
TP53 Mutation-Specific Dysregulation of Store-Operated Calcium Entry and Apoptotic Sensitivity in Triple-Negative Breast Cancer

 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Bioinformatics
2.2. Cell Culture
2.3. CACNA1D siRNA Knockdown
2.4. Real-Time PCR
2.5. Calcium Measurement
2.6. Proliferation Assay
2.7. Apoptosis Assay
2.8. Statistical Analysis
3. Results
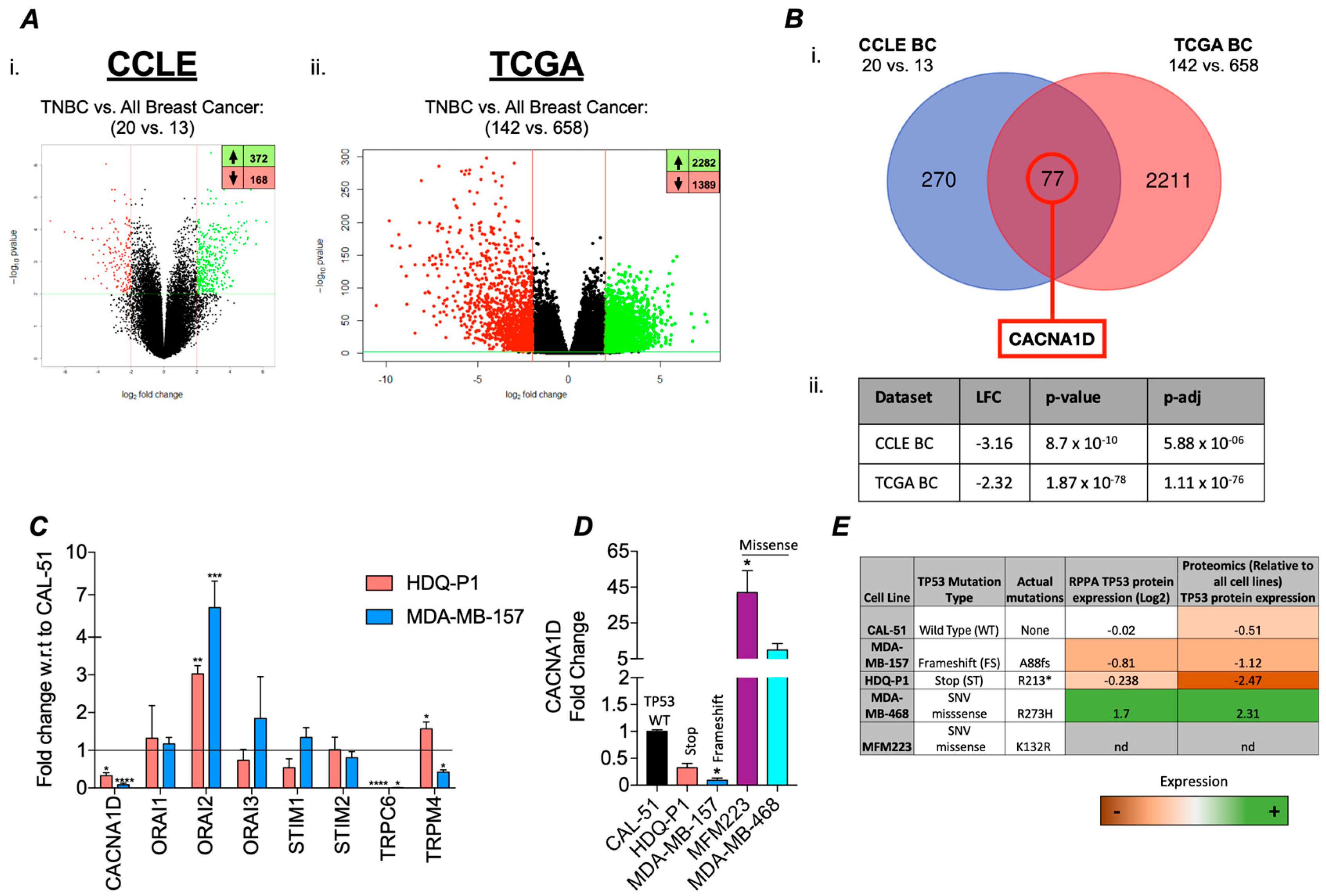
3.1. TP53 Mutations Associated with Differential Ca2+ Channel Gene Expression in TNBC
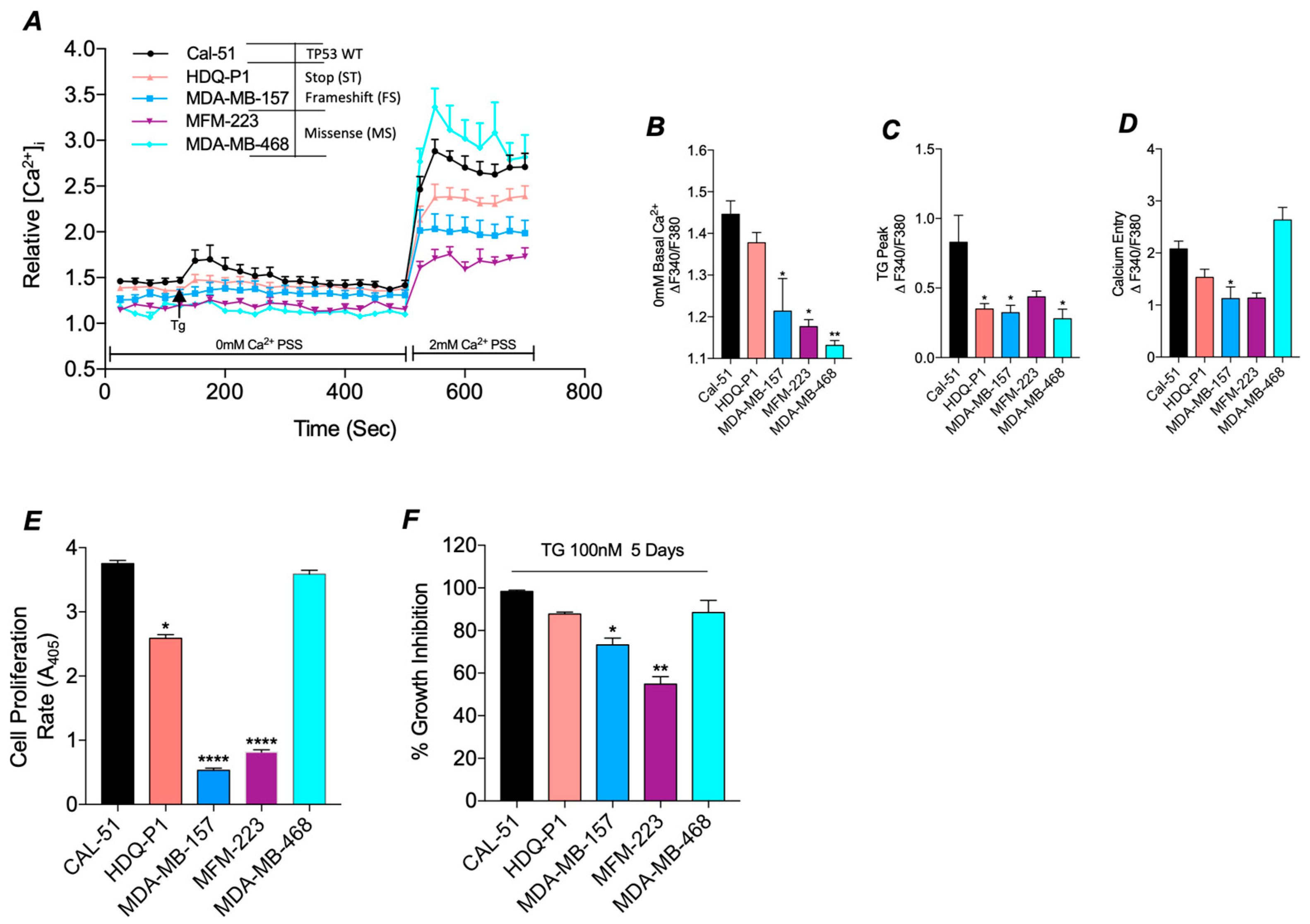
3.2. Mutant TP53 Impacts SOC and Proliferation in TNBC
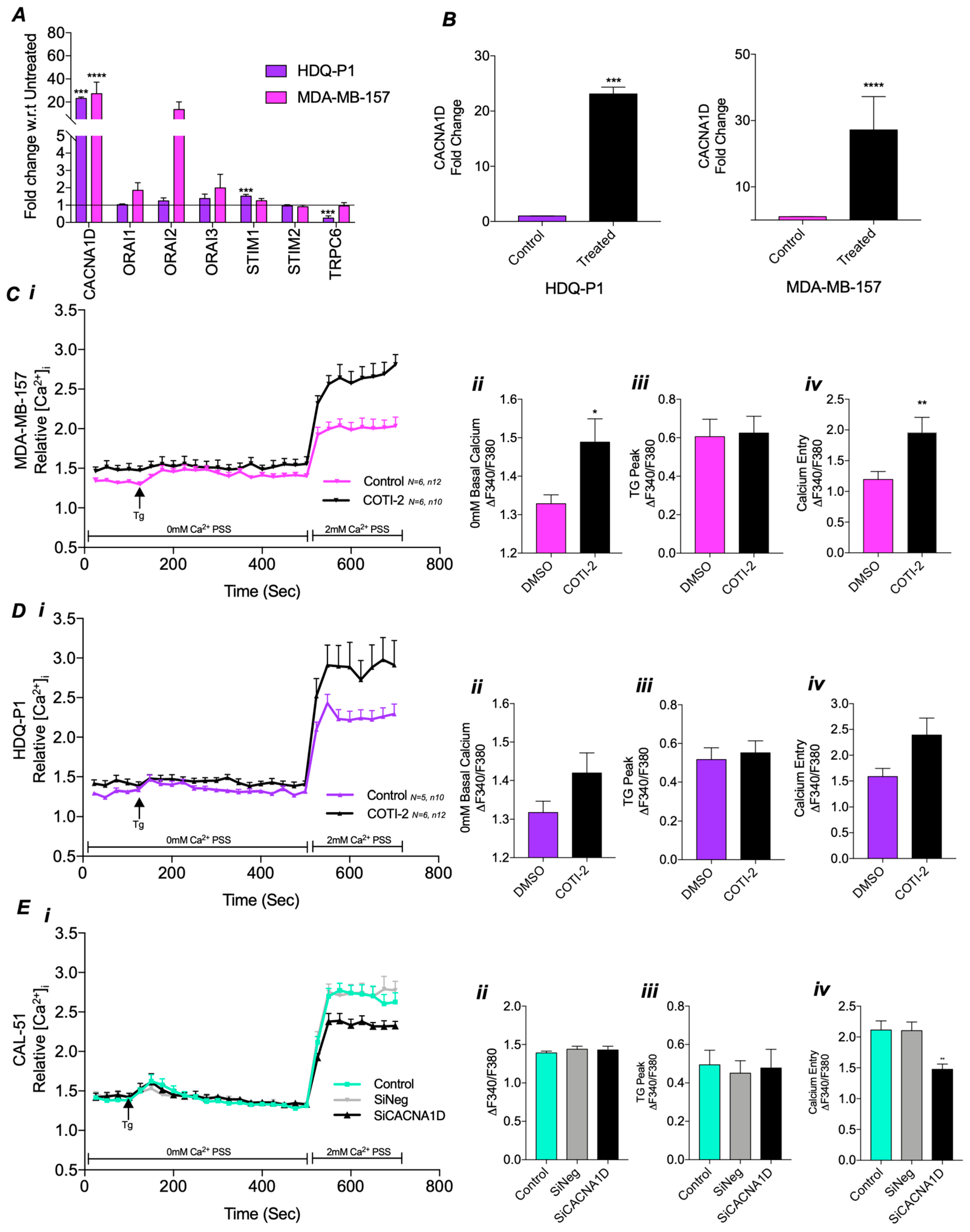
3.3. Mutant TP53 Modulates SOCE Through CACNA1D
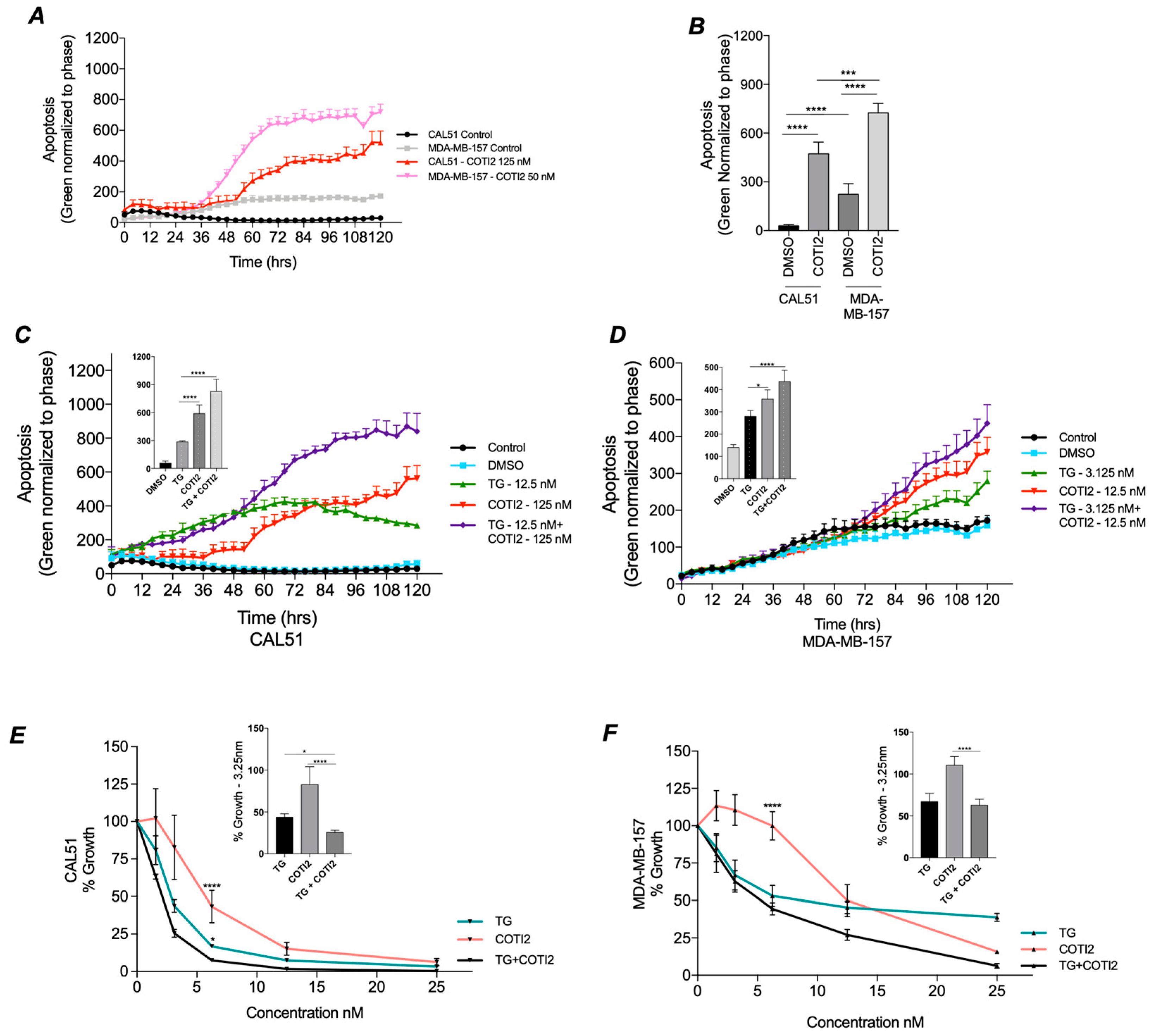
3.4. COTI-2 Increases Apoptosis and Sensitivity to Treatment in Both TP53 Wild-Type and Mutant TNBC
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AC | All Cancer |
| BC | Breast cancer |
| CCLE | Cancer Cell Line Encyclopaedia |
| DEA | Differential expression analyses |
| DEG | Differentially expressed genes |
| DGEA | Differential Gene Expression Analysis |
| ER | Estrogen receptor |
| FBS | Fetal Bovine Serum |
| FS | Frameshift |
| HER2 | Human epidermal growth factor receptor 2 |
| LFC | Log2 fold change |
| Mut | Mutant |
| PM | Plasma Membrane |
| PR | Progesterone receptor |
| SERCA | Sarcoendoplasmic Reticulum Calcium ATPase pump |
| SNV | Single Nucleotide Variant |
| SOC | Store operated current |
| SOCE | Store operated calcium entry |
| TCGA | The Cancer Genome Atlas |
| TG | Thapsigargin |
| TNBC | Triple negative breast cancer |
| TP53 | Tumour protein 53 |
| TRP | Transient receptor potential channel |
| TSG | Tumour suppressor gene |
| WT | Wild-type |
References
- Bray, F.; Laversanne, M.; Sung, H.; Ferlay, J.; Siegel, R.L.; Soerjomataram, I.; Jemal, A. Global Cancer Statistics 2022: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2024, 74, 229–263. [Google Scholar] [CrossRef] [PubMed]
- Bou Zerdan, M.; Ghorayeb, T.; Saliba, F.; Allam, S.; Bou Zerdan, M.; Yaghi, M.; Bilani, N.; Jaafar, R.; Nahleh, Z. Triple Negative Breast Cancer: Updates on Classification and Treatment in 2021. Cancers 2022, 14, 1253. [Google Scholar] [CrossRef] [PubMed]
- Bianchini, G.; Balko, J.M.; Mayer, I.A.; Sanders, M.E.; Gianni, L. Triple-Negative Breast Cancer: Challenges and Opportunities of a Heterogeneous Disease. Nat. Rev. Clin. Oncol. 2016, 13, 674–690. [Google Scholar] [CrossRef]
- Lakin, N.D.; Jackson, S.P. Regulation of P53 in Response to DNA Damage. Oncogene 1999, 18, 7644–7655. [Google Scholar] [CrossRef]
- Abuetabh, Y.; Wu, H.H.; Chai, C.; Al Yousef, H.; Persad, S.; Sergi, C.M.; Leng, R. DNA Damage Response Revisited: The P53 Family and Its Regulators Provide Endless Cancer Therapy Opportunities. Exp. Mol. Med. 2022, 54, 1658–1669. [Google Scholar] [CrossRef] [PubMed]
- Duffy, M.J.; Synnott, N.C.; Crown, J. Mutant P53 as a Target for Cancer Treatment. Eur. J. Cancer 2017, 83, 258–265. [Google Scholar] [CrossRef]
- Tate, J.G.; Bamford, S.; Jubb, H.C.; Sondka, Z.; Beare, D.M.; Bindal, N.; Boutselakis, H.; Cole, C.G.; Creatore, C.; Dawson, E.; et al. COSMIC: The Catalogue Of Somatic Mutations In Cancer. Nucleic Acids Res. 2019, 47, D941–D947. [Google Scholar] [CrossRef]
- Hu, J.; Cao, J.; Topatana, W.; Juengpanich, S.; Li, S.; Zhang, B.; Shen, J.; Cai, L.; Cai, X.; Chen, M. Targeting Mutant P53 for Cancer Therapy: Direct and Indirect Strategies. J. Hematol. Oncol. 2021, 14, 157. [Google Scholar] [CrossRef]
- Muller, P.A.J.; Vousden, K.H. P53 Mutations in Cancer. Nat. Cell Biol. 2013, 15, 2–8. [Google Scholar] [CrossRef]
- Mantovani, F.; Collavin, L.; Del Sal, G. Mutant P53 as a Guardian of the Cancer Cell. Cell Death Differ. 2019, 26, 199–212. [Google Scholar] [CrossRef]
- Duffy, M.J.; Crown, J. Drugging “Undruggable” Genes for Cancer Treatment: Are We Making Progress? Int. J. Cancer 2021, 148, 8–17. [Google Scholar] [CrossRef]
- Duffy, M.J.; Synnott, N.C.; O’Grady, S.; Crown, J. Targeting P53 for the Treatment of Cancer. Semin. Cancer Biol. 2022, 79, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Synnott, N.C.; Bauer, M.R.; Madden, S.; Murray, A.; Klinger, R.; O’Donovan, N.; O’Connor, D.; Gallagher, W.M.; Crown, J.; Fersht, A.R.; et al. Mutant P53 as a Therapeutic Target for the Treatment of Triple-Negative Breast Cancer: Preclinical Investigation with the Anti-P53 Drug, PK11007. Cancer Lett. 2018, 414, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.; Yang, M.; Bi, R.; Wang, Y.; Wang, C.; Wei, X.; Zhang, Z.; Xie, X.; Wei, W. Targeting Mutated P53 Dependency in Triple-Negative Breast Cancer Cells Through CDK7 Inhibition. Front. Oncol. 2021, 11, 664848. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Manero, G.; Goldberg, A.D.; Winer, E.S.; Altman, J.K.; Fathi, A.T.; Odenike, O.; Roboz, G.J.; Sweet, K.; Miller, C.; Wennborg, A.; et al. Eprenetapopt Combined with Venetoclax and Azacitidine in TP53-Mutated Acute Myeloid Leukaemia: A Phase 1, Dose-Finding and Expansion Study. Lancet Haematol. 2023, 10, e272–e283. [Google Scholar] [CrossRef]
- Riley, T.; Sontag, E.; Chen, P.; Levine, A. Transcriptional Control of Human P53-Regulated Genes. Nat. Rev. Mol. Cell Biol. 2008, 9, 402–412. [Google Scholar] [CrossRef]
- Wolter, K.G.; Hsu, Y.-T.; Smith, C.L.; Nechushtan, A.; Xi, X.-G.; Youle, R.J. Movement of Bax from the Cytosol to Mitochondria during Apoptosis. J. Cell Biol. 1997, 139, 1281–1292. [Google Scholar] [CrossRef]
- Eustace, A.J.; Lee, M.J.; Colley, G.; Roban, J.; Downing, T.; Buchanan, P.J. Aberrant Calcium Signalling Downstream of Mutations in TP53 and the PI3K/AKT Pathway Genes Promotes Disease Progression and Therapy Resistance in Triple Negative Breast Cancer. Cancer Drug Resist. 2022, 5, 560–576. [Google Scholar] [CrossRef]
- Prakriya, M.; Lewis, R.S. Store-Operated Calcium Channels. Physiol. Rev. 2015, 95, 1383–1436. [Google Scholar] [CrossRef]
- So, C.L.; Saunus, J.M.; Roberts-Thomson, S.J.; Monteith, G.R. Calcium Signalling and Breast Cancer. Semin. Cell Dev. Biol. 2019, 94, 74–83. [Google Scholar] [CrossRef]
- Bantsimba-Malanda, C.; Ahidouch, A.; Rodat-Despoix, L.; Ouadid-Ahidouch, H. Calcium Signal Modulation in Breast Cancer Aggressiveness. Cell Calcium 2023, 113, 102760. [Google Scholar] [CrossRef]
- Pratt, S.J.P.; Hernández-Ochoa, E.; Martin, S.S. Calcium Signaling: Breast Cancer’s Approach to Manipulation of Cellular Circuitry. Biophys. Rev. 2020, 12, 1343–1359. [Google Scholar] [CrossRef] [PubMed]
- Monteith, G.R.; Prevarskaya, N.; Roberts-Thomson, S.J. The Calcium–Cancer Signalling Nexus. Nat. Rev. Cancer 2017, 17, 367–380. [Google Scholar] [CrossRef]
- Romito, O.; Guéguinou, M.; Raoul, W.; Champion, O.; Robert, A.; Trebak, M.; Goupille, C.; Potier-Cartereau, M. Calcium Signaling: A Therapeutic Target to Overcome Resistance to Therapies in Cancer. Cell Calcium 2022, 108, 102673. [Google Scholar] [CrossRef] [PubMed]
- Kumari, N.; Pullaguri, N.; Rath, S.N.; Bajaj, A.; Sahu, V.; Ealla, K.K.R. Dysregulation of Calcium Homeostasis in Cancer and Its Role in Chemoresistance. Cancer Drug Resist 2024, 7, 11. [Google Scholar] [CrossRef]
- Bittremieux, M.; Bultynck, G. P53 and Ca2+ Signaling from the Endoplasmic Reticulum: Partners in Anti-Cancer Therapies. Oncoscience 2015, 2, 233–238. [Google Scholar] [CrossRef] [PubMed]
- Giorgi, C.; Bonora, M.; Sorrentino, G.; Missiroli, S.; Poletti, F.; Suski, J.M.; Galindo Ramirez, F.; Rizzuto, R.; Di Virgilio, F.; Zito, E.; et al. P53 at the Endoplasmic Reticulum Regulates Apoptosis in a Ca2+-Dependent Manner. Proc. Natl. Acad. Sci. USA 2015, 112, 1779–1784. [Google Scholar] [CrossRef]
- Madan, E.; Gogna, R.; Keppler, B.; Pati, U. P53 Increases Intra-Cellular Calcium Release by Transcriptional Regulation of Calcium Channel TRPC6 in GaQ3-Treated Cancer Cells. PLoS ONE 2013, 8, e71016. [Google Scholar] [CrossRef]
- Giorgi, C.; Bonora, M.; Missiroli, S.; Poletti, F.; Ramirez, F.G.; Morciano, G.; Morganti, C.; Pandolfi, P.P.; Mammano, F.; Pinton, P. Intravital Imaging Reveals P53-Dependent Cancer Cell Death Induced by Phototherapy via Calcium Signaling. Oncotarget 2014, 6, 1435–1445. [Google Scholar] [CrossRef]
- Wang, C.S.; Goulet, F.; Lavoie, J.; Drouin, R.; Auger, F.; Champetier, S.; Germain, L.; Têtu, B. Establishment and Characterization of a New Cell Line Derived from a Human Primary Breast Carcinoma. Cancer Genet. Cytogenet. 2000, 120, 58–72. [Google Scholar] [CrossRef]
- Lacroix, M.; Toillon, R.-A.; Leclercq, G. P53 and Breast Cancer, an Update. Endocr.-Relat. Cancer 2006, 13, 293–325. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, B.D.; Bauer, J.A.; Chen, X.; Sanders, M.E.; Chakravarthy, A.B.; Shyr, Y.; Pietenpol, J.A. Identification of Human Triple-Negative Breast Cancer Subtypes and Preclinical Models for Selection of Targeted Therapies. J. Clin. Investig. 2011, 121, 2750–2767. [Google Scholar] [CrossRef]
- Maleki Vareki, S.; Salim, K.Y.; Danter, W.R.; Koropatnick, J. Novel Anti-Cancer Drug COTI-2 Synergizes with Therapeutic Agents and Does Not Induce Resistance or Exhibit Cross-Resistance in Human Cancer Cell Lines. PLoS ONE 2018, 13, e0191766. [Google Scholar] [CrossRef] [PubMed]
- Salim, K.Y.; Vareki, S.M.; Danter, W.R.; San-Marina, S.; Koropatnick, J. Correction: COTI-2, a Novel Small Molecule That Is Active against Multiple Human Cancer Cell Lines in Vitro and in Vivo. Oncotarget 2017, 8, 60724. [Google Scholar] [CrossRef]
- Alam, M.; Rashid, S.; Fatima, K.; Adnan, M.; Shafie, A.; Akhtar, M.S.; Ganie, A.H.; Eldin, S.M.; Islam, A.; Khan, I.; et al. Biochemical Features and Therapeutic Potential of α-Mangostin: Mechanism of Action, Medicinal Values, and Health Benefits. Biomed. Pharmacother. 2023, 163, 114710. [Google Scholar] [CrossRef] [PubMed]
- Synnott, N.C.; O’Connell, D.; Crown, J.; Duffy, M.J. COTI-2 Reactivates Mutant P53 and Inhibits Growth of Triple-Negative Breast Cancer Cells. Breast Cancer Res Treat 2020, 179, 47–56. [Google Scholar] [CrossRef]
- O’Reilly, D.; Downing, T.; Kouba, S.; Potier-Cartereau, M.; McKenna, D.J.; Vandier, C.; Buchanan, P.J. CaV1.3 Enhanced Store Operated Calcium Promotes Resistance to Androgen Deprivation in Prostate Cancer. Cell Calcium 2022, 103, 102554. [Google Scholar] [CrossRef]
- Martin, A.; Clynes, M. Acid Phosphatase: Endpoint for in Vitro Toxicity Tests. Vitr. Cell. Dev. Biol.-Anim. 1991, 27, 183–184. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, T.; Su, W.; Dou, Z.; Zhao, D.; Jin, X.; Lei, H.; Wang, J.; Xie, X.; Cheng, B.; et al. Mutant P53 in Cancer: From Molecular Mechanism to Therapeutic Modulation. Cell Death Dis. 2022, 13, 974. [Google Scholar] [CrossRef]
- Pal, A.; Gonzalez-Malerva, L.; Eaton, S.; Xu, C.; Zhang, Y.; Grief, D.; Sakala, L.; Nwekwo, L.; Zeng, J.; Christensen, G.; et al. Multidimensional Quantitative Phenotypic and Molecular Analysis Reveals Neomorphic Behaviors of P53 Missense Mutants. npj Breast Cancer 2023, 9, 78. [Google Scholar] [CrossRef]
- Patergnani, S.; Danese, A.; Bouhamida, E.; Aguiari, G.; Previati, M.; Pinton, P.; Giorgi, C. Various Aspects of Calcium Signaling in the Regulation of Apoptosis, Autophagy, Cell Proliferation, and Cancer. Int. J. Mol. Sci. 2020, 21, 8323. [Google Scholar] [CrossRef] [PubMed]
- Ko, J.-H.; Ko, E.A.; Gu, W.; Lim, I.; Bang, H.; Zhou, T. Expression Profiling of Ion Channel Genes Predicts Clinical Outcome in Breast Cancer. Mol. Cancer 2013, 12, 106. [Google Scholar] [CrossRef]
- Lindemann, A.; Patel, A.A.; Silver, N.L.; Tang, L.; Liu, Z.; Wang, L.; Tanaka, N.; Rao, X.; Takahashi, H.; Maduka, N.K.; et al. COTI-2, A Novel Thiosemicarbazone Derivative, Exhibits Antitumor Activity in HNSCC through P53-Dependent and -Independent Mechanisms. Clin. Cancer Res. 2019, 25, 5650–5662. [Google Scholar] [CrossRef]
- Fourbon, Y.; Guéguinou, M.; Félix, R.; Constantin, B.; Uguen, A.; Fromont, G.; Lajoie, L.; Magaud, C.; Lecomte, T.; Chamorey, E.; et al. Ca2+ Protein Alpha 1D of CaV1.3 Regulates Intracellular Calcium Concentration and Migration of Colon Cancer Cells through a Non-Canonical Activity. Sci. Rep. 2017, 7, 14199. [Google Scholar] [CrossRef]
- Kappel, S.; Ross-Kaschitza, D.; Hauert, B.; Rother, K.; Peinelt, C. P53 Alters Intracellular Ca2+ Signaling through Regulation of TRPM4. Cell Calcium 2022, 104, 102591. [Google Scholar] [CrossRef] [PubMed]
- Dubois, C.; Vanden Abeele, F.; Prevarskaya, N. Targeting Apoptosis by the Remodelling of Calcium-Transporting Proteins in Cancerogenesis. FEBS J. 2013, 280, 5500–5510. [Google Scholar] [CrossRef]
- Moon, D.-O. Calcium’s Role in Orchestrating Cancer Apoptosis: Mitochondrial-Centric Perspective. Int. J. Mol. Sci. 2023, 24, 8982. [Google Scholar] [CrossRef] [PubMed]
- Zhu, G.; Pan, C.; Bei, J.-X.; Li, B.; Liang, C.; Xu, Y.; Fu, X. Mutant P53 in Cancer Progression and Targeted Therapies. Front. Oncol. 2020, 10, 595187. [Google Scholar] [CrossRef]
- Zhou, X.; Hao, Q.; Lu, H. Mutant P53 in Cancer Therapy-The Barrier or the Path. J. Mol. Cell Biol. 2019, 11, 293–305. [Google Scholar] [CrossRef]
- Mihara, M.; Erster, S.; Zaika, A.; Petrenko, O.; Chittenden, T.; Pancoska, P.; Moll, U.M. P53 Has a Direct Apoptogenic Role at the Mitochondria. Mol. Cell 2003, 11, 577–590. [Google Scholar] [CrossRef]
- Guo, Y.; Zhu, X.; Sun, X. COTI-2 Induces Cell Apoptosis in Pediatric Acute Lymphoblastic Leukemia via Upregulation of miR-203. Bioengineered 2020, 11, 201–208. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rabab, K.E.; Buchanan, P.J.; Colley, G.; White, A.; Murphy, A.; McCormack, C.; Eustace, A.J. TP53 Mutation-Specific Dysregulation of Store-Operated Calcium Entry and Apoptotic Sensitivity in Triple-Negative Breast Cancer. Cancers 2025, 17, 1614. https://doi.org/10.3390/cancers17101614
Rabab KE, Buchanan PJ, Colley G, White A, Murphy A, McCormack C, Eustace AJ. TP53 Mutation-Specific Dysregulation of Store-Operated Calcium Entry and Apoptotic Sensitivity in Triple-Negative Breast Cancer. Cancers. 2025; 17(10):1614. https://doi.org/10.3390/cancers17101614
Chicago/Turabian StyleRabab, Kaneez E., Paul J. Buchanan, Grace Colley, Anita White, Aisling Murphy, Chloe McCormack, and Alex J. Eustace. 2025. "TP53 Mutation-Specific Dysregulation of Store-Operated Calcium Entry and Apoptotic Sensitivity in Triple-Negative Breast Cancer" Cancers 17, no. 10: 1614. https://doi.org/10.3390/cancers17101614
APA StyleRabab, K. E., Buchanan, P. J., Colley, G., White, A., Murphy, A., McCormack, C., & Eustace, A. J. (2025). TP53 Mutation-Specific Dysregulation of Store-Operated Calcium Entry and Apoptotic Sensitivity in Triple-Negative Breast Cancer. Cancers, 17(10), 1614. https://doi.org/10.3390/cancers17101614