The Role of Macrophage Migration Inhibitory Factor (MIF) in Ultraviolet Radiation-Induced Carcinogenesis

Abstract
:
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. UV-Induced Inflammatory Cytokines in the Skin
3. Macrophage Migration Inhibitory Factor (MIF)
4. MIF Inhibits p53-Dependent Apoptotic Processes Following UV Radiation
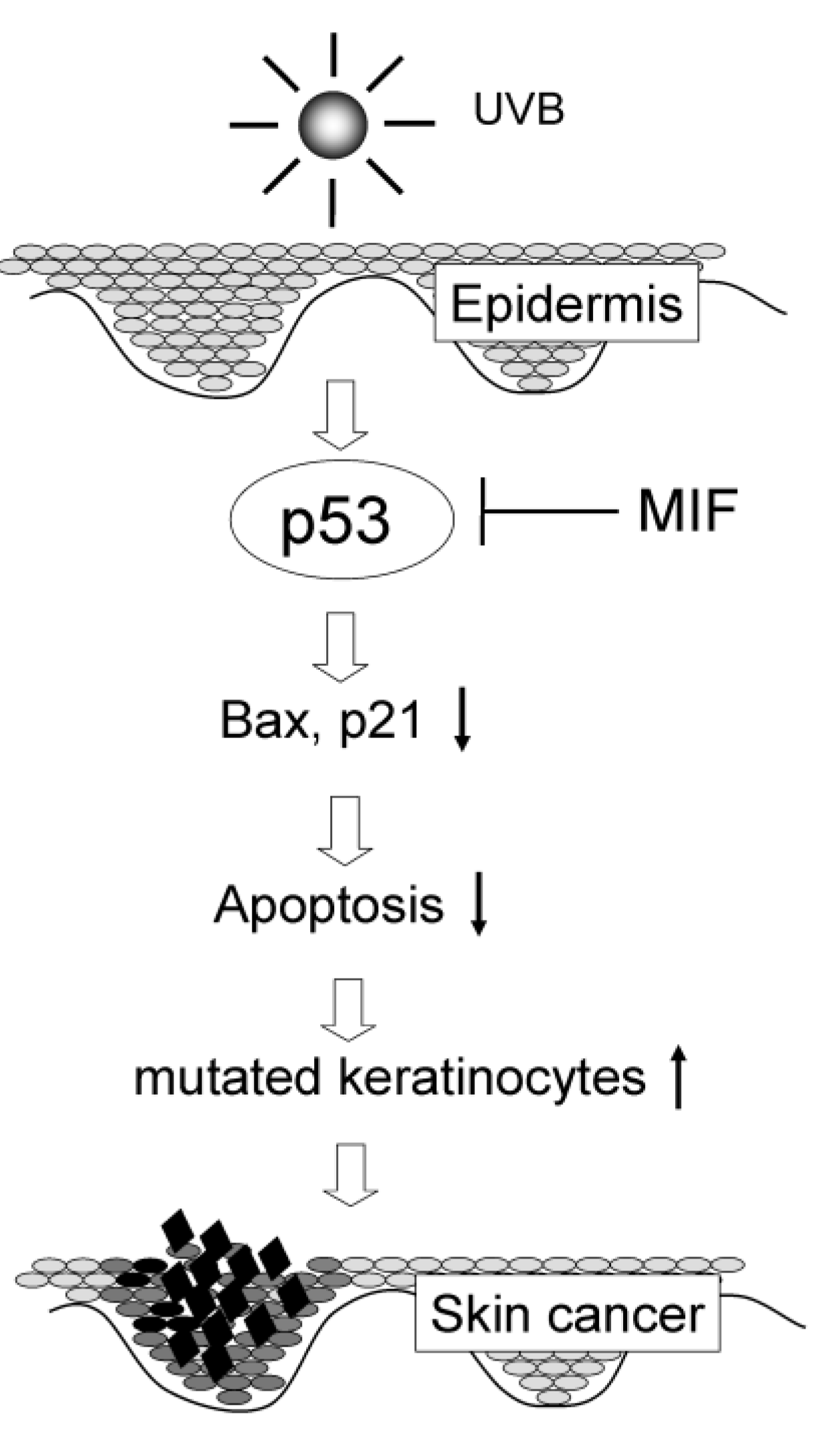
5. MIF Exerts Pro-neoplastic Activity after UV Radiation
6. MIF-Induced Skin Inflammation and Carcinogenesis
7. Conclusions
References
- Matsumura, Y.; Ananthaswamy, H.N. Short-term and long-term cellular and molecular events following UV irradiation of skin: implications for molecular medicine. Expert Rev. Mol. Med. 2002, 4, 1–22. [Google Scholar]
- Tenkate, T.D. Occupational exposure to ultraviolet radiation: a health risk assessment. Rev. Environ. Health 1999, 14, 187–209. [Google Scholar] [CrossRef]
- Ma, W.; Wlaschek, M.; Tantcheva-Poór, I.; Schneider, L.A.; Naderi, L.; Razi-Wolf, Z.; Schüller, J.; Scharffetter-Kochanek, K. Chronological ageing and photoageing of the fibroblasts and the dermal connective tissue. Clin. Exp. Dermatol. 2001, 26, 592–599. [Google Scholar] [CrossRef]
- Young, A.R. Cumulative effects of ultraviolet radiation on the skin: cancer and photoaging. Semin. Dermatol. 1990, 9, 25–31. [Google Scholar]
- Ridley, A.J.; Whiteside, J.R.; McMillan, T.J.; Allinson, S.L. Cellular and sub-cellular responses to UVA in relation to carcinogenesis. Int. J. Radiat. Biol. 2009, 85, 177–195. [Google Scholar] [CrossRef]
- Kraemer, K.H. Sunlight and skin cancer: another link revealed. Proc. Natl. Acad. Sci. USA 1997, 94, 11–14. [Google Scholar] [CrossRef]
- Fischer, S.M.; Lo, H.H.; Gordon, G.B.; Seibert, K.; Kelloff, G.; Lubet, R.A.; Conti, C.J. Chemopreventive activity of celecoxib, a specific cyclooxygenase-2 inhibitor, and indomethacin against ultraviolet light-induced skin carcinogenesis. Mol. Carcinog. 1999, 25, 231–240. [Google Scholar] [CrossRef]
- Pentland, A.P.; Schoggins, J.W.; Scott, G.A.; Khan, K.N.; Han, R. Reduction of UV-induced skin tumors in hairless mice by selective COX-2 inhibition. Carcinogenesis 1999, 20, 1939–1944. [Google Scholar] [CrossRef]
- Cleaver, J.E.; Crowley, E. UV damage, DNA repair and skin carcinogenesis. Front. Biosci. 2002, 7, d1024–d1043. [Google Scholar] [CrossRef]
- Ansel, J.C.; Luger, T.A.; Lowry, D.; Perry, P.; Roop, D.R.; Mountz, J.D. The expression and modulation of IL-1 alpha in murine keratinocytes. J. Immunol. 1988, 140, 2274–2278. [Google Scholar]
- Kupper, T.S.; Chua, A.O.; Flood, P.; McGuire, J.; Gubler, U. Interleukin 1 gene expression in cultured human keratinocytes is augmented by ultraviolet irradiation. J. Clin. Invest. 1987, 80, 430–436. [Google Scholar] [CrossRef]
- Kirnbauer, R.; Kock, A.; Neuner, P.; Forster, E.; Krutmann, J.; Urbanski, A.; Schauer, E.; Ansel, J.C.; Schwarz, T.; Luger, T.A. Regulation of epidermal cell interleukin-6 production by UV light and corticosteroids. J. Invest. Dermatol. 1991, 96, 484–489. [Google Scholar]
- Köck, A.; Schwarz, T.; Kirnbauer, R.; Urbanski, A.; Perry, P.; Ansel, J.C.; Luger, T.A. Human keratinocytes are a source for tumor necrosis factor alpha: evidence for synthesis and release upon stimulation with endotoxin or ultraviolet light. J. Exp. Med. 1990, 172, 1609–1614. [Google Scholar] [CrossRef]
- Dinarello, C.A.; Cannon, J.G.; Wolff, S.M.; Bernheim, H.A.; Beutler, B.; Cerami, A.; Figari, I.S.; Palladino, M.A., Jr.; O'Connor, J.V. Tumor necrosis factor (cachectin) is an endogenous pyrogen and induces production of interleukin 1. J. Exp. Med. 1986, 163, 1433–1450. [Google Scholar] [CrossRef]
- Schwarz, A.; Bhardwaj, R; Aragane, Y.; Mahnke, K.; Riemann, H.; Metze, D.; Luger, T.A.; Schwarz, T. Ultraviolet-B-induced apoptosis of keratinocytes: evidence for partial involvement of tumor necrosis factor-alpha in the formation of sunburn cells. J. Invest. Dermatol. 1995, 104, 922–997. [Google Scholar]
- Suganuma, M.; Okabe, S.; Kurusu, M.; Iida, N.; Ohshima, S.; Saeki, Y.; Kishimoto, T.; Fujiki, H. Discrete roles of cytokines, TNF-alpha, IL-1, IL-6 in tumor promotion and cell transformation. Int. J. Oncol. 2002, 20, 131–136. [Google Scholar]
- Bloom, B.R.; Bennett, B. Mechanism of a reaction in vitro associated with delayed-type hypersensitivity. Science 1966, 153, 80–82. [Google Scholar]
- David, J.R. Delayed hypersensitivity in vitro: its mediation by cell-free substances formed by lymphoid cell-antigen interaction. Proc. Natl. Acad. Sci. USA 1966, 56, 72–77. [Google Scholar] [CrossRef]
- Weiser, W.Y.; Temple, P.A.; Witek-Giannotti, J.S.; Remold, H.G.; Clark, S.C.; David, J.R. Molecular cloning of a cDNA encoding a human macrophage migration inhibitory factor. Proc. Natl. Acad. Sci. USA 1989, 86, 7522–7526. [Google Scholar] [CrossRef]
- Bernhagen, J.; Calandra, T.; Mitchell, R.A.; Martin, S.B.; Tracey, K.J.; Voelter, W.; Manogue, K.R.; Cerami, A.; Bucala, R. MIF is a pituitary-derived cytokine that potentiates lethal endotoxaemia. Nature 1993, 365, 756–759. [Google Scholar] [CrossRef]
- Calandra, T.; Bernhagen, J.; Mitchell, R.A.; Bucala, R. The macrophage is an important and previously unrecognized source of macrophage migration inhibitory factor. J. Exp. Med. 1994, 179, 1895–1902. [Google Scholar] [CrossRef]
- Lanahan, A.; Williams, J.B.; Sanders, L.K.; Nathans, D. Growth factor-induced delayed early response genes. Mol. Cell. Biol. 1992, 12, 3919–3929. [Google Scholar]
- Wistow, G.J.; Shaughnessy, M.P.; Lee, D.C.; Hodin, J.; Zelenka, P.S. A macrophage migration inhibitory factor is expressed in the differentiating cells of the eye lens. Proc. Natl. Acad. Sci. USA 1993, 90, 1272–1275. [Google Scholar] [CrossRef]
- Bucala, R. MIF re-discovered: pituitary hormone and glucocorticoid-induced regulator of cytokine production. Cytokine Growth Factor Rev. 1996, 7, 19–24. [Google Scholar] [CrossRef]
- Bacher, M.; Meinhardt, A.; Lan, H.Y.; Mu, W.; Metz, C.N.; Chesney, J.A.; Calandra, T.; Gemsa, D.; Donnelly, T.; Atkins, R.C.; Bucala, R. Migration inhibitory factor expression in experimentally induced endotoxemia. Am. J. Pathol. 1997, 150, 235–246. [Google Scholar]
- Nishihira, J. Macrophage migration inhibitory factor (MIF): its essential role in the immune system and cell growth. J. Interferon Cytokine Res. 2000, 20, 751–762. [Google Scholar] [CrossRef]
- Onodera, S.; Kaneda, K.; Mizue, Y.; Koyama, Y.; Fujinaga, M.; Nishihira, J. Macrophage migration inhibitory factor up-regulates expression of matrix metalloproteinases in synovial fibroblasts of rheumatoid arthritis. J. Biol. Chem. 2000, 275, 444–450. [Google Scholar]
- Calandra, T.; Bernhagen, J.; Metz, C.N.; Spiegel, L.A.; Bacher, M.; Donnelly, T.; Cerami, A.; Bucala, R. MIF as a glucocorticoid-induced modulator of cytokine production. Nature 1995, 377, 68–71. [Google Scholar] [CrossRef]
- Rosengren, E.; Bucala, R.; Aman, P.; Jacobsson, L.; Odh, G.; Metz, C.N.; Rorsman, H. The immunoregulatory mediator macrophage migration inhibitory factor (MIF) catalyzes a tautomerization reaction. Mol. Med. 1996, 2, 143–149. [Google Scholar]
- Roger, T.; David, J.; Glauser, M.P.; Calandra, T. MIF regulates innate immune responses through modulation of Toll-like receptor 4. Nature 2001, 414, 920–924. [Google Scholar] [CrossRef]
- Welford, S.M.; Bedogni, B.; Gradin, K.; Poellinger, L.; Broome Powell, M.; Giaccia, A.J. HIF1alpha delays premature senescence through the activation of MIF. Genes Dev. 2006, 20, 3366–3371. [Google Scholar] [CrossRef]
- Shi, X.; Leng, L.; Wang, T.; Wang, W.; Du, X.; Li, J.; McDonald, C.; Chen, Z.; Murphy, J.W.; Lolis, E.; Noble, P.; Knudson, W.; Bucala, R. CD44 is the signaling component of the macrophage migration inhibitory factor-CD74 receptor complex. Immunity 2006, 25, 595–606. [Google Scholar] [CrossRef]
- Leng, L.; Metz, C.N.; Fang, Y.; Xu, J.; Donnelly, S.; Baugh, J.; Delohery, T.; Chen, Y.; Mitchell, R.A.; Bucala, R. MIF signal transduction initiated by binding to CD74. J. Exp. Med. 2003, 197, 1467–1476. [Google Scholar] [CrossRef]
- Schwartz, V.; Lue, H.; Kraemer, S; Korbiel, J.; Krohn, R.; Ohl, K.; Bucala, R.; Weber, C.; Bernhagen, J. A functional heteromeric MIF receptor formed by CD74 and CXCR4. FEBS Lett. 2009, 583, 2749–2757. [Google Scholar] [CrossRef]
- Shimizu, T.; Ohkawara, A.; Nishihira, J.; Sakamoto, W. Identification of macrophage migration inhibitory factor (MIF) in human skin and its immunohistochemical localization. FEBS Lett. 1996, 381, 199–202. [Google Scholar] [CrossRef]
- Watanabe, H.; Shimizu, T.; Nishihira, J.; Abe, R.; Nakayama, T.; Taniguchi, M.; Sabe, H.; Ishibashi, T.; Shimizu, H. Ultraviolet A-induced production of matrix metalloproteinase-1 is mediated by macrophage migration inhibitory factor (MIF) in human dermal fibroblasts. J. Biol. Chem. 2004, 279, 1676–1683. [Google Scholar] [CrossRef]
- Shimizu, T.; Abe, R.; Ohkawara, A.; Nishihira, J. Ultraviolet B radiation upregulates the production of macrophage migration inhibitory factor (MIF) in human epidermal keratinocytes. J. Invest. Dermatol. 1999, 112, 210–215. [Google Scholar] [CrossRef]
- Zhao, Y.; Shimizu, T.; Nishihira, J.; Koyama, Y.; Kushibiki, T.; Honda, A.; Watanabe, H.; Abe, R.; Tabata, Y.; Shimizu, H. Tissue regeneration using macrophage migration inhibitory factor (MIF)-impregnated gelatin microbeads in cutaneous wounds. Am. J. Pathol. 2005, 167, 1519–1529. [Google Scholar] [CrossRef]
- Shimizu, T.; Abe, R.; Ohkawara, A.; Nishihira, J. Increased production of macrophage migration inhibitory factor by PBMCs of atopic dermatitis. J. Allergy Clin. Immunol. 1999, 104, 659–664. [Google Scholar] [CrossRef]
- Hamasaka, A.; Abe, R.; Koyama, Y.; Yoshioka, N.; Fujita, Y.; Hoshina, D.; Sasaki, M.; Hirasawa, T.; Onodera, S.; Ohshima, S.; Leng, L.; Bucala, R.; Nishihira, J.; Shimizu, T.; Shimizu, H. DNA-vaccination against macrophage migration inhibitory factor improves atopic dermatitis in murine models. J. Allergy Clin. Immunol. 2009, 124, 90–99. [Google Scholar] [CrossRef]
- Shimizu, T.; Abe, R.; Nakamura, H.; Ohkawara, A.; Suzuki, M.; Nishihira, J. High expression of macrophage migration inhibitory factor in human melanoma cells and its role in tumor cell growth and angiogenesis. Biochem. Biophys. Res. Commun. 1999, 264, 751–758. [Google Scholar] [CrossRef]
- Chesney, J.; Metz, C.; Bacher, M.; Peng, T.; Meinhardt, A.; Bucala, R. An essential role for macrophage migration inhibitory factor (MIF) in angiogenesis and the growth of a murine lymphoma. Mol. Med. 1999, 5, 181–191. [Google Scholar]
- Mitchell, R.A.; Liao, H.; Chesney, J.; Fingerle-Rowson, G.; Baugh, J.; David, J.; Bucala, R. Macrophage migration inhibitory factor (MIF) sustains macrophage proinflammatory function by inhibiting p53: regulatory role in the innate immune response. Proc. Natl. Acad. Sci. USA 2002, 99, 345–350. [Google Scholar]
- Hudson, J.D.; Shoaibi, M.A.; Maestro, R.; Carnero, A.; Hannon, G.J.; Beach, D.H. A proinflammatory cytokine inhibits p53 tumor suppressor activity. J. Exp. Med. 1999, 190, 1375–1382. [Google Scholar]
- Nemajerova, A.; Mena, P.; Fingerle-Rowson, G.; Moll, U.M.; Petrenko, O. Impaired DNA damage checkpoint response in MIF-deficient mice. EMBO J. 2007, 26, 987–997. [Google Scholar] [CrossRef]
- Levine, A.J. p53, the cellular gatekeeper for growth and division. Cell 1997, 88, 323–331. [Google Scholar] [CrossRef]
- Stenback, F.; Makinen, M.; Jussila, T. p53 expression in skin carcinogenesis and its relationship to cell proliferation and tumour growth. Eur. J. Cancer 1998, 34, 1415–1424. [Google Scholar] [CrossRef]
- Fingerle-Rowson, G.; Petrenko, O.; Metz, C.N.; Forsthuber, T.G.; Mitchell, R.; Huss, R.; Moll, U.; Muller, W.; Bucala, R. The p53-dependent effects of macrophage migration inhibitory factor revealed by gene targeting. Proc. Natl. Acad. Sci. USA 2003, 100, 9354–9359. [Google Scholar] [CrossRef]
- Shimizu, T. Role of macrophage migration inhibitory factor (MIF) in the skin. J. Dermatol. Sci. 2005, 37, 65–73. [Google Scholar] [CrossRef]
- Honda, A.; Abe, R.; Yoshihisa, Y.; Makino, T.; Matsunaga, K.; Nishihira, J.; Shimizu, H.; Shimizu, T. Deficient deletion of apoptotic cells by macrophage migration inhibitory factor (MIF) overexpression accelerates photocarcinogenesis. Carcinogenesis 2009, 30, 1597–1605. [Google Scholar] [CrossRef]
- Kitaichi, N.; Shimizu, T.; Yoshida, K.; Honda, A.; Yoshihisa, Y.; Kase, S.; Ohgami, K.; Norisugi, O.; Makino, T.; Nishihira, J.; Yamagishi, S.; Ohno, S. Macrophage migration inhibitory factor ameliorates UV-induced photokeratitis in mice. Exp. Eye Res. 2008, 86, 929–935. [Google Scholar] [CrossRef]
- Martin, J.; Duncan, F.J.; Keiser, T.; Shin, S.; Kusewitt, D.F.; Oberyszyn, T.; Satoskar, A.R.; VanBuskirk, A.M. Macrophage migration inhibitory factor (MIF) plays a critical role in pathogenesis of ultraviolet-B (UVB) -induced nonmelanoma skin cancer (NMSC). FASEB J. 2009, 23, 720–730. [Google Scholar] [CrossRef]
- Meyer-Siegler, K.; Fattor, R.A.; Hudson, P.B. Expression of macrophage migration inhibitory factor in the human prostate. Diagn. Mol. Pathol. 1998, 7, 44–50. [Google Scholar] [CrossRef]
- Legendre, H.; Decaestecker, C.; Nagy, N.; Hendlisz, A.; Schuring, M.P.; Salmon, I.; Gabius, H.J.; Pector, J.C.; Kiss, R. Prognostic values of galectin-3 and the macrophage migration inhibitory factor (MIF) in human colorectal cancers. Mod. Pathol. 2003, 16, 491–504. [Google Scholar] [CrossRef]
- Ren, Y.; Tsui, H.T.; Poon, R.T.; Ng, I.O.; Li, Z.; Chen, Y.; Jiang, G.; Lau, C.; Yu, W.C.; Bacher, M.; Fan, S.T. Macrophage migration inhibitory factor: roles in regulating tumor cell migration and expression of angiogenic factors in hepatocellular carcinoma. Int. J. Cancer 2003, 107, 22–29. [Google Scholar] [CrossRef]
- Kamimura, A.; Kamachi, M.; Nishihira, J.; Ogura, S.; Isobe, H.; Dosaka-Akita, H.; Ogata, A.; Shindoh, M.; Ohbuchi, T.; Kawakami, Y. Intracellular distribution of macrophage migration inhibitory factor predicts the prognosis of patients with adenocarcinoma of the lung. Cancer 2000, 89, 334–341. [Google Scholar] [CrossRef]
- Rogge, L. A genomic view of helper T cell subsets. Ann. N. Y. Acad. Sci. 2002, 975, 57–67. [Google Scholar] [CrossRef]
- Takahashi, N.; Nishihira, J.; Sato, Y.; Kondo, M.; Ogawa, H.; Ohshima, T.; Une, Y.; Todo, S. Involvement of macrophage migration inhibitory factor (MIF) in the mechanism of tumor cell growth. Mol. Med. 1998, 4, 707–714. [Google Scholar]
- Meyer-Siegler, K.L.; Iczkowski, K.A.; Leng, L.; Bucala, R.; Vera, P.L. Inhibition of macrophage migration inhibitory factor or its receptor (CD74) attenuates growth and invasion of DU-145 prostate cancer cells. J. Immunol. 2006, 177, 8730–8739. [Google Scholar]
- Abe, R.; Peng, T.; Sailors, J.; Bucala, R.; Metz, C.N. Regulation of the CTL response by macrophage migration inhibitory factor. J. Immunol. 2001, 166, 747–753. [Google Scholar]
- Hagemann, T.; Robinson, S.C.; Thompson, R.G.; Charles, K.; Kulbe, H.; Balkwill, F.R. Ovarian cancer cell-derived migration inhibitory factor enhances tumor growth, progression, and angiogenesis. Mol. Cancer Ther. 2007, 6, 1993–2002. [Google Scholar] [CrossRef]
- Moore, R.J.; Owens, D.M.; Stamp, G.; Arnott, C.; Burke, F.; East, N.; Holdsworth, H.; Turner, L.; Rollins, B.; Pasparakis, M.; Kollias, G.; Balkwill, F. Mice deficient in tumor necrosis factor-alpha are resistant to skin carcinogenesis. Nat. Med. 1999, 5, 828–831. [Google Scholar] [CrossRef]
- Wilgus, T.A.; Koki, A.T.; Zweifel, B.S.; Kusewitt, D.F.; Rubal, P.A.; Oberyszyn, T.M. Inhibition of cutaneous ultraviolet light B-mediated inflammation and tumor formation with topical celecoxib treatment. Mol. Carcinog. 2003, 38, 49–58. [Google Scholar] [CrossRef]
- Bach, J.P.; Rinn, B.; Meyer, B.; Dodel, R.; Bacher, M. Role of MIF in inflammation and tumorigenesis. Oncology 2008, 75, 127–133. [Google Scholar] [CrossRef]
- Toh, M.L.; Aeberli, D.; Lacey, D.; Yang, Y.; Santos, L.L.; Clarkson, M.; Sharma, L.; Clyne, C.; Morand, E.F. Regulation of IL-1 and TNF receptor expression and function by endogenous macrophage migration inhibitory factor. J. Immunol. 2006, 177, 4818–4825. [Google Scholar]
- Sanchez-Zamora, Y.; Terrazas, L.I.; Vilches-Flores, A.; Leal, E.; Juárez, I.; Whitacre, C.; Kithcart, A.; Pruitt, J.; Sielecki, T.; Satoskar, A.R.; Rodriguez-Sosa, M. Macrophage migration inhibitory factor is a therapeutic target in treatment of non-insulin-dependent diabetes mellitus. FASEB J. 2010, 24, 2583–2590. [Google Scholar] [CrossRef]
- Donnelly, S.C.; Haslett, C.; Reid, P.T.; Grant, I.S.; Wallace, W.A.; Metz, C.N.; Bruce, L.J.; Bucala, R. Regulatory role for macrophage migration inhibitory factor in acute respiratory distress syndrome. Nat. Med. 1997, 3, 320–323. [Google Scholar] [CrossRef]
© 2010 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Shimizu, T. The Role of Macrophage Migration Inhibitory Factor (MIF) in Ultraviolet Radiation-Induced Carcinogenesis. Cancers 2010, 2, 1555-1564. https://doi.org/10.3390/cancers2031555
Shimizu T. The Role of Macrophage Migration Inhibitory Factor (MIF) in Ultraviolet Radiation-Induced Carcinogenesis. Cancers. 2010; 2(3):1555-1564. https://doi.org/10.3390/cancers2031555
Chicago/Turabian StyleShimizu, Tadamichi. 2010. "The Role of Macrophage Migration Inhibitory Factor (MIF) in Ultraviolet Radiation-Induced Carcinogenesis" Cancers 2, no. 3: 1555-1564. https://doi.org/10.3390/cancers2031555
APA StyleShimizu, T. (2010). The Role of Macrophage Migration Inhibitory Factor (MIF) in Ultraviolet Radiation-Induced Carcinogenesis. Cancers, 2(3), 1555-1564. https://doi.org/10.3390/cancers2031555