

Tropisetron Suppresses Chronic Pancreatitis and Pancreatic Cancer by Blocking Interleukin 33 Expression

 , , , and
, , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Lines and Transfection
2.2. Small Molecule Screening
2.3. Quantitative PCR
2.4. Protein Studies
2.5. Animal Studies
2.6. Chronic Pancreatitis
2.7. Caerulein-Mediated Pancreatic Cancer
2.8. Tropisetron Treatment
2.9. Histology and Immunofluorescence
2.10. Statistical Analysis
3. Results
3.1. Tropisetron Is a Potent Inhibitor of IL-33 by Blocking IRF3 Phosphorylation
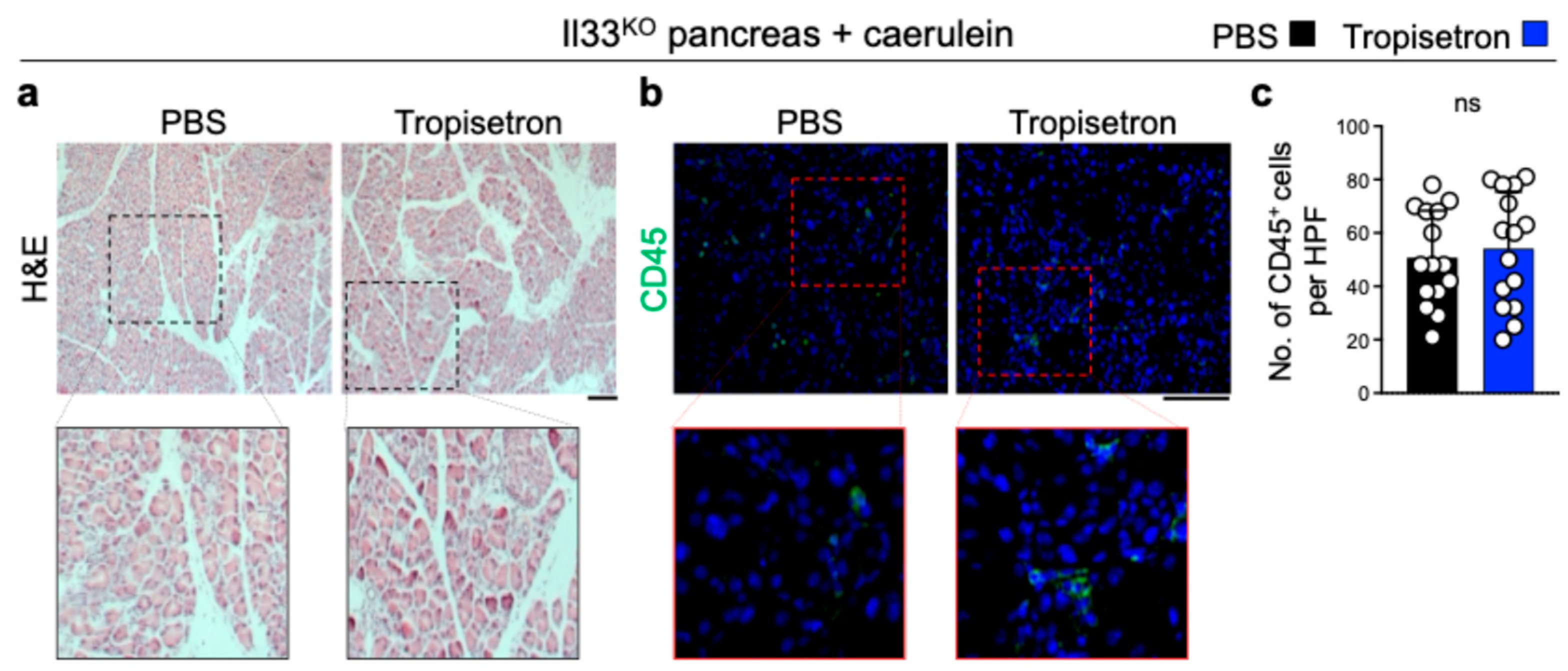
3.2. Tropisetron Alleviates Chronic Pancreatitis by Blocking IL-33
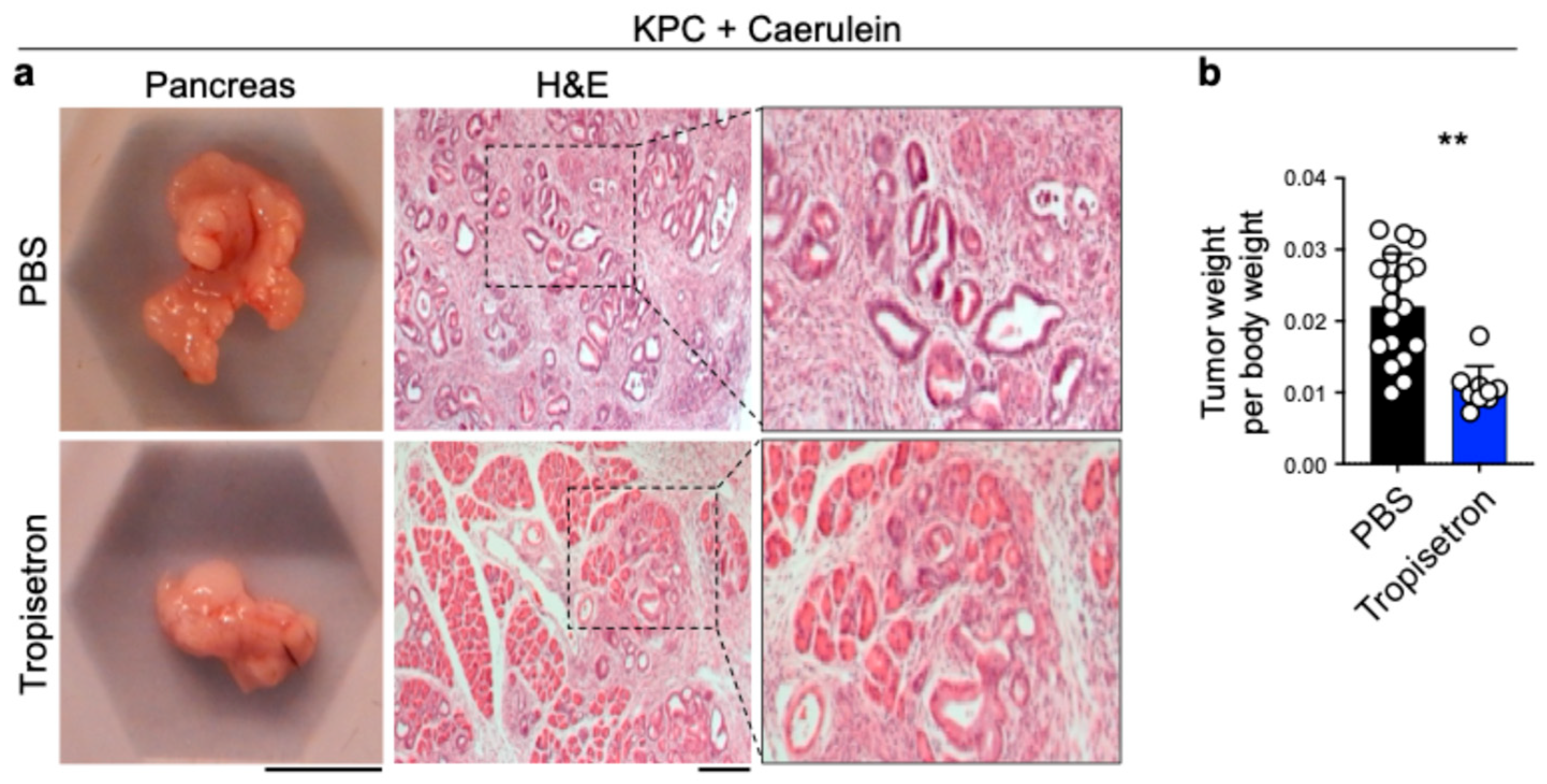
3.3. Tropisetron Prevents Pancreatic Cancer Development in Chronic Pancreatitis by Blocking IL-33
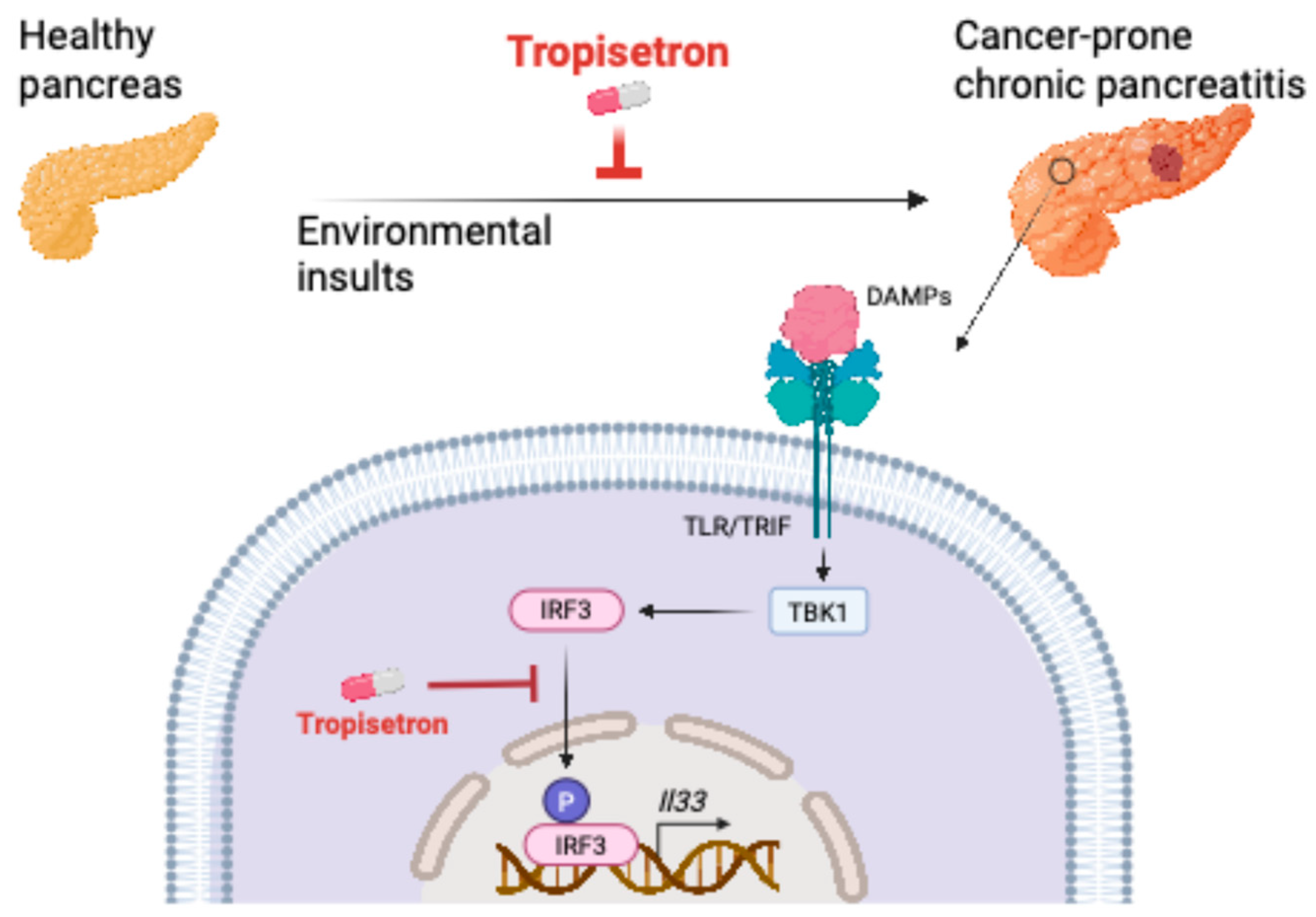
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhao, H.; Wu, L.; Yan, G.; Chen, Y.; Zhou, M.; Wu, Y.; Li, Y. Inflammation and tumor progression: Signaling pathways and targeted intervention. Signal Transduct. Target. Ther. 2021, 6, 263. [Google Scholar] [CrossRef] [PubMed]
- Grivennikov, S.I.; Greten, F.R.; Karin, M. Immunity, inflammation, and cancer. Cell 2010, 140, 883–899. [Google Scholar] [CrossRef] [PubMed]
- Cayrol, C.; Girard, J.P. Interleukin-33 (IL-33): A nuclear cytokine from the IL-1 family. Immunol. Rev. 2018, 281, 154–168. [Google Scholar] [CrossRef]
- Drake, L.Y.; Kita, H. IL-33: Biological properties, functions, and roles in airway disease. Immunol. Rev. 2017, 278, 173–184. [Google Scholar] [CrossRef]
- Ameri, A.H.; Tuchayi, S.M.; Zaalberg, A.; Park, J.H.; Ngo, K.H.; Li, T.; Lopez, E.; Colonna, M.; Lee, R.T.; Mino-Kenudson, M.; et al. IL-33/regulatory T cell axis triggers the development of a tumor-promoting immune environment in chronic inflammation. Proc. Natl. Acad. Sci. USA 2019, 116, 2646–2651. [Google Scholar] [CrossRef] [PubMed]
- Park, J.H.; Ameri, A.H.; Dempsey, K.E.; Conrad, D.N.; Kem, M.; Mino-Kenudson, M.; Demehri, S. Nuclear IL-33/SMAD signaling axis promotes cancer development in chronic inflammation. EMBO J. 2021, 40, e106151. [Google Scholar] [CrossRef]
- Yi, X.M.; Lian, H.; Li, S. Signaling and functions of interleukin-33 in immune regulation and diseases. Cell Insight 2022, 1, 100042. [Google Scholar] [CrossRef]
- Chan, B.C.L.; Lam, C.W.K.; Tam, L.S.; Wong, C.K. IL33: Roles in Allergic Inflammation and Therapeutic Perspectives. Front. Immunol. 2019, 10, 364. [Google Scholar] [CrossRef]
- Wang, S.; Ding, L.; Liu, S.S.; Wang, C.; Leng, R.X.; Chen, G.M.; Fan, Y.G.; Pan, H.F.; Ye, D.Q. IL-33: A potential therapeutic target in autoimmune diseases. J. Investig. Med. 2012, 60, 1151–1156. [Google Scholar] [CrossRef]
- Dong, Y.; Zhong, J.; Dong, L. IL-33 in Rheumatic Diseases. Front. Med. 2021, 8, 739489. [Google Scholar] [CrossRef]
- Drake, L.Y.; Prakash, Y.S. Contributions of IL-33 in Non-hematopoietic Lung Cells to Obstructive Lung Disease. Front. Immunol. 2020, 11, 1798. [Google Scholar] [CrossRef] [PubMed]
- Makrinioti, H.; Toussaint, M.; Jackson, D.J.; Walton, R.P.; Johnston, S.L. Role of interleukin 33 in respiratory allergy and asthma. Lancet Respir. Med. 2014, 2, 226–237. [Google Scholar] [CrossRef] [PubMed]
- Choi, M.R.; Sosman, J.A.; Zhang, B. The Janus Face of IL-33 Signaling in Tumor Development and Immune Escape. Cancers 2021, 13, 3281. [Google Scholar] [CrossRef]
- Fournie, J.J.; Poupot, M. The Pro-tumorigenic IL-33 Involved in Antitumor Immunity: A Yin and Yang Cytokine. Front. Immunol. 2018, 9, 2506. [Google Scholar] [CrossRef]
- Wasmer, M.H.; Krebs, P. The Role of IL-33-Dependent Inflammation in the Tumor Microenvironment. Front. Immunol. 2016, 7, 682. [Google Scholar] [CrossRef]
- Shen, J.X.; Liu, J.; Zhang, G.J. Interleukin-33 in Malignancies: Friends or Foes? Front. Immunol. 2018, 9, 3051. [Google Scholar] [CrossRef]
- Zhang, X.; Chen, W.; Zeng, P.; Xu, J.; Diao, H. The Contradictory Role of Interleukin-33 in Immune Cells and Tumor Immunity. Cancer Manag. Res. 2020, 12, 7527–7537. [Google Scholar] [CrossRef]
- Liu, J.; Shen, J.X.; Hu, J.L.; Huang, W.H.; Zhang, G.J. Significance of interleukin-33 and its related cytokines in patients with breast cancers. Front. Immunol. 2014, 5, 141. [Google Scholar] [CrossRef] [PubMed]
- Fang, M.; Li, Y.; Huang, K.; Qi, S.; Zhang, J.; Zgodzinski, W.; Majewski, M.; Wallner, G.; Gozdz, S.; Macek, P.; et al. IL33 Promotes Colon Cancer Cell Stemness via JNK Activation and Macrophage Recruitment. Cancer Res. 2017, 77, 2735–2745. [Google Scholar] [CrossRef]
- Yeoh, W.J.; Vu, V.P.; Krebs, P. IL-33 biology in cancer: An update and future perspectives. Cytokine 2022, 157, 155961. [Google Scholar] [CrossRef]
- Chen, S.F.; Nieh, S.; Jao, S.W.; Wu, M.Z.; Liu, C.L.; Chang, Y.C.; Lin, Y.S. The paracrine effect of cancer-associated fibroblast-induced interleukin-33 regulates the invasiveness of head and neck squamous cell carcinoma. J. Pathol. 2013, 231, 180–189. [Google Scholar] [CrossRef] [PubMed]
- Hodgson, A.; Wan, F. Interference with nuclear factor kappaB signaling pathway by pathogen-encoded proteases: Global and selective inhibition. Mol. Microbiol. 2016, 99, 439–452. [Google Scholar] [CrossRef] [PubMed]
- Polumuri, S.K.; Jayakar, G.G.; Shirey, K.A.; Roberts, Z.J.; Perkins, D.J.; Pitha, P.M.; Vogel, S.N. Transcriptional regulation of murine IL-33 by TLR and non-TLR agonists. J. Immunol. 2012, 189, 50–60. [Google Scholar] [CrossRef]
- Park, J.H.; Mortaja, M.; Son, H.G.; Zhao, X.; Sloat, L.M.; Azin, M.; Wang, J.; Collier, M.R.; Tummala, K.S.; Mandinova, A.; et al. Statin prevents cancer development in chronic inflammation by blocking interleukin 33 expression. Nat. Commun. 2024, 15, 4099. [Google Scholar] [CrossRef]
- Gandhi, S.; de la Fuente, J.; Murad, M.H.; Majumder, S. Chronic Pancreatitis Is a Risk Factor for Pancreatic Cancer, and Incidence Increases With Duration of Disease: A Systematic Review and Meta-analysis. Clin. Transl. Gastroenterol. 2022, 13, e00463. [Google Scholar] [CrossRef]
- Han, M.; Tran, T.P.T.; Oh, J.K. Chronic pancreatitis and cancer risk in a matched cohort study using national claims data in South Korea. Sci. Rep. 2022, 12, 5545. [Google Scholar] [CrossRef] [PubMed]
- Ilic, I.; Ilic, M. International patterns in incidence and mortality trends of pancreatic cancer in the last three decades: A joinpoint regression analysis. World J. Gastroenterol. 2022, 28, 4698–4715. [Google Scholar] [CrossRef]
- Adamska, A.; Domenichini, A.; Falasca, M. Pancreatic Ductal Adenocarcinoma: Current and Evolving Therapies. Int. J. Mol. Sci. 2017, 18, 1338. [Google Scholar] [CrossRef]
- Strickler, J.H.; Satake, H.; George, T.J.; Yaeger, R.; Hollebecque, A.; Garrido-Laguna, I.; Schuler, M.; Burns, T.F.; Coveler, A.L.; Falchook, G.S.; et al. Sotorasib in KRAS p.G12C-Mutated Advanced Pancreatic Cancer. N. Engl. J. Med. 2023, 388, 33–43. [Google Scholar] [CrossRef]
- Timmer, F.E.F.; Geboers, B.; Nieuwenhuizen, S.; Dijkstra, M.; Schouten, E.A.C.; Puijk, R.S.; de Vries, J.J.J.; van den Tol, M.P.; Bruynzeel, A.M.E.; Streppel, M.M.; et al. Pancreatic Cancer and Immunotherapy: A Clinical Overview. Cancers 2021, 13, 4138. [Google Scholar] [CrossRef]
- Hu, Z.I.; O’Reilly, E.M. Therapeutic developments in pancreatic cancer. Nat. Rev. Gastroenterol. Hepatol. 2024, 21, 7–24. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.R.; Plosker, G.L.; McTavish, D. Tropisetron. A review of its pharmacodynamic and pharmacokinetic properties, and therapeutic potential as an antiemetic. Drugs 1993, 46, 925–943. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.R.; Yen, T.H.; Lim, S.N.; Perng, M.D.; Lin, C.Y.; Su, M.Y.; Yeh, C.T.; Chiu, C.T. Granulocyte colony-stimulating factor reduces fibrosis in a mouse model of chronic pancreatitis. PLoS ONE 2014, 9, e116229. [Google Scholar] [CrossRef] [PubMed]
- Komar, H.M.; Serpa, G.; Kerscher, C.; Schwoegl, E.; Mace, T.A.; Jin, M.; Yang, M.C.; Chen, C.S.; Bloomston, M.; Ostrowski, M.C.; et al. Inhibition of Jak/STAT signaling reduces the activation of pancreatic stellate cells in vitro and limits caerulein-induced chronic pancreatitis in vivo. Sci. Rep. 2017, 7, 1787. [Google Scholar] [CrossRef]
- Collins, M.A.; Brisset, J.C.; Zhang, Y.; Bednar, F.; Pierre, J.; Heist, K.A.; Galban, C.J.; Galban, S.; di Magliano, M.P. Metastatic pancreatic cancer is dependent on oncogenic Kras in mice. PLoS ONE 2012, 7, e49707. [Google Scholar] [CrossRef]
- Murakami, S.; Shahbazian, D.; Surana, R.; Zhang, W.; Chen, H.; Graham, G.T.; White, S.M.; Weiner, L.M.; Yi, C. Yes-associated protein mediates immune reprogramming in pancreatic ductal adenocarcinoma. Oncogene 2017, 36, 1232–1244. [Google Scholar] [CrossRef]
- Natarajan, C.; Yao, S.Y.; Sriram, S. TLR3 Agonist Poly-IC Induces IL-33 and Promotes Myelin Repair. PLoS ONE 2016, 11, e0152163. [Google Scholar] [CrossRef]
- Zhao, X.J.; Dong, Q.; Bindas, J.; Piganelli, J.D.; Magill, A.; Reiser, J.; Kolls, J.K. TRIF and IRF-3 binding to the TNF promoter results in macrophage TNF dysregulation and steatosis induced by chronic ethanol. J. Immunol. 2008, 181, 3049–3056. [Google Scholar] [CrossRef]
- Voskuhl, R.R.; Peterson, R.S.; Song, B.; Ao, Y.; Morales, L.B.; Tiwari-Woodruff, S.; Sofroniew, M.V. Reactive astrocytes form scar-like perivascular barriers to leukocytes during adaptive immune inflammation of the CNS. J. Neurosci. 2009, 29, 11511–11522. [Google Scholar] [CrossRef]
- Park, S.J.; Kim, B.; Choi, S.; Balasubramaniam, S.; Lee, S.C.; Lee, J.Y.; Kim, H.S.; Kim, J.Y.; Kim, J.J.; Lee, Y.A.; et al. Imaging inflammation using an activated macrophage probe with Slc18b1 as the activation-selective gating target. Nat. Commun. 2019, 10, 1111. [Google Scholar] [CrossRef]
- Chen, S.M.; Chieng, W.W.; Huang, S.W.; Hsu, L.J.; Jan, M.S. The synergistic tumor growth-inhibitory effect of probiotic Lactobacillus on transgenic mouse model of pancreatic cancer treated with gemcitabine. Sci. Rep. 2020, 10, 20319. [Google Scholar] [CrossRef]
- Kawasaki, T.; Kawai, T. Toll-like receptor signaling pathways. Front. Immunol. 2014, 5, 461. [Google Scholar] [CrossRef] [PubMed]
- Spilman, P.; Descamps, O.; Gorostiza, O.; Peters-Libeu, C.; Poksay, K.S.; Matalis, A.; Campagna, J.; Patent, A.; Rao, R.; John, V.; et al. The multi-functional drug tropisetron binds APP and normalizes cognition in a murine Alzheimer’s model. Brain Res. 2014, 1551, 25–44. [Google Scholar] [CrossRef] [PubMed]
- Mousavizadeh, K.; Rahimian, R.; Fakhfouri, G.; Aslani, F.S.; Ghafourifar, P. Anti-inflammatory effects of 5-HT receptor antagonist, tropisetron on experimental colitis in rats. Eur. J. Clin. Investig. 2009, 39, 375–383. [Google Scholar] [CrossRef]
- Ozkan, A.; Yildiz, I.; Yuksel, L.; Apak, H.; Celkan, T. Tropisetron (Navoban) in the control of nausea and vomiting induced by combined cancer chemotherapy in children. Jpn. J. Clin. Oncol. 1999, 29, 92–95. [Google Scholar] [CrossRef] [PubMed]
- Nau, F., Jr.; Miller, J.; Saravia, J.; Ahlert, T.; Yu, B.; Happel, K.I.; Cormier, S.A.; Nichols, C.D. Serotonin 5-HT(2) receptor activation prevents allergic asthma in a mouse model. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2015, 308, L191–L198. [Google Scholar] [CrossRef]
- Wu, H.; Denna, T.H.; Storkersen, J.N.; Gerriets, V.A. Beyond a neurotransmitter: The role of serotonin in inflammation and immunity. Pharmacol. Res. 2019, 140, 100–114. [Google Scholar] [CrossRef]
- Cloez-Tayarani, I.; Petit-Bertron, A.F.; Venters, H.D.; Cavaillon, J.M. Differential effect of serotonin on cytokine production in lipopolysaccharide-stimulated human peripheral blood mononuclear cells: Involvement of 5-hydroxytryptamine2A receptors. Int. Immunol. 2003, 15, 233–240. [Google Scholar] [CrossRef]
- Herr, N.; Bode, C.; Duerschmied, D. The Effects of Serotonin in Immune Cells. Front. Cardiovasc. Med. 2017, 4, 48. [Google Scholar] [CrossRef]
- Vega Lde, L.; Munoz, E.; Calzado, M.A.; Lieb, K.; Candelario-Jalil, E.; Gschaidmeir, H.; Farber, L.; Mueller, W.; Stratz, T.; Fiebich, B.L. The 5-HT3 receptor antagonist tropisetron inhibits T cell activation by targeting the calcineurin pathway. Biochem. Pharmacol. 2005, 70, 369–380. [Google Scholar] [CrossRef]
- Koyama, Y.; Kondo, M.; Shimada, S. Building a 5-HT3A Receptor Expression Map in the Mouse Brain. Sci. Rep. 2017, 7, 42884. [Google Scholar] [CrossRef]
- Irving, H.; Turek, I.; Kettle, C.; Yaakob, N. Tapping into 5-HT(3) Receptors to Modify Metabolic and Immune Responses. Int. J. Mol. Sci. 2021, 22, 11910. [Google Scholar] [CrossRef] [PubMed]
- Houbracken, I.; Baeyens, L.; Ravassard, P.; Heimberg, H.; Bouwens, L. Gene delivery to pancreatic exocrine cells in vivo and in vitro. BMC Biotechnol. 2012, 12, 74. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Ye, E.; Huang, J.; Xu, M.; Zhang, J.; Zhang, J.; Cai, H.; Zhang, J. Cysteine-modified PEGylated nanoparticles for targeted delivery of methylprednisolone to pancreatitis. Eur. J. Pharm. Biopharm. 2024, 195, 114179. [Google Scholar] [CrossRef] [PubMed]
- Bear, A.S.; Vonderheide, R.H.; O’Hara, M.H. Challenges and Opportunities for Pancreatic Cancer Immunotherapy. Cancer Cell 2020, 38, 788–802. [Google Scholar] [CrossRef]
- Ju, Y.; Xu, D.; Liao, M.M.; Sun, Y.; Bao, W.D.; Yao, F.; Ma, L. Barriers and opportunities in pancreatic cancer immunotherapy. NPJ Precis. Oncol. 2024, 8, 199. [Google Scholar] [CrossRef]
- Kabacaoglu, D.; Ciecielski, K.J.; Ruess, D.A.; Algul, H. Immune Checkpoint Inhibition for Pancreatic Ductal Adenocarcinoma: Current Limitations and Future Options. Front. Immunol. 2018, 9, 1878. [Google Scholar] [CrossRef]
- Kawashima, S.; Togashi, Y. Resistance to immune checkpoint inhibitors and the tumor microenvironment. Exp. Dermatol. 2023, 32, 240–249. [Google Scholar] [CrossRef]
- Sammarco, E.; Rossetti, M.; Salfi, A.; Bonato, A.; Viacava, P.; Masi, G.; Galli, L.; Faviana, P. Tumor microenvironment and clinical efficacy of first line immunotherapy-based combinations in metastatic renal cell carcinoma. Med. Oncol. 2024, 41, 150. [Google Scholar] [CrossRef]
- Xiao, P.; Wan, X.; Cui, B.; Liu, Y.; Qiu, C.; Rong, J.; Zheng, M.; Song, Y.; Chen, L.; He, J.; et al. Interleukin 33 in tumor microenvironment is crucial for the accumulation and function of myeloid-derived suppressor cells. Oncoimmunology 2016, 5, e1063772. [Google Scholar] [CrossRef]
- Moral, J.A.; Leung, J.; Rojas, L.A.; Ruan, J.; Zhao, J.; Sethna, Z.; Ramnarain, A.; Gasmi, B.; Gururajan, M.; Redmond, D.; et al. ILC2s amplify PD-1 blockade by activating tissue-specific cancer immunity. Nature 2020, 579, 130–135. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bae, A.-N.; Mortaja, M.; Yeung, Y.; Huang, J.; Park, J.H.; Demehri, S. Tropisetron Suppresses Chronic Pancreatitis and Pancreatic Cancer by Blocking Interleukin 33 Expression. Cancers 2025, 17, 2087. https://doi.org/10.3390/cancers17132087
Bae A-N, Mortaja M, Yeung Y, Huang J, Park JH, Demehri S. Tropisetron Suppresses Chronic Pancreatitis and Pancreatic Cancer by Blocking Interleukin 33 Expression. Cancers. 2025; 17(13):2087. https://doi.org/10.3390/cancers17132087
Chicago/Turabian StyleBae, An-Na, Mahsa Mortaja, YeePui Yeung, Jiao Huang, Jong Ho Park, and Shadmehr Demehri. 2025. "Tropisetron Suppresses Chronic Pancreatitis and Pancreatic Cancer by Blocking Interleukin 33 Expression" Cancers 17, no. 13: 2087. https://doi.org/10.3390/cancers17132087
APA StyleBae, A.-N., Mortaja, M., Yeung, Y., Huang, J., Park, J. H., & Demehri, S. (2025). Tropisetron Suppresses Chronic Pancreatitis and Pancreatic Cancer by Blocking Interleukin 33 Expression. Cancers, 17(13), 2087. https://doi.org/10.3390/cancers17132087