

Identification of Deregulated Proteins in Mutated BRCA1/2 Breast and Ovarian Cancers for Vectorized Biologics

 , , , , and
, , , , and
Simple Summary
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Identification of BRCA1/BRCA2 Mutations in Breast and Ovarian Cancer Patients
2.2. Analysis of Molecular Function of the Selected Genes
2.3. Outcome and Prognosis Analysis
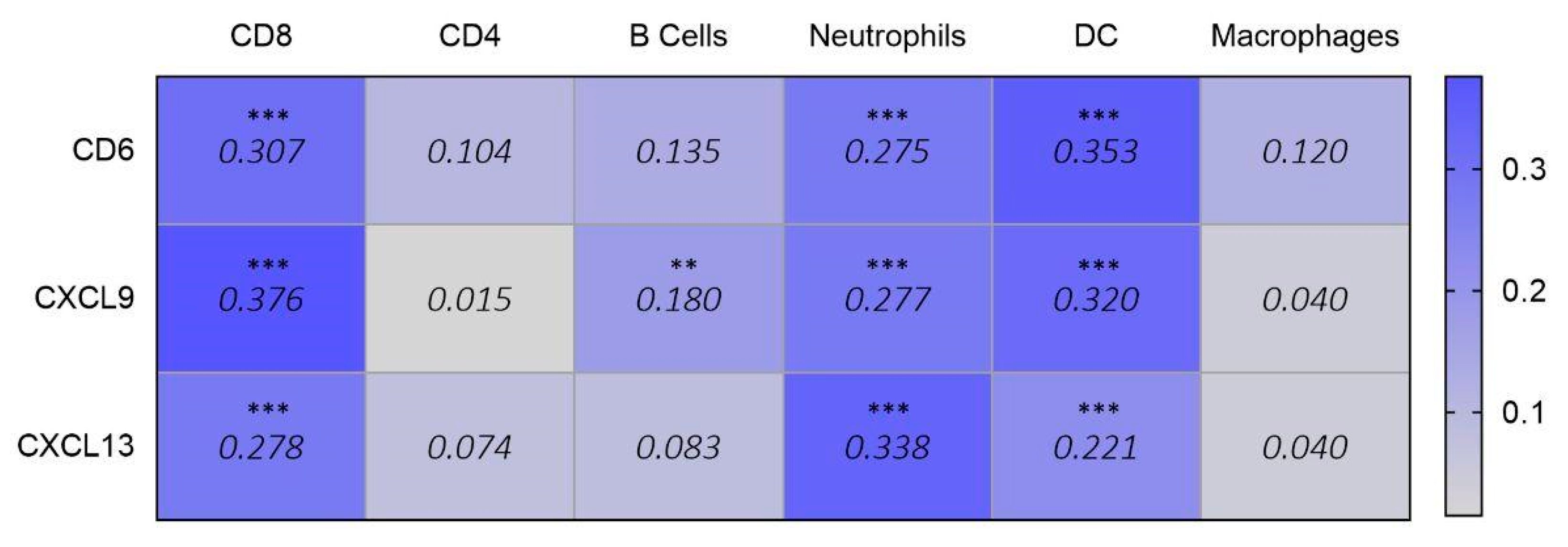
2.4. Immune Cell Infiltration and Gene Expression Correlation
2.5. Statistical Analysis
2.6. Graphical Design
3. Results
3.1. Flow Chart for the Selection and Identification of Genes
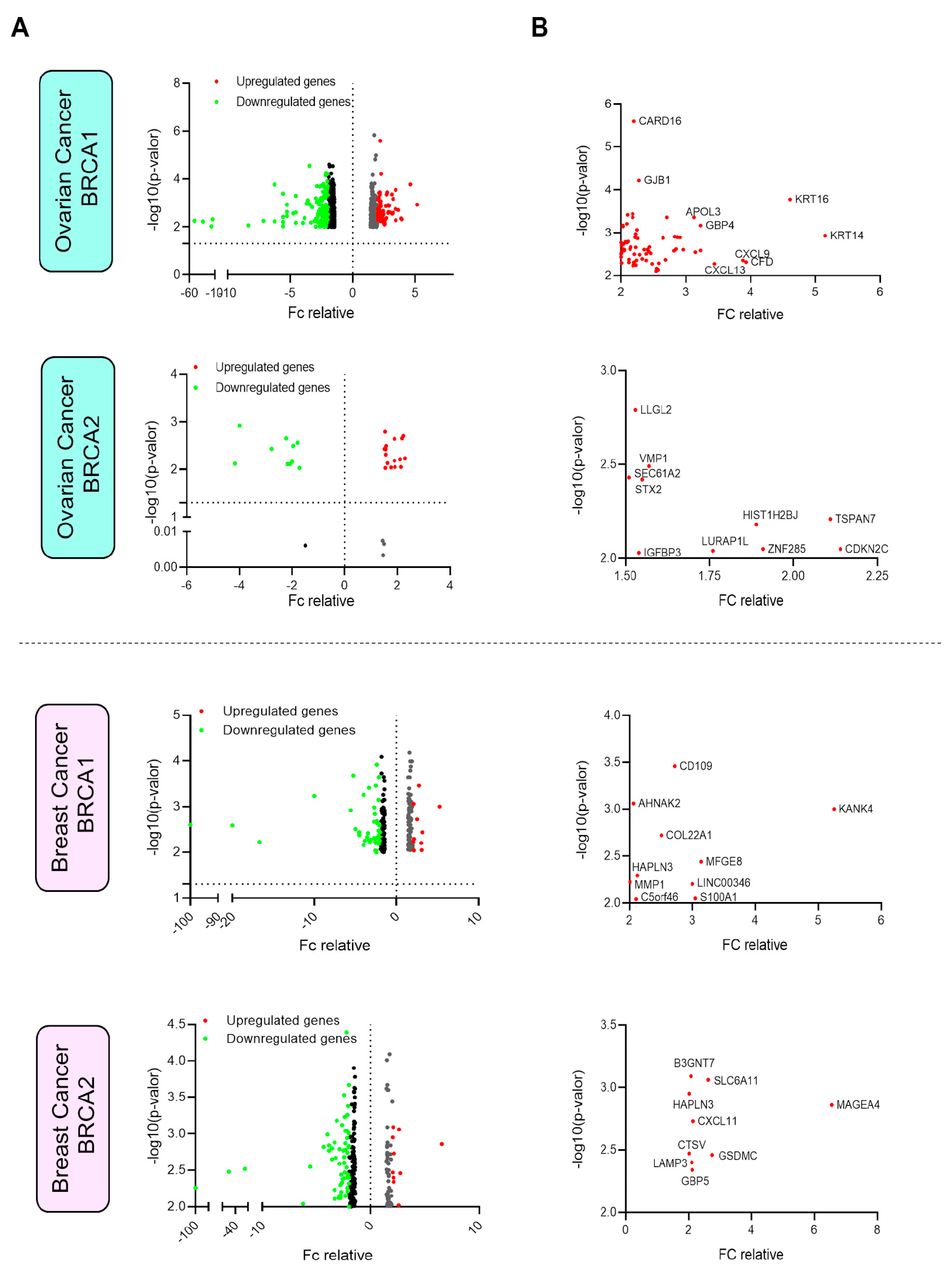
3.2. Selection of the Relevant Upregulated Genes in Breast and Ovarian Cancer
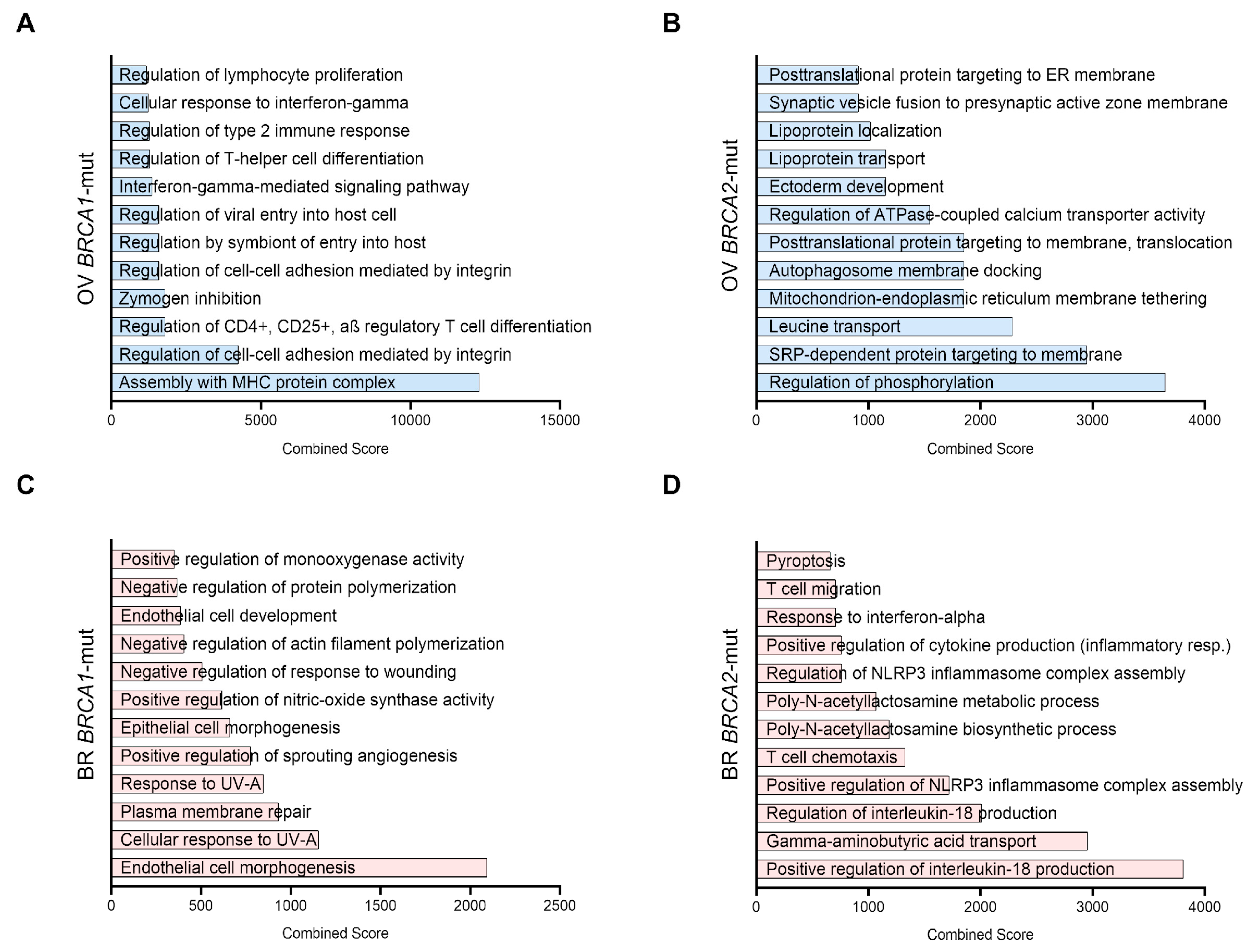
3.3. Gene-Set Enrichment Analysis of Upregulated Genes
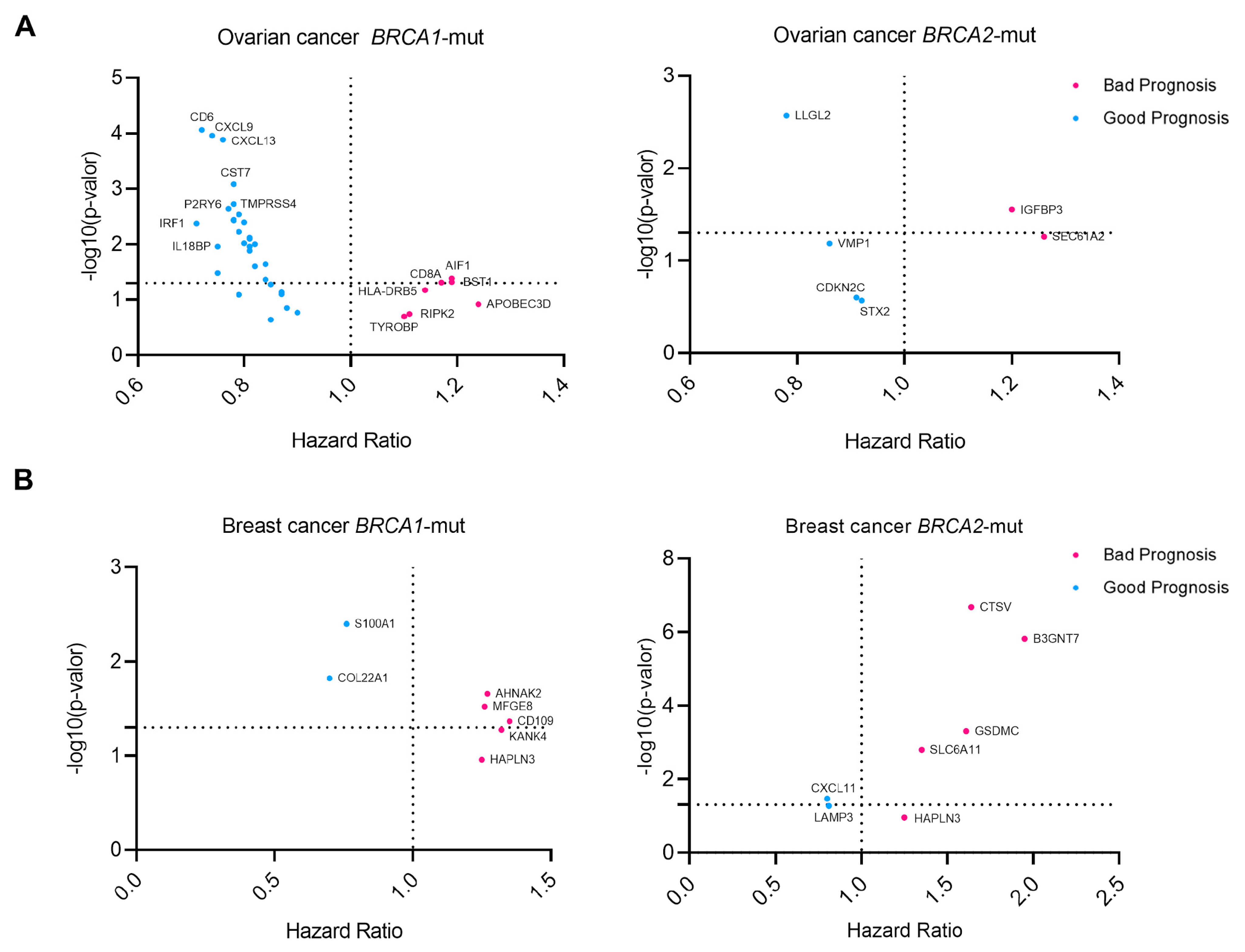
3.4. Outcome Analysis and Differential Expression of Upregulated Surfaceome Genes
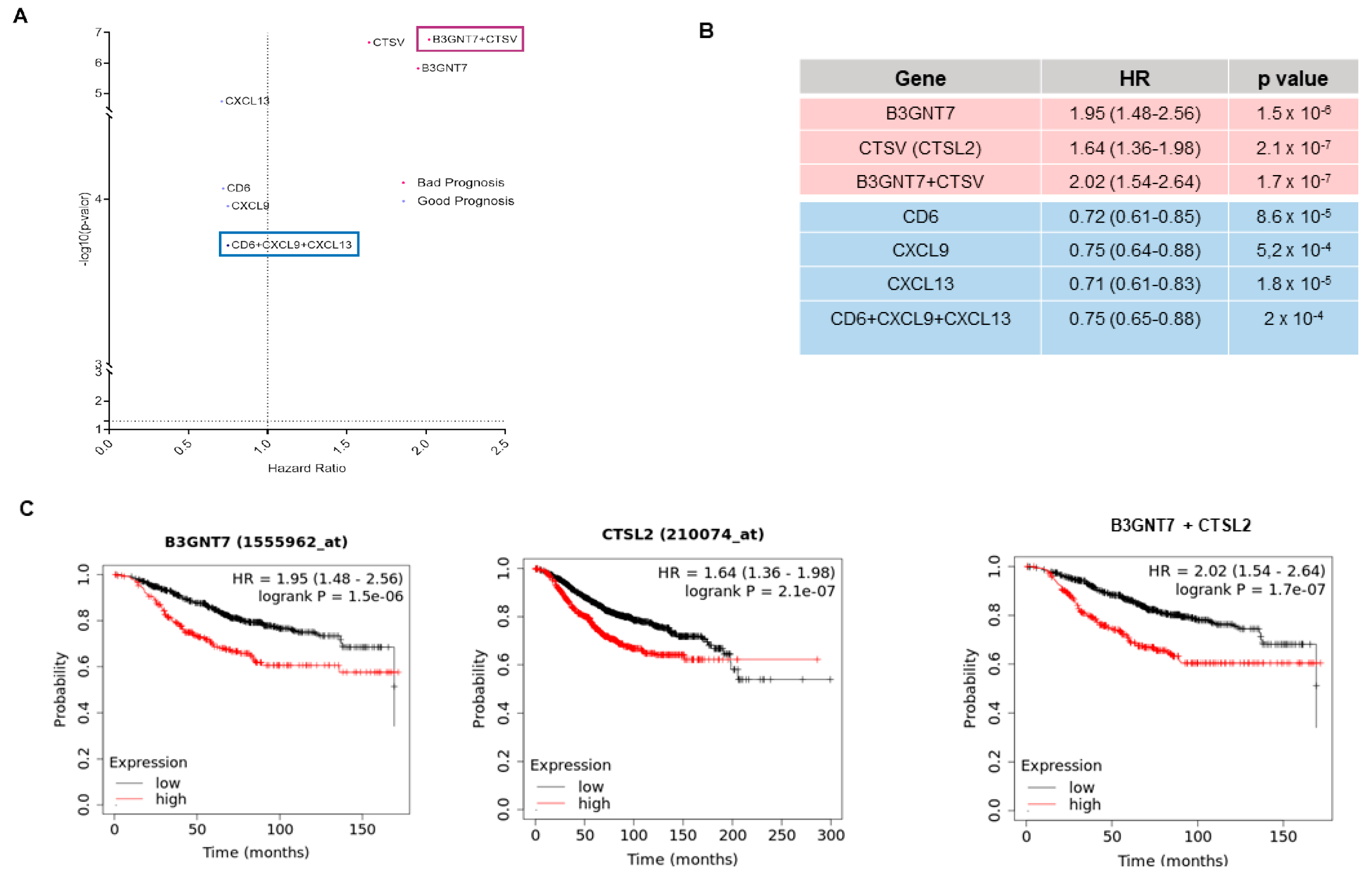
3.5. Identification of Outcome-Related Surfaceome Biomarkers
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kaplon, H.; Chenoweth, A.; Crescioli, S.; Reichert, J.M. Antibodies to Watch in 2022. MAbs 2022, 14, 2014296. [Google Scholar] [CrossRef] [PubMed]
- Musolino, A.; Gradishar, W.J.; Rugo, H.S.; Nordstrom, J.L.; Rock, E.P.; Arnaldez, F.; Pegram, M.D. Role of Fcγ Receptors in HER2-Targeted Breast Cancer Therapy. J. Immunother. Cancer 2022, 10, 3171. [Google Scholar] [CrossRef] [PubMed]
- Chan, I.S.; Ewald, A.J. The Changing Role of Natural Killer Cells in Cancer Metastasis. J. Clin. Investig. 2022, 132, e143762. [Google Scholar] [CrossRef]
- Türeci, Ӧzlem; Mitnacht-Kraus, R.; Wöll, S.; Yamada, T.; Sahin, U. Characterization of Zolbetuximab in Pancreatic Cancer Models. Oncoimmunology 2019, 8, e1523096. [Google Scholar] [CrossRef]
- Clynes, R.A.; Towers, T.L.; Presta, L.G.; Ravetch, J.V. Inhibitory Fc Receptors Modulate in Vivo Cytoxicity against Tumor Targets. Nat. Med. 2000, 6, 443–446. [Google Scholar] [CrossRef]
- Ocaña, A.; García-Alonso, S.; Amir, E.; Pandiella, A. Refining Early Antitumoral Drug Development. Trends Pharmacol. Sci. 2018, 39, 922–925. [Google Scholar] [CrossRef]
- Ocana, A.; Pandiella, A. Targeting HER Receptors in Cancer. Curr. Pharm. Des. 2013, 19, 808–817. [Google Scholar] [CrossRef]
- Díaz-Rodríguez, E.; Gandullo-Sánchez, L.; Ocaña, A.; Pandiella, A. Novel ADCs and Strategies to Overcome Resistance to Anti-HER2 ADCs. Cancers 2022, 14, 154. [Google Scholar] [CrossRef]
- García-Alonso, S.; Ocaña, A.; Pandiella, A. Resistance to Antibody–Drug Conjugates. Cancer Res. 2018, 78, 2159–2165. [Google Scholar] [CrossRef]
- Bardia, A.; Hurvitz, S.A.; Tolaney, S.M.; Loirat, D.; Punie, K.; Oliveira, M.; Brufsky, A.; Sardesai, S.D.; Kalinsky, K.; Zelnak, A.B.; et al. Sacituzumab Govitecan in Metastatic Triple-Negative Breast Cancer. N. Engl. J. Med. 2021, 384, 1529–1541. [Google Scholar] [CrossRef]
- García-Alonso, S.; Ocaña, A.; Pandiella, A. Trastuzumab Emtansine: Mechanisms of Action and Resistance, Clinical Progress, and Beyond. Trends Cancer 2020, 6, 130–146. [Google Scholar] [CrossRef]
- Juan, A.; del Mar Noblejas-López, M.; Arenas-Moreira, M.; Alonso-Moreno, C.; Ocaña, A. Options to Improve the Action of PROTACs in Cancer: Development of Controlled Delivery Nanoparticles. Front. Cell Dev. Biol. 2021, 9, 805336. [Google Scholar] [CrossRef]
- McClain, M.R.; Palomaki, G.E.; Nathanson, K.L.; Haddow, J.E. Adjusting the Estimated Proportion of Breast Cancer Cases Associated with BRCA1 and BRCA2 Mutations: Public Health Implications. Genet. Med. 2005, 7, 28–33. [Google Scholar] [CrossRef]
- Mateo, J.; Lord, C.J.; Serra, V.; Tutt, A.; Balmaña, J.; Castroviejo-Bermejo, M.; Cruz, C.; Oaknin, A.; Kaye, S.B.; De Bono, J.S. A Decade of Clinical Development of PARP Inhibitors in Perspective. Ann. Oncol. 2019, 30, 1437. [Google Scholar] [CrossRef]
- Lord, C.J.; Ashworth, A. PARP Inhibitors: The First Synthetic Lethal Targeted Therapy. Science 2017, 355, 1152. [Google Scholar] [CrossRef]
- Lord, C.J.; Tutt, A.N.J.; Ashworth, A. Synthetic Lethality and Cancer Therapy: Lessons Learned from the Development of PARP Inhibitors. Annu. Rev. Med. 2015, 66, 455–470. [Google Scholar] [CrossRef]
- Huang, A.; Garraway, L.A.; Ashworth, A.; Weber, B. Synthetic Lethality as an Engine for Cancer Drug Target Discovery. Nat. Rev. Drug Discov. 2019, 19, 23–38. [Google Scholar] [CrossRef]
- Kim, D.; Nam, H.J. PARP Inhibitors: Clinical Limitations and Recent Attempts to Overcome Them. Int. J. Mol. Sci. 2022, 23, 8412. [Google Scholar] [CrossRef]
- Bausch-Fluck, D.; Goldmann, U.; Müller, S.; van Oostrum, M.; Müller, M.; Schubert, O.T.; Wollscheid, B. The in Silico Human Surfaceome. Proc. Natl. Acad. Sci. USA 2018, 115, E10988–E10997. [Google Scholar] [CrossRef]
- Lánczky, A.; Győrffy, B. Web-Based Survival Analysis Tool Tailored for Medical Research (KMplot): Development and Implementation. J. Med. Internet Res. 2021, 23, e27633. [Google Scholar] [CrossRef]
- Li, T.; Fan, J.; Wang, B.; Traugh, N.; Chen, Q.; Liu, J.S.; Li, B.; Liu, X.S. TIMER: A Web Server for Comprehensive Analysis of Tumor-Infiltrating Immune Cells. Cancer Res. 2017, 77, e108. [Google Scholar] [CrossRef]
- Li, T.; Fu, J.; Zeng, Z.; Cohen, D.; Li, J.; Chen, Q.; Li, B.; Liu, X.S. TIMER2.0 for Analysis of Tumor-Infiltrating Immune Cells. Nucleic Acids Res. 2020, 48, W509. [Google Scholar] [CrossRef]
- Jin, H.; Wang, L.; Bernards, R. Rational Combinations of Targeted Cancer Therapies: Background, Advances and Challenges. Nat. Rev. Drug Discov. 2022, 22, 213–234. [Google Scholar] [CrossRef]
- Varma Shrivastav, S.; Bhardwaj, A.; Pathak, K.A.; Shrivastav, A. Insulin-Like Growth Factor Binding Protein-3 (IGFBP-3): Unraveling the Role in Mediating IGF-Independent Effects Within the Cell. Front. Cell Dev. Biol. 2020, 8, 286. [Google Scholar] [CrossRef]
- Gurrea-Rubio, M.; Fox, D.A. The Dual Role of CD6 as a Therapeutic Target in Cancer and Autoimmune Disease. Front. Med. 2022, 9, 1026521. [Google Scholar] [CrossRef]
- Velasco-de Andrés, M.; Casadó-Llombart, S.; Català, C.; Leyton-Pereira, A.; Lozano, F.; Aranda, F. Soluble CD5 and CD6: Lymphocytic Class I Scavenger Receptors as Immunotherapeutic Agents. Cells 2020, 9, 2589. [Google Scholar] [CrossRef]
- Hsieh, C.H.; Jian, C.Z.; Lin, L.I.; Low, G.S.; Ou, P.Y.; Hsu, C.; Ou, D.L. Potential Role of CXCL13/CXCR5 Signaling in Immune Checkpoint Inhibitor Treatment in Cancer. Cancers 2022, 14, 294. [Google Scholar] [CrossRef]
- Reschke, R.; Gajewski, T.F. CXCL9 and CXCL10 Bring the Heat to Tumors. Sci. Immunol. 2022, 7, eabq6509. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sanvicente, A.; Nieto-Jiménez, C.; Morafraile, E.C.; Díaz-Tejeiro, C.; Barberán, V.G.; Segura, P.P.; Balázs, G.; Ocaña, A. Identification of Deregulated Proteins in Mutated BRCA1/2 Breast and Ovarian Cancers for Vectorized Biologics. Cancers 2025, 17, 2208. https://doi.org/10.3390/cancers17132208
Sanvicente A, Nieto-Jiménez C, Morafraile EC, Díaz-Tejeiro C, Barberán VG, Segura PP, Balázs G, Ocaña A. Identification of Deregulated Proteins in Mutated BRCA1/2 Breast and Ovarian Cancers for Vectorized Biologics. Cancers. 2025; 17(13):2208. https://doi.org/10.3390/cancers17132208
Chicago/Turabian StyleSanvicente, Adrián, Cristina Nieto-Jiménez, Esther Cabañas Morafraile, Cristina Díaz-Tejeiro, Vanesa García Barberán, Pedro Pérez Segura, Győrffy Balázs, and Alberto Ocaña. 2025. "Identification of Deregulated Proteins in Mutated BRCA1/2 Breast and Ovarian Cancers for Vectorized Biologics" Cancers 17, no. 13: 2208. https://doi.org/10.3390/cancers17132208
APA StyleSanvicente, A., Nieto-Jiménez, C., Morafraile, E. C., Díaz-Tejeiro, C., Barberán, V. G., Segura, P. P., Balázs, G., & Ocaña, A. (2025). Identification of Deregulated Proteins in Mutated BRCA1/2 Breast and Ovarian Cancers for Vectorized Biologics. Cancers, 17(13), 2208. https://doi.org/10.3390/cancers17132208