1. Introduction
Industrial development is associated with an increasing energy demand. This concern affects all branches of industry, with transportation ranked at the top. Growing production of cars comes with further higher fuel consumption. Unavoidable depletion of fossil fuels, which are commonly used as main energy sources, poses both economic and ecological hazards. A great alternative to fossil fuels is offered by biofuels [
1]. Biofuels, like bioethanol, are produced from crops which are renewable sources; however, growing crops especially for fuel production may cause further practical problems due to competition between biofuel and food crops in the cultivation area. A solution to this problem is the production of second-generation bioethanol (2G) [
1] from cellulose-rich plant biomass, predominantly agriculture wastes, which is connected with significantly reduced costs. One of the substrates available for 2G bioethanol production is sweet sorghum bagasse (SSB), a lignocellulosic material left after the sucrose-rich juice has been extracted from the plants [
2]. Sweet sorghum itself is a plant able to grow in fifth class soil due to its deep root system and resistance to drought as well as many plant diseases and pests. These features make sorghum the fifth most cultivated crop in the world, resulting in SSB being an inexpensive fermentation substrate source [
3,
4,
5]. Ethanol produced from sweet sorghum is safer for the environment due to low sulfur content, low biological and chemical oxygen demand, and a high octane rating. While annual ethanol output from sweet sorghum depends on several factors, including genetic background, season, soil quality, and other environmental factors, the annual ethanol production from sweet sorghum crop is estimated at 8000 L/ha [
6].
Obtaining a suitable substrate for the production of 2G bioethanol is merely the initial step; however, a further major concern is saccharification of cellulose, since fermenting yeasts are unable to use sugar polymers. Substrates used in this process (i.e., SSB) need to be chemically/physically pretreated and then enzymatically hydrolyzed prior to the fermentation step. An initial pretreatment is essential to make raw materials accessible to the hydrolyzing enzymes [
7]. This step involves mostly acidic or alkaline treatment of the substrate, although such methods as microwave radiation, sonification, spray drying, gamma radiation, and pyrolysis are employed as well [
8]. The second step of treatment, widely covered in this article, includes enzymatic hydrolysis, typically performed with the use of commercial enzymatic preparations. Cellulase preparations available on the market derive from different microorganisms, such as:
Aspergillus sp.,
Trichoderma sp. (
T. longibrachiatum,
T. reesei,
T. viride, and
T. harzianum),
Humicola sp. (
H. insolens and
H. griseathermoidea), or
Bacillus sp. [
9]. These preparations are costly, for example, the price of CTec2 according to the data of one of the largest distributors of enzyme preparations is €120 per 50 mL (Sigma-Aldrich, St. Louis, MO, USA). This article introduces a new enzymatic preparation from
Trichoderma citrinoviride C1, produced in a less complex and low-cost process involving two simple steps, as a promising alternative to commercial counterparts.
The efficiency of a cellulase preparation, whether commercial or not, depends on such process parameters as pH, temperature, and substrate concentration. To achieve its peak efficiency, the saccharification process conditions require optimization. The objective of this study was to produce a new cellulase preparation but also to optimize hydrolysis parameters of sweet sorghum waste biomass (pH, temperature, and substrate concentration), as well as to assess their usefulness in the process of substrate pretreatment for ethanol fermentation. The efficiency results obtained in this research were compared to those acquired with a commercially viable enzyme for cellulosic ethanol production (Cellic® CTec2) to establish applicability of the enzymatic preparation produced in the study.
2. Results
Biosynthesis of cellulolytic and xylanolytic enzymes was performed in 10-day solid state fermentation of
T. citrinoviride C1 with confirmed capability for extracellular synthesis of these hydrolases, in culture medium based on a mixture of wheat bran and sweet beet pulp. The enzymatic activity determined in the post-culture liquid reached 2.37 U mL
−1 and 6.52 U mL
−1 for cellulases and xylanases, respectively (
Table 1).
The ultrafiltration step resulted in a more than three-fold increase in enzymatic activity per volume unit. The concentrated preparation exhibited the activity of cellulases (8.99 U mL
−1) and xylanases (21.26 U mL
−1), whereas the enzymatic protein recovery yield was not lower than 80% and the degree of purification was not higher than 1.6 (
Table 1).
The low degree of purification was due to the single-stage purification process; however, the determined enzymatic activity of the preparation was at a satisfactory level, which allowed to use it for the enzymatic hydrolysis of sorghum waste biomass. In the commercial enzyme cellulolytic activity of 185.42 U mL−1 was determined; however, both preparations were applied for the process of hydrolysis at similar doses of 2.15 U of endocellulases (CMCases) per 10 g of substrate.
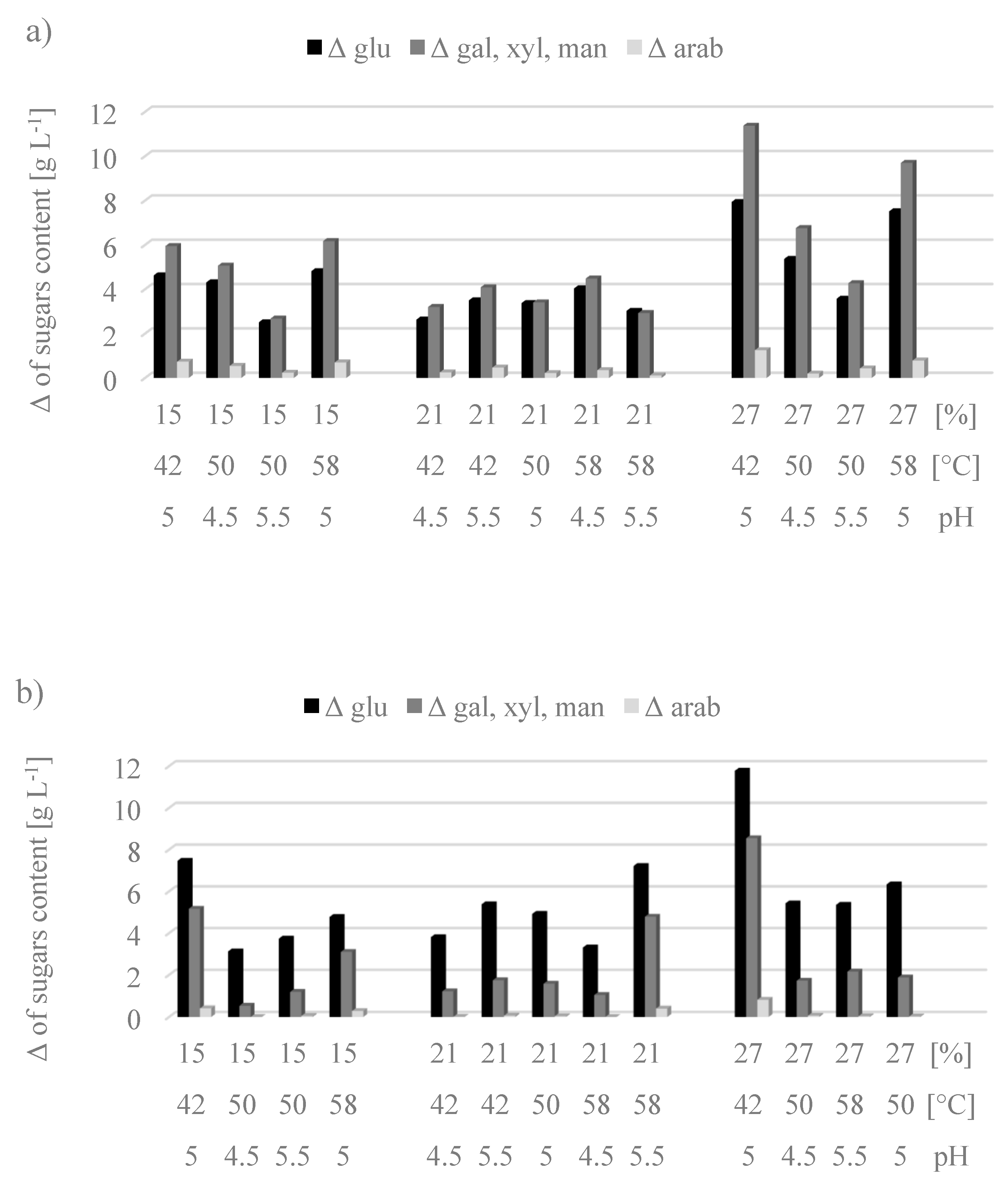
The increase in sugar concentration (Δ) after the enzymatic hydrolysis process in relation to their amount after the applied prehydrolysis process is presented in
Figure 1. Enzymatic saccharification was carried out under various conditions (substrate concentration, temperature, and pH) with the synthesized enzyme C1 (a) and the commercial enzyme preparation CTec2 (b). Concentrations of a mixture of galactose + xylose + mannose and of arabinose were higher in the hydrolysates made with C1 preparation, compared to the commercial CTec2 preparation, except for the variant with the medium concentration of the substrate (21%) hydrolyzed at 58 °C and pH 5.5. In turn, in hydrolysates produced with the CTec2 preparation higher concentrations of glucose were obtained as a result of dominant cellulolytic activity. This had a direct impact on the efficiency of the hydrolysis process calculated in relation to glucose, and in the experiment involving CTec2 preparation, its value ranged from 9.25% (run 10) to 16.93% (run 1) (data not shown).
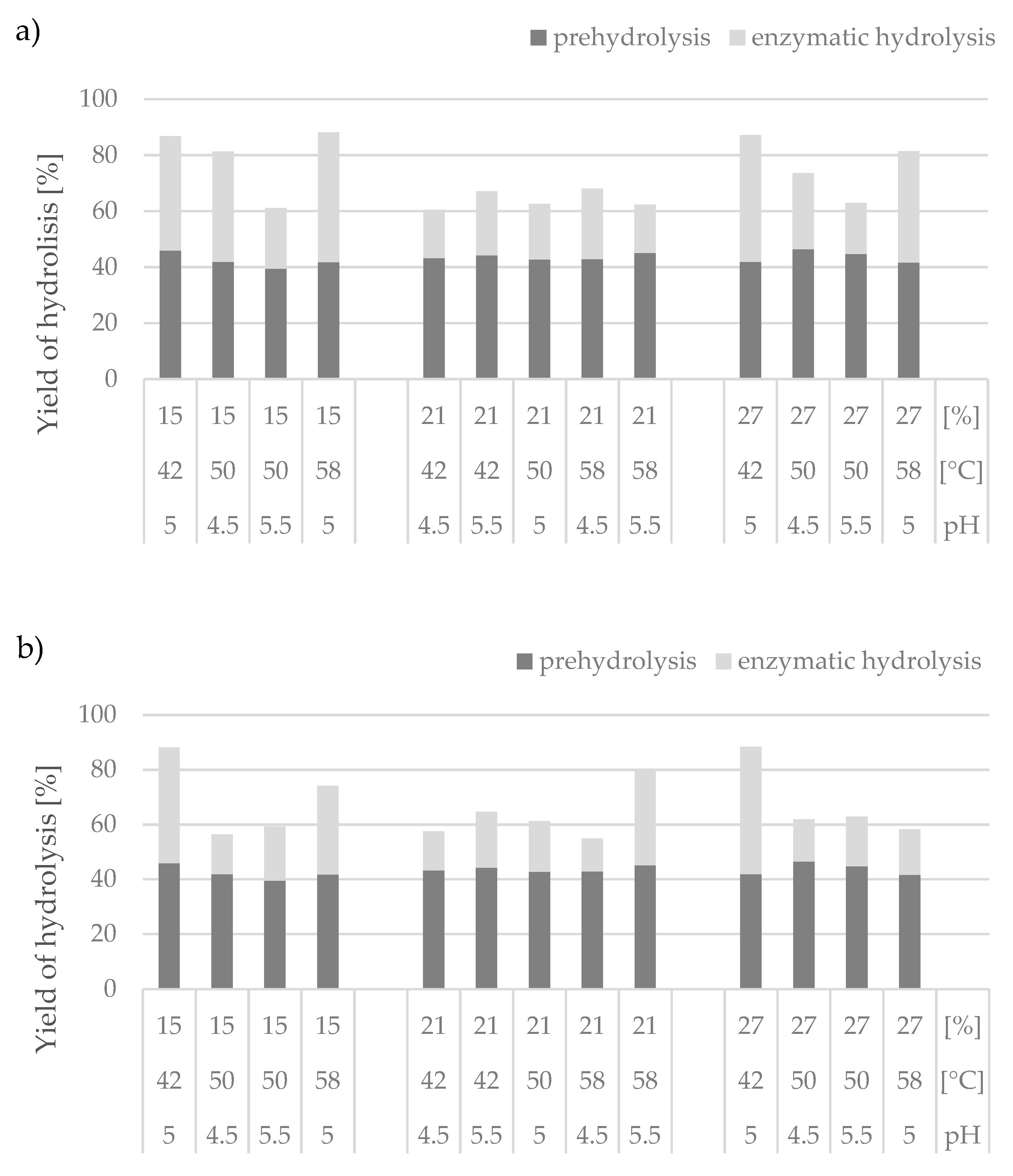
Considering the effectiveness of enzymatic hydrolysis performed with the preparation obtained under laboratory conditions, the variants with the maximum analyzed concentration of the substrate at pH 5.0 were the most advantageous. The sugars were released to hydrolysates as a result of prehydrolysis and enzymatic hydrolysis processes. The efficiency of the first step ranged from 39 to 46%. In turn, the efficiency of the enzymatic hydrolysis processes depended mainly on the process conditions, but the preparation used had no influence here because the efficiency in both processes ranged from 17% to 46% (
Figure 2).
Employing separate enzymatic hydrolysis and fermentation in ethanol production from lignocellulosic materials enables better adjustment of process conditions to these, conditions optimal for the effective action of both enzymatic preparations and
Saccharomyces cerevisiae yeast, compared to the processes involving simultaneous hydrolysis and fermentation.
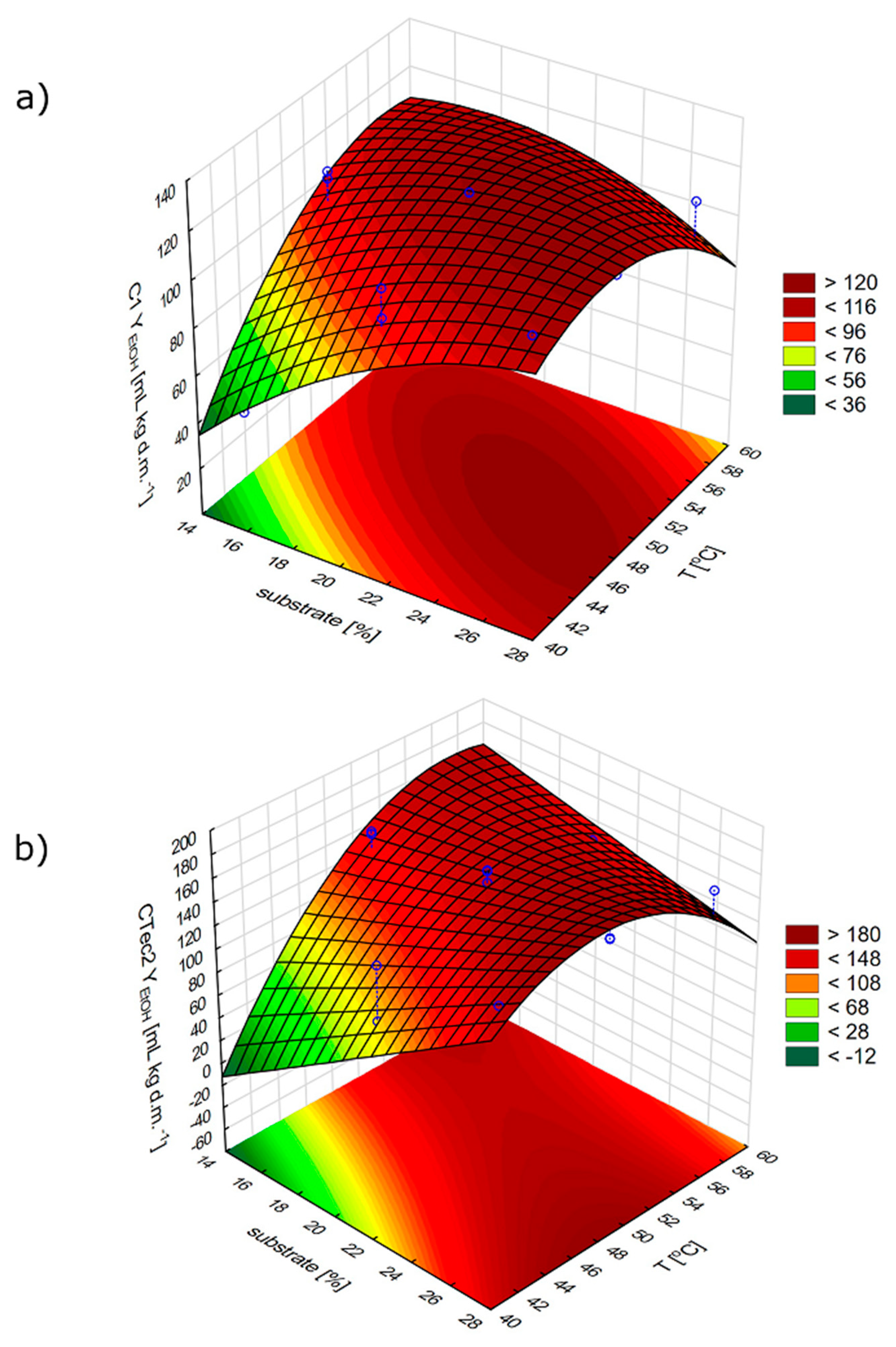
Figure 3 presents response surface models depicting the effect of substrate concentration and temperature on the yield of ethanol fermentation performed with C1 and CTec2 enzymes at the stage of substrate pretreatment.
Statistically significant effects of substrate concentration and process temperature were demonstrated, whereas there was no significant impact of pH value change, expressed both in the linear and quadratic effect, on the yield of ethanol fermentation process.
Ethanol yield after completed fermentation of the hydrolysates obtained with C1 and CTec2 preparations, depending on the experience option, ranged from 40.2 to 122.5 and from 3.5 to 165.3 mL kg d.m.
−1, respectively (
Table 2). Taking into account that the yeasts used in the ethanol fermentation process from the total sugar content in hydrolysates were only able to use hexoses, the theoretical efficiency of the process was relatively high (
Table 2), up to approximately 57% when using C1 and up to 76% for CTec2 preparation.
In terms of sugars consumed, fermentation of hydrolysates obtained with the C1 preparation allowed to obtain 66 to 280 mL of ethanol, while fermentation of CTec2 hydrolysates, from 6 to 377 mL of ethanol, calculated per kg of sugars (
Figure S1). Regardless of the enzymatic preparation, the highest bioethanol yields were achieved at substrate concentration of 21% and hydrolysis conducted at a temperature of 50 °C and pH 5.0.
The course of prediction profiles plotted for the proposed experimental model (
Figure S2) enabled the establishment of the optimal process conditions. The determined conditions for the application of the C1 preparation were as follows: Substrate concentration 22.6%, temperature 50.5 °C, and pH 5.1; they allowed achieving ethanol yield at 122.5 mL kg d.m.
−1. In the case of the commercial CTec2 preparation, the respective conditions were: 27%, 52.7 °C, and pH 5.2, and predicted ethanol yield was at 170.8 mL kg d.m.
−1.
Both obtained regression models were analyzed with ANOVA to verify their correctness. Significance of the models was confirmed by achieved F-values and adherence of models to the measured data was described by coefficients of determination (R
2). For the process with C1 enzyme preparation F-value was 5.01 and R
2 0.7358. For the hydrolysis with CTec2 F-value reached 7.83 and R
2 0.8132 (
Table S1). Furthermore, the “lack of fit” tests were insignificant for both models, which further proved their correctness.
The empirical relationship between predicted ethanol yield and the variables substrate concentration and temperature was expressed as a polynomial equation:
and
3. Discussion
Reduction in the amount of generated wastes is contemporarily an urgent goal to achieve. Some waste materials may be successfully used as substrates in biotechnological processes. This is the case with, e.g., lignocellulosic waste biomass produced in large volumes as a result of agricultural activity or as a waste product in food processing, thus posing a great problem to the natural environment protection due to a lack of its management strategies. Part of the waste biomass is spent as feedstuff; however, its vast amounts significantly surpass the needs in this respect. On the other hand, this waste material offers an excellent source of carbon and energy to microorganisms, which has been exploited for years. One of the waste biomass management sectors may be production of hydrolytic enzymes and second-generation bioethanol. Due to their composition, such lignocellulosic waste materials as vegetable and fruit peels, fruit pomace, sugar cane, pomace from oil production, wheat straw, wheat bran, and sugar beet pulp are often utilized for cultivation of bacteria and filamentous fungi aimed at the synthesis of cellulases, xylanases, lipases, and pectinases [
10,
11,
12,
13,
14]. A tendency has been recently observed for the use of these waste materials in solid media cultures, which appeared to be superior in terms of cost-effectiveness, environment-friendly character, and yield compared to submerged cultures [
15,
16,
17].
Lignocellulosic waste materials, as an inexpensive and readily available source of carbon and energy, are commonly used as substrates in the production of enzymes, in particular cellulolytic and xylanolytic. In the present study, a mixture of sugar beet pulp and wheat bran was used for the biosynthesis of cellulolytic and xylanolytic enzymes in a solid state culture, which allowed achieving enzymatic activity at 21.5 U g
−1 and 59.27 U g
−1 medium, respectively. Using the solid medium composed of sugar beet pulp only and the same strain of filamentous fungi, Piegza et al. [
18] obtained cellulolytic activity at 6.0 U g
−1, which may indicate that a mixture of substrates is a better inducer of cellulolytic enzymes compared to a single substrate. Similar observations were made by Mala et al. [
19], who proved that the use of two substrates for lipases production enhanced their biosynthesis by 36%.
From among other carbon sources (wheat bran, rice straw, sugar cane pulp, wheat straw) used by Idris et al. [
20], wheat bran turned out to be the most advantageous inducer of cellulase production by
Trichoderma reesei RUT C-30, which allowed for an exceptionally high activity of 298.4 U g
−1. The selection of the waste material for the process of biosynthesis is dependent on its availability in a specific country; hence, in this study a mixture of beet pulp and wheat bran was used, which represent raw materials commonly found in Poland as food industry by-products. Currently, numerous publications are available on the application of microorganisms in the biosynthesis of cellulases and xylanases. To summarize their contribution, we can conclude that most common producers are saprophytes isolated primarily from soil, organic residues found in forests and composts, which are classified mainly among bacteria, actinomycetes and filamentous fungi [
21,
22]. In addition, the white-rot basidiomycetes, or Shiitake medicinal mushroom
Lentinus edodes (higher Basidiomycetes) are also known as cellulase producers [
23,
24].
In scientific works, the most frequently discussed are issues related to the pretreatment of lignocellulose, the use of various carbon sources (hexoses, pentoses) and various techniques (separate hydrolysis and fermentation, simultaneous saccharification and fermentation, simultaneous saccharification and co-fermentation, and consolidated bioprocessing) promoting high ethanol yield and cost-efficiency of the entire bioprocess [
25].
In addition, the enzymatic preparation used in the production of second-generation (2G) ethanol is a significant determinant of the yield and profitability of the entire process. Many commercial preparations are available on the market; however, their excessive cost prevents their industrial-scale application. This urges the need for developing a technology that would exploit an inexpensive preparation, often produced with filamentous fungi
Trichoderma sp. Some authors have suggested that when pure glucose was used to produce the enzymatic preparation, its price would account for even 50% of preparation production cost [
26]. The C1 preparation was produced using lignocellulosic waste materials, which substantially increases its attractiveness. Despite a single-stage ultrafiltration process, the enzymatic activity of the preparation did not differ significantly from results reported by other authors [
27,
28]. Using a membrane with the cut-off point of 10 kDa in the concentration process enabled retention of (hemi)cellulolytic proteins in the concentrated syrup. The use of such a membrane in our study allowed for over 80% recovery of determined enzymes. A marginally lower recovery percentage was obtained by Qi et al. [
29] who managed to retain 78% of proteins in the preparation after the ultrafiltration process. Mores et al. [
30], who also used a 10 kDa cut-off membrane, confirmed that it allowed for an effective recovery of cellulolytic enzymes. These examples demonstrate the right choice of this type of membrane for the production of a concentrated preparation.
Substrate pretreatment, involving breakdown of the compact structure of lignocellulose, is known to be a crucial stage in 2G bioethanol production, determining the effectiveness of the entire process. In the above study, likewise in research of Głąb et al. [
31], the substrate pretreatment involved the acidic pre-hydrolysis. In turn, Cao et al. [
32] compared various methods for sorghum biomass pre-hydrolysis and achieved the best result upon the coupled use of a diluted sodium hydroxide solution and hot hydrogen peroxide. However, the application of multiple chemical reagents has an adverse effect on the natural environment and, therefore, the least interfering methods should be looked after. It is also worthy of notice that the outcomes of hydrolysis and fermentation are affected by inhibitors produced during substrate pretreatment. Treating the lignocellulosic substrate with acid causes primarily hemicellulose degradation to monosaccharides, but additionally induces synthesis of such compounds as phenols, furfural or 5-hydroxymethylfurfural, which may inhibit activity of enzymes and microorganisms in the fermentation process. Teixeira et al. [
33] analyzed the composition of inhibitors, i.e., furfural, 5-hydroxymethylfurfural (HMF) and acetic acid, in acidic, basic, and enzymatic hydrolysates of sugar cane pulp. The highest concentrations of HMF and furfural were determined in acidic hydrolysates obtained with 7% sulfuric acid (VI). These authors concluded that the accumulation of both these compounds increased proportionally with the increasing acid concentration. In turn, the enzymatic hydrolysates analyzed in their study had a high concentration of acetic acid. In recent years, various authors have proposed natural pretreatment methods which—when optimized—could offer an alternative to the chemical methods [
34,
35].
Undoubtedly, a key element for reaching a high bioethanol yield from plant waste is the stage of enzymatic hydrolysis. Its conditions should be adjusted individually to the specific substrate. Performing enzymatic hydrolysis with a specified dose of endocellulases allowed for comparing the effectiveness of C1 preparation with that of the commercial CTec2 preparation. This comparative analysis demonstrated a diverse effect of both preparations on process yield due to the presence of accompanying enzymes.
Ethanol yields obtained during fermentation of the hydrolysates produced with the laboratory and commercial enzymatic preparation were consistent with those reported in literature, even though conditions of the fermentation process had not been optimized. Employing acidic pre-hydrolysis of sorghum waste biomass, Gibbons et al. [
36] achieved ethanol yield at 141 g kg
−1 dry sorghum (178.5 mL kg
−1 dry sorghum). Kim and Day [
37] produced 97 g (122.8 mL) of ethanol from one kg of sweet sorghum dry matter. In turn, Salvi et al. [
38] achieved from 210 to 250 g (from 265.8 to 316.4 mL) of ethanol kg
−1 dry sorghum biomass using a diluted ammonia pretreatment (28%
v/
v ammonium hydroxide at 160 °C for 1 h) and enzymatic hydrolysis (Spezyme CP and Novozyme 188). Furthermore, impregnation of sweet sorghum bagasse 2.5% SO
2 at a temperature of 200 °C for 7.5 min coupled with enzymatic hydrolysis (Spezyme CP and Novozyme 188) resulted in ethanol yield of 153 g (193.7 mL) from kg of substrate dry matter [
39].
Future investigations should attempt to simplify the technological process and to minimize the inhibitory effect of hydrolysis products, accumulated in the medium through the simultaneous saccharification and fermentation (SSF) or semi-simultaneous saccharification and fermentation (SSSF) [
40] with the use of the enzymatic preparation obtained under laboratory conditions in our study. In turn, conditions of the fermentation process could be improved by supplementing fermentation media with components that demonstrate a positive impact on yeast condition [
41,
42]. In addition, due to the synthesis of pentoses and significant amounts of xylose upon hemicelluloses hydrolysis, it would seem advisable to make use of unconventional microorganisms capable of ethanol fermentation of pentoses.
4. Materials and Methods
4.1. Microorganisms
The enzymatic preparation used in the study was produced using Trichoderma citrinoviride C1 fungus strain with confirmed capability to produce cellulolytic and xylanolytic enzymes. The strain originated from the local culture collection of the Department of Biotechnology and Food Microbiology of the Wroclaw University of Environmental and Life Sciences (Wroclaw, Poland), stored in a PDA (potato dextrose agar; Merck Millipore, Burlington, MA, USA) medium at 4 °C.
The process of ethanol fermentation was performed using dried culture of industrial distillery yeast Saccharomyces cerevisiae SIHA Active Yeast 6 strain DF 639 (EATON, Irvine, CA, USA) stored as recommended.
4.2. Waste Biomass of Sweet Sorghum
The experimental material included sweet sorghum (Sorghum bicolor (L.) Moench) of hybrid type Sucrosorgo 506, grown at the Research and Experimental Station of the Wrocław University of Environmental and Life Sciences (Pawłowice, Poland), characterized by 23.95% of dry matter, of which 41.49% was cellulose and 21.61% hemicellulose.
4.3. Production of the Enzymatic Preparation
As an inducer of cellulo- and xylanolytic enzymes, two waste raw materials were used: Sugar beet pulp (Sugar mill, Wałbrzych, Poland) and wheat bran (Department of Fermentation and Cereals Technology, Wrocław University of Environmental and Life Sciences). A sterile culture medium composed of sugar beet pulp and wheat bran (1:1) was inoculated with a suspension of T. citrinoviride C1 strain spores to the initial density of 5 × 106 spores g−1 medium. The solid state culture of 70% initial moisture was run at a temperature of 25 °C for 10 days under 24 h lighting in a KBWF 720 growth chamber (Binder GmbH, Tuttlingen, Germany). It was terminated by extraction using 0.05 M acetate buffer with pH 4.8 and agitated at room temperature for 30 min (160 rpm).
The enzymatic preparation (C1) was obtained by a four-fold concentration of the post-culture extract via ultrafiltration at a temperature of 4 °C. Ultrafiltration was performed using the Labscale TFF System (Merck Millipore, Burlington, MA, USA) with a Pellicon XL Ultrafiltration Module Biomax (polyethylsulfone membrane) with the cut-off point of 10 kDa.
4.4. Acidic Pre-Hydrolysis of Sweet Sorghum Biomass
Disintegrated waste biomass of sweet sorghum was subjected to high-temperature acidic pre-hydrolysis (2% sulfuric acid (VI), 150 °C, 1 h, WTB Binder dryer, Binder GmbH, Tuttlingen, Germany). The waste material was used in various concentrations (15, 21, and 27%
w/
w) according to the adopted optimization model (see section Results,
Table 2). The pre-hydrolysates were cooled, their pH was adjusted to values of 4.5, 5.0, and 5.5, using 1M solution of sodium hydroxide, and used in the process of enzymatic hydrolysis.
4.5. Optimization of the Enzymatic Hydrolysis of Sweet Sorghum Waste Biomass
Enzymatic hydrolysis was conducted in Erlenmayer flasks (100 mL) incubated in a GFL 1083 water bath (Bionovo, Legnica, Poland) at temperatures of 42 °C, 50 °C, and 58 °C at 120 rpm for 48 h (see section Results,
Table 2) using the concentrated preparation C1 in a dose corresponding to 2.15 U of endocellulases (CMCases) per 10 g of substrate. To determine the effectiveness of the enzymatic preparation obtained under laboratory conditions, a commercial enzymatic preparation Cellic
® CTec2 (further referred to as CTec2) (Novozymes, Kobenhavn, Denmark) was used for hydrolysis under analogous conditions.
The response surface methodology (RSM) was used to optimize conditions of enzymatic hydrolysis and to evaluate its usability for the process of ethanol fermentation. The RSM enables determining the main effects and interactions between selected independent variables (substrate concentration X
1, temperature X
2, and pH X
3) on the yield of ethanol fermentation process (dependent variable). Three levels (−1, 0, +1) were adopted in the Box–Behnken design for each of the independent variables (X
1: 15%, 21%, 27%; X
2: 42 °C, 47 °C, 58 °C; and X
3: 4.5, 5.0, 5.5). The detailed experimental layout is presented in
Table 2.
The following polynomial equation was fitted to describe a correlation between independent variables, including linear (β
1, β
2, β
3), quadratic (β
11, β
22, β
33), and interaction terms (β
12, β
13, β
23) and the response (Y).
Results obtained were subjected to the regression analysis and to the analysis of variance (ANOVA), using Statistica software ver. 12.5 (StatSoft, Kraków, Poland). The effects were summarized and insignificant terms, with confidence interval (p) higher than 0.5, were removed from the model.
4.6. Ethanol Fermentation Process
The hydrolysates were subjected to periodic ethanol fermentation in Erlenmayer flasks (100 mL) with industrial distillery yeast Saccharomyces cerevisiae SIHA Active Yeast 6 (EATON, Irvine, CA, USA) used in the concentration of 0.2 g L−1, rehydrated in sterile distilled water right before the process. The fermentation process was performed in Binder incubator (BINDER, Tuttlingen, Germany) at a temperature of 37 °C for 72 h.
4.7. Analytical Analyses
4.7.1. Enzyme Activity Assay and Protein Concentration Determination
The enzymatic activities of cellulases (CMCases, EC 3.2.1.4) and endoxylanases (EC 3.2.1.8) in extracts from solid state cultures, as well as in the concentrated preparation C1 and commercial Cellic CTec2, were determined according to the methods described in the literature [
43], using DNS reagent [
44]. The unit of enzyme activity was expressed as the amount of enzyme which liberates 1 μmol of glucose or xylose per minute per 1 mL (U mL
−1). To determine specific activity (U mg
−1), protein concentration was determined with Lowry’s method [
45].
4.7.2. HPLC Analysis
Concentrations of glucose (glu), arabinose (arab), and a mixture of galactose + xylose + mannose (gal, xyl, man) were determined with high-performance liquid chromatography (HPLC) in the Dionex Ultimate 3000 system (Thermo Scientific, Waltham, MA, USA), using an Aminex HPX-87H column (Bio-Rad, EATON, Irvine, CA, USA) coupled with a UV (λ = 210 nm) and IR detectors. Analyses were carried out at a temperature of 35 °C, with 0.01 M sulfuric acid (VI) used as the mobile phase at the flow rate of 0.6 mL min−1.
Hydrolysis yield expressed in % was calculated as follows:
Ethanol concentration in the post-fermentation liquids was determined with the liquid chromatography using a Shimadzu Prominence system with RID detector–10A (Shimadzu, Kyoto, Japan). Samples centrifuged at 15,000 rpm for 10 min were filtered using a syringe filter (pore diameter 0.22 µm). Separation conditions were as follows: Security Guard Cartridges Carbo-Pb pre-column (4 × 3.0 mm ID) (Phenomenex, Torrance, MA, USA), Rezex ROA-Organic Acid H+ column (300 × 7.8 mm) (Phenomenex, Torrance, MA, USA), eluent −0.005 M H2SO4, mobile phase flow rate −0.6 mL min−1, temperature 60 °C, and injection volume −20 μL. Chromatograms were analyzed in Chromax 10 software (Pol-Lab, Poland).
Process yield was expressed in mL of 100% ethanol per kg of substrate dry matter (mL kg d.m.−1) and in mL of 100% ethanol per kg of total sugars content (mL kg sugars−1). According to the stoichiometric equations of the ethanol fermentation process of pentoses and hexoses, the theoretical ethanol yield was calculated from the total amount of sugars in the hydrolysates (% of theoretical yield).






{kind=link}
{kind=link}
{kind=link}
{kind=link}