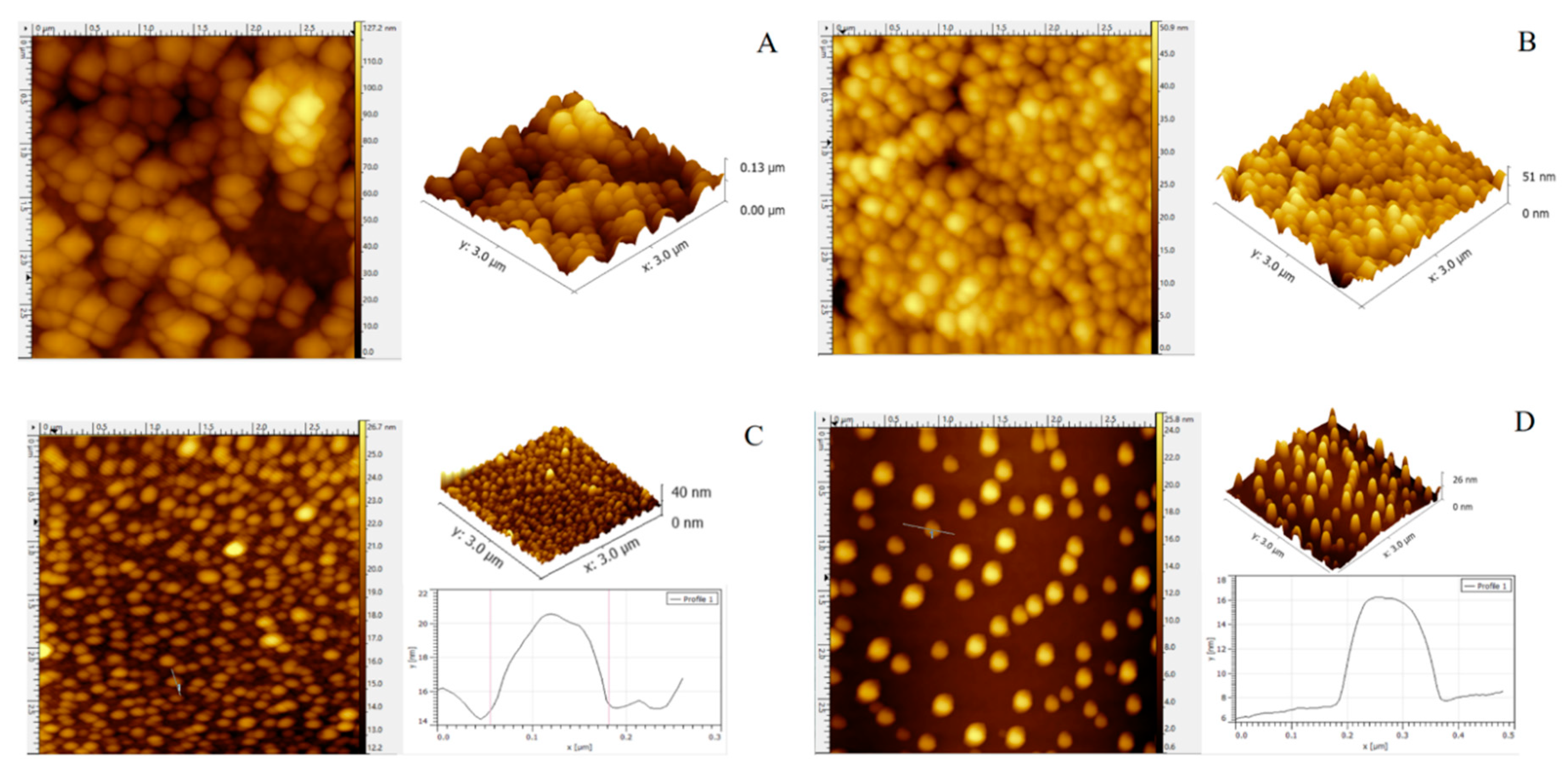
2.4. Atomic Force Microscopy Imaging
2.5. Enzyme Activity Analysis
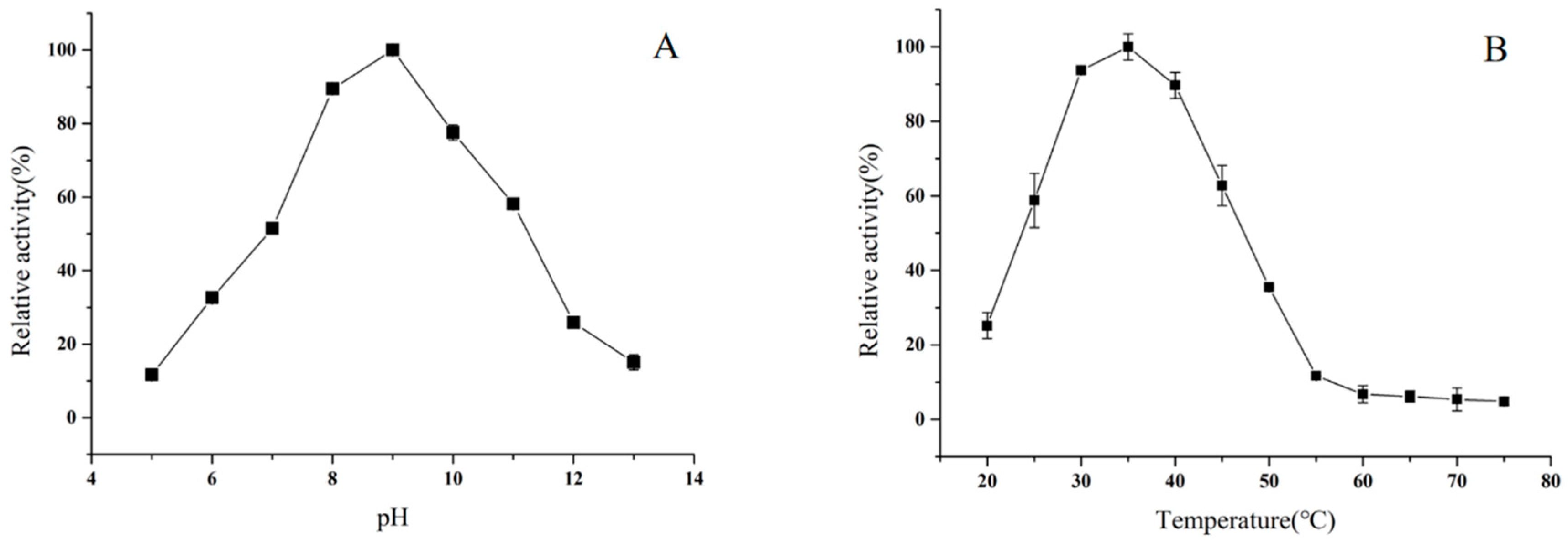
2.6. Effects of pH and Temperature on Enzyme Activity
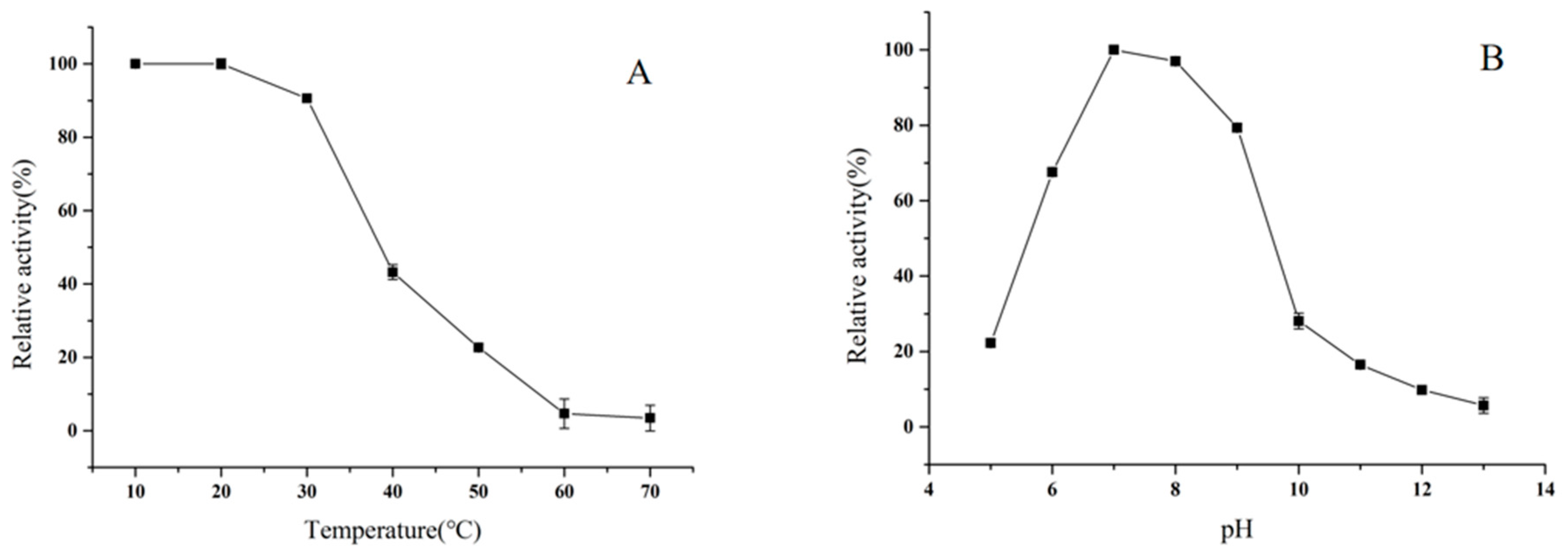
2.7. Effects of pH and Temperature on Enzyme Stability
3. Materials and Methods
3.1. Bioinformatic Analysis of the ota8 Gene and the BpTA-2 Protein
3.2. Materials
3.3. Gene Expression and Enzyme Purification
3.4. SDS-PAGE and Gel Filtration Chromatography
3.5. Circular Dichroism Spectroscopy
3.6. Atomic Force Microscopy
3.7. Enzyme Activity Assay
3.8. Effects of pH and Temperature on Enzyme Activity and Stability
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Schmidt, N.G.; Simon, R.C.; Kroutil, W. Biocatalytic asymmetric synthesis of optically pure aromatic propargylic amines employing ω-transaminases. Adv. Synth. Catal. 2015, 357, 1815–1821. [Google Scholar] [CrossRef]
- Svedendahl, M.; Branneby, C.; Lindberg, L.; Berglund, P. Reversed enantiopreference of an ω-transaminase by a single-point mutation. ChemCatChem 2010, 2, 976–980. [Google Scholar] [CrossRef]
- Wu, X.; Fei, M.; Chen, Y.; Wang, Z.; Chen, Y. Enzymatic synthesis of L-norephedrine by coupling recombinant pyruvate decarboxylase and ω-transaminase. Appl. Microbiol. Biotechnol. 2014, 98, 7399–7408. [Google Scholar] [CrossRef] [PubMed]
- Guo, F.; Berglund, P. Transaminase biocatalysis: Optimization and application. Green Chem. 2017, 19, 333–360. [Google Scholar] [CrossRef]
- Ghislieri, D.; Turner, N.J. Biocatalytic approaches to the synthesis of enantiomerically pure chiral amines. Top. Catal. 2014, 57, 284–300. [Google Scholar] [CrossRef]
- Kohls, H.; Steffen-Munsberg, F.; Höhne, M. Recent achievements in developing the biocatalytic toolbox for chiral amine synthesis. Curr. Opin. Chem. Biol. 2014, 19, 180–192. [Google Scholar] [CrossRef]
- Fuchs, M.; Farnberger, J.E.; Kroutil, W. The industrial age of biocatalytic transamination. Eur. J. Org. Chem. 2015, 32, 6965–6982. [Google Scholar] [CrossRef]
- Simon, R.C.; Richter, N.; Busto, E.; Kroutil, W. Recent developments of cascade reactions involving ω-transaminases. ACS Catal. 2014, 4, 129–143. [Google Scholar] [CrossRef]
- Narancic, T.; Almahboub, S.A.; O’Connor, K.E. Unnatural amino acids: Production and biotechnological potential. World J. Microb. Biotechnol. 2019, 35, 67. [Google Scholar] [CrossRef]
- Martin, A.R.; DiSanto, R.; Plotnikov, I.; Kamat, S.; Shonnard, D.; Pannuri, S. Improved activity and thermostability of (S)-aminotransferase by error-prone polymerase chain reaction for the production of a chiral amine. Biochem. Eng. J. 2007, 37, 246–255. [Google Scholar] [CrossRef]
- Citoler, J.; Derrington, S.R.; Galman, J.L.; Bevinakatti, H.; Turner, N.J. A biocatalytic cascade for the conversion of fatty acids to fatty amines. Green Chem. 2019, 21, 4932–4935. [Google Scholar] [CrossRef]
- Bezsudnova, E.Y.; Stekhanova, T.N.; Popinako, A.V.; Rakitina, T.V.; Nikolaeva, A.Y.; Boyko, K.M.; Popov, V.O. Diaminopelargonic acid transaminase from Psychrobacter cryohalolentis is active towards (S)-(-)-1-phenylethylamine, aldehydes and α-diketones. Appl. Microbiol. Biotechnol. 2018, 102, 9621–9633. [Google Scholar] [CrossRef] [PubMed]
- Ferrandi, E.E.; Monti, D. Amine transaminases in chiral amines synthesis: Recent advances and challenges. World J. Microbiol. Biotechnol. 2018, 34, 13. [Google Scholar] [CrossRef] [PubMed]
- Savile, C.K.; Janey, J.M.; Mundorff, E.C.; Moore, J.C.; Tam, S.; Jarvis, W.R.; Colbeck, J.C.; Krebber, A.; Fleitz, F.J.; Brands, J.; et al. Biocatalytic asymmetric synthesis of chiral amines from ketones applied to sitagliptin manufacture. Science 2010, 329, 305–309. [Google Scholar] [CrossRef] [PubMed]
- Zhai, L.; Yang, S.; Lai, Y.; Meng, D.; Tian, Q.; Guan, Z.; Cai, Y.; Liao, X. Effect of residue substitution via site-directed mutagenesis on activity and steroselectivity of transaminase BpTA from Bacillus pumilus W3 for sitafloxacin hydrate intermediate. Int. J. Biol. Macromol. 2019, 137, 732–740. [Google Scholar] [CrossRef] [PubMed]
- Markossian, K.A.; Golub, N.V.; Kleymenov, S.Y.; Muranov, K.O.; Sholukh, M.V.; Kurganov, B.I. Effect of α-crystallin on thermostability of mitochondrial aspartate aminotransferase. Int. J. Biol. Macromol. 2009, 44, 441–446. [Google Scholar] [CrossRef]
- Patil, M.D.; Grogan, G.; Bommarius, A.; Yun, H. Recent advances in ω-transaminase-mediated biocatalysis for the enantioselective synthesis of chiral amines. Catalysts 2018, 8, 254. [Google Scholar] [CrossRef]
- Molnár, Z.; Farkas, E.; Lakó, Á.; Erdélyi, B.; Kroutil, W.; Vértessy, B.G.; Paizs, C.; Poppe, L. Immobilized whole-cell transaminase biocatalysts for continuous-flow kinetic resolution of amines. Catalysts 2019, 9, 438. [Google Scholar] [CrossRef]
- Grishin, N.V.; Phillips, M.A.; Goldsmith, E.J. Modeling of the spatial structure of eukaryotic ornithine decarboxylases. Protein Sci. 1995, 4, 1291–1304. [Google Scholar] [CrossRef]
- Mueser, T.C.; Drago, V.; Kovalevsky, A.; Dajnowicz, S. Pyridoxal 5′-phosphate dependent reactions: Analyzing the mechanism of aspartate aminotransferase. Method Enzymol. 2020, 634, 333–359. [Google Scholar] [CrossRef]
- Schneider, G.; Käck, H.; Lindqvist, Y. The manifold of vitamin B6 dependent enzymes. Structure 2000, 8, R1–R6. [Google Scholar] [CrossRef]
- Steffen-Munsberg, F.; Vickers, C.; Kohls, H.; Land, H.; Mallin, H.; Nobili, A.; Skalden, L.; Bergh, T.; Joosten, H.; Berglund, P.; et al. Bioinformatic analysis of a PLP-dependent enzyme superfamily suitable for biocatalytic applications. Biotechnol. Adv. 2015, 33, 566–604. [Google Scholar] [CrossRef]
- Inoue, K.; Kuramitsu, S.; Aki, K.; Watanabe, Y.; Takagi, T.; Nishigai, M.; Ikai, A.; Kagamiyama, H. Branched-chain amino acid aminotransferase of Escherichia coli: Overproduction and properties1. J. Biochem. 1988, 104, 777–784. [Google Scholar] [CrossRef] [PubMed]
- Tanizawa, K.; Masu, Y.; Asano, S.; Tanaka, H.; Soda, K. Thermostable D-amino acid aminotransferase from a thermophilic Bacillus species. J. Biol. Chem. 1989, 264, 2445–2449. [Google Scholar]
- Iwasaki, A.; Matsumoto, K.; Hasegawa, J.; Yasohara, Y. A novel transaminase, (R)-amine:pyruvate aminotransferase, from Arthrobacter sp. KNK168 (FERM BP-5228): Purification, characterization, and gene cloning. Appl. Microbiol. Biotechnol. 2012, 93, 1563–1573. [Google Scholar] [CrossRef] [PubMed]
- Bezsudnova, E.Y.; Popov, V.O.; Boyko, K.M. Structural insight into the substrate specificity of PLP fold type IV transaminases. Appl. Microbiol. Biotechnol. 2020, 104, 2343–2357. [Google Scholar] [CrossRef]
- Mathew, S.; Nadarajan, S.P.; Chung, T.; Park, H.H.; Yun, H. Biochemical characterization of thermostable ω-transaminase from Sphaerobacter thermophilus and its application for producing aromatic β- and γ-amino acids. Enzym. Microb. Technol. 2016, 87–88, 52–60. [Google Scholar] [CrossRef]
- Kim, G.H.; Jeon, H.; Khobragade, T.P.; Patil, M.D.; Sung, S.; Yoon, S.; Won, Y.; Choi, I.S.; Yun, H. Enzymatic synthesis of sitagliptin intermediate using a novel ω-transaminase. Enzym. Microb. Technol. 2019, 120, 52–60. [Google Scholar] [CrossRef]
- Han, S.W.; Park, E.S.; Dong, J.Y.; Shin, J.S. Mechanism-guided engineering of ω-transaminase to accelerate reductive amination of ketones. Adv. Synth. Catal. 2015, 357, 1732–1740. [Google Scholar] [CrossRef]
- Scheidt, T.; Land, H.; Anderson, M.; Chen, Y.; Berglund, P.; Yi, D.; Fessner, W.D. Fluorescence-based kinetic assay for high-throughput discovery and engineering of stereoselective ω-transaminases. Adv. Synth. Catal. 2015, 357, 1721–1731. [Google Scholar] [CrossRef]
- Hohne, M.; Schatzle, S.; Jochens, H.; Robins, K.; Bornscheuer, U.T. Rational assignment of key motifs for function guides in silico enzyme identification. Nat. Chem. Biol. 2010, 6, 807–813. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Chen, X.; Zhang, D.; Wu, Q.; Zhu, D. Characterization of (R)-selective amine transaminases identified by in silico motif sequence blast. Appl. Microbiol. Biotechnol. 2015, 99, 2613–2621. [Google Scholar] [CrossRef] [PubMed]
- Sayer, C.; Martinez-Torres, R.J.; Richter, N.; Isupov, M.N.; Hailes, H.C.; Littlechild, J.A.; Ward, J.M. The substrate specificity, enantioselectivity and structure of the (R)-selective amine: Pyruvate transaminase from Nectria haematococca. FEBS J. 2014, 281, 2240–2253. [Google Scholar] [CrossRef] [PubMed]
- Zhai, L.; Ren, R.; Meng, D.; Tian, Q.; Guan, Z.; Cai, Y.; Liao, X. Comparison of aminotransferases of three Bacillus strains Bacillus altitudinis W3, Bacillus velezensis SYBC H47, and Bacillus amyloliquefaciens YP6 via genome analysis and bioinformatics. J. Appl. Genet. 2019, 60, 427–430. [Google Scholar] [CrossRef]
- Zhai, L.; Yang, S.; Lai, Y.; Meng, D.; Tian, Q.; Guan, Z.; Cai, Y.; Liao, X. Mining of aminotransferase gene ota3 from Bacillus pumilus W3 via genome analysis, gene cloning and expressing for compound bioamination. Gene 2019, 686, 21–28. [Google Scholar] [CrossRef]
- Gao, S.; Su, Y.; Zhao, L.; Li, G.; Zheng, G. Characterization of a (R)-selective amine transaminase from Fusarium oxysporum. Process Biochem. 2017, 63, 130–136. [Google Scholar] [CrossRef]
- Bezsudnova, E.Y.; Dibrova, D.V.; Nikolaeva, A.Y.; Rakitina, T.V.; Popov, V.O. Identification of branched-chain amino acid aminotransferases active towards (R)-(+)-1-phenylethylamine among PLP fold type IV transaminases. J. Biotechnol. 2018, 271, 26–28. [Google Scholar] [CrossRef]
- Cerioli, L.; Planchestainer, M.; Cassidy, J.; Tessaro, D.; Paradisi, F. Characterization of a novel amine transaminase from Halomonas Elongate. J. Mol. Catal. B Enzym. 2015, 120, 141–150. [Google Scholar] [CrossRef]
- Kaulmann, U.; Smithies, K.; Smith, M.E.; Hailes, H.C.; Ward, J.M. Substrate spectrum of ω-transaminase from Chromobacterium violaceum DSM30191 and its potential for biocatalysis. Enzyme Microb. Technol. 2007, 41, 628–637. [Google Scholar] [CrossRef]
- Shin, J.S.; Yun, H.; Jang, J.W.; Park, I.; Kim, B.G. Purification, characterization, and molecular cloning of a novel amine: Pyruvate transaminase from Vibrio fluvialis JS17. Appl. Microbiol. Biotechnol. 2003, 61, 463–471. [Google Scholar] [CrossRef]
- Guan, Z.; Cai, Y.; Zhang, Y.; Zhao, H.; Liao, X. Complete genome sequence of Bacillus pumilus W3: A strain exhibiting high laccase activity. J. Biotechnol. 2015, 207, 8–9. [Google Scholar] [CrossRef] [PubMed]
- Crooks, G.E.; Hon, G.; Chandonia, J.M.; Brenner, S.E. WebLogo: A sequence logo generator. Genome Res. 2004, 14, 1188–1190. [Google Scholar] [CrossRef] [PubMed]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Meng, D.; Tian, Q.P.; Yang, S.L.; Deng, H.X.; Guan, Z.B.; Cai, Y.J.; Liao, X.R. Characterization of a novel carboxylesterase from Bacillus velezensis SYBC H47 and its application in degradation of phthalate esters. J. Biosci. Bioeng. 2020, 129, 588–594. [Google Scholar] [CrossRef] [PubMed]
- Miles, A.J.; Wallace, B.A. CDtoolX, a Downloadable Software Package for Processing and Analyses of Circular Dichroism Spectroscopic Data. Protein Sci. 2018, 27, 1717–1722. [Google Scholar] [CrossRef] [PubMed]
- Whitmore, L.; Wallace, B.A. Protein Secondary Structure Analyses from Circular Dichroism Spectroscopy: Methods and Reference Databases. Biopolymers 2008, 89, 392–400. [Google Scholar] [CrossRef]
- Whitmore, L.; Wallace, B.A. DICHROWEB, an online server for protein secondary structure analyses from circular dichroism spectroscopic data. Nucleic Acids Res. 2004, 32, 668–673. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | (R)-α-Phenethylamine | (S)-α-Phenethylamine |
|---|---|---|
| Activity (U/mg) a | Activity (U/mg) a | |
| Purified BpTA-2 | 2.1207 ± 0.20 | 0.0011 ± 0.19 |
| Purified BpTA-1 [35] | 1.1760 ± 0.19 | 0.0005 ± 0.15 |
| ATFo [36] | 1.3920 | 0.0004 |
| T. terrenum Tam [37] | 0.15 | n.d. |
| H. ochraceum Tam [37] | 0.10 | n.d. |
| HEWT [38] | <0.0705 | 1.4946 |
| C. violaceum ω-Tam [39] | <0.0564 | 0.97854 |
| V. fluvialis ω-Tam [40] | <0.0141 | 1.5792 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhai, L.; Xie, Z.; Tian, Q.; Guan, Z.; Cai, Y.; Liao, X. Structural and Functional Analysis of the Only Two Pyridoxal 5′-Phosphate-Dependent Fold Type IV Transaminases in Bacillus altitudinis W3. Catalysts 2020, 10, 1308. https://doi.org/10.3390/catal10111308
Zhai L, Xie Z, Tian Q, Guan Z, Cai Y, Liao X. Structural and Functional Analysis of the Only Two Pyridoxal 5′-Phosphate-Dependent Fold Type IV Transaminases in Bacillus altitudinis W3. Catalysts. 2020; 10(11):1308. https://doi.org/10.3390/catal10111308
Chicago/Turabian StyleZhai, Lixin, Zihao Xie, Qiaopeng Tian, Zhengbing Guan, Yujie Cai, and Xiangru Liao. 2020. "Structural and Functional Analysis of the Only Two Pyridoxal 5′-Phosphate-Dependent Fold Type IV Transaminases in Bacillus altitudinis W3" Catalysts 10, no. 11: 1308. https://doi.org/10.3390/catal10111308
APA StyleZhai, L., Xie, Z., Tian, Q., Guan, Z., Cai, Y., & Liao, X. (2020). Structural and Functional Analysis of the Only Two Pyridoxal 5′-Phosphate-Dependent Fold Type IV Transaminases in Bacillus altitudinis W3. Catalysts, 10(11), 1308. https://doi.org/10.3390/catal10111308