Platelet Integrins in Tumor Metastasis: Do They Represent a Therapeutic Target?


Abstract
:1. The Role of Platelets in Hemostasis and Tumor Metastasis
2. The Repertoire and Function of Integrins at the Platelet Surface
2.1. The Platelet β1 Integrins
2.1.1. α2β1
2.1.2. α5β1
2.1.3. α6β1
2.2. The Platelet β3 Integrins
2.2.1. αIIbβ3
2.2.2. αvβ3
3. The Role of Platelet Integrins in the Interplay with Tumor Cells and in Tumor Metastasis
3.1. The β1 Integrins
3.2. The β3 Integrins
4. Is Targeting Platelet Integrins a Potentially Promising Anti-Metastatic Strategy?
5. Conclusions
Acknowledgments
Authors Contributions
Conflicts of Interest
References
- Versteeg, H.H.; Heemskerk, J.W.; Levi, M.; Reitsma, P.H. New fundamentals in hemostasis. Physiol. Rev. 2013, 93, 327–358. [Google Scholar] [CrossRef] [PubMed]
- Savage, B.; Saldivar, E.; Ruggeri, Z.M. Initiation of platelet adhesion by arrest onto fibrinogen or translocation on von Willebrand factor. Cell 1996, 84, 289–297. [Google Scholar] [CrossRef]
- Ikeda, Y.; Handa, M.; Kawano, K.; Kamata, T.; Murata, M.; Araki, Y.; Anbo, H.; Kawai, Y.; Watanabe, K.; Itagaki, I.; et al. The role of von Willebrand factor and fibrinogen in platelet aggregation under varying shear stress. J. Clin. Investig. 1991, 87, 1234–1240. [Google Scholar] [CrossRef] [PubMed]
- Reininger, A.J. Platelet function under high shear conditions. Hamostaseologie 2009, 29, 21–24. [Google Scholar] [PubMed]
- Bergmeier, W.; Hynes, R.O. Extracellular matrix proteins in hemostasis and thrombosis. Cold Spring Harb. Perspect. Biol. 2012, 4, a005132. [Google Scholar] [CrossRef] [PubMed]
- Nieswandt, B.; Watson, S.P. Platelet-collagen interaction: Is GPVI the central receptor? Blood 2003, 102, 449–461. [Google Scholar] [CrossRef] [PubMed]
- Hechler, B.; Gachet, C. Purinergic Receptors in Thrombosis and Inflammation. Arterioscler. Thromb. Vasc. Biol. 2015, 35, 2307–2315. [Google Scholar] [CrossRef] [PubMed]
- Shattil, S.J.; Newman, P.J. Integrins: Dynamic scaffolds for adhesion and signaling in platelets. Blood 2004, 104, 1606–1615. [Google Scholar] [CrossRef] [PubMed]
- Jackson, S.P.; Nesbitt, W.S.; Kulkarni, S. Signaling events underlying thrombus formation. J. Thromb. Haemost. 2003, 1, 1602–1612. [Google Scholar] [CrossRef] [PubMed]
- Nurden, A.T. Platelets, inflammation and tissue regeneration. Thromb. Haemost. 2011, 105, S13–S33. [Google Scholar] [CrossRef] [PubMed]
- Rondina, M.T.; Garraud, O. Emerging evidence for platelets as immune and inflammatory effector cells. Front. Immunol. 2014, 5, 653. [Google Scholar] [CrossRef] [PubMed]
- Walsh, T.G.; Metharom, P.; Berndt, M.C. The functional role of platelets in the regulation of angiogenesis. Platelets 2015, 26, 199–211. [Google Scholar] [CrossRef] [PubMed]
- Mammadova-Bach, E.; Mangin, P.; Lanza, F.; Gachet, C. Platelets in cancer. From basic research to therapeutic implications. Hamostaseologie 2015, 35, 325–336. [Google Scholar] [CrossRef] [PubMed]
- Leblanc, R.; Peyruchaud, O. Metastasis: New functional implications of platelets and megakaryocytes. Blood 2016, 128, 24–31. [Google Scholar] [CrossRef] [PubMed]
- Varki, A. Trousseau’s syndrome: Multiple definitions and multiple mechanisms. Blood 2007, 110, 1723–1729. [Google Scholar] [CrossRef] [PubMed]
- Gasic, G.J.; Gasic, T.B.; Stewart, C.C. Antimetastatic effects associated with platelet reduction. Proc. Natl. Acad. Sci. USA 1968, 61, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Rothwell, P.M.; Wilson, M.; Price, J.F.; Belch, J.F.; Meade, T.W.; Mehta, Z. Effect of daily aspirin on risk of cancer metastasis: A study of incident cancers during randomised controlled trials. Lancet 2012, 379, 1591–1601. [Google Scholar] [CrossRef]
- Shiao, J.; Thomas, K.M.; Rahimi, A.S.; Rao, R.; Yan, J.; Xie, X.-J.; DaSilva, M.; Spangler, A.; Leitch, M.; Wooldridge, R.; et al. Aspirin/antiplatelet agent use improves disease-free survival and reduces the risk of distant metastases in Stage II and III triple-negative breast cancer patients. Breast Cancer Res. Treat. 2017, 161, 463–471. [Google Scholar] [CrossRef] [PubMed]
- Rothwell, P.M.; Fowkes, F.G.; Belch, J.F.; Ogawa, H.; Warlow, C.P.; Meade, T.W. Effect of daily aspirin on long-term risk of death due to cancer: Analysis of individual patient data from randomised trials. Lancet 2011, 377, 31–41. [Google Scholar] [CrossRef]
- Suzuki-Inoue, K.; Kato, Y.; Inoue, O.; Kaneko, M.K.; Mishima, K.; Yatomi, Y.; Yamazaki, Y.; Narimatsu, H.; Ozaki, Y. Involvement of the snake toxin receptor CLEC-2, in podoplanin-mediated platelet activation, by cancer cells. J. Biol. Chem. 2007, 282, 25993–26001. [Google Scholar] [CrossRef] [PubMed]
- Gong, L.; Cai, Y.; Zhou, X.; Yang, H. Activated platelets interact with lung cancer cells through P-selectin glycoprotein ligand-1. Pathol. Oncol. Res. 2012, 18, 989–996. [Google Scholar] [CrossRef] [PubMed]
- Boukerche, H.; Berthier-Vergnes, O.; Tabone, E.; Dore, J.F.; Leung, L.L.; McGregor, J.L. Platelet-melanoma cell interaction is mediated by the glycoprotein IIb-IIIa complex. Blood 1989, 74, 658–663. [Google Scholar] [PubMed]
- Mammadova-Bach, E.; Zigrino, P.; Brucker, C.; Bourdon, C.; Freund, M.; de Arcangelis, A.; Abrams, S.I.; Orend, G.; Gachet, C.; Mangin, P.H. Platelet integrin α6β1 controls lung metastasis through direct binding to cancer cell-derived ADAM9. JCI Insight 2016, 1, e88245. [Google Scholar] [CrossRef] [PubMed]
- Tesfamariam, B. Involvement of platelets in tumor cell metastasis. Pharmacol. Ther. 2016, 157, 112–119. [Google Scholar] [CrossRef] [PubMed]
- Placke, T.; Kopp, H.G.; Salih, H.R. The wolf in sheep’s clothing: Platelet-derived “pseudo self” impairs cancer cell “missing self” recognition by NK cells. Oncoimmunology 2012, 1, 557–559. [Google Scholar] [CrossRef] [PubMed]
- Nieswandt, B.; Hafner, M.; Echtenacher, B.; Mannel, D.N. Lysis of tumor cells by natural killer cells in mice is impeded by platelets. Cancer Res. 1999, 59, 1295–1300. [Google Scholar] [PubMed]
- Kopp, H.G.; Placke, T.; Salih, H.R. Platelet-derived transforming growth factor-beta down-regulates NKG2D thereby inhibiting natural killer cell antitumor reactivity. Cancer Res. 2009, 69, 7775–7783. [Google Scholar] [CrossRef] [PubMed]
- Labelle, M.; Begum, S.; Hynes, R.O. Direct signaling between platelets and cancer cells induces an epithelial-mesenchymal-like transition and promotes metastasis. Cancer Cell 2011, 20, 576–590. [Google Scholar] [CrossRef] [PubMed]
- Dardik, R.; Kaufmann, Y.; Savion, N.; Rosenberg, N.; Shenkman, B.; Varon, D. Platelets mediate tumor cell adhesion to the subendothelium under flow conditions: Involvement of platelet GPIIb-IIIa and tumor cell α(v) integrins. Int. J. Cancer 1997, 70, 201–207. [Google Scholar] [CrossRef]
- Schumacher, D.; Strilic, B.; Sivaraj, K.K.; Wettschureck, N.; Offermanns, S. Platelet-derived nucleotides promote tumor-cell transendothelial migration and metastasis via P2Y2 receptor. Cancer Cell 2013, 24, 130–137. [Google Scholar] [CrossRef] [PubMed]
- Sawicki, G.; Sanders, E.J.; Salas, E.; Wozniak, M.; Rodrigo, J.; Radomski, M.W. Localization and translocation of MMP-2 during aggregation of human platelets. Thromb. Haemost. 1998, 80, 836–839. [Google Scholar] [PubMed]
- Choi, J.H.; Kim, H.; Kim, H.S.; Um, S.H.; Choi, J.W.; Oh, B.K. MMP-2 detective silicon nanowire biosensor using enzymatic cleavage reaction. J. Biomed. Nanotechnol. 2013, 9, 732–735. [Google Scholar] [CrossRef] [PubMed]
- Gay, L.J.; Felding-Habermann, B. Contribution of platelets to tumour metastasis. Nat. Rev. Cancer 2011, 11, 123–134. [Google Scholar] [CrossRef] [PubMed]
- Jurasz, P.; Alonso-Escolano, D.; Radomski, M.W. Platelet—Cancer interactions: Mechanisms and pharmacology of tumour cell-induced platelet aggregation. Br. J. Pharmacol. 2004, 143, 819–826. [Google Scholar] [CrossRef] [PubMed]
- Erpenbeck, L.; Schon, M.P. Deadly allies: The fatal interplay between platelets and metastasizing cancer cells. Blood 2010, 115, 3427–3436. [Google Scholar] [CrossRef] [PubMed]
- Varga-Szabo, D.; Pleines, I.; Nieswandt, B. Cell adhesion mechanisms in platelets. Arterioscler. Thromb. Vasc. Biol. 2008, 28, 403–412. [Google Scholar] [CrossRef] [PubMed]
- Bennett, J.S. Regulation of integrins in platelets. Biopolymers 2015, 104, 323–333. [Google Scholar] [CrossRef] [PubMed]
- Takagi, J.; Springer, T.A. Integrin activation and structural rearrangement. Immunol. Rev. 2002, 186, 141–163. [Google Scholar] [CrossRef] [PubMed]
- Ruggeri, Z.M. Platelet adhesion under flow. Microcirculation 2009, 16, 58–83. [Google Scholar] [CrossRef] [PubMed]
- Ruggeri, Z.M.; Mendolicchio, G.L. Adhesion mechanisms in platelet function. Circ. Res. 2007, 100, 1673–1685. [Google Scholar] [CrossRef] [PubMed]
- Guidetti, G.F.; Canobbio, I.; Torti, M. PI3K/Akt in platelet integrin signaling and implications in thrombosis. Adv. Biol. Regul. 2015, 59, 36–52. [Google Scholar] [CrossRef] [PubMed]
- Nieswandt, B.; Varga-Szabo, D.; Elvers, M. Integrins in platelet activation. J. Thromb. Haemost. 2009, 7, 206–209. [Google Scholar] [CrossRef] [PubMed]
- Bianconi, D.; Unseld, M.; Prager, G.W. Integrins in the Spotlight of Cancer. Int. J. Mol. Sci. 2016, 17, 2037. [Google Scholar] [CrossRef] [PubMed]
- Schaff, M.; Tang, C.; Maurer, E.; Bourdon, C.; Receveur, N.; Eckly, A.; Hechler, B.; Arnold, C.; de Arcangelis, A.; Nieswandt, B.; et al. Integrin α6β1 is the main receptor for vascular laminins and plays a role in platelet adhesion, activation, and arterial thrombosis. Circulation 2013, 128, 541–552. [Google Scholar] [CrossRef] [PubMed]
- Zutter, M.M.; Santoro, S.A. Widespread histologic distribution of the α2β1 integrin cell-surface collagen receptor. Am. J. Pathol. 1990, 137, 113–120. [Google Scholar] [PubMed]
- Madamanchi, A.; Santoro, S.A.; Zutter, M.M. α2β1 Integrin. Adv. Exp. Med. Biol. 2014, 819, 41–60. [Google Scholar] [CrossRef] [PubMed]
- Langsenlehner, U.; Renner, W.; Yazdani-Biuki, B.; Eder, T.; Wascher, T.C.; Paulweber, B.; Clar, H.; Hofmann, G.; Samonigg, H.; Krippl, P. Integrin α2 and β3 gene polymorphisms and breast cancer risk. Breast Cancer Res. Treat. 2006, 97, 67–72. [Google Scholar] [CrossRef] [PubMed]
- Gerger, A.; Hofmann, G.; Langsenlehner, U.; Renner, W.; Weitzer, W.; Wehrschutz, M.; Wascher, T.; Samonigg, H.; Krippl, P. Integrin α2 and β3 gene polymorphisms and colorectal cancer risk. Int. J. Colorectal Dis. 2009, 24, 159–163. [Google Scholar] [CrossRef] [PubMed]
- Ayala, F.; Corral, J.; Gonzalez-Conejero, R.; Sanchez, I.; Moraleda, J.M.; Vicente, V. Genetic polymorphisms of platelet adhesive molecules: Association with breast cancer risk and clinical presentation. Breast Cancer Res. Treat. 2003, 80, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Sarratt, K.L.; Chen, H.; Zutter, M.M.; Santoro, S.A.; Hammer, D.A.; Kahn, M.L. GPVI and α2β1 play independent critical roles during platelet adhesion and aggregate formation to collagen under flow. Blood 2005, 106, 1268–1277. [Google Scholar] [CrossRef] [PubMed]
- Di Paola, J.; Jugessur, A.; Goldman, T.; Reiland, J.; Tallman, D.; Sayago, C.; Murray, J.C. Platelet glycoprotein I(b) α and integrin α2β1polymorphisms: Gene frequencies and linkage disequilibrium in a population diversity panel. J. Thromb. Haemost. 2005, 3, 1511–1521. [Google Scholar] [CrossRef] [PubMed]
- Inoue, O.; Suzuki-Inoue, K.; Dean, W.L.; Frampton, J.; Watson, S.P. Integrin α2β1 mediates outside-in regulation of platelet spreading on collagen through activation of Src kinases and PLCgamma2. J. Cell Biol. 2003, 160, 769–780. [Google Scholar] [CrossRef] [PubMed]
- Kehrel, B.; Balleisen, L.; Kokott, R.; Mesters, R.; Stenzinger, W.; Clemetson, K.J.; van de Loo, J. Deficiency of intact thrombospondin and membrane glycoprotein Ia in platelets with defective collagen-induced aggregation and spontaneous loss of disorder. Blood 1988, 71, 1074–1078. [Google Scholar] [PubMed]
- Nieuwenhuis, H.K.; Akkerman, J.W.; Houdijk, W.P.; Sixma, J.J. Human blood platelets showing no response to collagen fail to express surface glycoprotein Ia. Nature 1985, 318, 470–472. [Google Scholar] [CrossRef] [PubMed]
- Holtkotter, O.; Nieswandt, B.; Smyth, N.; Muller, W.; Hafner, M.; Schulte, V.; Krieg, T.; Eckes, B. Integrin α2-deficient mice develop normally, are fertile, but display partially defective platelet interaction with collagen. J. Biol. Chem. 2002, 277, 10789–10794. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Pappan, L.K.; Grenache, D.G.; Li, Z.; Tollefsen, D.M.; Santoro, S.A.; Zutter, M.M. The contributions of the α2β1 integrin to vascular thrombosis in vivo. Blood 2003, 102, 3652–3657. [Google Scholar] [CrossRef] [PubMed]
- Kuijpers, M.J.; Pozgajova, M.; Cosemans, J.M.; Munnix, I.C.; Eckes, B.; Nieswandt, B.; Heemskerk, J.W. Role of murine integrin α2β1 in thrombus stabilization and embolization: Contribution of thromboxane A2. Thromb. Haemost. 2007, 98, 1072–1080. [Google Scholar] [CrossRef] [PubMed]
- Kunicki, T.J. The role of platelet collagen receptor (glycoprotein Ia/IIa; integrin α2β1) polymorphisms in thrombotic disease. Curr. Opin. Hematol. 2001, 8, 277–285. [Google Scholar] [CrossRef] [PubMed]
- Kasirer-Friede, A.; Kahn, M.L.; Shattil, S.J. Platelet integrins and immunoreceptors. Immunol. Rev. 2007, 218, 247–264. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.T.; Rayburn, H.; Hynes, R.O. Embryonic mesodermal defects in α5 integrin-deficient mice. Development 1993, 119, 1093–1105. [Google Scholar] [PubMed]
- Schaffner, F.; Ray, A.M.; Dontenwill, M. Integrin α5β1, the Fibronectin Receptor, as a Pertinent Therapeutic Target in Solid Tumors. Cancers 2013, 5, 27–47. [Google Scholar] [CrossRef] [PubMed]
- Jung, A.C.; Ray, A.M.; Ramolu, L.; Macabre, C.; Simon, F.; Noulet, F.; Blandin, A.F.; Renner, G.; Lehmann, M.; Choulier, L.; et al. Caveolin-1-negative head and neck squamous cell carcinoma primary tumors display increased epithelial to mesenchymal transition and prometastatic properties. Oncotarget 2015, 6, 41884–41901. [Google Scholar] [CrossRef] [PubMed]
- Bell-McGuinn, K.M.; Matthews, C.M.; Ho, S.N.; Barve, M.; Gilbert, L.; Penson, R.T.; Lengyel, E.; Palaparthy, R.; Gilder, K.; Vassos, A.; et al. A phase II, single-arm study of the anti-α5β1 integrin antibody volociximab as monotherapy in patients with platinum-resistant advanced epithelial ovarian or primary peritoneal cancer. Gynecol. Oncol. 2011, 121, 273–279. [Google Scholar] [CrossRef] [PubMed]
- Beumer, S.; MJ, I.J.; de Groot, P.G.; Sixma, J.J. Platelet adhesion to fibronectin in flow: Dependence on surface concentration and shear rate, role of platelet membrane glycoproteins GP IIb/IIIa and VLA-5, and inhibition by heparin. Blood 1994, 84, 3724–3733. [Google Scholar] [PubMed]
- Zaidi, T.N.; McIntire, L.V.; Farrell, D.H.; Thiagarajan, P. Adhesion of platelets to surface-bound fibrinogen under flow. Blood 1996, 88, 2967–2972. [Google Scholar] [PubMed]
- McCarty, O.J.; Zhao, Y.; Andrew, N.; Machesky, L.M.; Staunton, D.; Frampton, J.; Watson, S.P. Evaluation of the role of platelet integrins in fibronectin-dependent spreading and adhesion. J. Thromb. Haemost. 2004, 2, 1823–1833. [Google Scholar] [CrossRef] [PubMed]
- Maurer, E.; Schaff, M.; Receveur, N.; Bourdon, C.; Mercier, L.; Nieswandt, B.; Dubois, C.; Jandrot-Perrus, M.; Goetz, J.G.; Lanza, F.; et al. Fibrillar cellular fibronectin supports efficient platelet aggregation and procoagulant activity. Thromb. Haemost. 2015, 114, 1175–1188. [Google Scholar] [CrossRef] [PubMed]
- Hallmann, R.; Horn, N.; Selg, M.; Wendler, O.; Pausch, F.; Sorokin, L.M. Expression and function of laminins in the embryonic and mature vasculature. Physiol. Rev. 2005, 85, 979–1000. [Google Scholar] [CrossRef] [PubMed]
- Georges-Labouesse, E.; Messaddeq, N.; Yehia, G.; Cadalbert, L.; Dierich, A.; Le Meur, M. Absence of integrin α6 leads to epidermolysis bullosa and neonatal death in mice. Nat. Genet. 1996, 13, 370–373. [Google Scholar] [CrossRef] [PubMed]
- Hogg, N.; Bates, P.A. Genetic analysis of integrin function in man: LAD-1 and other syndromes. J. Int. Soc. Matrix Biol. 2000, 19, 211–222. [Google Scholar] [CrossRef]
- Wewer, U.M.; Shaw, L.M.; Albrechtsen, R.; Mercurio, A.M. The integrin α6β1 promotes the survival of metastatic human breast carcinoma cells in mice. Am. J. Pathol. 1997, 151, 1191–1198. [Google Scholar] [PubMed]
- Sroka, I.C.; Anderson, T.A.; McDaniel, K.M.; Nagle, R.B.; Gretzer, M.B.; Cress, A.E. The laminin binding integrin α6β1 in prostate cancer perineural invasion. J. Cell. Physiol. 2010, 224, 283–288. [Google Scholar] [CrossRef] [PubMed]
- Lathia, J.D.; Gallagher, J.; Heddleston, J.M.; Wang, J.; Eyler, C.E.; Macswords, J.; Wu, Q.; Vasanji, A.; McLendon, R.E.; Hjelmeland, A.B.; et al. Integrin α6 regulates glioblastoma stem cells. Cell Stem Cell 2010, 6, 421–432. [Google Scholar] [CrossRef] [PubMed]
- Lamb, L.E.; Zarif, J.C.; Miranti, C.K. The androgen receptor induces integrin α6β1 to promote prostate tumor cell survival via NF-κB and Bcl-xL Independently of PI3K signaling. Cancer Res. 2011, 71, 2739–2749. [Google Scholar] [CrossRef] [PubMed]
- Ports, M.O.; Nagle, R.B.; Pond, G.D.; Cress, A.E. Extracellular engagement of α6 integrin inhibited urokinase-type plasminogen activator-mediated cleavage and delayed human prostate bone metastasis. Cancer Res. 2009, 69, 5007–5014. [Google Scholar] [CrossRef] [PubMed]
- Hindriks, G.; Ijsseldijk, M.J.; Sonnenberg, A.; Sixma, J.J.; de Groot, P.G. Platelet adhesion to laminin: Role of Ca2+ and Mg2+ ions, shear rate, and platelet membrane glycoproteins. Blood 1992, 79, 928–935. [Google Scholar] [PubMed]
- Inoue, O.; Suzuki-Inoue, K.; McCarty, O.J.; Moroi, M.; Ruggeri, Z.M.; Kunicki, T.J.; Ozaki, Y.; Watson, S.P. Laminin stimulates spreading of platelets through integrin α6β1-dependent activation of GPVI. Blood 2006, 107, 1405–1412. [Google Scholar] [CrossRef] [PubMed]
- Sonnenberg, A.; Modderman, P.W.; Hogervorst, F. Laminin receptor on platelets is the integrin VLA-6. Nature 1988, 336, 487–489. [Google Scholar] [CrossRef] [PubMed]
- Nigatu, A.; Sime, W.; Gorfu, G.; Geberhiwot, T.; Anduren, I.; Ingerpuu, S.; Doi, M.; Tryggvason, K.; Hjemdahl, P.; Patarroyo, M. Megakaryocytic cells synthesize and platelets secrete α5-laminins, and the endothelial laminin isoform laminin 10 (α5β1γ1) strongly promotes adhesion but not activation of platelets. Thromb. Haemost. 2006, 95, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Geberhiwot, T.; Ingerpuu, S.; Pedraza, C.; Neira, M.; Lehto, U.; Virtanen, I.; Kortesmaa, J.; Tryggvason, K.; Engvall, E.; Patarroyo, M. Blood platelets contain and secrete laminin-8 (α4β1γ1) and adhere to laminin-8 via α6β1 integrin. Exp. Cell Res. 1999, 253, 723–732. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.C.; Chang, H.H.; Lin, C.T.; Lo, S.J. The integrin α6β1 modulation of PI3K and Cdc42 activities induces dynamic filopodium formation in human platelets. J. Biomed. Sci. 2005, 12, 881–898. [Google Scholar] [CrossRef] [PubMed]
- Ill, C.R.; Engvall, E.; Ruoslahti, E. Adhesion of platelets to laminin in the absence of activation. J. Cell Biol. 1984, 99, 2140–2145. [Google Scholar] [CrossRef] [PubMed]
- Pan, L.; Zhao, Y.; Yuan, Z.; Qin, G. Research advances on structure and biological functions of integrins. SpringerPlus 2016, 5, 1094. [Google Scholar] [CrossRef] [PubMed]
- Grossi, I.M.; Hatfield, J.S.; Fitzgerald, L.A.; Newcombe, M.; Taylor, J.D.; Honn, K.V. Role of tumor cell glycoproteins immunologically related to glycoproteins Ib and IIb/IIIa in tumor cell-platelet and tumor cell-matrix interactions. FASEB J. 1988, 2, 2385–2395. [Google Scholar] [PubMed]
- Honn, K.V.; Chen, Y.Q.; Timar, J.; Onoda, J.M.; Hatfield, J.S.; Fligiel, S.E.; Steinert, B.W.; Diglio, C.A.; Grossi, I.M.; Nelson, K.K.; et al. αIIbβ 3 integrin expression and function in subpopulations of murine tumors. Exp. Cell Res. 1992, 201, 23–32. [Google Scholar] [CrossRef]
- Timar, J.; Trikha, M.; Szekeres, K.; Bazaz, R.; Honn, K. Expression and function of the high affinity αIIbβ3 integrin in murine melanoma cells. Clin. Exp. Metastasis 1998, 16, 437–445. [Google Scholar] [CrossRef] [PubMed]
- Desgrosellier, J.S.; Cheresh, D.A. Integrins in cancer: Biological implications and therapeutic opportunities. Nat. Rev. Cancer 2010, 10, 9–22. [Google Scholar] [CrossRef] [PubMed]
- Scatena, M.; Almeida, M.; Chaisson, M.L.; Fausto, N.; Nicosia, R.F.; Giachelli, C.M. NF-κB mediates α3 integrin-induced endothelial cell survival. J. Cell Biol. 1998, 141, 1083–1093. [Google Scholar] [CrossRef] [PubMed]
- Morimoto, K.; Satoh-Yamaguchi, K.; Hamaguchi, A.; Inoue, Y.; Takeuchi, M.; Okada, M.; Ikeda, W.; Takai, Y.; Imai, T. Interaction of cancer cells with platelets mediated by Necl-5/poliovirus receptor enhances cancer cell metastasis to the lungs. Oncogene 2008, 27, 264–273. [Google Scholar] [CrossRef] [PubMed]
- Khan, Z.; Marshall, J.F. The role of integrins in TGF-β activation in the tumour stroma. Cell Tissue Res. 2016, 365, 657–673. [Google Scholar] [CrossRef] [PubMed]
- Nierodzik, M.L.; Kajumo, F.; Karpatkin, S. Effect of thrombin treatment of tumor cells on adhesion of tumor cells to platelets in vitro and tumor metastasis in vivo. Cancer Res. 1992, 52, 3267–3272. [Google Scholar] [PubMed]
- Wagner, C.L.; Mascelli, M.A.; Neblock, D.S.; Weisman, H.F.; Coller, B.S.; Jordan, R.E. Analysis of GPIIb/IIIa receptor number by quantification of 7E3 binding to human platelets. Blood 1996, 88, 907–914. [Google Scholar] [PubMed]
- Nurden, A.T. Glanzmann thrombasthenia. Orphanet J. Rare Dis. 2006, 1, 10. [Google Scholar] [CrossRef] [PubMed]
- Yousuf, O.; Bhatt, D.L. The evolution of antiplatelet therapy in cardiovascular disease. Nat. Rev. Cardiol. 2011, 8, 547–559. [Google Scholar] [CrossRef] [PubMed]
- Watson, S.P.; Auger, J.M.; McCarty, O.J.; Pearce, A.C. GPVI and integrin αIIbβ3 signaling in platelets. J. Thromb. Haemost. 2005, 3, 1752–1762. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Xiang, B.; Ye, S.; Chrzanowska-Wodnicka, M.; Morris, A.J.; Gartner, T.K.; Whiteheart, S.W.; White, G.C., 2nd; Smyth, S.S.; Li, Z. Distinct roles for Rap1b protein in platelet secretion and integrin αIIbβ3 outside-in signaling. J. Biol. Chem. 2011, 286, 39466–39477. [Google Scholar] [CrossRef] [PubMed]
- Goschnick, M.W.; Lau, L.M.; Wee, J.L.; Liu, Y.S.; Hogarth, P.M.; Robb, L.M.; Hickey, M.J.; Wright, M.D.; Jackson, D.E. Impaired “outside-in” integrin αIIbβ3 signaling and thrombus stability in TSSC6-deficient mice. Blood 2006, 108, 1911–1918. [Google Scholar] [CrossRef] [PubMed]
- Law, D.A.; DeGuzman, F.R.; Heiser, P.; Ministri-Madrid, K.; Killeen, N.; Phillips, D.R. Integrin cytoplasmic tyrosine motif is required for outside-in αIIbβ3 signalling and platelet function. Nature 1999, 401, 808–811. [Google Scholar] [CrossRef] [PubMed]
- Phillips, D.R.; Prasad, K.S.; Manganello, J.; Bao, M.; Nannizzi-Alaimo, L. Integrin tyrosine phosphorylation in platelet signaling. Curr. Opin. Cell Biol. 2001, 13, 546–554. [Google Scholar] [CrossRef]
- Takizawa, H.; Nishimura, S.; Takayama, N.; Oda, A.; Nishikii, H.; Morita, Y.; Kakinuma, S.; Yamazaki, S.; Okamura, S.; Tamura, N.; et al. Lnk regulates integrin αIIbβ3 outside-in signaling in mouse platelets, leading to stabilization of thrombus development in vivo. J. Clin. Investig. 2010, 120, 179–190. [Google Scholar] [CrossRef] [PubMed]
- Valiyaveettil, M.; Kar, N.; Ashraf, M.Z.; Byzova, T.V.; Febbraio, M.; Podrez, E.A. Oxidized high-density lipoprotein inhibits platelet activation and aggregation via scavenger receptor BI. Blood 2008, 111, 1962–1971. [Google Scholar] [CrossRef] [PubMed]
- Paul, B.Z.; Vilaire, G.; Kunapuli, S.P.; Bennett, J.S. Concurrent signaling from Galphaq- and Galphai-coupled pathways is essential for agonist-induced αvβ3 activation on human platelets. J. Thromb. Haemost. 2003, 1, 814–820. [Google Scholar] [CrossRef] [PubMed]
- Mor-Cohen, R. Disulfide Bonds as Regulators of Integrin Function in Thrombosis and Hemostasis. Antioxid. Redox Signal. 2016, 24, 16–31. [Google Scholar] [CrossRef] [PubMed]
- Mahimkar, R.M.; Visaya, O.; Pollock, A.S.; Lovett, D.H. The disintegrin domain of ADAM9: A ligand for multiple beta1 renal integrins. Biochem. J. 2005, 385, 461–468. [Google Scholar] [CrossRef] [PubMed]
- Karpatkin, S.; Pearlstein, E.; Ambrogio, C.; Coller, B.S. Role of adhesive proteins in platelet tumor interaction in vitro and metastasis formation in vivo. J. Clin. Investig. 1988, 81, 1012–1019. [Google Scholar] [CrossRef] [PubMed]
- Nierodzik, M.L.; Plotkin, A.; Kajumo, F.; Karpatkin, S. Thrombin stimulates tumor-platelet adhesion in vitro and metastasis in vivo. J. Clin. Investig. 1991, 87, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Qian, W.; Tao, L.; Wang, Y.; Zhang, F.; Li, M.; Huang, S.; Wang, A.; Chen, W.; Yue, Z.; Chen, L.; et al. Downregulation of Integrins in Cancer Cells and Anti-Platelet Properties Are Involved in Holothurian Glycosaminoglycan-Mediated Disruption of the Interaction of Cancer Cells and Platelets in Hematogenous Metastasis. J. Vasc. Res. 2015, 52, 197–209. [Google Scholar] [CrossRef] [PubMed]
- Chopra, H.; Hatfield, J.S.; Chang, Y.S.; Grossi, I.M.; Fitzgerald, L.A.; O’Gara, C.Y.; Marnett, L.J.; Diglio, C.A.; Taylor, J.D.; Honn, K.V. Role of tumor cytoskeleton and membrane glycoprotein IRGpIIb/IIIa in platelet adhesion to tumor cell membrane and tumor cell-induced platelet aggregation. Cancer Res. 1988, 48, 3787–3800. [Google Scholar] [PubMed]
- Humphries, M.J.; Yamada, K.M.; Olden, K. Investigation of the biological effects of anti-cell adhesive synthetic peptides that inhibit experimental metastasis of B16-F10 murine melanoma cells. J. Clin. Investig. 1988, 81, 782–790. [Google Scholar] [CrossRef] [PubMed]
- McCarty, O.J.; Mousa, S.A.; Bray, P.F.; Konstantopoulos, K. Immobilized platelets support human colon carcinoma cell tethering, rolling, and firm adhesion under dynamic flow conditions. Blood 2000, 96, 1789–1797. [Google Scholar] [PubMed]
- Gawaz, M.P.; Loftus, J.C.; Bajt, M.L.; Frojmovic, M.M.; Plow, E.F.; Ginsberg, M.H. Ligand bridging mediates integrin αIIbβ3 (platelet GPIIB-IIIA) dependent homotypic and heterotypic cell-cell interactions. J. Clin. Investig. 1991, 88, 1128–1134. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Liu, Y.; Gao, Y.; Shen, J.; Zheng, S.; Wei, M.; Zeng, X. Modified heparins inhibit integrin α(IIb)β(3) mediated adhesion of melanoma cells to platelets in vitro and in vivo. Int. J. Cancer 2009, 125, 2058–2065. [Google Scholar] [CrossRef] [PubMed]
- Lonsdorf, A.S.; Kramer, B.F.; Fahrleitner, M.; Schonberger, T.; Gnerlich, S.; Ring, S.; Gehring, S.; Schneider, S.W.; Kruhlak, M.J.; Meuth, S.G.; et al. Engagement of αIIbβ3 (GPIIb/IIIa) with αvβ3 integrinD mediates interaction of melanoma cells with platelets: A connection to hematogenous metastasis. J. Biol. Chem. 2012, 287, 2168–2178. [Google Scholar] [CrossRef] [PubMed]
- Bastida, E.; Escolar, G.; Almirall, L.; Ordinas, A. Platelet activation induced by a human neuroblastoma tumor cell line is reduced by prior administration of ticlopidine. Thromb. Haemost. 1986, 55, 333–337. [Google Scholar] [PubMed]
- Heinmoller, E.; Weinel, R.J.; Heidtmann, H.H.; Salge, U.; Seitz, R.; Schmitz, I.; Muller, K.M.; Zirngibl, H. Studies on tumor-cell-induced platelet aggregation in human lung cancer cell lines. J. Cancer Res. Clin. Oncol. 1996, 122, 735–744. [Google Scholar] [CrossRef] [PubMed]
- Boukerche, H.; Berthier-Vergnes, O.; Penin, F.; Tabone, E.; Lizard, G.; Bailly, M.; McGregor, J.L. Human melanoma cell lines differ in their capacity to release ADP and aggregate platelets. Br. J. Haematol. 1994, 87, 763–772. [Google Scholar] [CrossRef] [PubMed]
- Alonso-Escolano, D.; Strongin, A.Y.; Chung, A.W.; Deryugina, E.I.; Radomski, M.W. Membrane type-1 matrix metalloproteinase stimulates tumour cell-induced platelet aggregation: Role of receptor glycoproteins. Br. J. Pharmacol. 2004, 141, 241–252. [Google Scholar] [CrossRef] [PubMed]
- Jurasz, P.; Sawicki, G.; Duszyk, M.; Sawicka, J.; Miranda, C.; Mayers, I.; Radomski, M.W. Matrix metalloproteinase 2 in tumor cell-induced platelet aggregation: Regulation by nitric oxide. Cancer Res. 2001, 61, 376–382. [Google Scholar] [PubMed]
- Grignani, G.; Pacchiarini, L.; Almasio, P.; Pagliarino, M.; Gamba, G.; Rizzo, S.C.; Ascari, E. Characterization of the platelet-aggregating activity of cancer cells with different metastatic potential. Int. J. Cancer 1986, 38, 237–244. [Google Scholar] [CrossRef] [PubMed]
- Pacchiarini, L.; Zucchella, M.; Milanesi, G.; Tacconi, F.; Bonomi, E.; Canevari, A.; Grignani, G. Thromboxane production by platelets during tumor cell-induced platelet activation. Invasion Metastasis 1991, 11, 102–109. [Google Scholar] [PubMed]
- Tzanakakis, G.N.; Krambovitis, E.; Tsatsakis, A.M.; Vezeridis, M.P. The preventive effect of ketoconazole on experimental metastasis from a human pancreatic carcinoma may be related to its effect on prostaglandin synthesis. Int. J. Gastrointest. Cancer 2002, 32, 23–30. [Google Scholar] [CrossRef]
- De Leval, X.; Benoit, V.; Delarge, J.; Julemont, F.; Masereel, B.; Pirotte, B.; Merville, M.P.; David, J.L.; Dogne, J.M. Pharmacological evaluation of the novel thromboxane modulator BM-567 (II/II). Effects of BM-567 on osteogenic sarcoma-cell-induced platelet aggregation. Prostaglandins Leukot. Essent. Fat. Acids 2003, 68, 55–59. [Google Scholar] [CrossRef]
- Steinert, B.W.; Tang, D.G.; Grossi, I.M.; Umbarger, L.A.; Honn, K.V. Studies on the role of platelet eicosanoid metabolism and integrin αIIbβ3 in tumor-cell-induced platelet aggregation. Int. J. Cancer 1993, 54, 92–101. [Google Scholar] [CrossRef] [PubMed]
- Amirkhosravi, A.; Amaya, M.; Siddiqui, F.; Biggerstaff, J.P.; Meyer, T.V.; Francis, J.L. Blockade of GpIIb/IIIa inhibits the release of vascular endothelial growth factor (VEGF) from tumor cell-activated platelets and experimental metastasis. Platelets 1999, 10, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Bastida, E.; Almirall, L.; Jamieson, G.A.; Ordinas, A. Cell surface sialylation of two human tumor cell lines and its correlation with their platelet-activating activity. Cancer Res. 1987, 47, 1767–1770. [Google Scholar] [PubMed]
- Grossi, I.M.; Fitzgerald, L.A.; Kendall, A.; Taylor, J.D.; Sloane, B.F.; Honn, K.V. Inhibition of human tumor cell induced platelet aggregation by antibodies to platelet glycoproteins Ib and IIb/IIIa. Proc. Soc. Exp. Biol. Med. 1987, 186, 378–383. [Google Scholar] [CrossRef] [PubMed]
- Sheu, J.R.; Lin, C.H.; Chung, J.L.; Teng, C.M.; Huang, T.F. Triflavin, an Arg-Gly-Asp containing snake venom peptide, inhibits aggregation of human platelets induced by human hepatoma cell line. Thromb. Res. 1992, 66, 679–691. [Google Scholar] [CrossRef]
- Sheu, J.R.; Lin, C.H.; Peng, H.C.; Teng, C.M.; Huang, T.F. Triflavin, an Arg-Gly-Asp-containing peptide, inhibits tumor cell-induced platelet aggregation. Jpn. J. Cancer Res. 1993, 84, 1062–1071. [Google Scholar] [CrossRef] [PubMed]
- Lian, L.; Li, W.; Li, Z.Y.; Mao, Y.X.; Zhang, Y.T.; Zhao, Y.M.; Chen, K.; Duan, W.M.; Tao, M. Inhibition of MCF-7 breast cancer cell-induced platelet aggregation using a combination of antiplatelet drugs. Oncol. Lett. 2013, 5, 675–680. [Google Scholar] [CrossRef] [PubMed]
- Bastida, E.; Almirall, L.; Ordinas, A. Tumor-cell-induced platelet aggregation is a glycoprotein-dependent and lipoxygenase-associated process. Int. J. Cancer 1987, 39, 760–763. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.W.; Hsieh, P.W.; Chang, Y.T.; Lu, M.H.; Huang, T.F.; Chong, K.Y.; Liao, H.R.; Cheng, J.C.; Tseng, C.P. Identification of a novel platelet antagonist that binds to CLEC-2 and suppresses podoplanin-induced platelet aggregation and cancer metastasis. Oncotarget 2015, 6, 42733–42748. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, C.A.; Deighton, N.M.; Jones, C.R.; Reid, J.L. Changes in rabbit platelet α and β adrenoceptor number and platelet aggregation. Eur. J. Pharmacol. 1986, 130, 145–149. [Google Scholar] [CrossRef]
- Oleksowicz, L.; Mrowiec, Z.; Schwartz, E.; Khorshidi, M.; Dutcher, J.P.; Puszkin, E. Characterization of tumor-induced platelet aggregation: The role of immunorelated GPIb and GPIIb/IIIa expression by MCF-7 breast cancer cells. Thromb. Res. 1995, 79, 261–274. [Google Scholar] [CrossRef]
- Cavanaugh, P.G.; Sloane, B.F.; Honn, K.V. Role of the coagulation system in tumor-cell-induced platelet aggregation and metastasis. Haemostasis 1988, 18, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Jung, G.; Millon-Collard, R.; Abecassis, J. Procoagulant and aggregating platelet activities of human mammary tumor cells. Bull. Cancer 1991, 78, 253–260. [Google Scholar] [PubMed]
- Pacchiarini, L.; Zucchella, M.; Eynard, A.R.; Grignani, G. Tumor cells induce platelet aggregation and intraplatelet calcium ion movements. Platelets 1993, 4, 275–279. [Google Scholar] [CrossRef] [PubMed]
- Heinmoller, E.; Schropp, T.; Kisker, O.; Simon, B.; Seitz, R.; Weinel, R.J. Tumor cell-induced platelet aggregation in vitro by human pancreatic cancer cell lines. Scand. J. Gastroenterol. 1995, 30, 1008–1016. [Google Scholar] [CrossRef] [PubMed]
- Sugimoto, Y.; Watanabe, M.; Oh-hara, T.; Sato, S.; Isoe, T.; Tsuruo, T. Suppression of experimental lung colonization of a metastatic variant of murine colon adenocarcinoma 26 by a monoclonal antibody 8F11 inhibiting tumor cell-induced platelet aggregation. Cancer Res. 1991, 51, 921–925. [Google Scholar] [PubMed]
- Okoshi, H.; Hakomori, S.; Nisar, M.; Zhou, Q.H.; Kimura, S.; Tashiro, K.; Igarashi, Y. Cell membrane signaling as target in cancer therapy. II: Inhibitory effect of N,N,N-trimethylsphingosine on metastatic potential of murine B16 melanoma cell line through blocking of tumor cell-dependent platelet aggregation. Cancer Res. 1991, 51, 6019–6024. [Google Scholar] [PubMed]
- Mahalingam, M.; Ugen, K.E.; Kao, K.J.; Klein, P.A. Functional role of platelets in experimental metastasis studied with cloned murine fibrosarcoma cell variants. Cancer Res. 1988, 48, 1460–1464. [Google Scholar] [PubMed]
- Tang, D.G.; Onoda, J.M.; Steinert, B.W.; Grossi, I.M.; Nelson, K.K.; Umbarger, L.; Diglio, C.A.; Taylor, J.D.; Honn, K.V. Phenotypic properties of cultured tumor cells: Integrin αIIbβ3 expression, tumor-cell-induced platelet aggregation, and tumor-cell adhesion to endothelium as important parameters of experimental metastasis. Int. J. Cancer 1993, 54, 338–347. [Google Scholar] [CrossRef] [PubMed]
- Medina, C.; Jurasz, P.; Santos-Martinez, M.J.; Jeong, S.S.; Mitsky, T.; Chen, R.; Radomski, M.W. Platelet aggregation-induced by caco-2 cells: Regulation by matrix metalloproteinase-2 and adenosine diphosphate. J. Pharmacol. Exp. Ther. 2006, 317, 739–745. [Google Scholar] [CrossRef] [PubMed]
- Kunita, A.; Kashima, T.G.; Morishita, Y.; Fukayama, M.; Kato, Y.; Tsuruo, T.; Fujita, N. The platelet aggregation-inducing factor aggrus/podoplanin promotes pulmonary metastasis. Am. J. Pathol. 2007, 170, 1337–1347. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.D.; Lee, Y.J.; Baik, J.S.; Han, J.Y.; Lee, C.G.; Heo, K.; Park, Y.S.; Kim, J.S.; Ji, H.D.; Park, S.I.; et al. Baicalein inhibits agonist- and tumor cell-induced platelet aggregation while suppressing pulmonary tumor metastasis via cAMP-mediated VASP phosphorylation along with impaired MAPKs and PI3K-Akt activation. Biochem. Pharmacol. 2014, 92, 251–265. [Google Scholar] [CrossRef] [PubMed]
- Amirkhosravi, A.; Mousa, S.A.; Amaya, M.; Blaydes, S.; Desai, H.; Meyer, T.; Francis, J.L. Inhibition of tumor cell-induced platelet aggregation and lung metastasis by the oral GpIIb/IIIa antagonist XV454. Thromb. Haemost. 2003, 90, 549–554. [Google Scholar] [CrossRef] [PubMed]
- Bakewell, S.J.; Nestor, P.; Prasad, S.; Tomasson, M.H.; Dowland, N.; Mehrotra, M.; Scarborough, R.; Kanter, J.; Abe, K.; Phillips, D.; et al. Platelet and osteoclast β3 integrins are critical for bone metastasis. Proc. Nat. Acad. Sci. USA 2003, 100, 14205–14210. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Dang, S.; Hong, T.; Tang, J.; Fan, J.; Bu, D.; Sun, Y.; Wang, Z.; Wisniewski, T. A humanized single-chain antibody against β3 integrin inhibits pulmonary metastasis by preferentially fragmenting activated platelets in the tumor microenvironment. Blood 2012, 120, 2889–2898. [Google Scholar] [CrossRef] [PubMed]
- Trikha, M.; Zhou, Z.; Timar, J.; Raso, E.; Kennel, M.; Emmell, E.; Nakada, M.T. Multiple roles for platelet GPIIb/IIIa and αvβ3 integrins in tumor growth, angiogenesis, and metastasis. Cancer Res. 2002, 62, 2824–2833. [Google Scholar] [PubMed]
- Schror, K.; Weber, A.A. Comparative pharmacology of GP IIb/IIIa antagonists. J. Thromb. Haemost. 2003, 15, 71–80. [Google Scholar] [CrossRef]
- Echtler, K.; Konrad, I.; Lorenz, M.; Schneider, S.; Hofmaier, S.; Plenagl, F.; Stark, K.; Czermak, T.; Tirniceriu, A.; Eichhorn, M.; et al. Platelet GPIIb supports initial pulmonary retention but inhibits subsequent proliferation of melanoma cells during hematogenic metastasis. PloS ONE 2017, 12, e0172788. [Google Scholar] [CrossRef] [PubMed]
- Ugen, K.E.; Mahalingam, M.; Klein, P.A.; Kao, K.J. Inhibition of tumor cell-induced platelet aggregation and experimental tumor metastasis by the synthetic Gly-Arg-Gly-Asp-Ser peptide. J. Nat. Cancer Inst. 1988, 80, 1461–1466. [Google Scholar] [CrossRef] [PubMed]
- Soszka, T.; Knudsen, K.A.; Beviglia, L.; Rossi, C.; Poggi, A.; Niewiarowski, S. Inhibition of murine melanoma cell-matrix adhesion and experimental metastasis by albolabrin, an RGD-containing peptide isolated from the venom of Trimeresurus albolabris. Exp. Cell Res. 1991, 196, 6–12. [Google Scholar] [CrossRef]
- Schneider, D.J. Anti-platelet therapy: Glycoprotein IIb-IIIa antagonists. Br. J. Clin. Pharmacol. 2011, 72, 672–682. [Google Scholar] [CrossRef] [PubMed]
- Chew, D.P.; Bhatt, D.L.; Sapp, S.; Topol, E.J. Increased mortality with oral platelet glycoprotein IIb/IIIa antagonists: A meta-analysis of phase III multicenter randomized trials. Circulation 2001, 103, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Bassler, N.; Loeffler, C.; Mangin, P.; Yuan, Y.; Schwarz, M.; Hagemeyer, C.E.; Eisenhardt, S.U.; Ahrens, I.; Bode, C.; Jackson, S.P.; et al. A mechanistic model for paradoxical platelet activation by ligand-mimetic αIIbβ3 (GPIIb/IIIa) antagonists. Arterioscler. Thromb. Vasc. Biol. 2007, 27, e9–e15. [Google Scholar] [CrossRef] [PubMed]
- Hagemeyer, C.E.; Peter, K. Targeting the platelet integrin GPIIb/IIIa. Curr. Pharm. Des. 2010, 16, 4119–4133. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, P.C.; Peter, K. GPIIb/IIIa inhibitors: From bench to bedside and back to bench again. Thromb. Haemost. 2012, 107, 808–814. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, M.; Meade, G.; Stoll, P.; Ylanne, J.; Bassler, N.; Chen, Y.C.; Hagemeyer, C.E.; Ahrens, I.; Moran, N.; Kenny, D.; et al. Conformation-specific blockade of the integrin GPIIb/IIIa: A novel antiplatelet strategy that selectively targets activated platelets. Circ. Res. 2006, 99, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Vootukuri, S.; Shang, Y.; Negri, A.; Jiang, J.K.; Nedelman, M.; Diacovo, T.G.; Filizola, M.; Thomas, C.J.; Coller, B.S. RUC-4: A novel αIIbβ3 antagonist for prehospital therapy of myocardial infarction. Arterioscler. Thromb. Vasc. Biol. 2014, 34, 2321–2329. [Google Scholar] [CrossRef] [PubMed]
- Zhu, G.; Zhang, Q.; Reddy, E.C.; Carrim, N.; Chen, Y.; Xu, X.R.; Xu, M.; Wang, Y.; Hou, Y.; Ma, L.; et al. The integrin PSI domain has an endogenous thiol isomerase function and is a novel target for antiplatelet therapy. Blood 2017, 129, 1840–1854. [Google Scholar] [CrossRef] [PubMed]
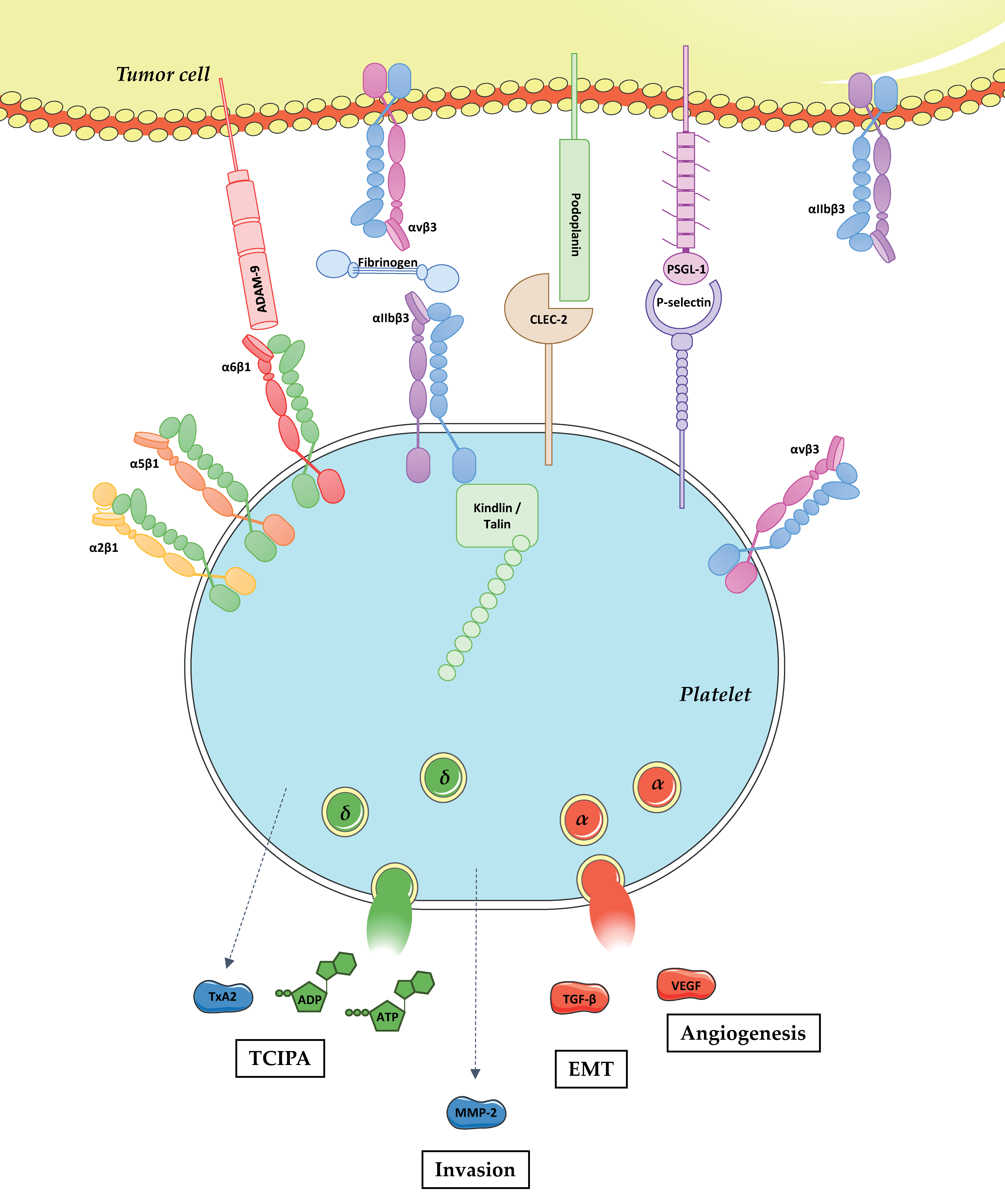

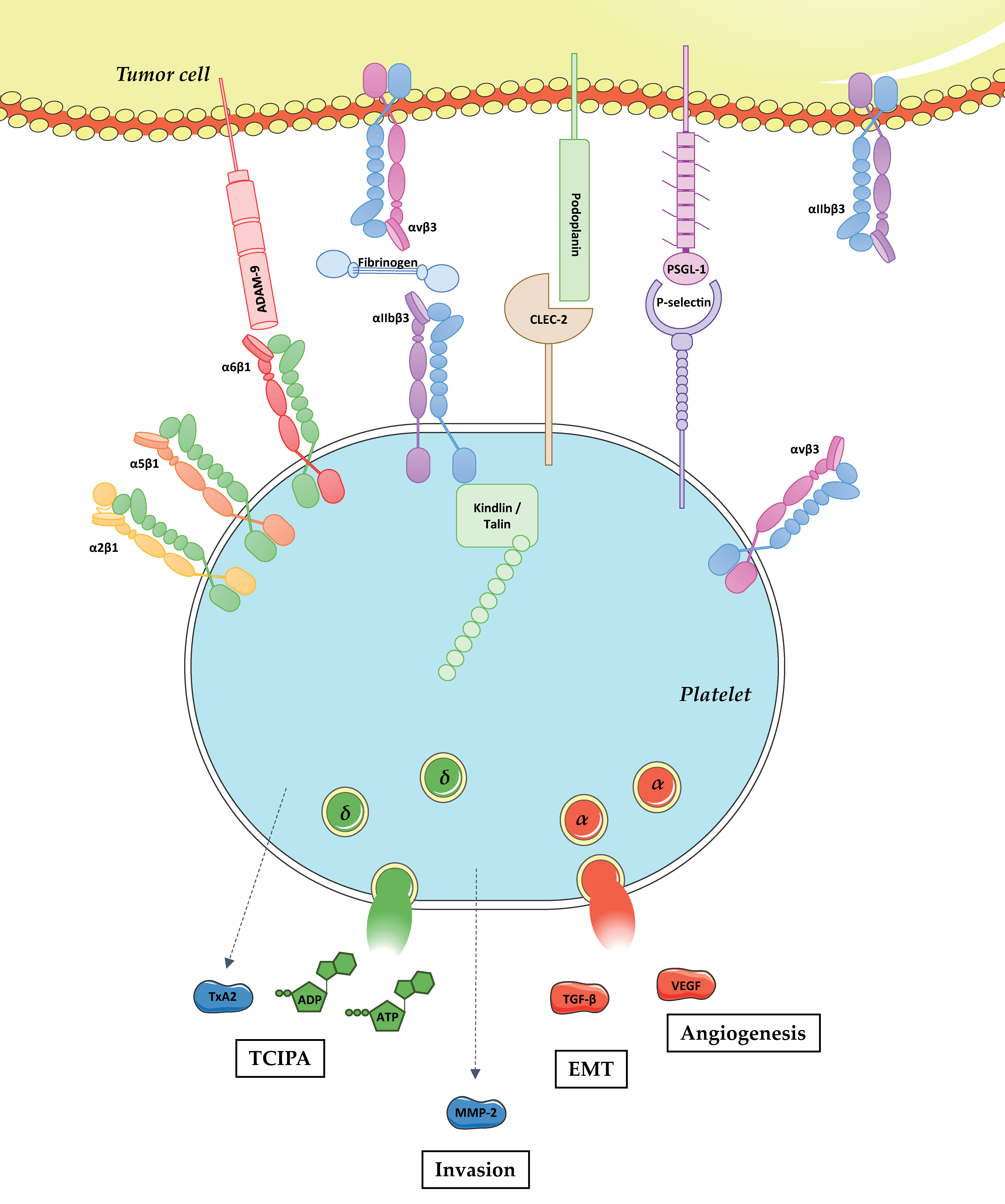{kind=link}
| Name | Nature of the Agent | Use | Inhibition of Platelet Aggregation | Inhibition of in Vivo Thrombus Formation | Activatory Effect on αIIbβ3 | Effect on Bleeding |
|---|---|---|---|---|---|---|
| Abciximab (ReoPro®) | chimeric Fab fragment derived from the murine monoclonal antibody 7E3 | Clinically used | √ | √ | √ | √ |
| Tirofiban (Aggrastat®) | non-peptide agent based on the RGD sequence | Clinically used | √ | √ | √ | √ |
| Eptifibatide (Integrilin®) | KGD-containing cyclic heptapeptide | Clinically used | √ | √ | √ | √ |
| RUC-4 | Low-molecular weight molecule | Used in pre-clinical studies | √ | √ | X | Not evaluated |
| scFv MA2 | Single-chain antibody directed against the activated form of αIIbβ3 | Used in pre-clinical studies | √ | √ | X | No impact on mouse tail-bleeding time |
| mAb anti-PSI | Monoclonal antibody against the β3 PSI domain | Used in pre-clinical studies | √ | √ | X | No impact on mouse tail-bleeding time |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lavergne, M.; Janus-Bell, E.; Schaff, M.; Gachet, C.; Mangin, P.H. Platelet Integrins in Tumor Metastasis: Do They Represent a Therapeutic Target? Cancers 2017, 9, 133. https://doi.org/10.3390/cancers9100133
Lavergne M, Janus-Bell E, Schaff M, Gachet C, Mangin PH. Platelet Integrins in Tumor Metastasis: Do They Represent a Therapeutic Target? Cancers. 2017; 9(10):133. https://doi.org/10.3390/cancers9100133
Chicago/Turabian StyleLavergne, Marion, Emily Janus-Bell, Mathieu Schaff, Christian Gachet, and Pierre H. Mangin. 2017. "Platelet Integrins in Tumor Metastasis: Do They Represent a Therapeutic Target?" Cancers 9, no. 10: 133. https://doi.org/10.3390/cancers9100133
APA StyleLavergne, M., Janus-Bell, E., Schaff, M., Gachet, C., & Mangin, P. H. (2017). Platelet Integrins in Tumor Metastasis: Do They Represent a Therapeutic Target? Cancers, 9(10), 133. https://doi.org/10.3390/cancers9100133