
Impact on the Transcriptome of Proton Beam Irradiation Targeted at Healthy Cardiac Tissue of Mice
 , ,
, ,  ,
,  , ,
, , 
 ,
,  ,
,  ,
,  and
and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
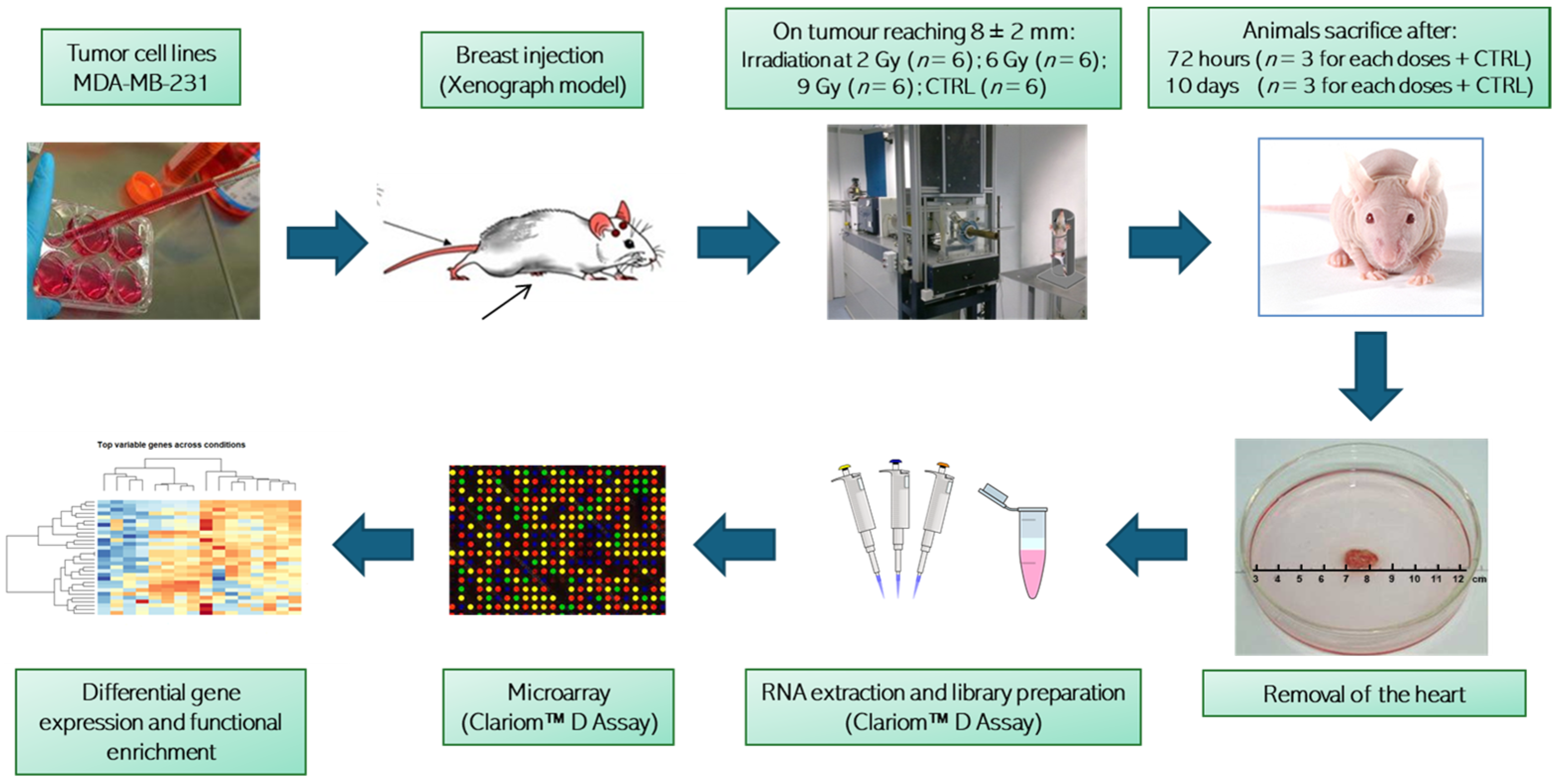
2.1. Ethics Statement and Animal Model
2.2. Animal Radiation Treatment
2.3. RNA Extraction and Microarray
2.4. Computational Analysis
2.4.1. Data Preprocessing
2.4.2. Differential Expression Analysis
2.4.3. Functional Enrichment Analysis
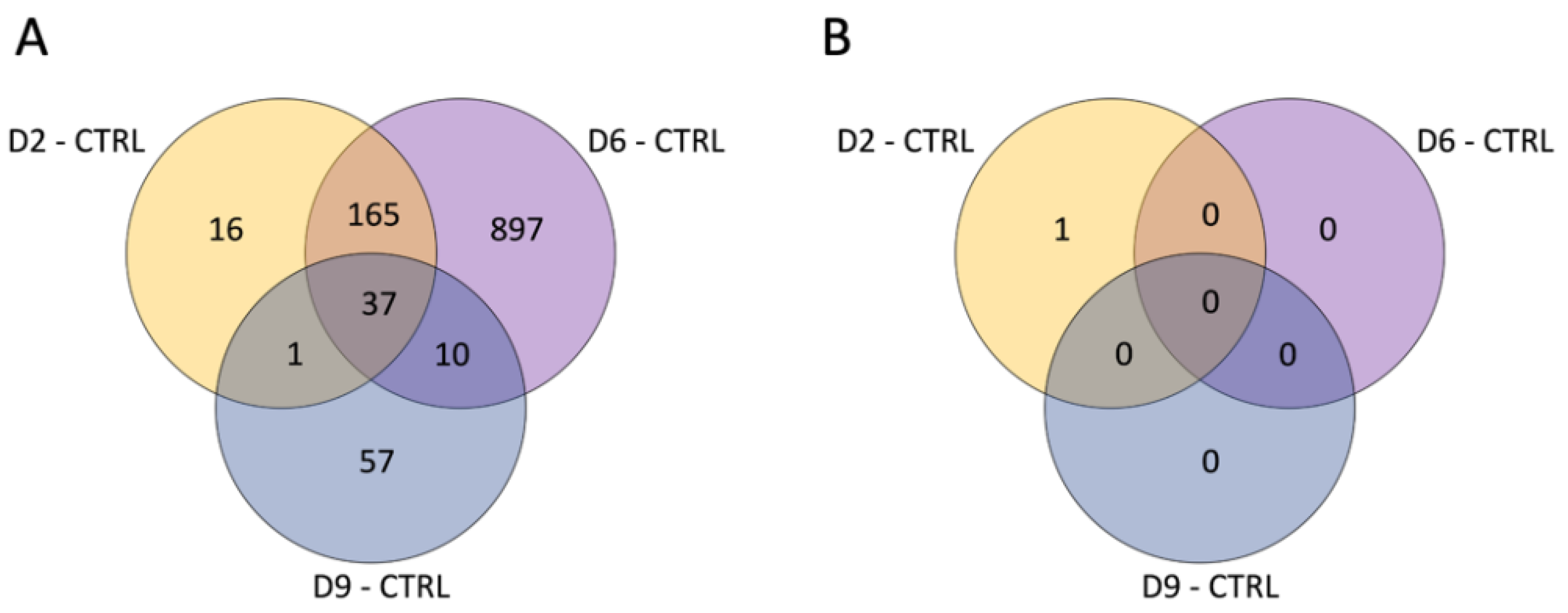
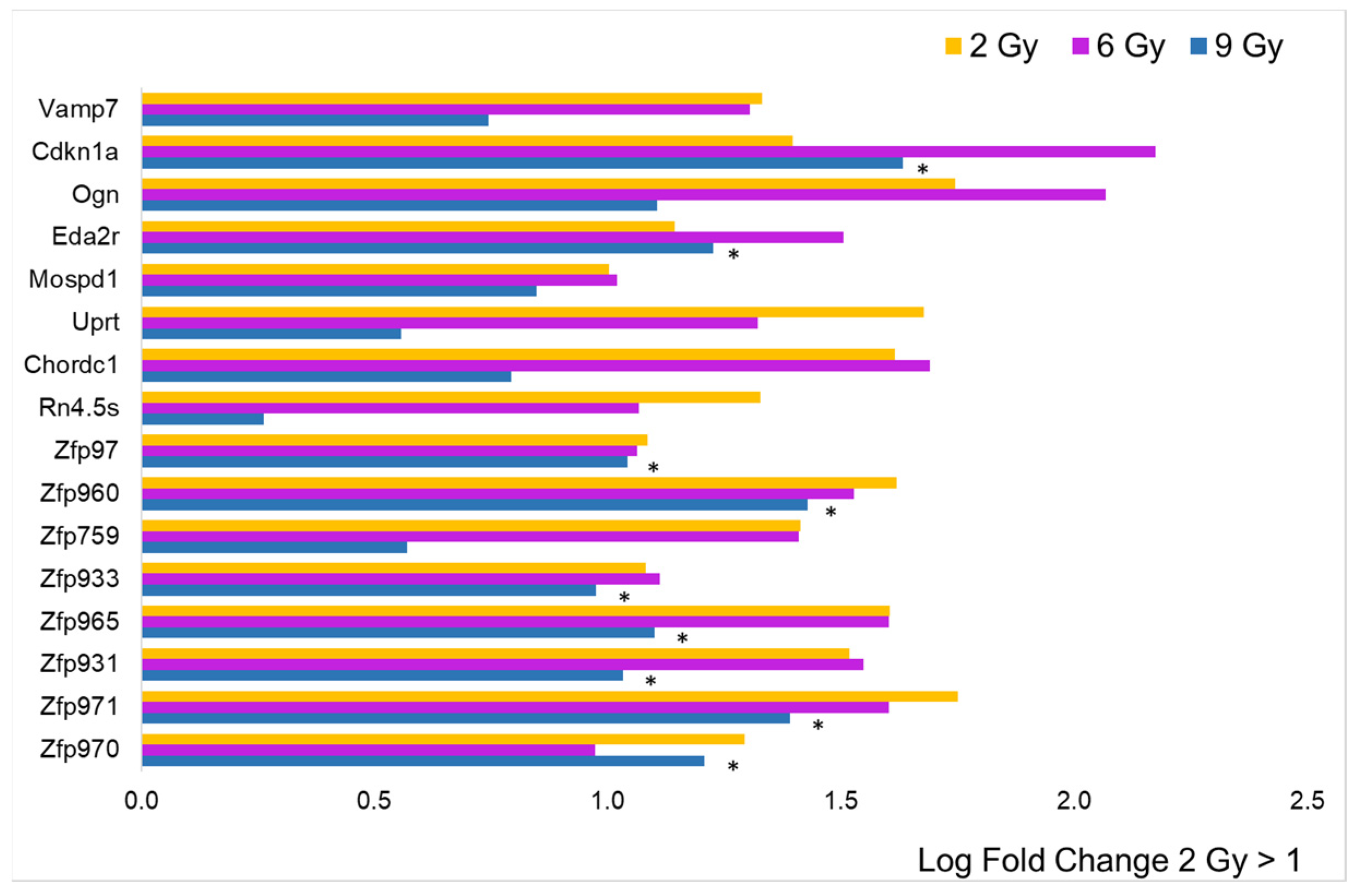
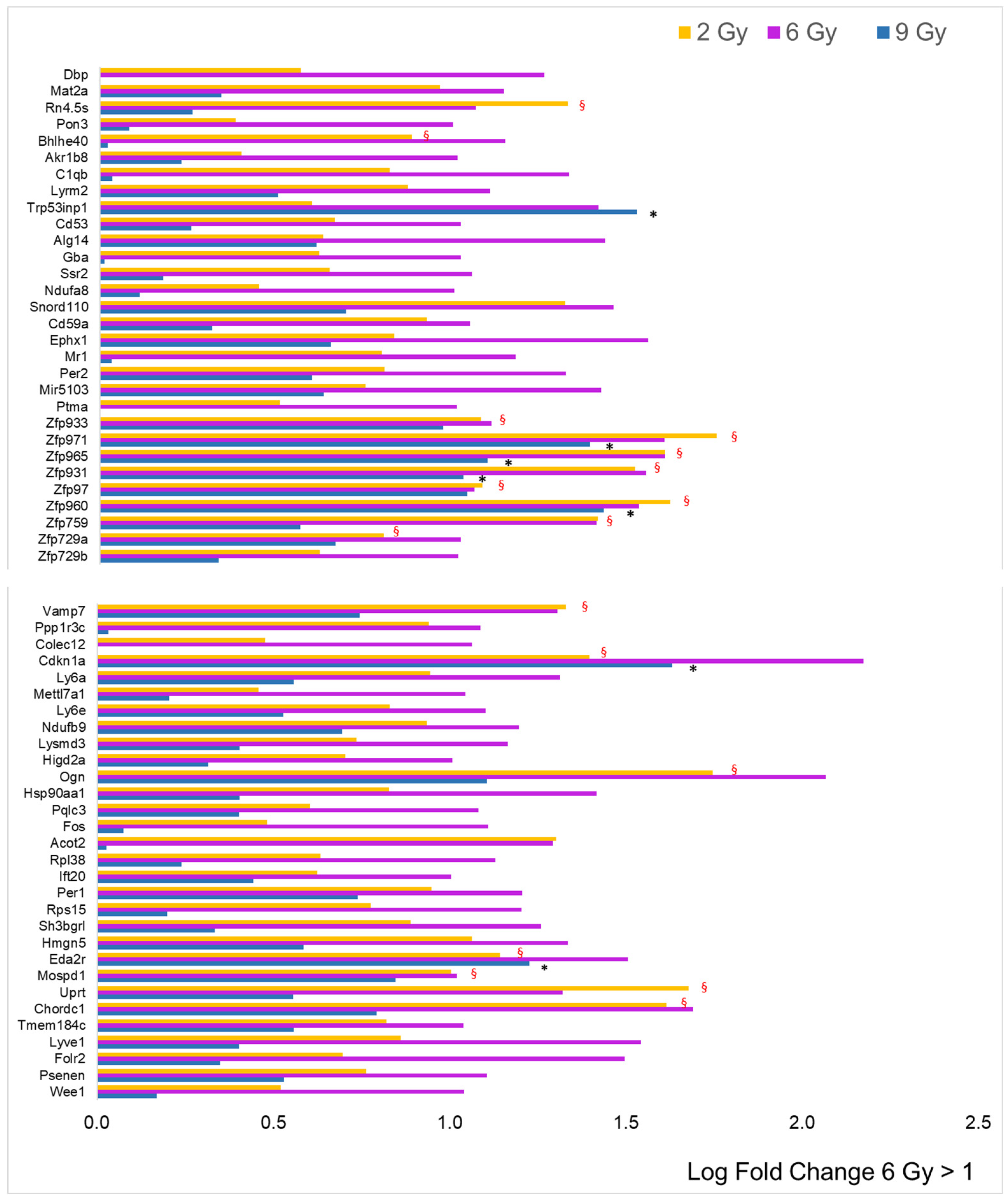
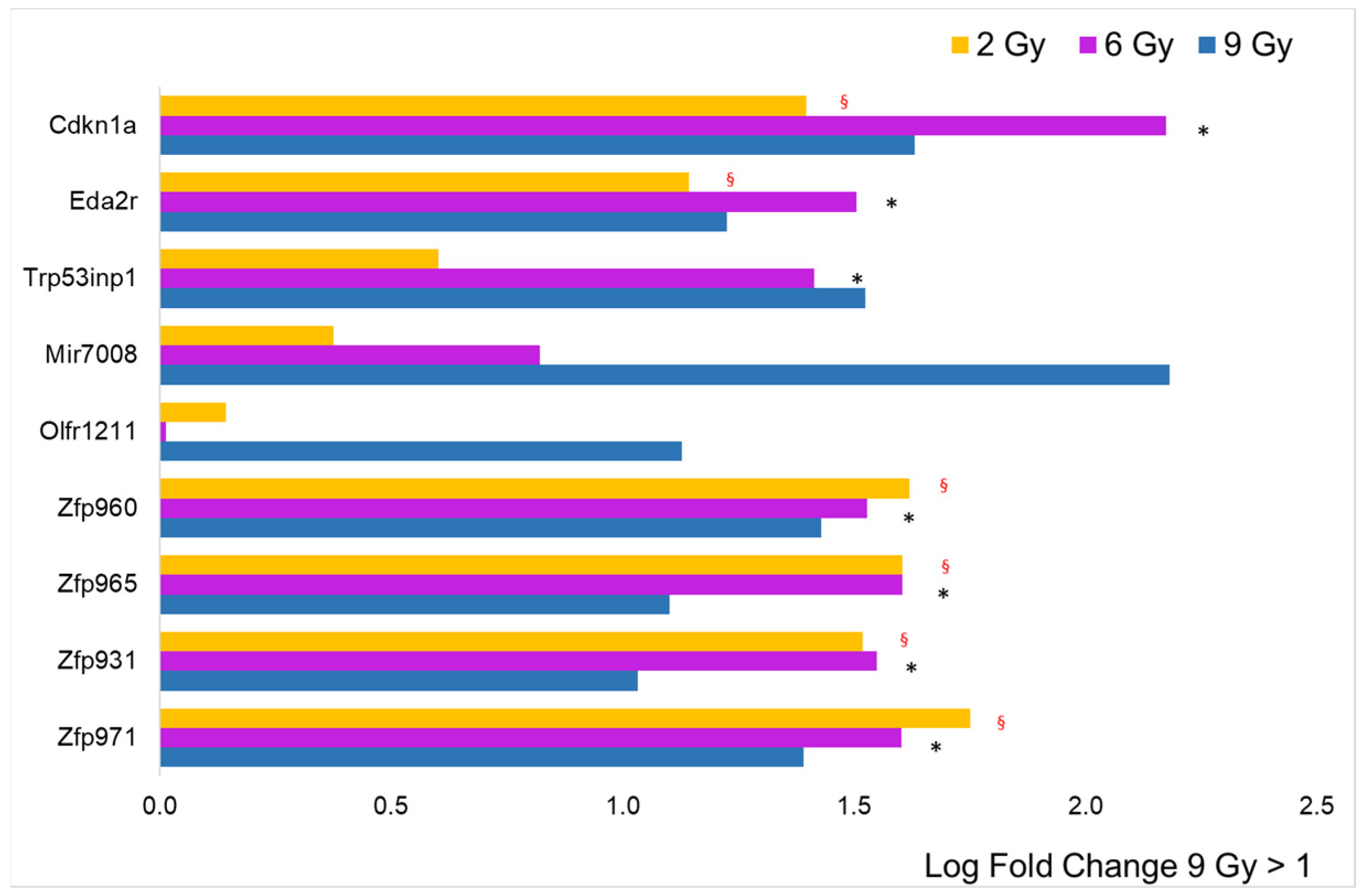
3. Results
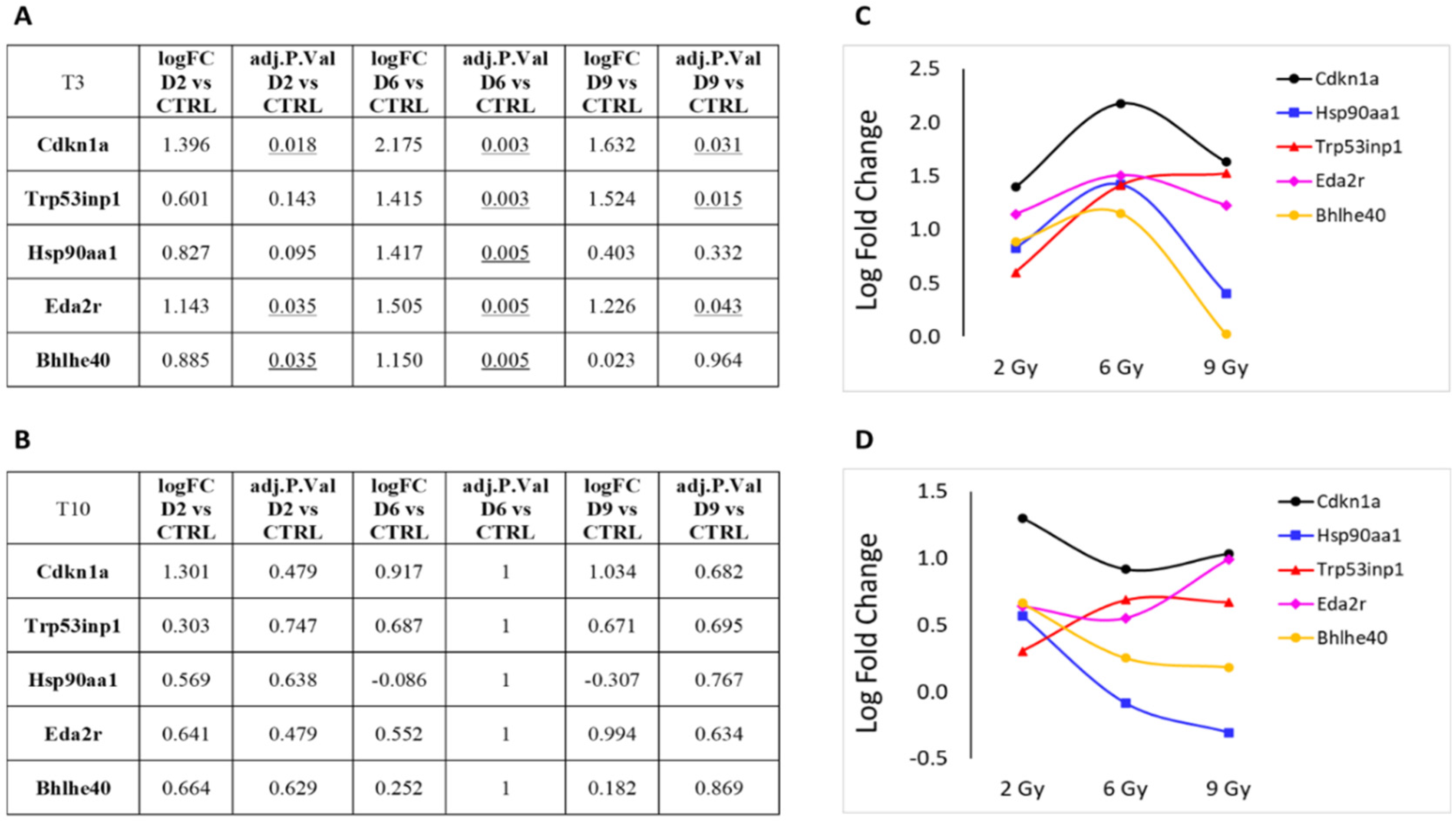
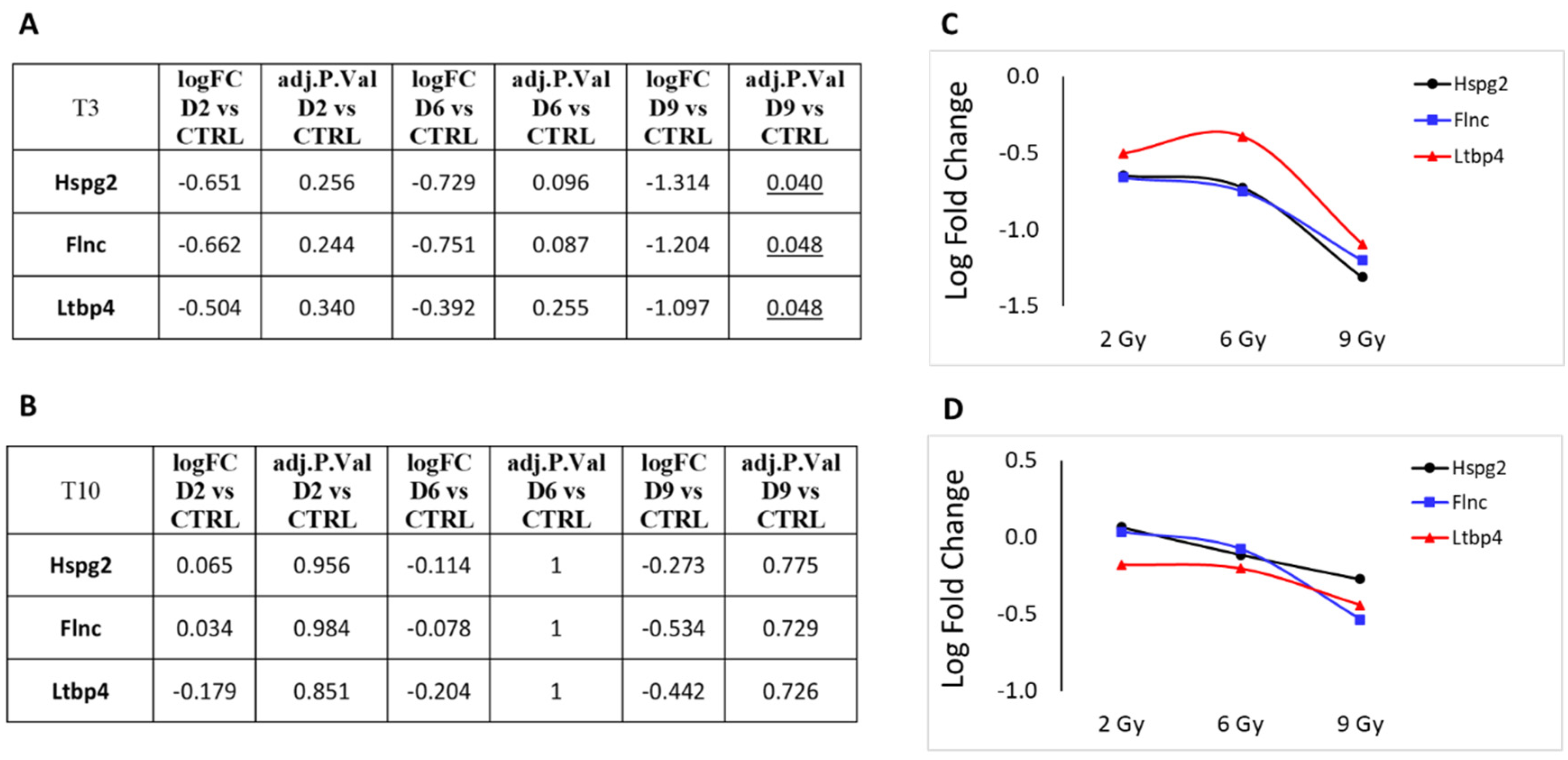
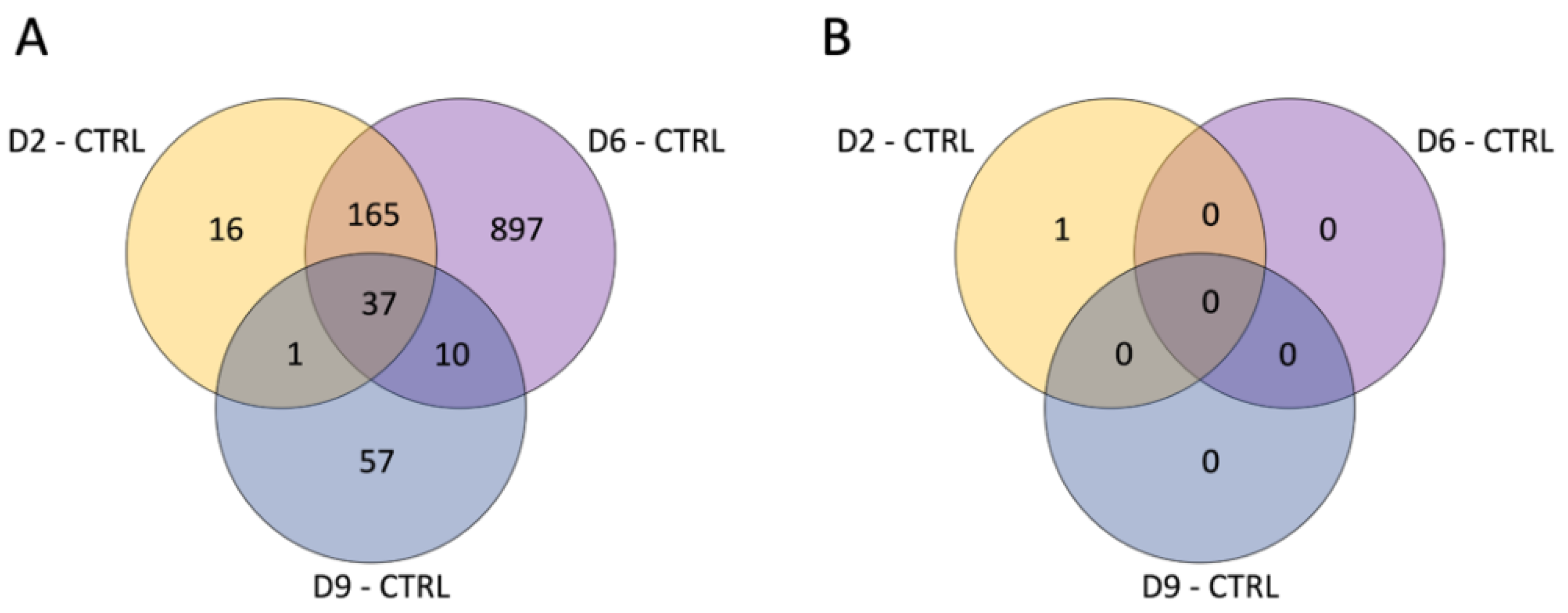
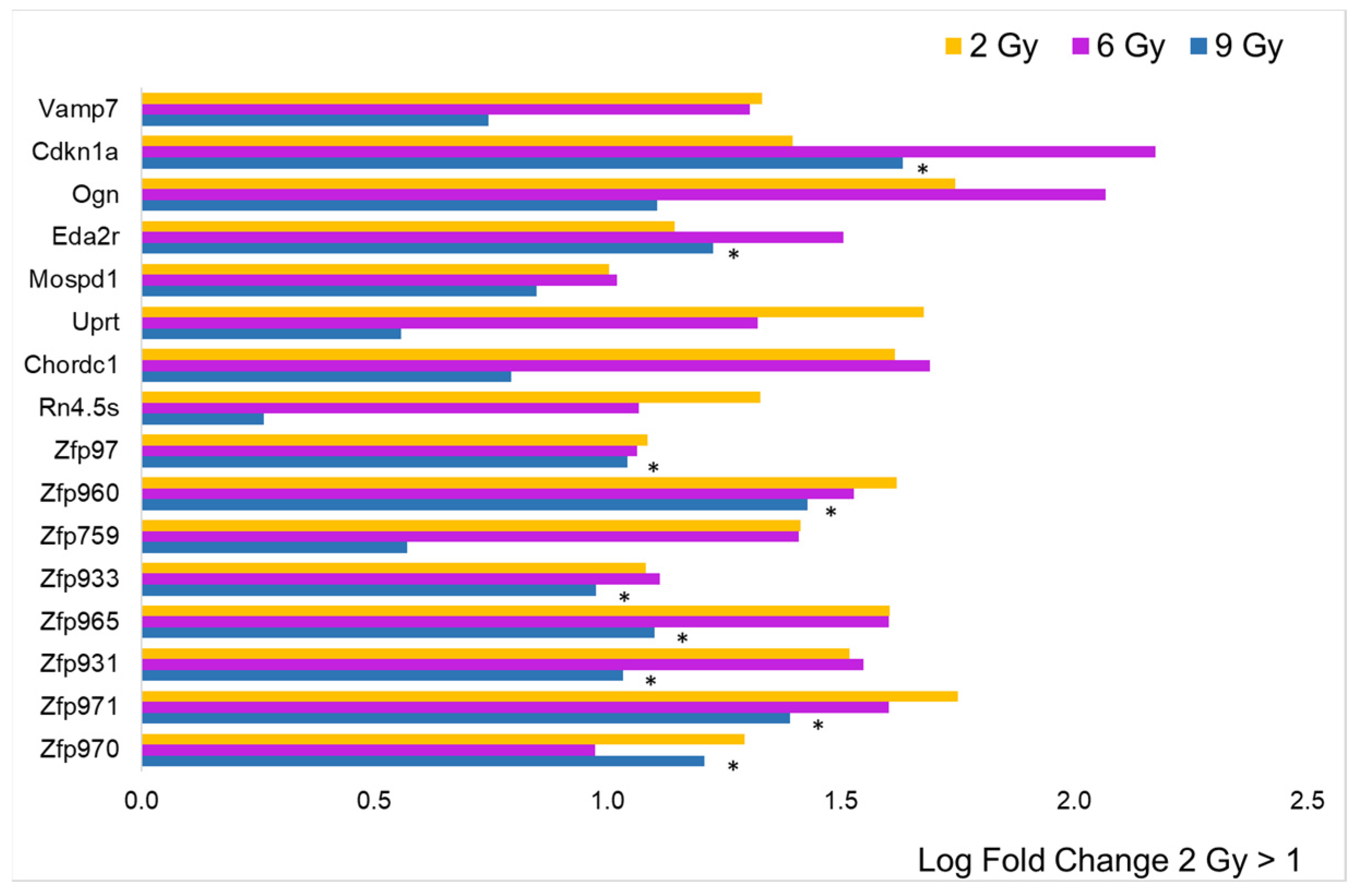
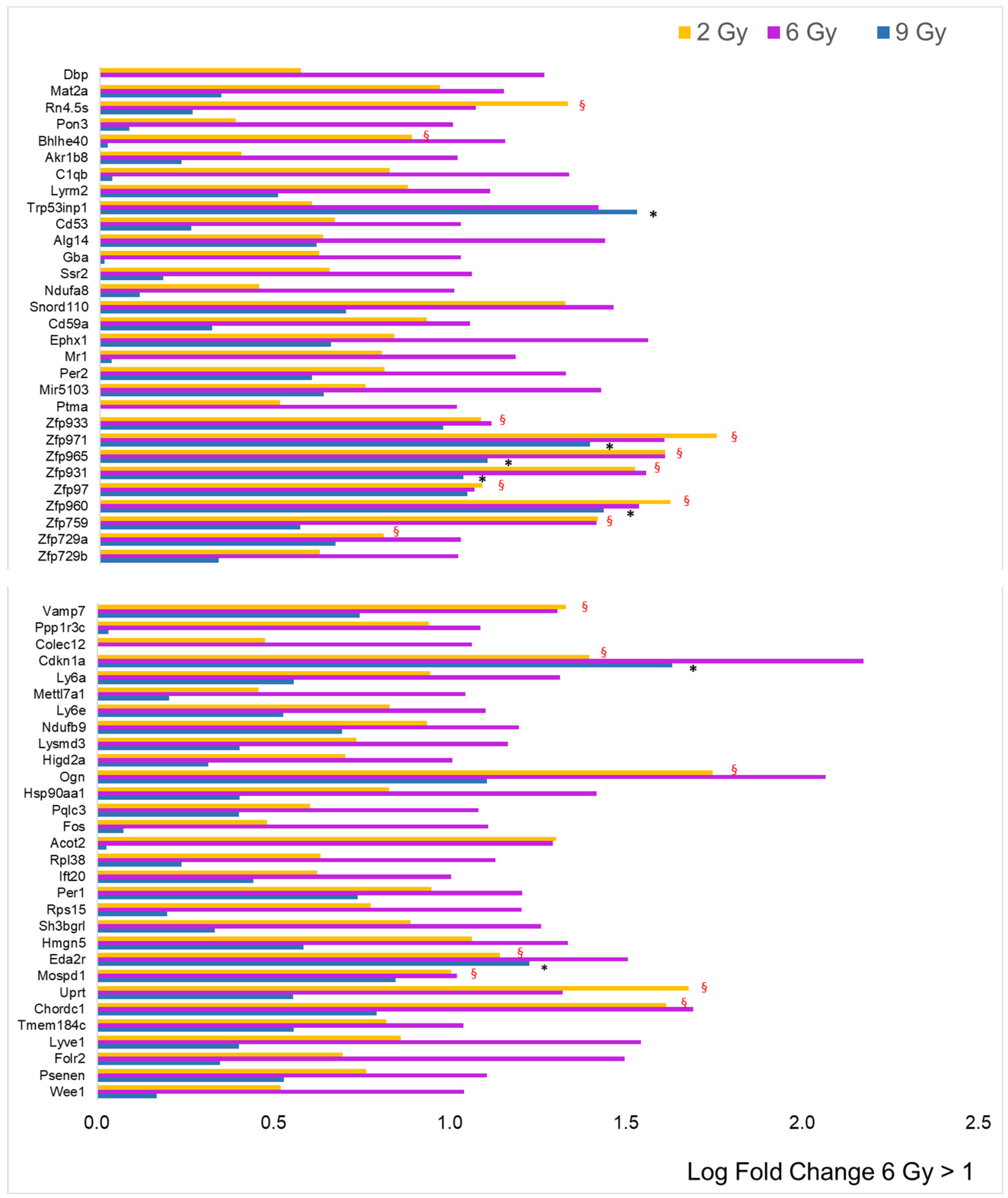
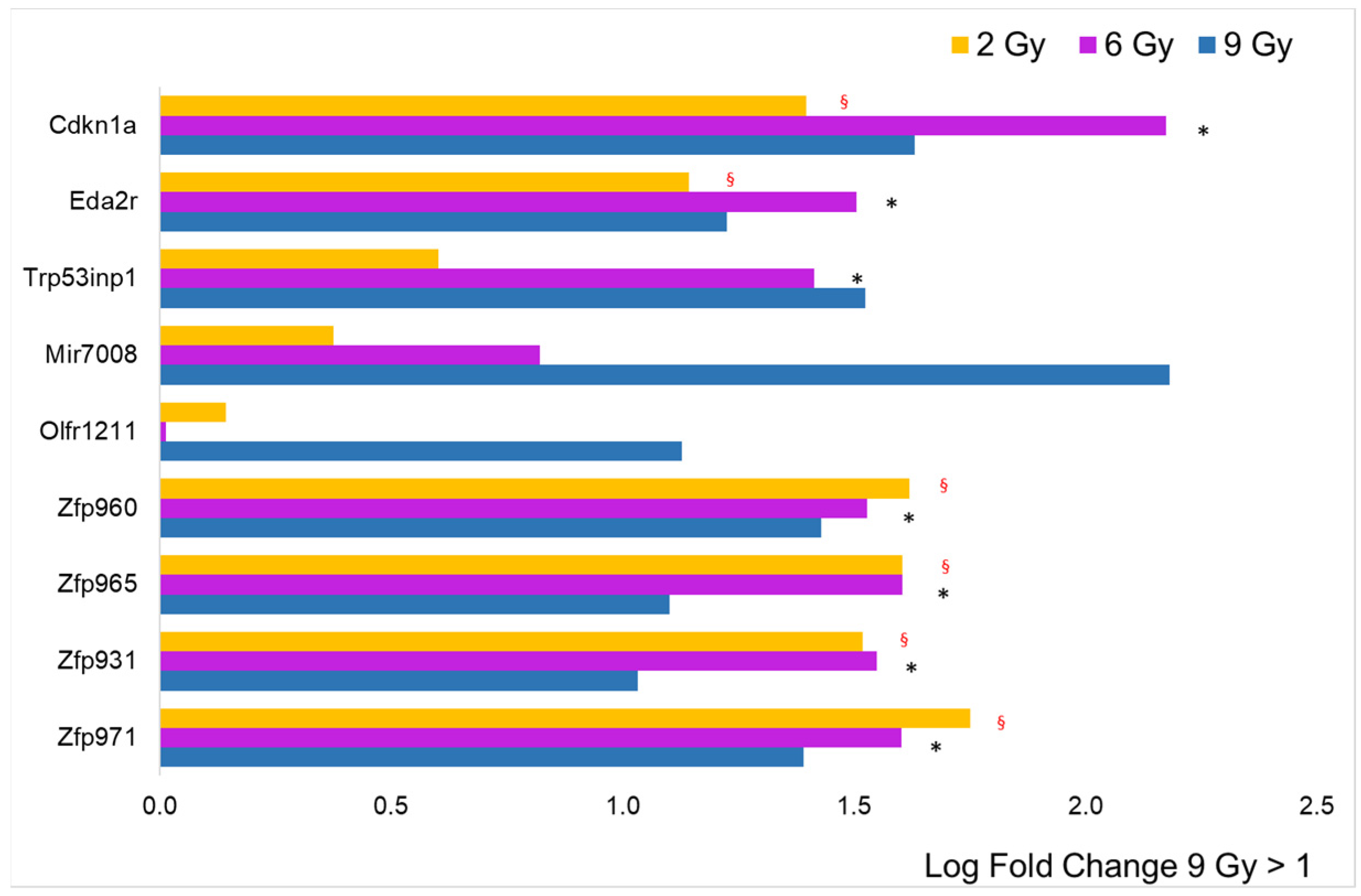
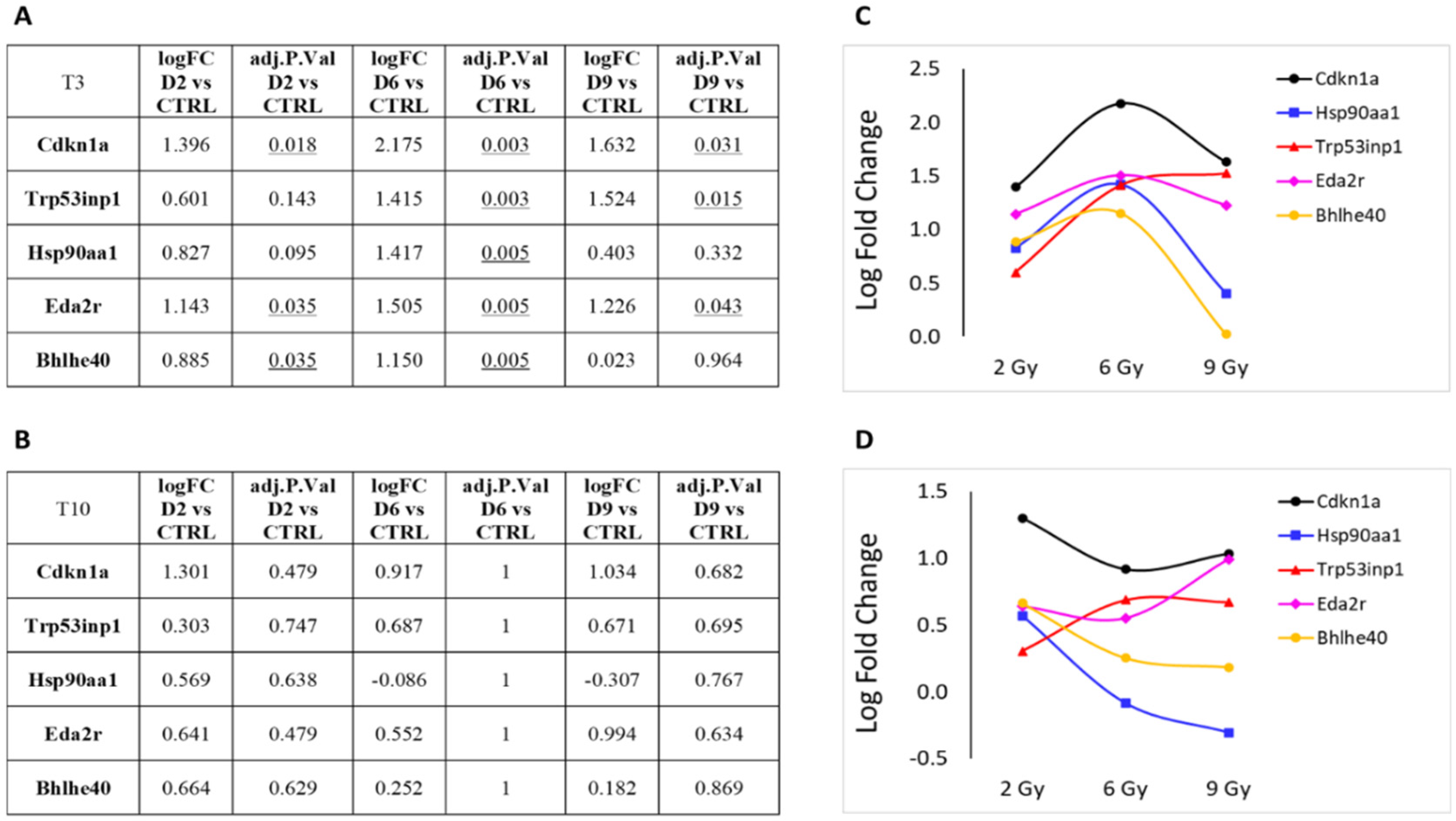
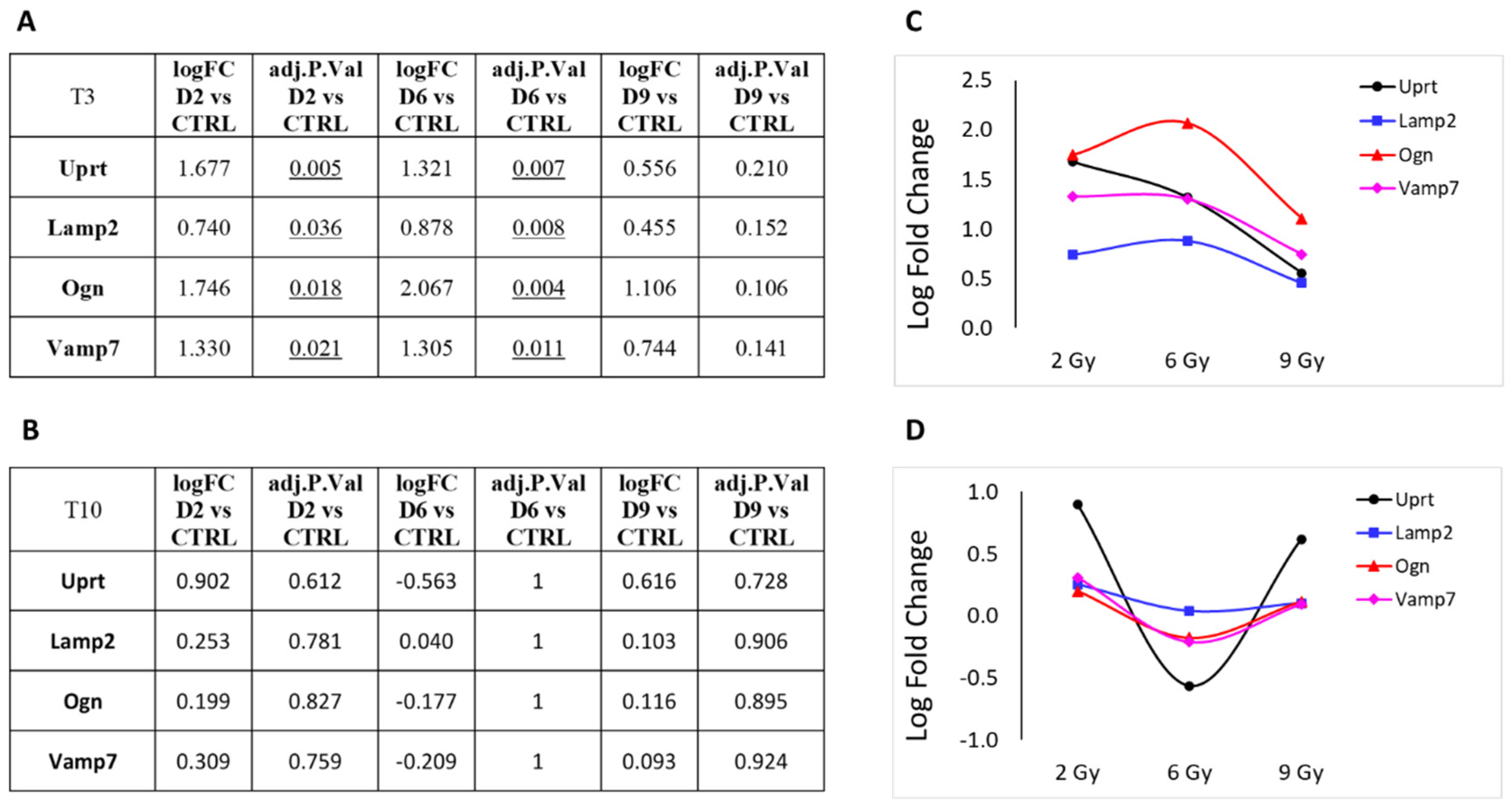
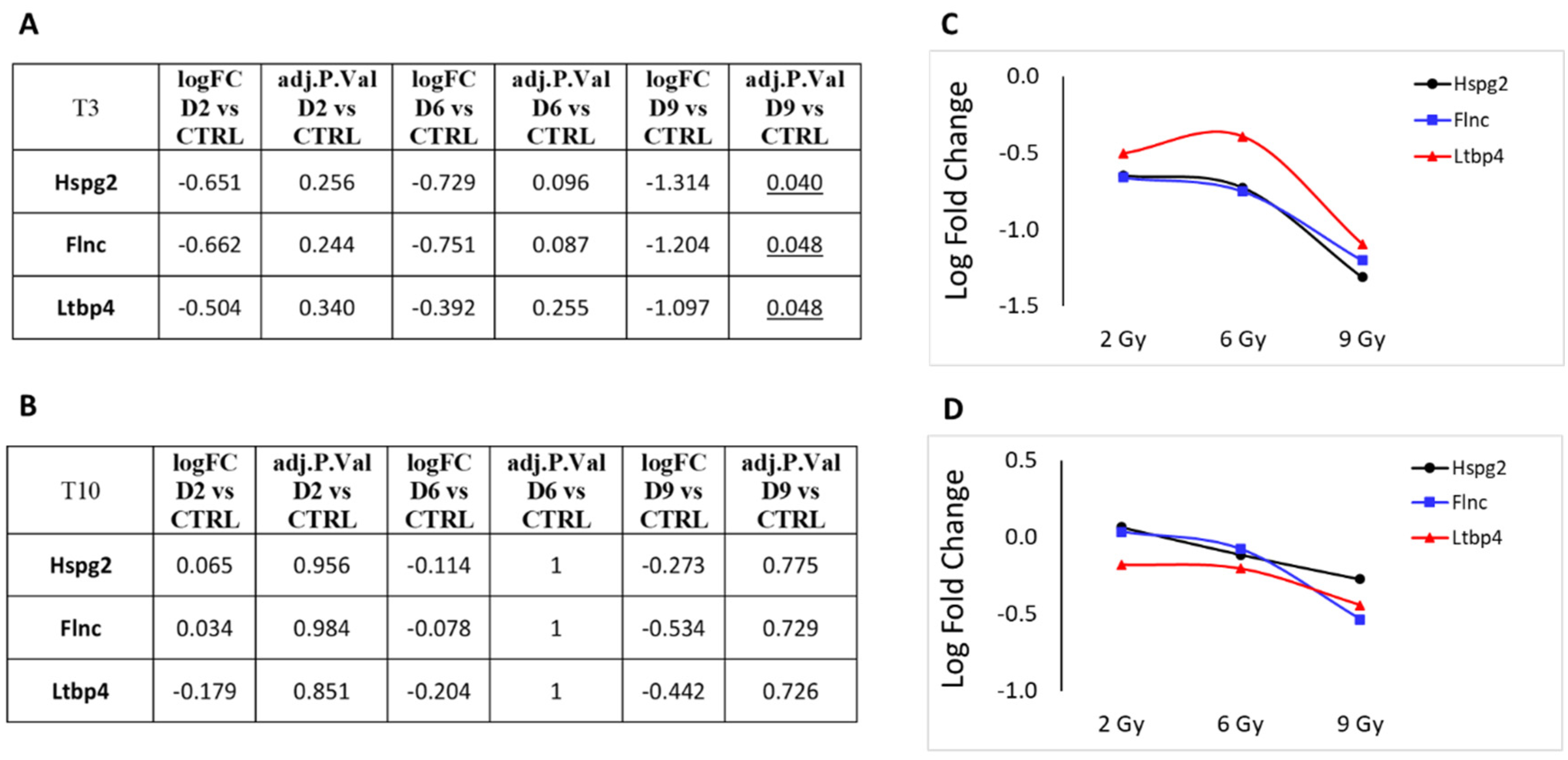
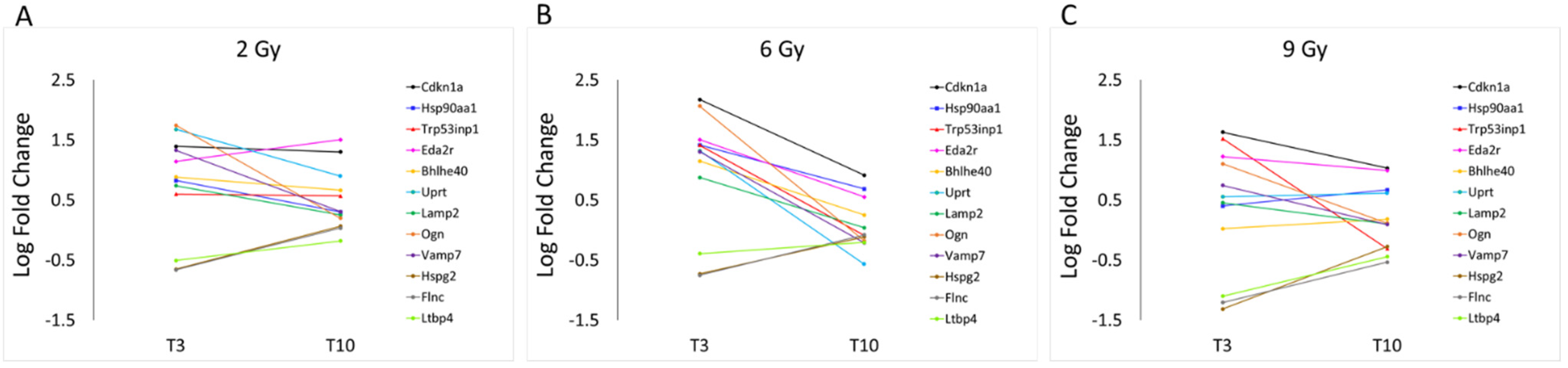
3.1. Differential Expression Analysis
3.2. Focus on Protein-Coding Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Menezes, K.M.; Wang, H.; Hada, M.; Saganti, P.B. Radiation Matters of the Heart: A Mini Review. Front. Cardiovasc. Med. 2018, 5, 83. [Google Scholar] [CrossRef] [PubMed]
- Taylor, C.W.; Nisbet, A.; McGale, P.; Darby, S.C. Cardiac Exposures in Breast Cancer Radiotherapy: 1950s–1990s. Int. J. Radiat. Oncol. 2007, 69, 1484–1495. [Google Scholar] [CrossRef] [PubMed]
- EBCTCG; Darby, S.; McGale, P.; Correa, C.; Taylor, C.; Arriagada, R.; Clarke, M.; Cutter, D.; Davies, C.; Ewertz, M.; et al. Effect of radiotherapy after breast-conserving surgery on 10-year recurrence and 15-year breast cancer death: Meta-analysis of individual patient data for 10,801 women in 17 randomised trials. Lancet 2011, 378, 1707–1716. [Google Scholar] [PubMed]
- Bartelink, H.; Maingon, P.; Poortmans, P.; Weltens, C.; Fourquet, A.; Jager, J.; Schinagl, D.; Oei, B.; Rodenhuis, C.; Horiot, J.-C.; et al. Whole-breast irradiation with or without a boost for patients treated with breast-conserving surgery for early breast cancer: 20-year follow-up of a randomised phase 3 trial. Lancet Oncol. 2015, 16, 47–56. [Google Scholar] [CrossRef]
- Chowdhary, M.; Lee, A.; Gao, S.; Wang, D.; Barry, P.N.; Diaz, R.; Bagadiya, N.R.; Park, H.S.; Yu, J.B.; Wilson, L.D.; et al. Is Proton Therapy a “Pro” for Breast Cancer? A Comparison of Proton vs. Non-proton Radiotherapy Using the National Cancer Database. Front. Oncol. 2019, 8, 678. [Google Scholar] [CrossRef] [PubMed]
- Taylor, C.W.; Wang, Z.; Macaulay, E.; Jagsi, R.; Duane, F.; Darby, S.C. Exposure of the Heart in Breast Cancer Radiation Therapy: A Systematic Review of Heart Doses Published During 2003 to 2013. Int. J. Radiat. Oncol. 2015, 93, 845–853. [Google Scholar] [CrossRef] [PubMed]
- Mast, M.E.; Vredeveld, E.J.; Credoe, H.M.; van Egmond, J.; Heijenbrok, M.W.; Hug, E.B.; Kalk, P.; van Kempen-Harteveld, L.M.L.; Korevaar, E.W.; van der Laan, H.P.; et al. Whole breast proton irradiation for maximal reduction of heart dose in breast cancer patients. Breast Cancer Res. Treat. 2014, 148, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Tommasino, F.; Durante, M.; D’Avino, V.; Liuzzi, R.; Conson, M.; Farace, P.; Palma, G.; Schwarz, M.; Cella, L.; Pacelli, R. Model-based approach for quantitative estimates of skin, heart, and lung toxicity risk for left-side photon and proton irradiation after breast-conserving surgery. Acta Oncol. 2017, 56, 730–736. [Google Scholar] [CrossRef] [PubMed]
- Bravatà, V.; Cammarata, F.P.; Minafra, L.; Pisciotta, P.; Scazzone, C.; Manti, L.; Savoca, G.; Petringa, G.; Cirrone, G.A.P.; Cuttone, G.; et al. Proton-irradiated breast cells: Molecular points of view. J. Radiat. Res. 2019, 60, 451–465. [Google Scholar] [CrossRef] [PubMed]
- Ricciotti, E.; Sarantopoulou, D.; Grant, G.R.; Sanzari, J.K.; Krigsfeld, G.S.; Kiliti, A.J.; Kennedy, A.R.; Grosser, T. Distinct vascular genomic response of proton and gamma radiation-A pilot investigation. PLoS ONE 2019, 14, e0207503. [Google Scholar] [CrossRef] [PubMed]
- Garikipati, V.N.S.; Arakelyan, A.; Blakely, E.A.; Chang, P.Y.; Truongcao, M.M.; Cimini, M.; Malaredy, V.; Bajpai, A.; Addya, S.; Bisserier, M.; et al. Long-Term Effects of Very Low Dose Particle Radiation on Gene Expression in the Heart: Degenerative Disease Risks. Cells 2021, 10, 387. [Google Scholar] [CrossRef] [PubMed]
- Laschinsky, L.; Baumann, M.; Beyreuther, E.; Enghardt, W.; Kaluza, M.; Karsch, L.; Lessmann, E.; Naumburger, D.; Nicolai, M.; Richter, C.; et al. Radiobiological Effectiveness of Laser Accelerated Electrons in Comparison to Electron Beams from a Conventional Linear Accelerator. J. Radiat. Res. 2012, 53, 395–403. [Google Scholar] [CrossRef] [PubMed]
- Willers, H.; Allen, A.; Grosshans, D.; McMahon, S.J.; von Neubeck, C.; Wiese, C.; Vikram, B. Toward A variable RBE for proton beam therapy. Radiother. Oncol. 2018, 128, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Workman, P.; Aboagye, E.O.; Balkwill, F.; Balmain, A.; Bruder, G.; Chaplin, D.J.; Double, J.A.; Everitt, J.; Farningham, D.A.H.; Glennie, M.J.; et al. Guidelines for the welfare and use of animals in cancer research. Br. J. Cancer 2010, 102, 1555–1577. [Google Scholar] [CrossRef] [PubMed]
- Bravatà, V.; Cammarata, F.P.; Minafra, L.; Musso, R.; Pucci, G.; Spada, M.; Fazio, I.; Russo, G.; Forte, G.I. Gene Expression Profiles Induced by High-dose Ionizing Radiation in MDA-MB-231 Triple-negative Breast Cancer Cell Line. Cancer Genom. Proteom. 2019, 16, 257–266. [Google Scholar] [CrossRef]
- Cammarata, F.P.; Forte, G.I.; Broggi, G.; Bravatà, V.; Minafra, L.; Pisciotta, P.; Calvaruso, M.; Tringali, R.; Tomasello, B.; Torrisi, F.; et al. Molecular Investigation on a Triple Negative Breast Cancer Xenograft Model Exposed to Proton Beams. Int. J. Mol. Sci. 2020, 21, 6337. [Google Scholar] [CrossRef] [PubMed]
- Simonetti, G.; Padella, A.; do Valle, I.F.; Fontana, M.C.; Fonzi, E.; Bruno, S.; Baldazzi, C.; Guadagnuolo, V.; Manfrini, M.; Ferrari, A.; et al. Aneuploid acute myeloid leukemia exhibits a signature of genomic alterations in the cell cycle and protein degradation machinery. Cancer 2019, 125, 712–725. [Google Scholar] [CrossRef] [PubMed]
- Irizarry, R.A.; Bolstad, B.M.; Collin, F.; Cope, L.M.; Hobbs, B.; Speed, T.P. Summaries of Affymetrix GeneChip probe level data. Nucleic Acids Res. 2003, 31, e15. [Google Scholar] [CrossRef] [PubMed]
- Klaus, B.; Reisenauer, S. An end to end workflow for differential gene expression using Affymetrix microarrays. F1000Research 2016, 5, 1384. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef]
- Raudvere, U.; Kolberg, L.; Kuzmin, I.; Arak, T.; Adler, P.; Peterson, H.; Vilo, J. g:Profiler: A web server for functional enrichment analysis and conversions of gene lists (2019 update). Nucleic Acids Res. 2019, 47, W191–W198. [Google Scholar] [CrossRef] [PubMed]
- Urban, Z.; Hucthagowder, V.; Schürmann, N.; Todorovic, V.; Zilberberg, L.; Choi, J.; Sens, C.; Brown, C.W.; Clark, R.D.; Holland, K.E.; et al. Mutations in LTBP4 Cause a Syndrome of Impaired Pulmonary, Gastrointestinal, Genitourinary, Musculoskeletal, and Dermal Development. Am. J. Hum. Genet. 2009, 85, 593–605. [Google Scholar] [CrossRef] [PubMed]
- Deckx, S.; Heymans, S.; Papageorgiou, A. The diverse functions of osteoglycin: A deceitful dwarf, or a master regulator of disease? FASEB J. 2016, 30, 2651–2661. [Google Scholar] [CrossRef]
- Deckx, S.; Heggermont, W.; Carai, P.; Rienks, M.; Dresselaers, T.; Himmelreich, U.; van Leeuwen, R.; Lommen, W.; van der Velden, J.; Gonzalez, A.; et al. Osteoglycin prevents the development of age-related diastolic dysfunction during pressure overload by reducing cardiac fibrosis and inflammation. Matrix Biol. 2018, 66, 110–124. [Google Scholar] [CrossRef] [PubMed]
- Laity, J.H.; Lee, B.M.; Wright, P.E. Zinc finger proteins: New insights into structural and functional diversity. Curr. Opin. Struct. Biol. 2001, 11, 39–46. [Google Scholar] [CrossRef]
- Yang, H.; Lu, Y.; Zheng, Y.; Yu, X.; Xia, X.; He, X.; Feng, W.; Xing, L.; Ling, Z. shRNA-mediated silencing of ZFX attenuated the proliferation of breast cancer cells. Cancer Chemother. Pharmacol. 2014, 73, 569–576. [Google Scholar] [CrossRef] [PubMed]
- Machnik, M.; Cylwa, R.; Kiełczewski, K.; Biecek, P.; Liloglou, T.; Mackiewicz, A.; Oleksiewicz, U. The expression signature of cancer-associated KRAB-ZNF factors identified in TCGA pan-cancer transcriptomic data. Mol. Oncol. 2019, 13, 701–724. [Google Scholar] [CrossRef]
- Sobocińska, J.; Molenda, S.; Machnik, M.; Oleksiewicz, U. KRAB-ZFP Transcriptional Regulators Acting as Oncogenes and Tumor Suppressors: An Overview. Int. J. Mol. Sci. 2021, 22, 2212. [Google Scholar] [CrossRef] [PubMed]
- Galeaz, C.; Totis, C.; Bisio, A. Radiation Resistance: A Matter of Transcription Factors. Front. Oncol. 2021, 11, 662840. [Google Scholar] [CrossRef] [PubMed]
- Wade Harper, J. The p21 Cdk-interacting protein Cip1 is a potent inhibitor of G1 cyclin-dependent kinases. Cell 1993, 75, 805–816. [Google Scholar] [CrossRef] [PubMed]
- Antoccia, A.; Sgura, A.; Berardinelli, F.; Cavinato, M.; Cherubini, R.; Gerardi, S.; Tanzarella, C. Cell Cycle Perturbations and Genotoxic Effects in Human Primary Fibroblasts Induced by Low-energy Protons and X/γ-rays. J. Radiat. Res. 2009, 50, 457–468. [Google Scholar] [CrossRef] [PubMed]
- Licursi, V.; Wang, W.; Di Nisio, E.; Cammarata, F.P.; Acquaviva, R.; Russo, G.; Manti, L.; Cestelli Guidi, M.; Fratini, E.; Kamel, G.; et al. Transcriptional modulations induced by proton irradiation in mice skin in function of adsorbed dose and distance. J. Radiat. Res. Appl. Sci. 2021, 14, 260–270. [Google Scholar] [CrossRef]
- Sertorio, M.; Nowrouzi, A.; Akbarpour, M.; Chetal, K.; Salomonis, N.; Brons, S.; Mascia, A.; Ionascu, D.; McCauley, S.; Kupneski, T.; et al. Differential transcriptome response to proton versus X-ray radiation reveals novel candidate targets for combinatorial PT therapy in lymphoma. Radiother. Oncol. 2021, 155, 293–303. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, S.; Bassler, N.; Grzanka, L.; Swakon, J.; Olko, P.; Andreassen, C.N.; Overgaard, J.; Alsner, J.; Sørensen, B.S. Differential gene expression in primary fibroblasts induced by proton and cobalt-60 beam irradiation. Acta Oncol. 2017, 56, 1406–1412. [Google Scholar] [CrossRef]
- Paganetti, H.; Niemierko, A.; Ancukiewicz, M.; Gerweck, L.E.; Goitein, M.; Loeffler, J.S.; Suit, H.D. Relative biological effectiveness (RBE) values for proton beam therapy. Int. J. Radiat. Oncol. 2002, 53, 407–421. [Google Scholar] [CrossRef] [PubMed]
- United Nations Scientific Committee on the Effects of Atomic Radiation. Biological mechanisms relevant to dose–response relationships. In Sources, Effects and Risks of Ionizing Radiation; Scientific Annex C: Biological mechanisms Relevant for the Inference of Cancer Risks from Low-Dose and Low-Dose-Rate Radiation; United Nations: New York, NY, USA, 2022; Volume III, pp. 12–137. [Google Scholar] [CrossRef]
- Giantsoudi, D.; Adams, J.; MacDonald, S.M.; Paganetti, H. Proton Treatment Techniques for Posterior Fossa Tumors: Consequences for Linear Energy Transfer and Dose-Volume Parameters for the Brainstem and Organs at Risk. Int. J. Radiat. Oncol. 2017, 97, 401–410. [Google Scholar] [CrossRef]
- McNamara, A.; Willers, H.; Paganetti, H. Modelling variable proton relative biological effectiveness for treatment planning. Br. J. Radiol. 2020, 93, 20190334. [Google Scholar] [CrossRef] [PubMed]
- Newhauser, W.D.; Zhang, R. The physics of proton therapy. Phys. Med. Biol. 2015, 60, R155–R209. [Google Scholar] [CrossRef]
- Friedrich, T. Proton RBE dependence on dose in the setting of hypofractionation. Br. J. Radiol. 2020, 93, 20190291. [Google Scholar] [CrossRef] [PubMed]
- Girdhani, S.; Sachs, R.; Hlatky, L. Biological Effects of Proton Radiation: What We Know and Don’t Know. Radiat. Res. 2013, 179, 257–272. [Google Scholar] [CrossRef]
- Vitti, E.T.; Parsons, J.L. The Radiobiological Effects of Proton Beam Therapy: Impact on DNA Damage and Repair. Cancers 2019, 11, 946. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | Term_Name | Term_ID | Adjusted_p_Value | Intersections |
|---|---|---|---|---|
| GO:MF | RNA polymerase II transcription regulatory region sequence-specific DNA binding | GO:0000977 | 0.002695 | GM14295, ZFP970, GM14403, GM14322, ZFP971, GM14393, GM14399, GM14325, GM14326, ZFP931, ZFP965, GM14305, ZFP933, ZFP759, ZFP960, ZFP97, GM2026, ZFP938, ZFP935, ZFP729A, BHLHE40 |
| GO:MF | DNA-binding transcription factor activity | GO:0003700 | 0.000000 | GM14295, ZFP970, GM14403, GM14322, ZFP971, GM14393, GM14399, GM14325, GM14326, ZFP931, ZFP965, GM14305, ZFP933, ZFP759, ZFP960, ZFP97, GM2026, ZFP938, ZFP935, ZFP729A, BHLHE40 |
| GO:BP | regulation of transcription by RNA polymerase II | GO:0006357 | 0.001250 | GM14295, ZFP970, GM14403, GM14322, ZFP971 GM14393, GM14399, GM14325, GM14326, ZFP931, ZFP965, GM14305, ZFP933, MOSPD1, ZFP759, ZFP960, ZFP97, GM2026, ZFP938, ZFP935, ZFP729A, BHLHE40 |
| REAC | Gene expression (Transcription) | REAC:R-MMU-74160 | 0.000409 | GM14322, ZFP971, GM14325, ZFP931, CDKN1A, GM2026, ZFP938, GTF3C6, ZFP729A |
| (a) | ||||
|---|---|---|---|---|
| Source | Term_Name | Term_ID | Adjusted_p_Value | Intersections |
| GO:MF | RNA polymerase II transcription regulatory region sequence-specific DNA binding | GO:0000977 | 0.000569582 | GM14393, GM14399, GM14325, GM14326, ZFP931, ZFP965, GM14305, ZFP933, BHLHE40, DBP, PER1, FOS, ZFP759, ZFP729A, ZFP729B, ZFP960, ZFP97, MAX, XBP1, ZFP955B, ZFP760, ZFP953, ZFP935, ZFP72, ZFP712, ZFP273, ZFP938, ZFP433, FP930, ZFP975, ZFP84 |
| GO:MF | Transcription cis-regulatory region binding | GO:0000976 | 0.00256767 | GM14399, GM14326, GM14325, ZFP931, ZFP965, GM14305, ZFP933, BHLHE40, DBP, PER1, FOS, ZFP759, ZFP729A, ZFP729B, ZFP960, ZFP97, MAX, XBP1, ZFP955B, ZFP760, ZFP953, ZFP935, ZFP72, ZFP712, ZFP273, ZFP938, ZFP433, ZFP930, ZFP975, ZFP84, M14393 |
| GO:BP | Cellular amide metabolic process | GO:0043603 | 0.00049512 | RPS15, PER1, RPL38, ACOT2, ACOT10, RPL17, IMPACT, RPL15, PDHB, ABHD4, RPS29, DLD, HMGN5, EIF2S3X, RBM3, GSTA4, ABCE1, PSENEN, EIF3K, NGRN, MCEE, EIF4E3, GSTK1, RPL29, MRPS17, SCP2, GSTM4, GBA, EIF2A |
| GO:BP | Establishment of protein localization | GO:0045184 | 0.019966747 | HSP90AA1, CDKN1A, PPP1R3C, ATAD1, ANXA1, Y IPF5, CRIPT, APOD, SNAP29, STK3, ENY 2, BTF3, BCAP29, VPS25, IFT20, XBP1, MDM2, PEX3, PTTG1IP, RAB9, SNX12, LAMP2, EMD, TIMM8B, VPS35, FOLR2, RAB6A, GOLT1B, EXOC4, CHMP5, SSR3, UFM1, SEC62 |
| GO:BP | Oxidative phosphorylation | GO:0006119 | 0.021330016 | COX7A2L, NDUFB9, COX7C, UQCRB, DLD, SDHD, RHOA, STOML2 |
| KEGG | Oxidative phosphorylation | KEGG:00190 | 0.039889377 | COX7A2L, ATP6V0E, NDUFB9, COX7C, UQCRB, SDHD, ATP6V0E2, NDUFC1 |
| KEGG | Drug metabolism—cytochrome P450 | KEGG:00982 | 0.043325466 | MAOB, GSTA4, GSTK1, UGT2B38, UGT2B5, GSTM4 |
| (b) | ||||
| Source | Term_Name | Term_ID | Adjusted_p_Value | Intersections |
| GO:BP | Cellular response to radiation | GO:0071478 | 0.008787 | SWI5, MTCH2, HSPA5, IFI207, COPS9 |
| GO:CC | Myelin sheath | GO:0043209 | 0.037942 | TUBB4B, ATP5C1, CD59A, HSPA5 |
| KEGG | Chemical carcinogenesis—reactive oxygen species | KEGG:05208 | 0.033798 | NFE2L2, NDUFA8, ATP5C1, EPHX1 |
| Source | Term_Name | Term_ID | Adjusted_p_Value | Intersections |
|---|---|---|---|---|
| GO:MF | DNA-binding transcription factor activity, RNA polymerase II-specific | GO:0000981 | 0.0008 | ZFP971, GM14393, GM14325, ZFP931, ZFP965, GM14305, ZFP960 |
| GO:MF | DNA-binding transcription factor activity | GO:0003700 | 0.0011 | ZFP971, GM14393, GM14325, ZFP931, ZFP965, GM14305, ZFP960 |
| GO:MF | Transcription regulator activity | GO:0140110 | 0.0071 | ZFP971, GM14393, GM14325, ZFP931, ZFP965, GM14305, ZFP960 |
| KEGG | Proteoglycans in cancer | KEGG:05205 | 0.0312 | HSPG2, FLNC, CDKN1A |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sala, C.; Tarozzi, M.; Simonetti, G.; Pazzaglia, M.; Cammarata, F.P.; Russo, G.; Acquaviva, R.; Cirrone, G.A.P.; Petringa, G.; Catalano, R.; et al. Impact on the Transcriptome of Proton Beam Irradiation Targeted at Healthy Cardiac Tissue of Mice. Cancers 2024, 16, 1471. https://doi.org/10.3390/cancers16081471
Sala C, Tarozzi M, Simonetti G, Pazzaglia M, Cammarata FP, Russo G, Acquaviva R, Cirrone GAP, Petringa G, Catalano R, et al. Impact on the Transcriptome of Proton Beam Irradiation Targeted at Healthy Cardiac Tissue of Mice. Cancers. 2024; 16(8):1471. https://doi.org/10.3390/cancers16081471
Chicago/Turabian StyleSala, Claudia, Martina Tarozzi, Giorgia Simonetti, Martina Pazzaglia, Francesco Paolo Cammarata, Giorgio Russo, Rosaria Acquaviva, Giuseppe Antonio Pablo Cirrone, Giada Petringa, Roberto Catalano, and et al. 2024. "Impact on the Transcriptome of Proton Beam Irradiation Targeted at Healthy Cardiac Tissue of Mice" Cancers 16, no. 8: 1471. https://doi.org/10.3390/cancers16081471
APA StyleSala, C., Tarozzi, M., Simonetti, G., Pazzaglia, M., Cammarata, F. P., Russo, G., Acquaviva, R., Cirrone, G. A. P., Petringa, G., Catalano, R., Elia, V. C., Fede, F., Manti, L., Castellani, G., Remondini, D., & Zironi, I. (2024). Impact on the Transcriptome of Proton Beam Irradiation Targeted at Healthy Cardiac Tissue of Mice. Cancers, 16(8), 1471. https://doi.org/10.3390/cancers16081471