Immune Regulatory Activity of Vitamin D3 in Head and Neck Cancer


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
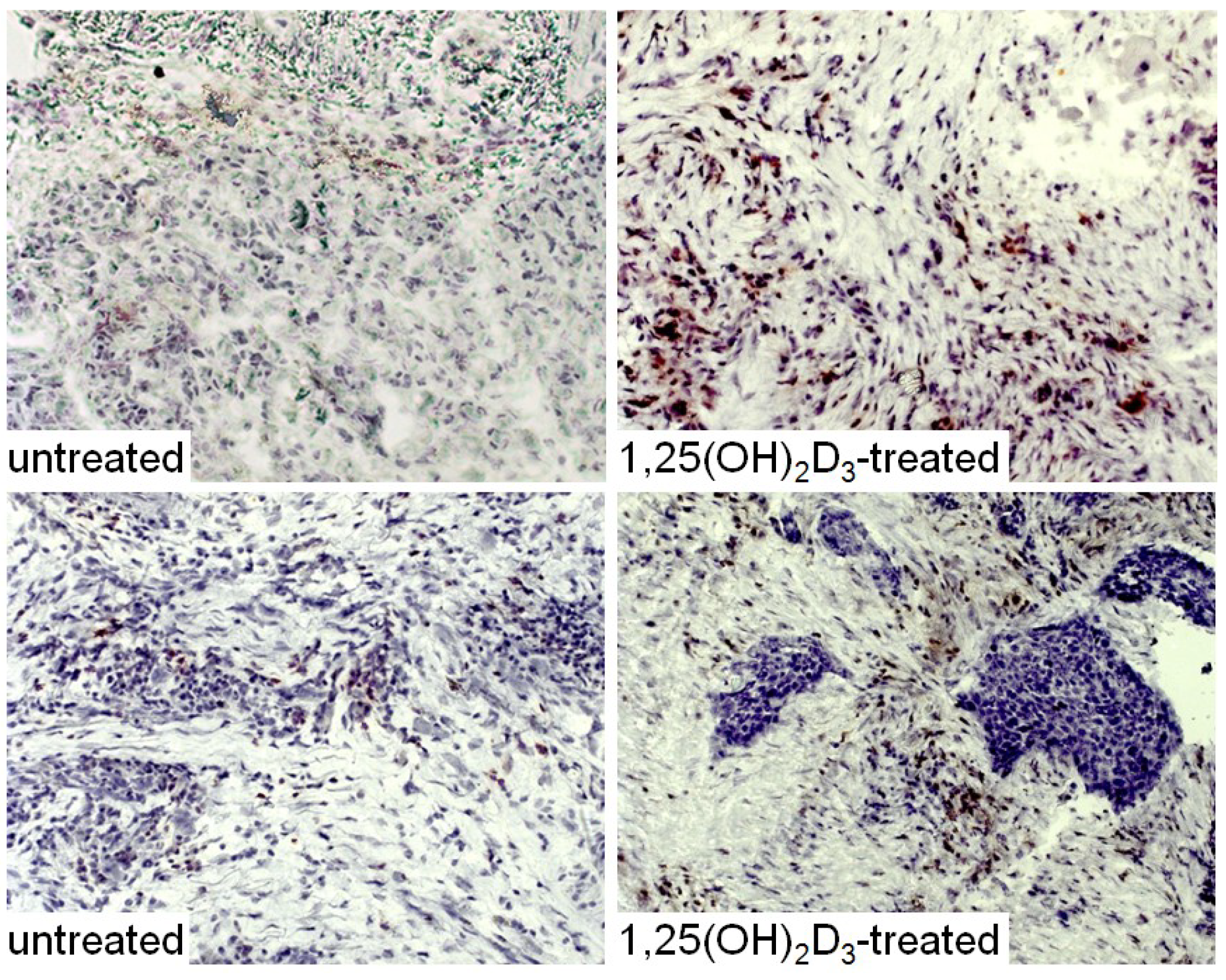{kind=link}
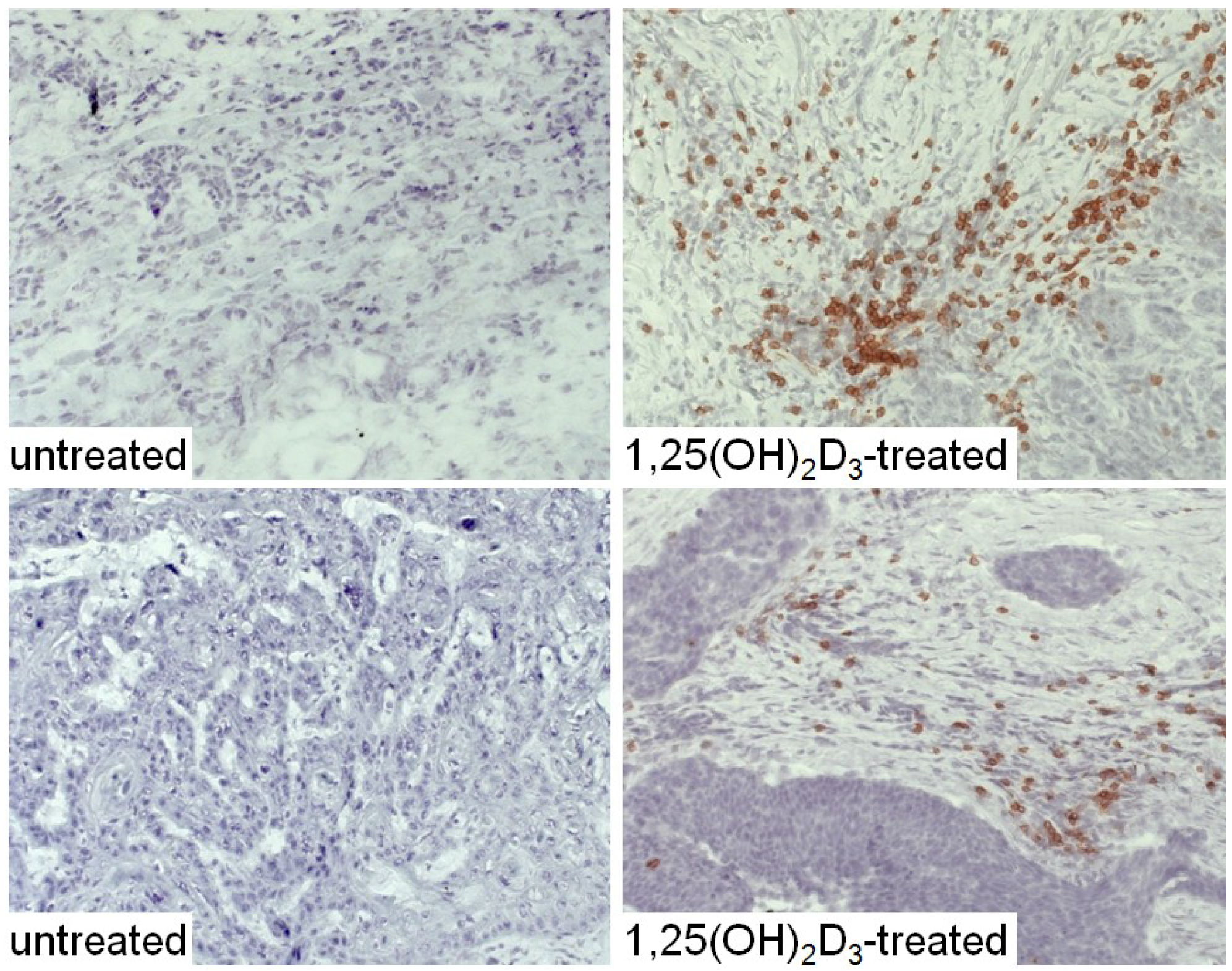{kind=link}
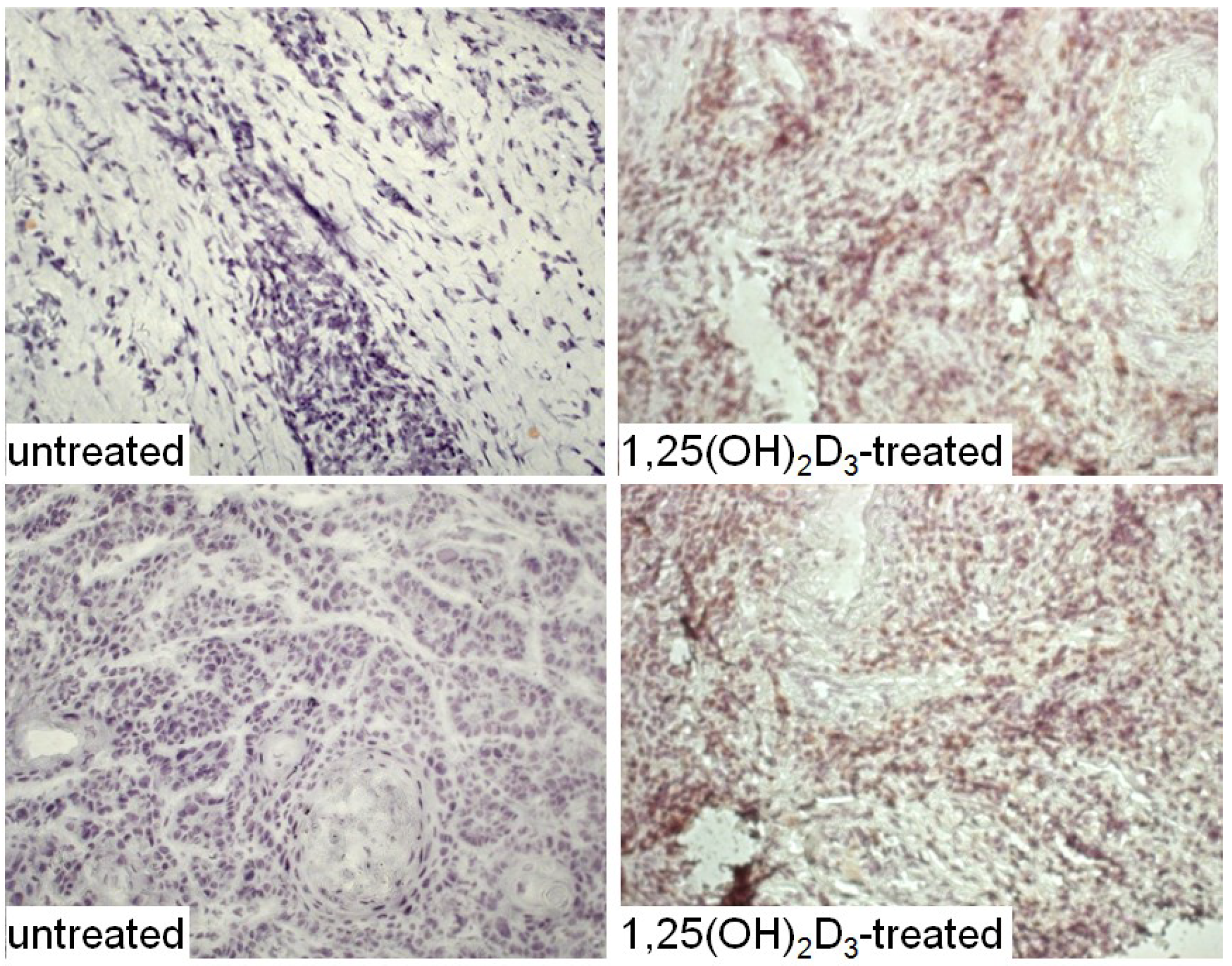{kind=link}
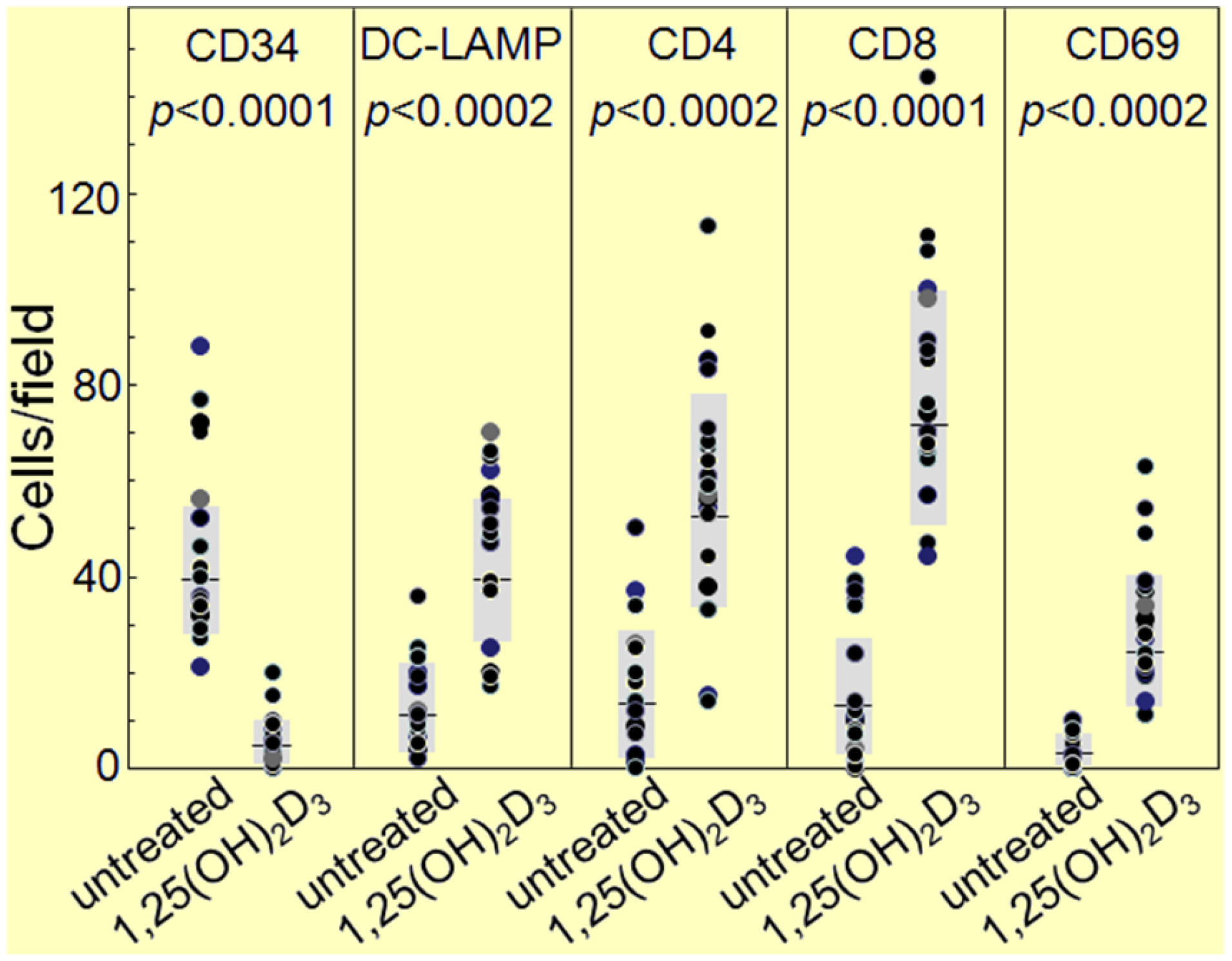{kind=link}
Abstract
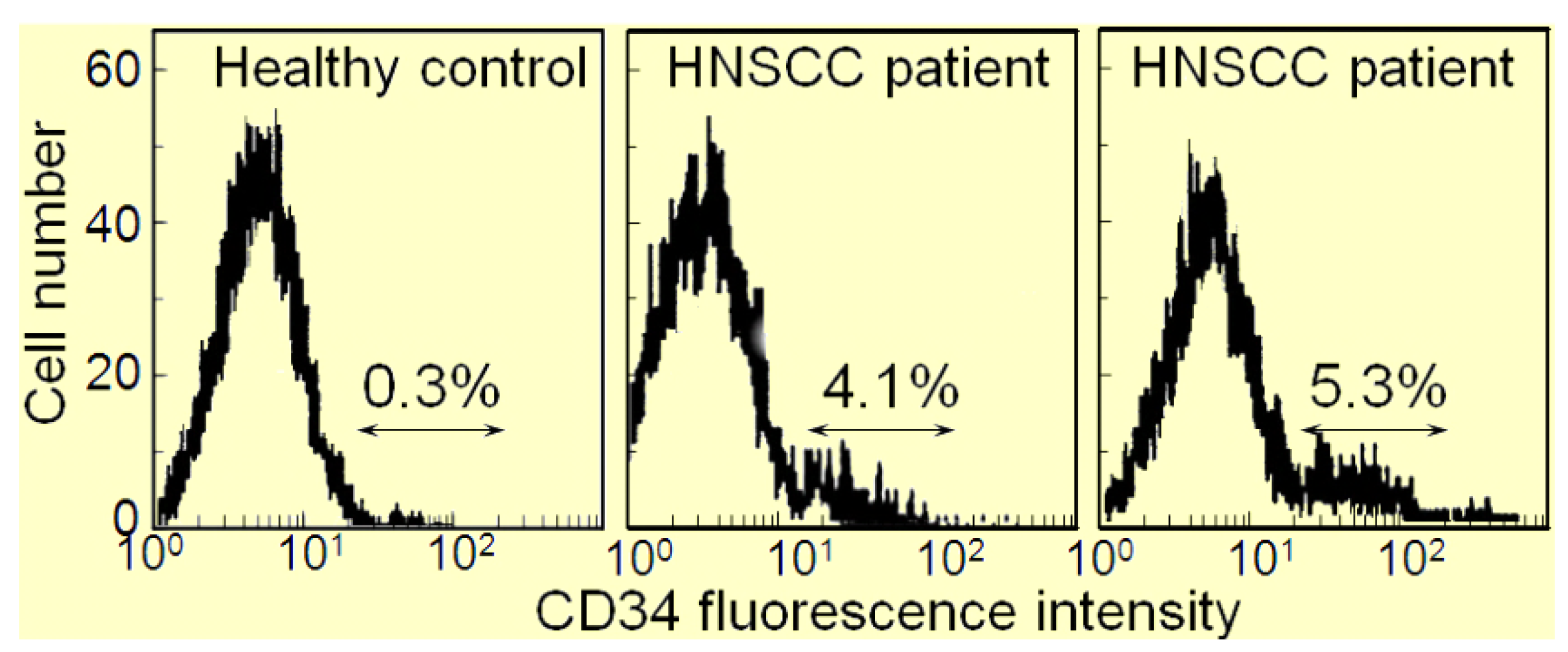
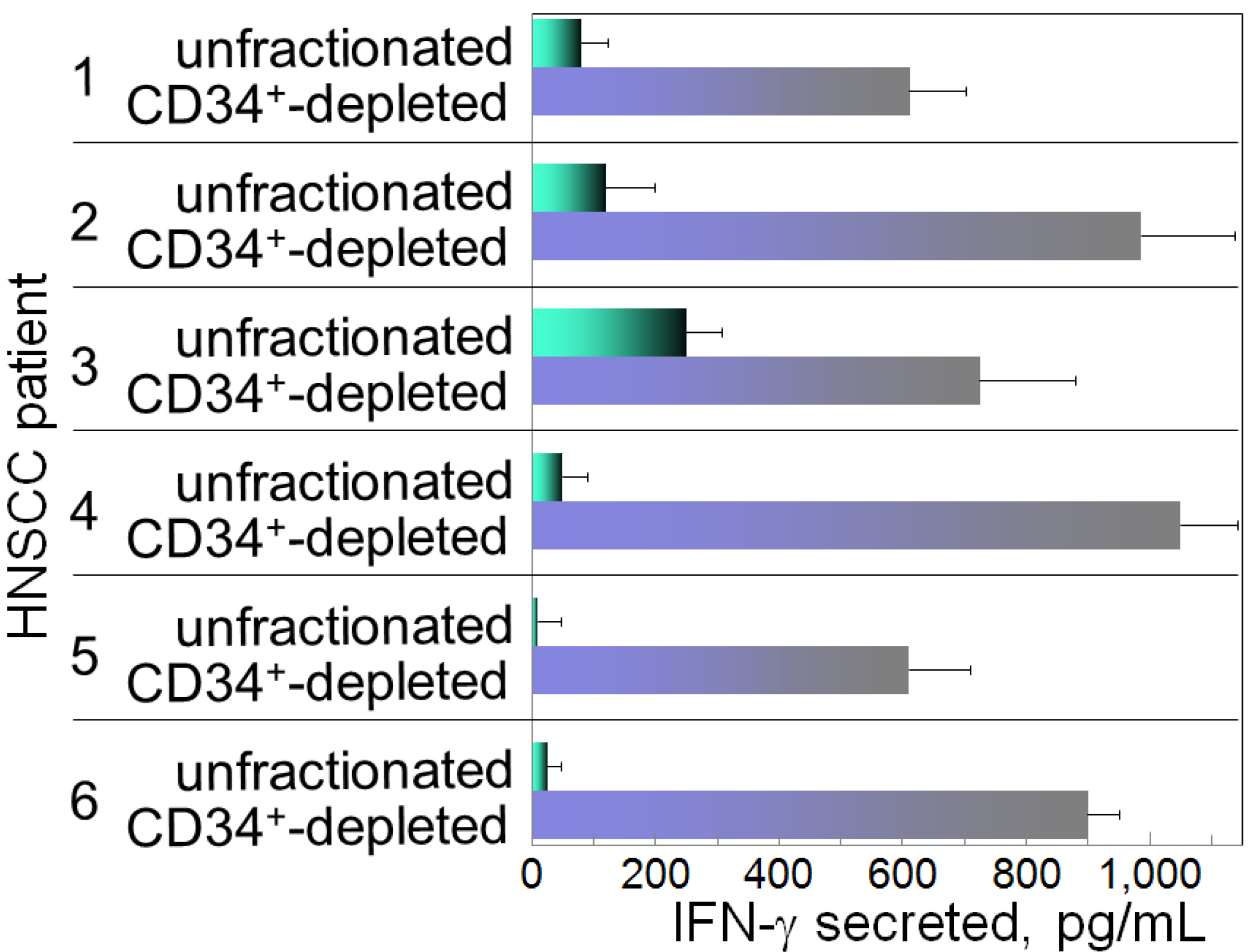
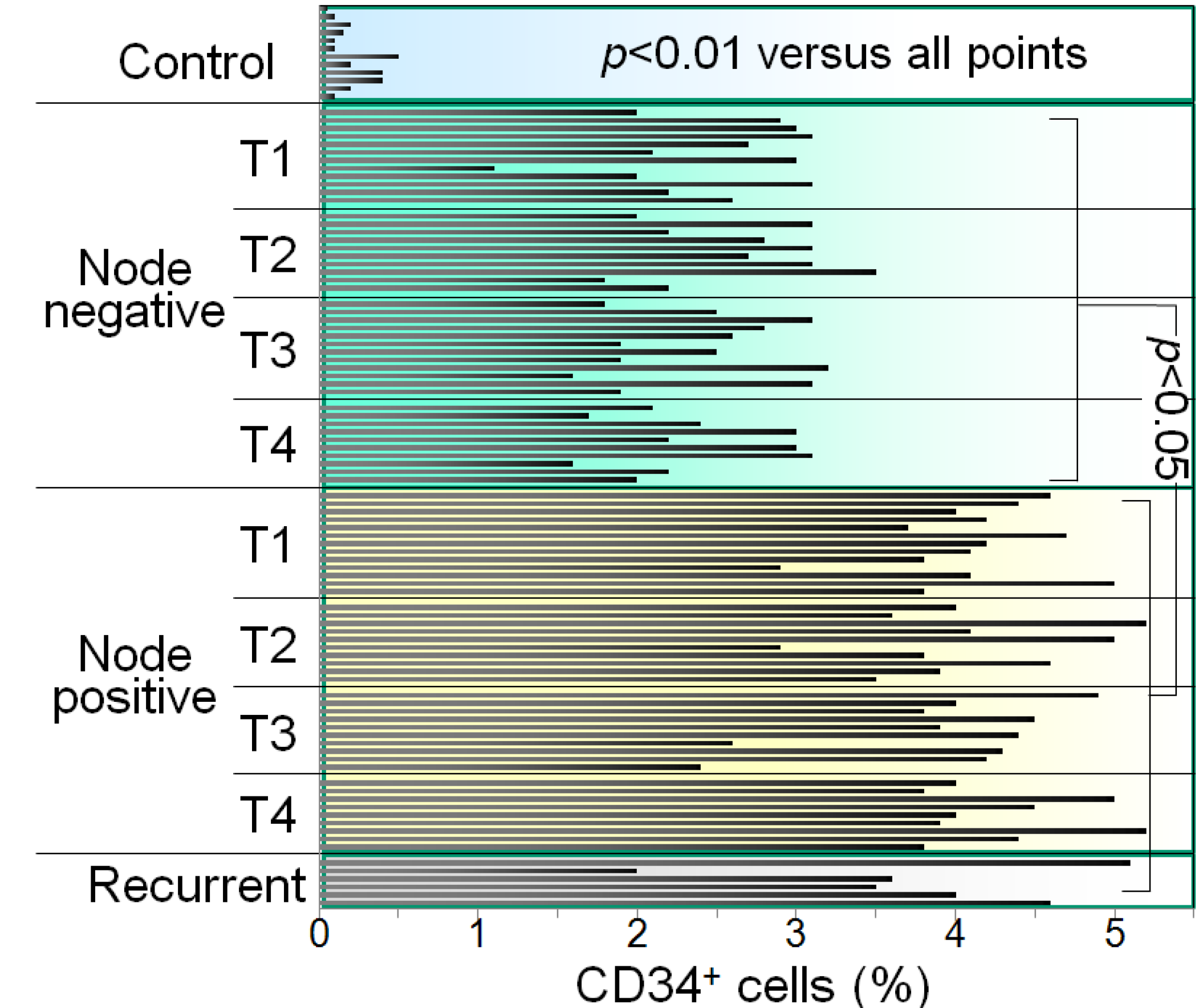
:1. Induction of Immune Inhibitory Cells by HNSCC
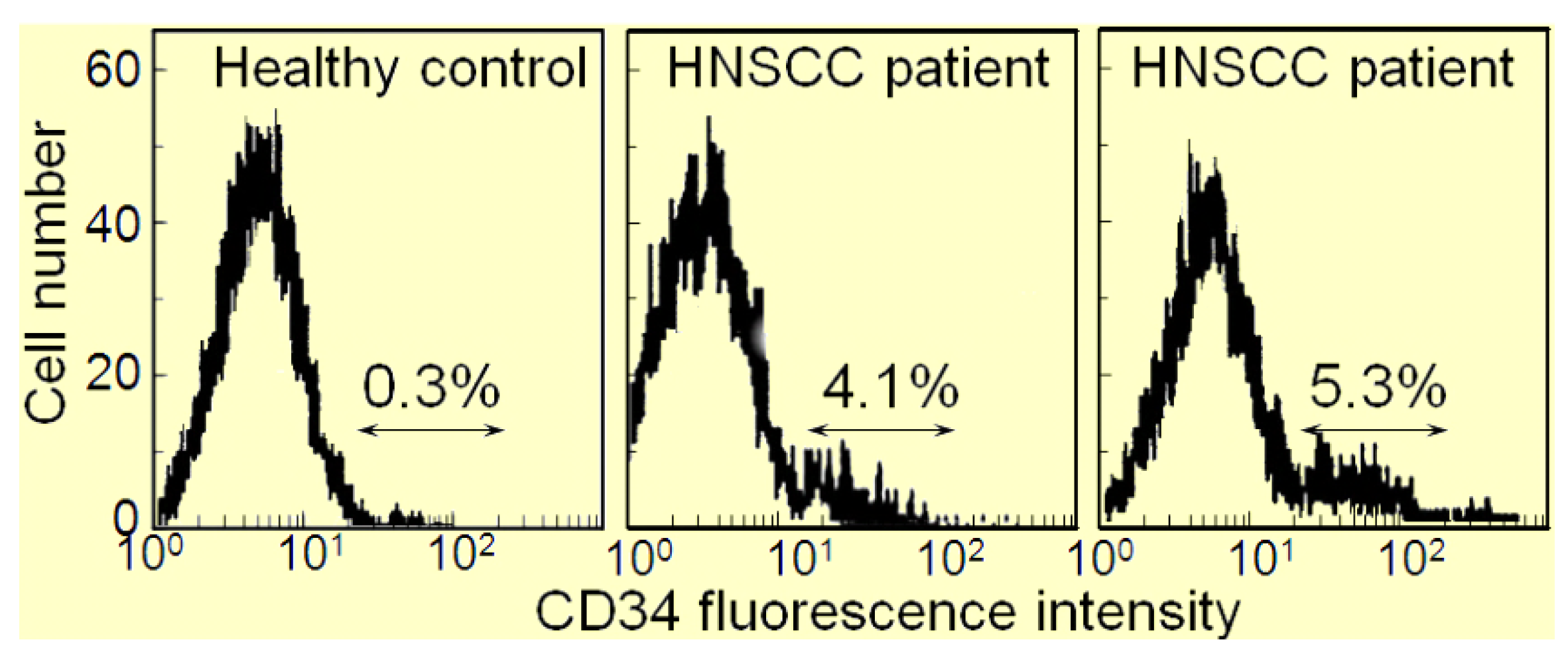
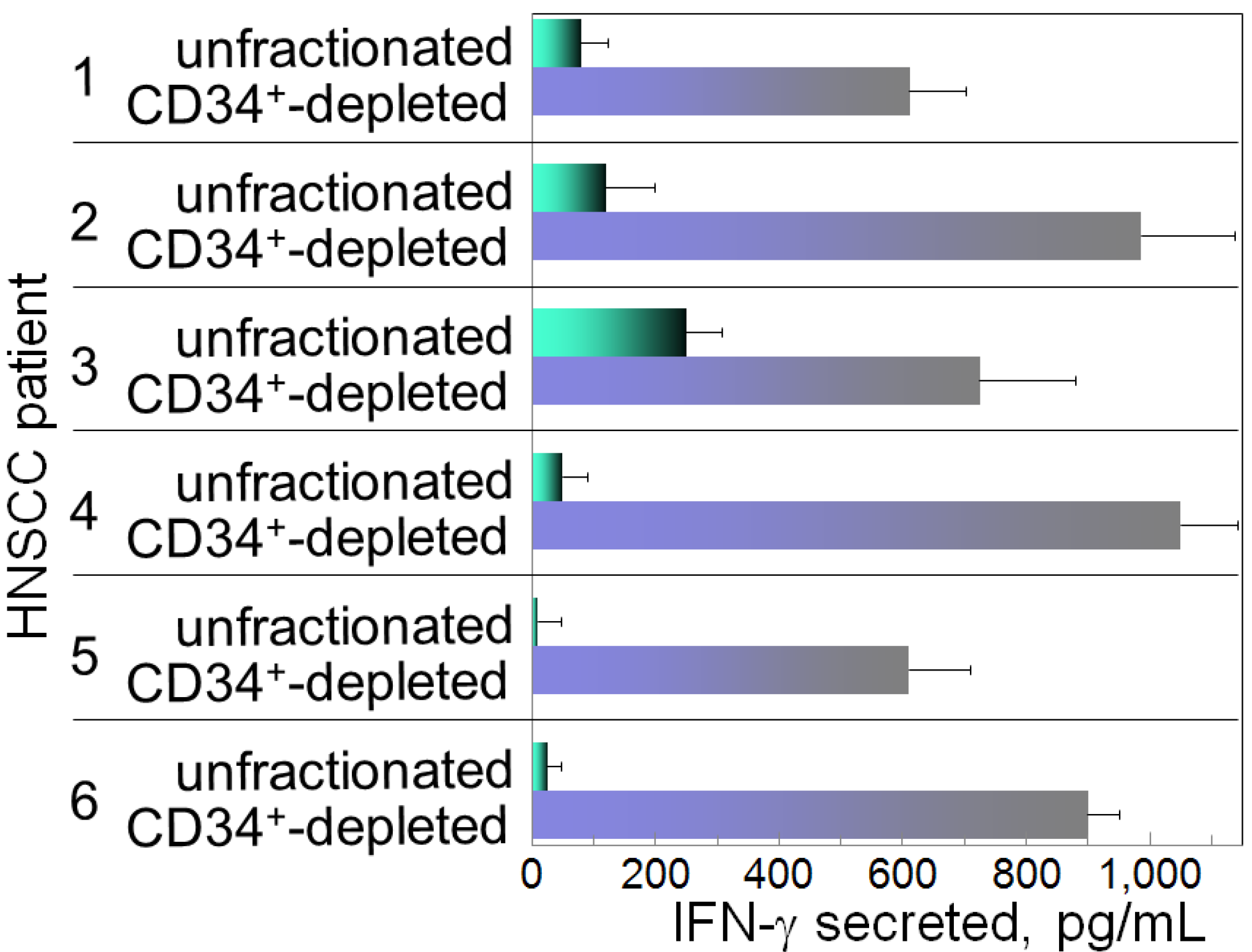
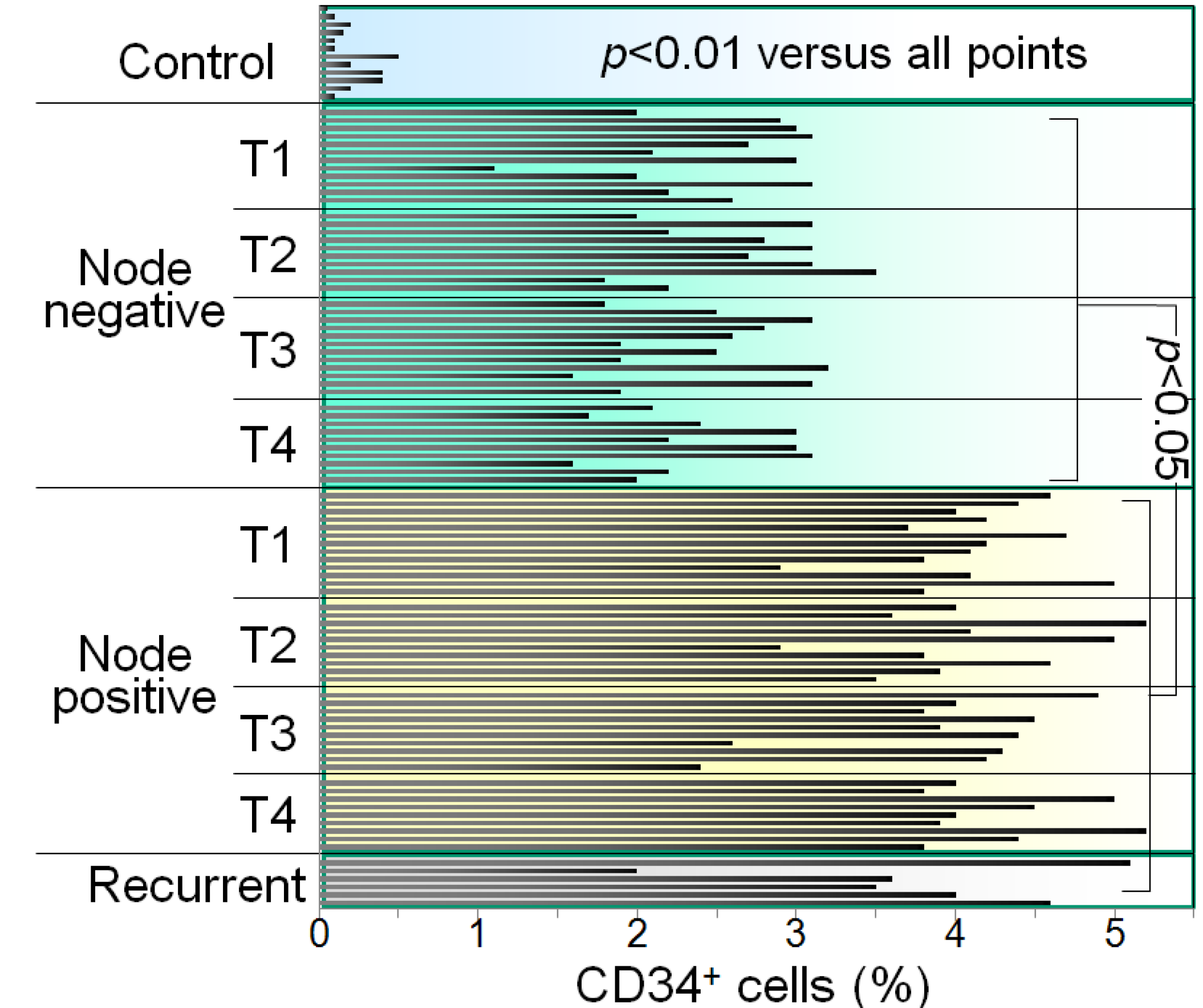
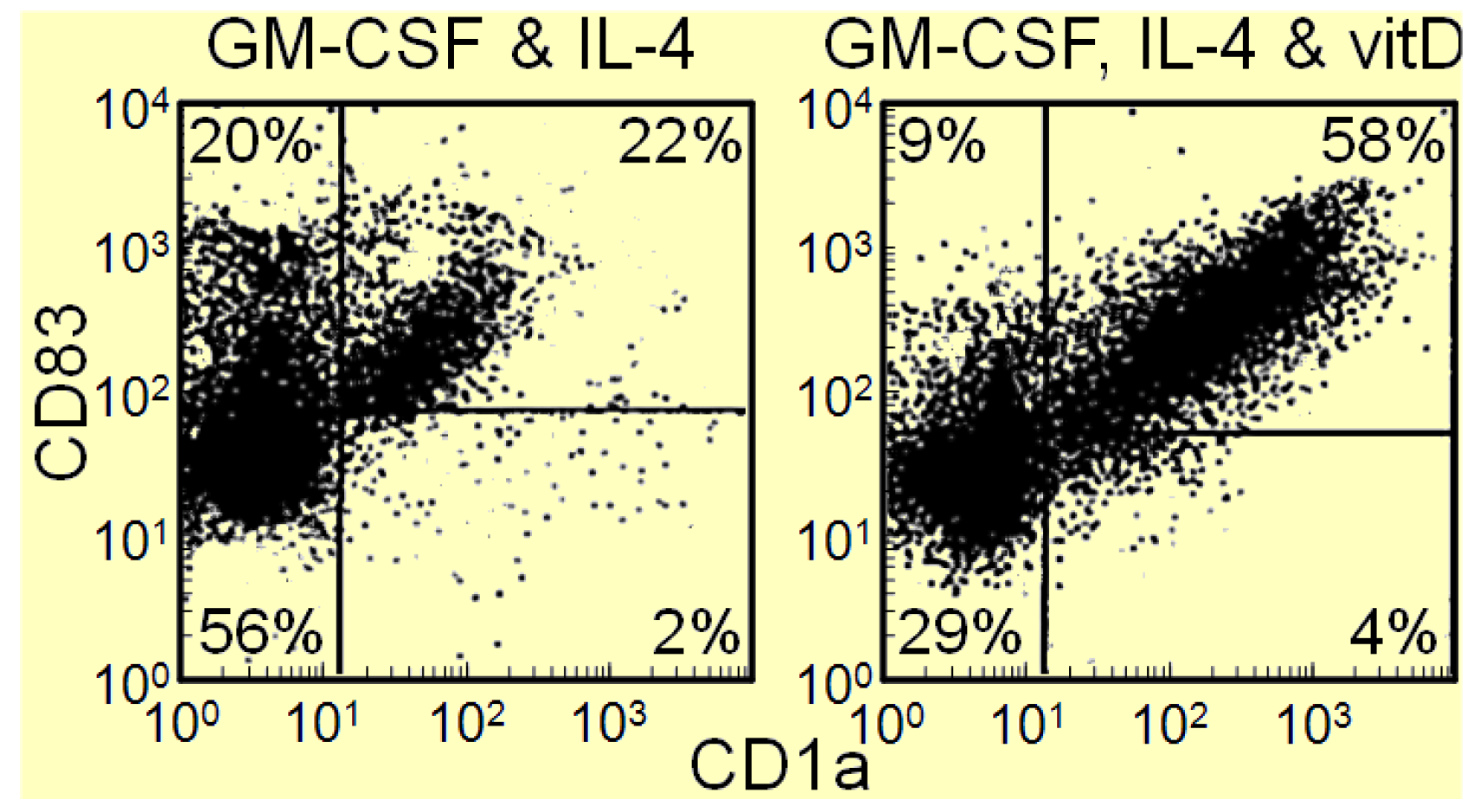
2. Overcoming Cancer-Induced Immune Inhibitory Mechanisms with Vitamin D
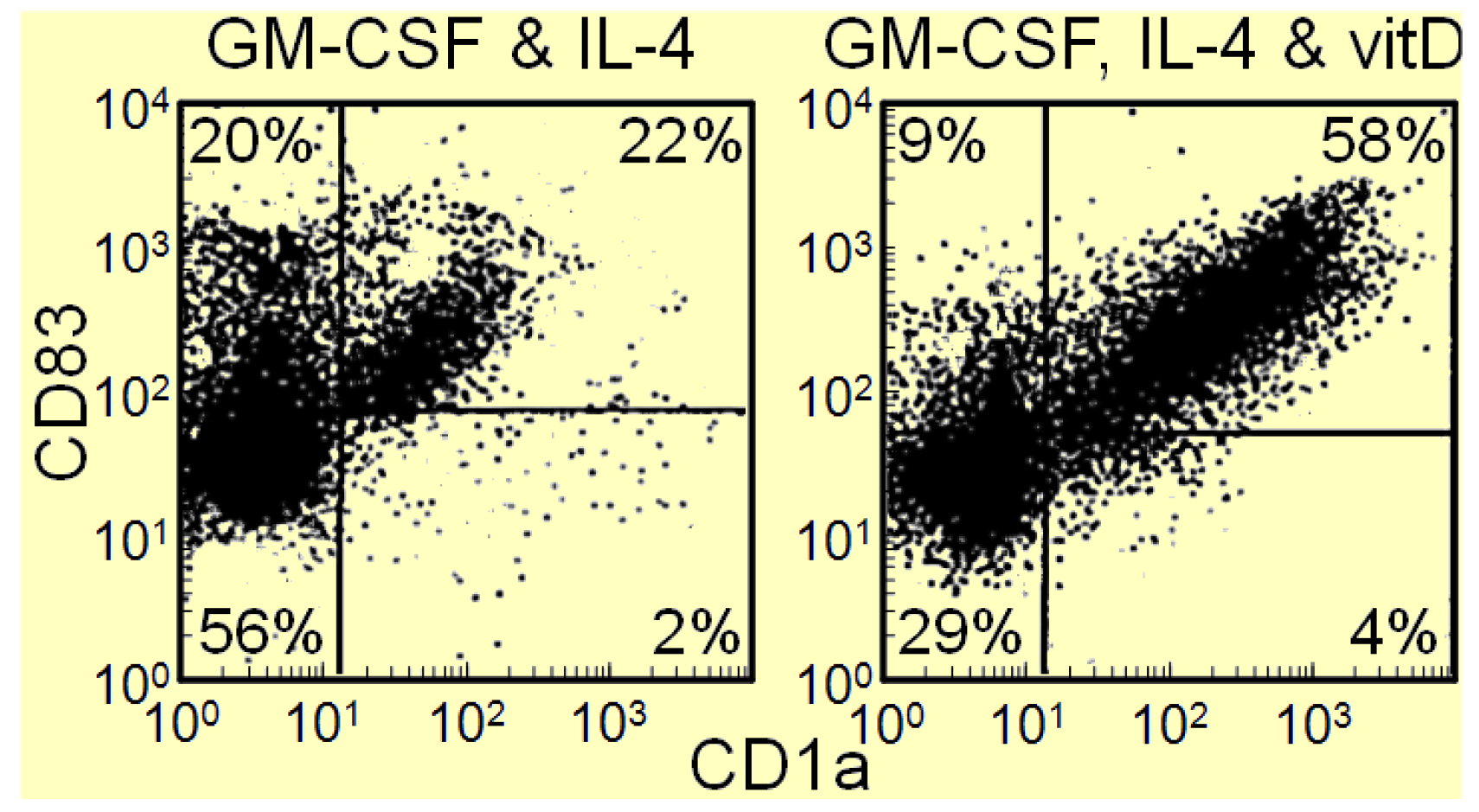
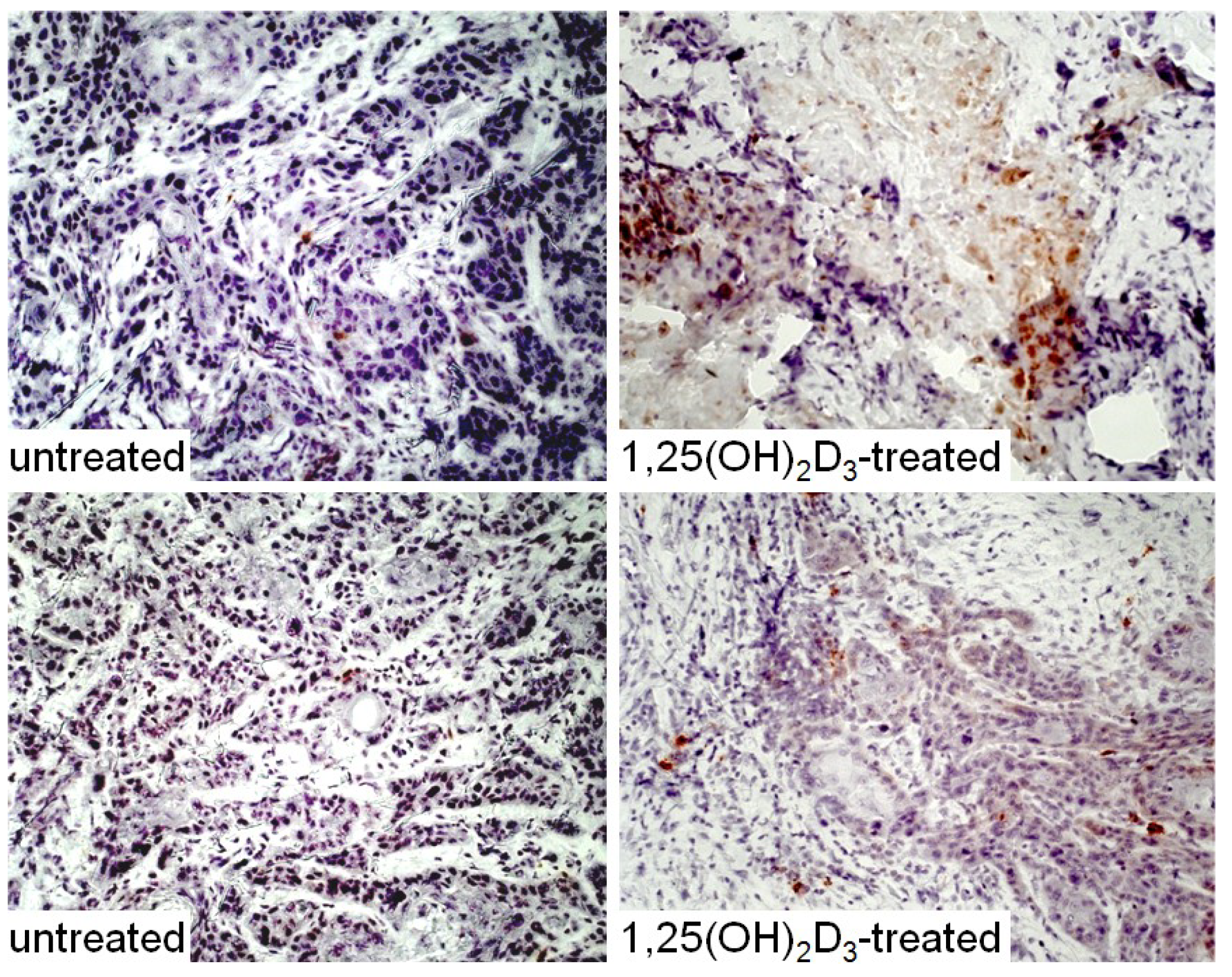
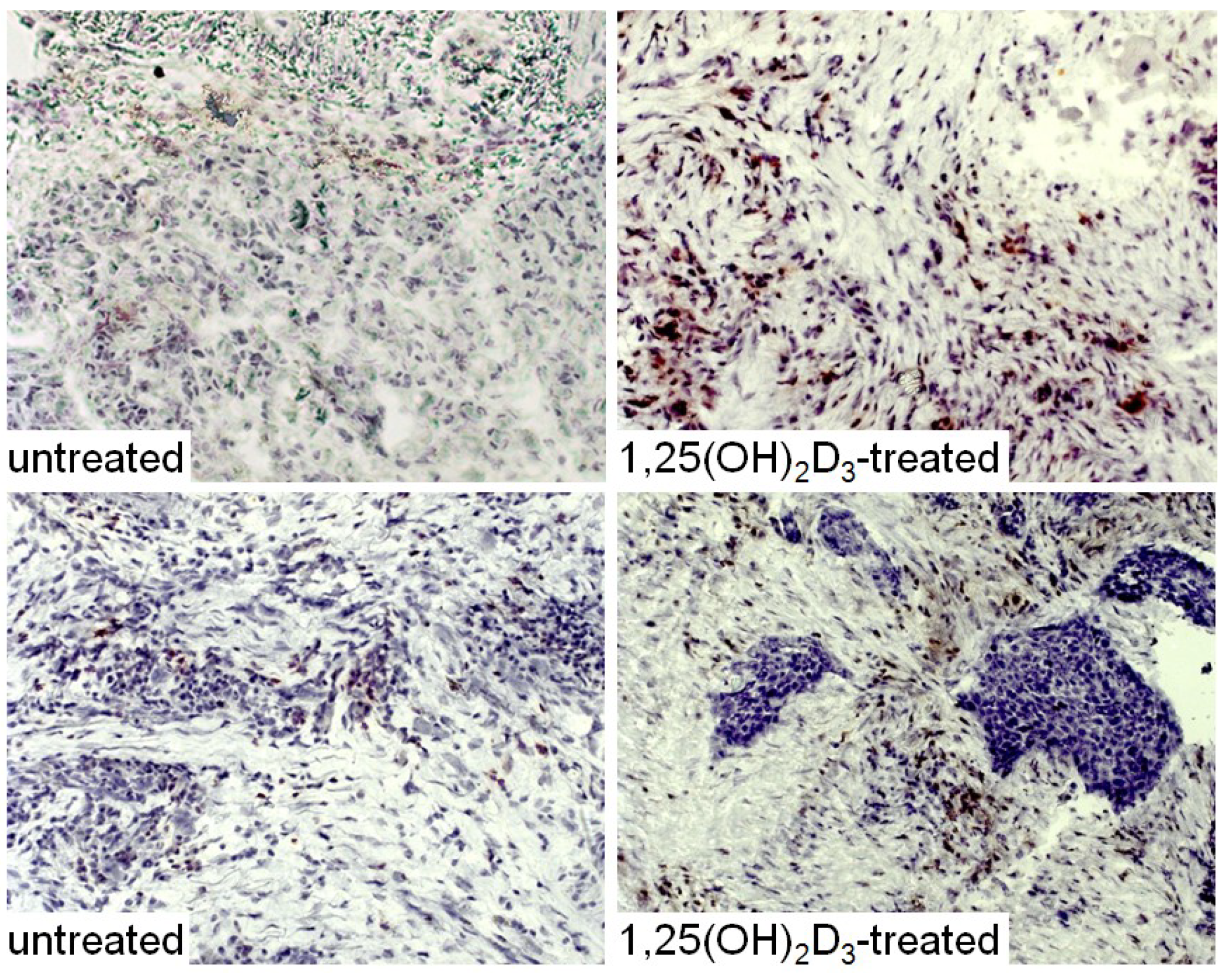
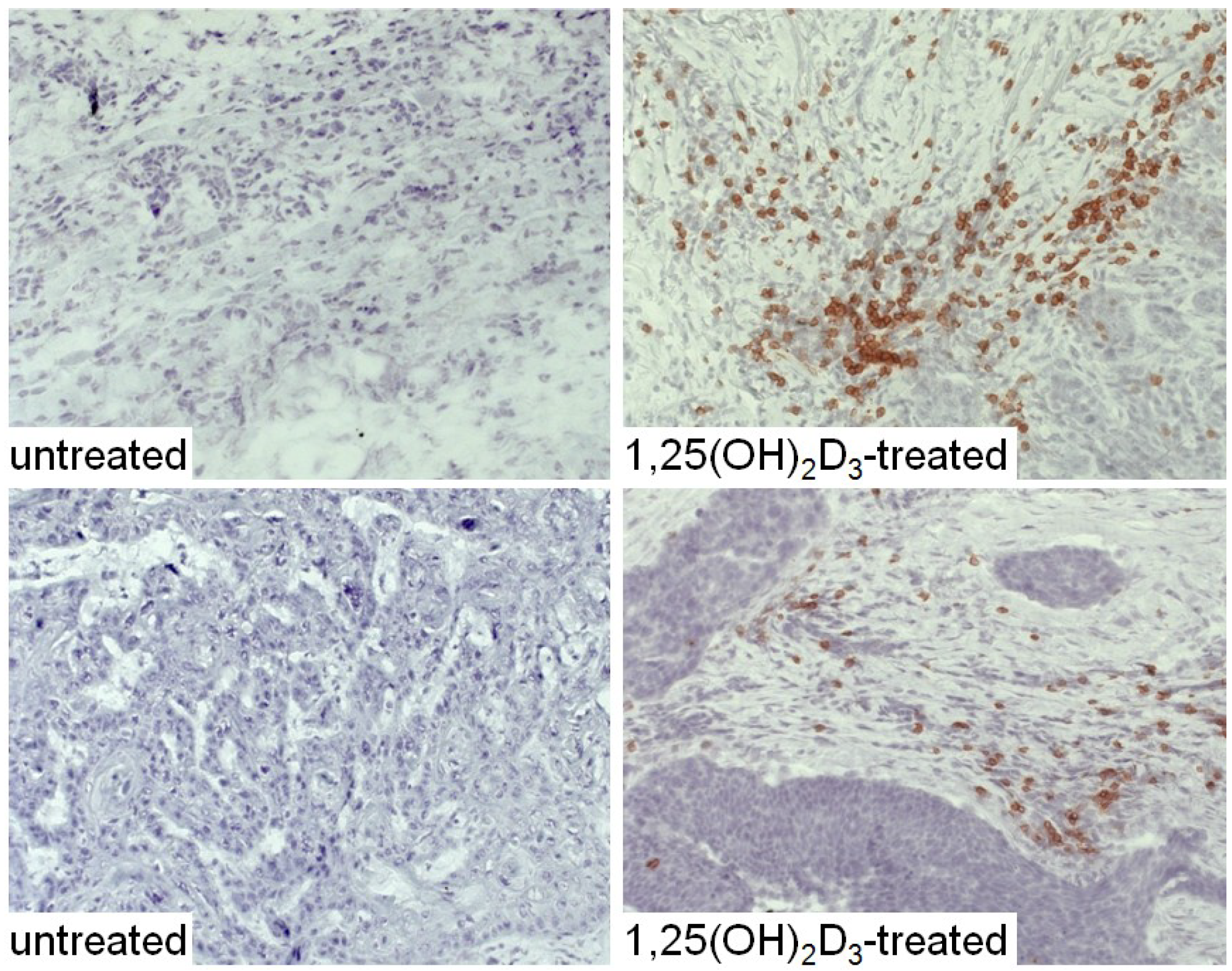
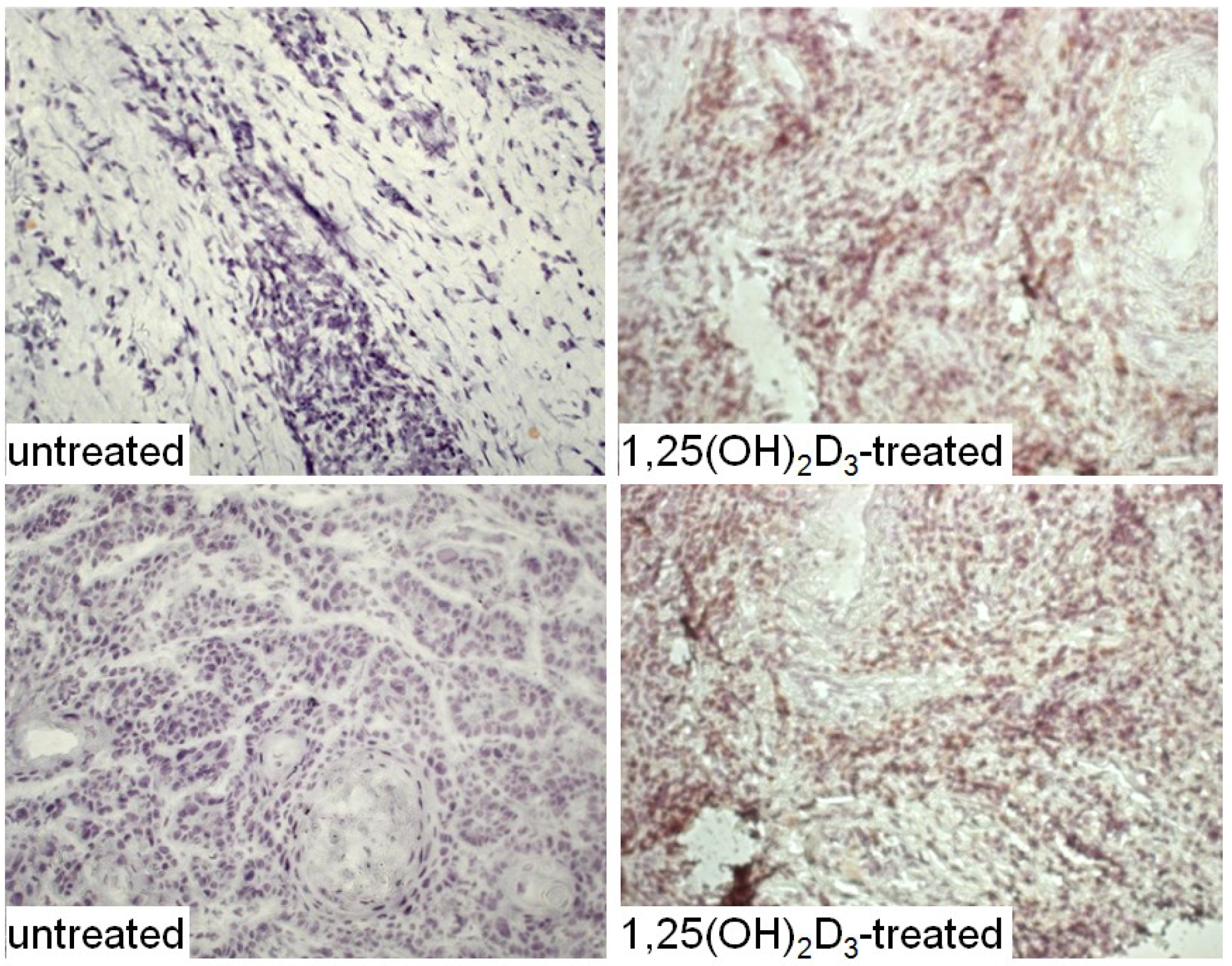
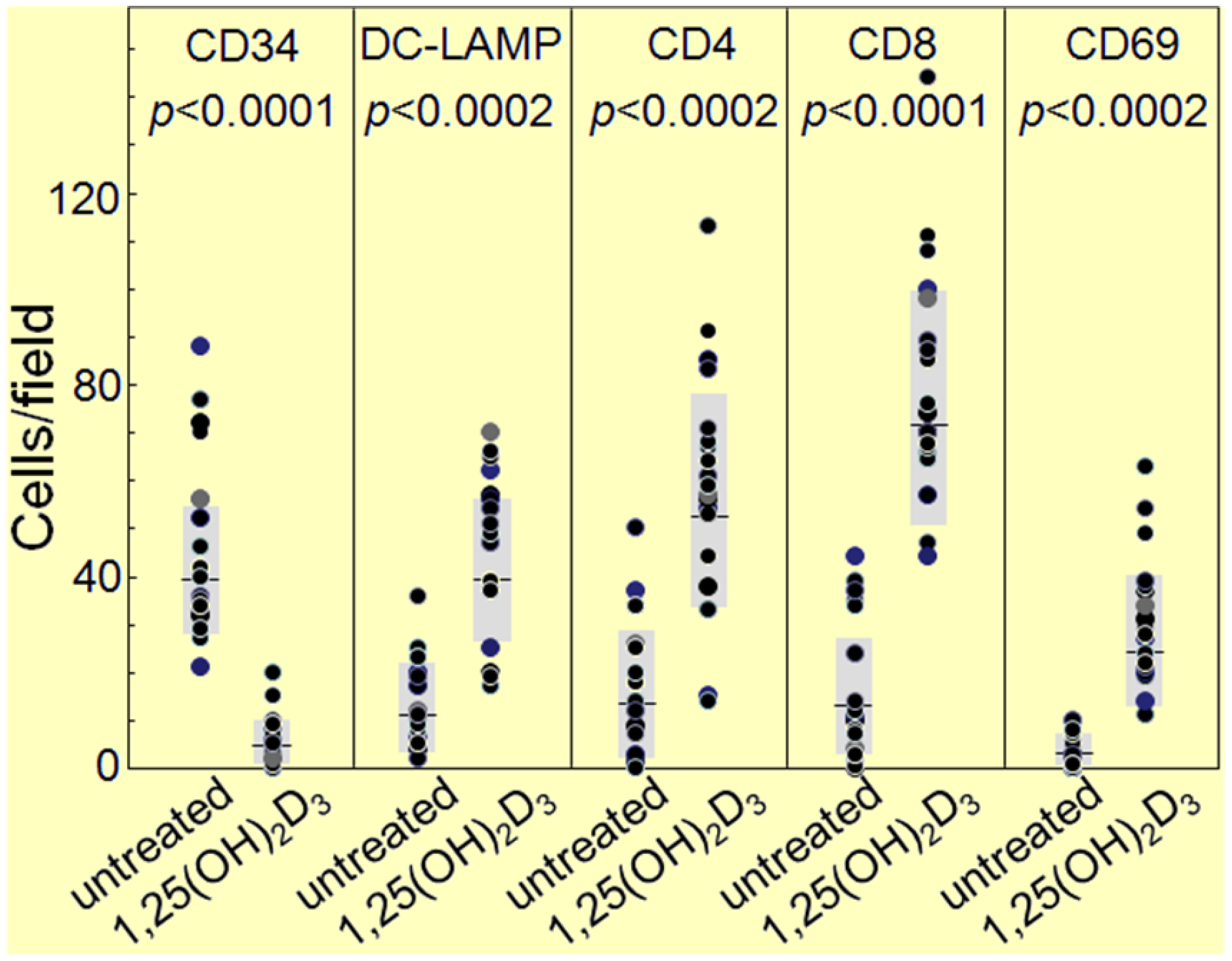
3. Interplay of Vitamin D and COX-2 in Immune Modulation in Cancer
4. Additional Immune Modulatory Mechanisms of Vitamin D That Can Impact on Cancer Development and Progression
5. Conclusions
Acknowledgments
Conflicts of Interest
References
- National Cancer Institute. Surveillance epidemiology and end results. Available online: http://seer.cancer.gov/statfacts/html/oralcav.html/ (accessed on 21 May 2013).
- Heimdal, J.H.; Aarstad, H.J.; Olofsson, J. Peripheral blood T-lymphocyte and monocyte function and survival in patients with head and neck carcinoma. Laryngoscope 2000, 110, 402–407. [Google Scholar] [CrossRef]
- Meneses, A.; Verastegui, E.; Barrera, J.L.; de la Garza, J.; Hadden, J.W. Lymph node histology in head and neck cancer: Impact of immunotherapy with IRX-2. Int. Immunol. 2003, 3, 1083–1091. [Google Scholar] [CrossRef]
- Shibuya, T.Y.; Wei, W.Z.; Zormeier, M.; Ensley, J.; Sakr, W.; Mathog, R.H.; Meleca, R.J.; Yoo, G.H.; June, C.H.; Levine, B.L.; et al. Anti-CD3/anti-CD28 bead stimulation overcomes CD3 unresponsiveness in patients with head and neck squamous cell carcinoma. Arch. Otolaryngol. 2000, 126, 473–479. [Google Scholar] [CrossRef]
- Almand, B.; Resser, J.R.; Lindman, B.; Nadaf, S.; Clark, J.I.; Kwon, E.D.; Carbone, D.P.; Gabrilovich, D.I. Clinical significance of defective dendritic cell differentiation in cancer. Clin. Cancer Res. 2000, 6, 1755–1766. [Google Scholar]
- Egan, J.E.; Quadrini, K.J.; Santiago-Schwarz, F.; Hadden, J.W.; Brandwein, H.J.; Signorelli, K.L. IRX-2, a novel in vivo immunotherapeutic, induces maturation and activation of human dendritic cells in vitro. J. Immunother. 2007, 30, 624–633. [Google Scholar] [CrossRef]
- Kacani, L.; Wurm, M.; Schwentner, I.; Andrle, J.; Schennach, H.; Sprinzl, G.M. Maturation of dendritic cells in the presence of living, apoptotic and necrotic tumour cells derived from squamous cell carcinoma of head and neck. Oral Oncol. 2005, 41, 17–24. [Google Scholar] [CrossRef]
- Heimdal, J.H.; Aarstad, H.J.; Klementsen, B.; Olofsson, J. Peripheral blood mononuclear cell (PBMC) responsiveness in patients with head and neck cancer in relation to tumour stage and prognosis. Acta OtoLaryngol. 1999, 119, 281–284. [Google Scholar] [CrossRef]
- Millrud, C.R.; Mansson Kvarnhammar, A.; Uddman, R.; Bjornsson, S.; Riesbeck, K.; Cardell, L.O. The activation pattern of blood leukocytes in head and neck squamous cell carcinoma is correlated to survival. PLoS One 2012, 7, e51120. [Google Scholar]
- Russell, S.M.; Lechner, M.G.; Gong, L.; Megiel, C.; Liebertz, D.J.; Masood, R.; Correa, A.J.; Han, J.; Puri, R.K.; Sinha, U.K.; et al. USC-HN2, a new model cell line for recurrent oral cavity squamous cell carcinoma with immunosuppressive characteristics. Oral Oncol. 2011, 47, 810–817. [Google Scholar] [CrossRef]
- Wild, C.A.; Brandau, S.; Lindemann, M.; Lotfi, R.; Hoffmann, T.K.; Lang, S.; Bergmann, C. Toll-like receptors in regulatory T cells of patients with head and neck cancer. Arch. Otolaryngol. 2010, 136, 1253–1259. [Google Scholar] [CrossRef]
- Bergmann, C.; Strauss, L.; Wang, Y.; Szczepanski, M.J.; Lang, S.; Johnson, J.T.; Whiteside, T.L. T regulatory type 1 cells in squamous cell carcinoma of the head and neck: Mechanisms of suppression and expansion in advanced disease. Clin. Cancer Res. 2008, 14, 3706–3715. [Google Scholar] [CrossRef]
- Strauss, L.; Bergmann, C.; Gooding, W.; Johnson, J.T.; Whiteside, T.L. The frequency and suppressor function of CD4+CD25highFoxp3+ T cells in the circulation of patients with squamous cell carcinoma of the head and neck. Clin. Cancer Res. 2007, 13, 6301–6311. [Google Scholar] [CrossRef]
- Tsujikawa, T.; Yaguchi, T.; Ohmura, G.; Ohta, S.; Kobayashi, A.; Kawamura, N.; Fujita, T.; Nakano, H.; Shimada, T.; Takahashi, T.; et al. Autocrine and paracrine loops between cancer cells and macrophages promote lymph node metastasis via CCR4/CCL22 in head and neck squamous cell carcinoma. Int. J. Cancer 2013, 132, 2755–2766. [Google Scholar] [CrossRef]
- Kulbersh, J.S.; Day, T.A.; Gillespie, M.B.; Young, M.R. 1α,25-Dihydroxyvitamin D3 to skew intratumoral levels of immune inhibitory CD34+ progenitor cells into dendritic cells. Otolaryngol. Head Neck Surg. 2009, 140, 235–240. [Google Scholar] [CrossRef]
- Gasparoto, T.H.; de Souza, M.; Benevides, L.; de Melo, E.J.; Costa, M.R.; Damante, J.H.; Ikoma, M.R.; Garlet, G.P.; Cavassani, K.A.; da Silva, J.S.; et al. Patients with oral squamous cell carcinoma are characterized by increased frequency of suppressive regulatory T cells in the blood and tumor microenvironment. Cancer Immunol. Immunother. 2010, 59, 819–828. [Google Scholar] [CrossRef]
- Lathers, D.M.R.; Achille, N.; Kolesiak, K.; Hulett, K.; Sparano, A.; Young, M.R.I. Increased levels of immune inhibitory CD34+ progenitor cells in the peripheral blood of patients with node positive head and neck cancer and the ability of the CD34+ cells to differentiate into antigen presenting dendritic cells. Clin. Cancer Res. 2001, 125, 205–212. [Google Scholar]
- Pandit, R.; Lathers, D.M.; Beal, N.M.; Garrity, T.; Young, M.R.I. CD34+ immune suppressive cells in the peripheral blood of patients with head and neck cancer. Ann. Otol. Rhinol. Laryngol. 2000, 109, 749–754. [Google Scholar]
- Young, M.R.I.; Petruzzelli, G.; Kolesiak, K.; Achille, N.; Lathers, D.M.; Gabrilovich, D. Human squamous cell carcinomas of the head and neck chemoattract immune suppressive CD34+ progenitor cells. Human Immunol. 2001, 62, 332–341. [Google Scholar] [CrossRef]
- Schuler, P.J.; Borger, V.; Bolke, E.; Habermehl, D.; Matuschek, C.; Wild, C.A.; Greve, J.; Bas, M.; Schilling, B.; Bergmann, C.; et al. Dendritic cell generation and CD4+ CD25high FOXP3+ regulatory T cells in human head and neck carcinoma during radio-chemotherapy. Eur. J. Med. Res. 2011, 16, 57–62. [Google Scholar] [CrossRef]
- Buttigliero, C.; Monagheddu, C.; Petroni, P.; Saini, A.; Dogliotti, L.; Ciccone, G.; Berruti, A. Prognostic role of vitamin d status and efficacy of vitamin D supplementation in cancer patients: A systematic review. Oncologist 2011, 16, 1215–1227. [Google Scholar] [CrossRef]
- Meier, J.D.; Enepekides, D.J.; Poirier, B.; Bradley, C.A.; Albala, J.S.; Farwell, D.G. Treatment with 1-α,25-dihydroxyvitamin D3 (vitamin D3) to inhibit carcinogenesis in the hamster buccal pouch model. Arch. Otolaryngol. 2007, 133, 1149–1152. [Google Scholar] [CrossRef]
- Wiers, K.M.; Lathers, D.M.; Wright, M.A.; Young, M.R.I. Vitamin D3 treatment to diminish the levels of immune suppressive CD34+ cells increases the effectiveness of adoptive immunotherapy. J. Immunother. 2000, 23, 115–124. [Google Scholar] [CrossRef]
- Bolland, M.J.; Grey, A.; Gamble, G.D.; Reid, I.R. Calcium and vitamin D supplements and health outcomes: A reanalysis of the Women’s Health Initiative (WHI) limited-access data set. Am. J. Clin. Nutr. 2011, 94, 1144–1149. [Google Scholar] [CrossRef]
- Ma, Y.; Zhang, P.; Wang, F.; Yang, J.; Liu, Z.; Qin, H. Association between vitamin D and risk of colorectal cancer: A systematic review of prospective studies. J. Clin. Oncol. 2011, 29, 3775–3782. [Google Scholar] [CrossRef]
- Vrieling, A.; Hein, R.; Abbas, S.; Schneeweiss, A.; Flesch-Janys, D.; Chang-Claude, J. Serum 25-hydroxyvitamin D and postmenopausal breast cancer survival: A prospective patient cohort study. Breast Cancer Res. 2011, 13, R74. [Google Scholar] [CrossRef]
- Chiang, K.C.; Yeh, C.N.; Chen, H.Y.; Lee, J.M.; Juang, H.H.; Chen, M.F.; Takano, M.; Kittaka, A.; Chen, T.C. 19-Nor-2α-(3-hydroxypropyl)-1α,25-dihydroxyvitamin D3 (MART-10) is a potent cell growth regulator with enhanced chemotherapeutic potency in liver cancer cells. Steroids 2011, 76, 1513–1519. [Google Scholar] [CrossRef]
- Kovalenko, P.L.; Zhang, Z.; Yu, J.G.; Li, Y.; Clinton, S.K.; Fleet, J.C. Dietary vitamin D and vitamin D receptor level modulate epithelial cell proliferation and apoptosis in the prostate. Cancer Prev. Res. 2011, 4, 1617–1725. [Google Scholar] [CrossRef]
- Lee, L.R.; Teng, P.N.; Nguyen, H.; Hood, B.L.; Kavandi, L.; Wang, G.; Turbov, J.M.; Thaete, L.G.; Hamilton, C.A.; Maxwell, G.L.; et al. Progesterone enhances calcitriol antitumor activity by upregulating vitamin D receptor expression and promoting apoptosis in endometrial cancer cells. Cancer Prev. Res. 2013, 6, 731–743. [Google Scholar] [CrossRef]
- Cantorna, M.T.; Yu, S.; Bruce, D. The paradoxical effects of vitamin D on type 1 mediated immunity. Mol. Aspects Med. 2008, 29, 369–375. [Google Scholar] [CrossRef]
- Chun, R.F.; Adams, J.S.; Hewison, M. Back to the future: A new look at “old” vitamin D. J. Endocrinol. 2008, 198, 261–269. [Google Scholar] [CrossRef]
- Anand, S.P.; Selvaraj, P.; Narayanan, P.R. Effect of 1,25 dihydroxyvitamin D3 on intracellular IFN-γ and TNF-α positive T cell subsets in pulmonary tuberculosis. Cytokine 2009, 45, 105–110. [Google Scholar] [CrossRef]
- Imazeki, I.; Matsuzaki, J.; Tsuji, K.; Nishimura, T. Immunomodulating effect of vitamin D3 derivatives on type 1 cellular immunity. Biomed. Res. 2006, 27, 1–9. [Google Scholar] [CrossRef]
- Correale, J.; Ysrraelit, M.C.; Gaitan, M.I. Vitamin D-mediated immune regulation in multiple sclerosis. J. Neurol. Sci. 2011, 311, 23–31. [Google Scholar] [CrossRef]
- Joshi, S.; Pantalena, L.C.; Liu, X.K.; Gaffen, S.L.; Liu, H.; Rohowsky-Kochan, C.; Ichiyama, K.; Yoshimura, A.; Steinman, L.; Christakos, S.; et al. 1,25-dihydroxyvitamin D3 ameliorates Th17 autoimmunity via transcriptional modulation of interleukin-17A. Mol. Cell. Biol. 2011, 31, 3653–3669. [Google Scholar]
- Ritterhouse, L.L.; Crowe, S.R.; Niewold, T.B.; Kamen, D.L.; Macwana, S.R.; Roberts, V.C.; Dedeke, A.B.; Harley, J.B.; Scofield, R.H.; Guthridge, J.M.; et al. Vitamin D deficiency is associated with an increased autoimmune response in healthy individuals and in patients with systemic lupus erythematosus. Ann. Rheum. Dis. 2011, 70, 1569–1574. [Google Scholar] [CrossRef]
- Yazici, G.; Yildiz, F.; Iskit, A.; Erdemli, E.; Surucu, S.; Firat, P.; Hayran, M.; Ozyigit, G.; Cengiz, M. The effect of vitamin D prophylaxis on radiation induced pulmonary damage. J. Radiat. Res. 2011, 52, 616–621. [Google Scholar] [CrossRef]
- Almand, B.; Clark, J.I.; Nikitina, E.; van Beynen, J.; English, N.R.; Knight, S.C.; Carbone, D.P.; Gabrilovich, D.I. Increased production of immature myeloid cells in cancer patients: A mechanism of immunosuppression in cancer. J. Immunol. 2001, 166, 678–689. [Google Scholar]
- Cheng, X.; Zhao, X.; Khurana, S.; Bruggeman, L.A.; Kao, H.Y. Microarray analyses of glucocorticoid and vitamin D3 target genes in differentiating cultured human podocytes. PLoS One 2013, 8, e60213. [Google Scholar]
- Li, J.; Jin, D.; Fu, S.; Mei, G.; Zhou, J.; Lei, L.; Yu, B.; Wang, G. Insulin-like growth factor binding protein-3 modulates osteoblast differentiation via interaction with Vitamin D receptor. Biochem. Biophys. Res. Commun. 2013, 436, 632–637. [Google Scholar] [CrossRef]
- Garrity, T.; Pandit, R.; Wright, M.A.; Benefield, J.; Young, M.R.I. Increased presence of CD34+ cells in the peripheral blood of head and neck cancer patients and their differentiation into dendritic cells. Int. J. Cancer 1997, 73, 663–669. [Google Scholar] [CrossRef]
- Lathers, D.M.; Lubbers, E.; Wright, M.A.; Young, M.R.I. Dendritic cell differentiation pathways of CD34+ cells from the peripheral blood of head and neck cancer patients. J. Leukoc. Biol. 1999, 65, 623–628. [Google Scholar]
- Ginde, A.A.; Liu, M.C.; Camargo, C.A., Jr. Demographic differences and trends of vitamin D insufficiency in the US population, 1988–2004. Arch. Int. Med. 2009, 169, 626–632. [Google Scholar] [CrossRef]
- Godar, D.E.; Pope, S.J.; Grant, W.B.; Holick, M.F. Solar UV doses of adult Americans and vitamin D3 production. Dermatoendocrinol. 2011, 3, 243–250. [Google Scholar] [CrossRef]
- Walsh, J.E.; Clark, A.M.; Day, T.A.; Gillespie, M.B.; Young, M.R. Use of 1α,25-dihydroxyvitamin D3 treatment to stimulate immune infiltration into head and neck squamous cell carcinoma. Human Immunol. 2010, 71, 659–665. [Google Scholar] [CrossRef]
- Moreno, J.; Krishnan, A.V.; Peehl, D.M.; Feldman, D. Mechanisms of vitamin D-mediated growth inhibition in prostate cancer cells: Inhibition of the prostaglandin pathway. Anticancer Res. 2006, 26, 2525–2530. [Google Scholar]
- Thill, M.; Fischer, D.; Hoellen, F.; Kelling, K.; Dittmer, C.; Landt, S.; Salehin, D.; Diedrich, K.; Friedrich, M.; Becker, S. Prostaglandin metabolising enzymes and PGE2 are inversely correlated with vitamin D receptor and 25(OH)2D3 in breast cancer. Anticancer Res. 2010, 30, 1673–1679. [Google Scholar]
- Thill, M.; Fischer, D.; Kelling, K.; Hoellen, F.; Dittmer, C.; Hornemann, A.; Salehin, D.; Diedrich, K.; Friedrich, M.; Becker, S. Expression of vitamin D receptor (VDR), cyclooxygenase-2 (COX-2) and 15-hydroxyprostaglandin dehydrogenase (15-PGDH) in benign and malignant ovarian tissue and 25-hydroxycholecalciferol (25(OH2)D3) and prostaglandin E2 (PGE2) serum level in ovarian cancer patients. J. Steroid Biochem. Mol. Biol. 2010, 121, 387–390. [Google Scholar] [CrossRef]
- Dong, J.; Wong, S.L.; Lau, C.W.; Liu, J.; Wang, Y.X.; He, Z.D.; Ng, C.F.; Chen, Z.Y.; Yao, X.; Xu, A.; et al. Calcitriol restores renovascular function in estrogen-deficient rats through downregulation of cyclooxygenase-2 and the thromboxane-prostanoid receptor. Kidney Int. 2013, 84, 54–63. [Google Scholar] [CrossRef]
- Renkonen, J.; Wolff, H.; Paavonen, T. Expression of cyclo-oxygenase-2 in human tongue carcinoma and its precursor lesions. Virchows Arch. 2002, 440, 594–597. [Google Scholar] [CrossRef]
- Mulligan, J.K.; Day, T.A.; Gillespie, M.B.; Rosenzweig, S.A.; Young, M.R. Secretion of vascular endothelial growth factor by oral squamous cell carcinoma cells skews endothelial cells to suppress T-cell functions. Human Immunol. 2009, 70, 375–382. [Google Scholar] [CrossRef]
- Wanebo, H.J.; Riley, T.; Karz, D.; Pace, R.C.; Johns, M.E.; Cantrell, R.W. Indomethacin sensitive suppressor-cell activity in head and neck cancer patients. The role of the adherent mononuclear cell. Cancer 1988, 61, 462–474. [Google Scholar] [CrossRef]
- Dovedi, S.J.; Kirby, J.A.; Davies, B.R.; Leung, H.; Kelly, J.D. Celecoxib has potent antitumour effects as a single agent and in combination with BCG immunotherapy in a model of urothelial cell carcinoma. Eur. Urol. 2008, 54, 621–630. [Google Scholar] [CrossRef]
- Young, M.R.; Newby, M. Differential induction of macrophage prostaglandin E2 secretion and suppressor activity by Lewis lung carcinoma variants. In Prostaglandins and Cancer; Garaci, E., Santoro, M.G., Eds.; Springer-Verlag: Berlin Heidelberg, Germany, 1987; pp. 273–276. [Google Scholar]
- Gillespie, M.B.; Moody, M.W.; Lee, F.S.; Poole, L.J.; Hornig, J.D.; Lathers, D.; Young, M.R.; Day, T.A. Head and neck cancer recurrence and mortality in nonselective cyclooxygenase inhibitor users. Arch. Otolaryngol. 2007, 133, 28–31. [Google Scholar] [CrossRef]
- Lang, S.; Lauffer, L.; Clausen, C.; Lohr, I.; Schmitt, B.; Holzel, D.; Wollenberg, B.; Gires, O.; Kastenbauer, E.; Zeidler, R. Impaired monocyte function in cancer patients: Restoration with a cyclooxygenase-2 inhibitor. FASEB J. 2003, 17, 286–288. [Google Scholar]
- Lin, R.; Nagai, Y.; Sladek, R.; Bastien, Y.; Ho, J.; Petrecca, K.; Sotiropoulou, G.; Diamandis, E.P.; Hudson, T.J.; White, J.H. Expression profiling in squamous carcinoma cells reveals pleiotropic effects of vitamin D3 analog EB1089 signaling on cell proliferation, differentiation, and immune system regulation. Mol. Endocrinol. 2002, 16, 1243–1256. [Google Scholar] [CrossRef]
- De Costa, A.M.; Schuyler, C.A.; Walker, D.D.; Young, M.R. Characterization of the evolution of immune phenotype during the development and progression of squamous cell carcinoma of the head and neck. Cancer Immunol. Immunother. 2011, 61, 927–939. [Google Scholar]
- Kesselring, R.; Thiel, A.; Pries, R.; Trenkle, T.; Wollenberg, B. Human Th17 cells can be induced through head and neck cancer and have a functional impact on HNSCC development. Br. J. Cancer 2010, 103, 1245–1254. [Google Scholar]
- Gnerlich, J.L.; Mitchem, J.B.; Weir, J.S.; Sankpal, N.V.; Kashiwagi, H.; Belt, B.A.; Porembka, M.R.; Herndon, J.M.; Eberlein, T.J.; Goedegebuure, P.; et al. Induction of Th17 cells in the tumor microenvironment improves survival in a murine model of pancreatic cancer. J. Immunol. 2010, 185, 4063–4071. [Google Scholar] [CrossRef]
- Maruyama, T.; Kono, K.; Mizukami, Y.; Kawaguchi, Y.; Mimura, K.; Watanabe, M.; Izawa, S.; Fujii, H. Distribution of Th17 cells and FoxP3+ regulatory T cells in tumor-infiltrating lymphocytes, tumor-draining lymph nodes and peripheral blood lymphocytes in patients with gastric cancer. Cancer Sci. 2010, 101, 1947–1954. [Google Scholar] [CrossRef]
- Berger, H.; Vegran, F.; Chikh, M.; Gilardi, F.; Ladoire, S.; Bugaut, H.; Mignot, G.; Chalmin, F.; Bruchard, M.; Derangere, V.; et al. SOCS3 transactivation by PPARγ prevents IL-17-driven cancer growth. Cancer Res. 2013, 73, 3578. [Google Scholar] [CrossRef]
- Bessler, H.; Djaldetti, M. 1α,25-Dihydroxyvitamin D3 modulates the interaction between immune and colon cancer cells. Biomed. Pharmacother. 2012, 66, 428–432. [Google Scholar] [CrossRef]
- Olsen, K.S.; Rylander, C.; Brustad, M.; Aksnes, L.; Lund, E. Plasma 25 hydroxyvitamin D level and blood gene expression profiles: A cross-sectional study of the Norwegian Women and Cancer Post-genome Cohort. Eur. J. Clin. Nutr. 2013, 67, 773–778. [Google Scholar] [CrossRef]
- Fleet, J.C.; DeSmet, M.; Johnson, R.; Li, Y. Vitamin D and cancer: A review of molecular mechanisms. Biochem. J. 2012, 441, 61–76. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Young, M.R.I.; Day, T.A. Immune Regulatory Activity of Vitamin D3 in Head and Neck Cancer. Cancers 2013, 5, 1072-1085. https://doi.org/10.3390/cancers5031072
Young MRI, Day TA. Immune Regulatory Activity of Vitamin D3 in Head and Neck Cancer. Cancers. 2013; 5(3):1072-1085. https://doi.org/10.3390/cancers5031072
Chicago/Turabian StyleYoung, M. Rita I., and Terry A. Day. 2013. "Immune Regulatory Activity of Vitamin D3 in Head and Neck Cancer" Cancers 5, no. 3: 1072-1085. https://doi.org/10.3390/cancers5031072
APA StyleYoung, M. R. I., & Day, T. A. (2013). Immune Regulatory Activity of Vitamin D3 in Head and Neck Cancer. Cancers, 5(3), 1072-1085. https://doi.org/10.3390/cancers5031072