
Transforming Growth Factor Beta 2 (TGFB2) mRNA Levels, in Conjunction with Interferon-Gamma Receptor Activation of Interferon Regulatory Factor 5 (IRF5) and Expression of CD276/B7-H3, Are Therapeutically Targetable Negative Prognostic Markers in Low-Grade Gliomas

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Comparing mRNA Expression in LGG Tumors and Normal Brain Samples
2.2. Kaplan–Meier OS Analysis Comparing Subsets of LGG Patients Stratified According to mRNA Expression Levels of TGFB2 and Target Genes in the TME
2.3. Comparing the Independent Effects on Hazard Ratios (HR) of High Levels of TGFB2 mRNA Levels Controlling for Target Gene Expression Levels, Age, and Interaction between TGFB2 and the Target Gene
2.4. Correlation of TGFB2 and Immune Cell Infiltration
2.5. OS Impact in LGG Patients upon Demethylation of the Target Genes
3. Results
3.1. LGG Tumors Exhibit Augmented mRNA Expression Levels for TGFB Ligands, Interferon-Gamma Receptors, Downstream Signaling Molecules, and Macrophage Markers Compared to Normal Brain Samples
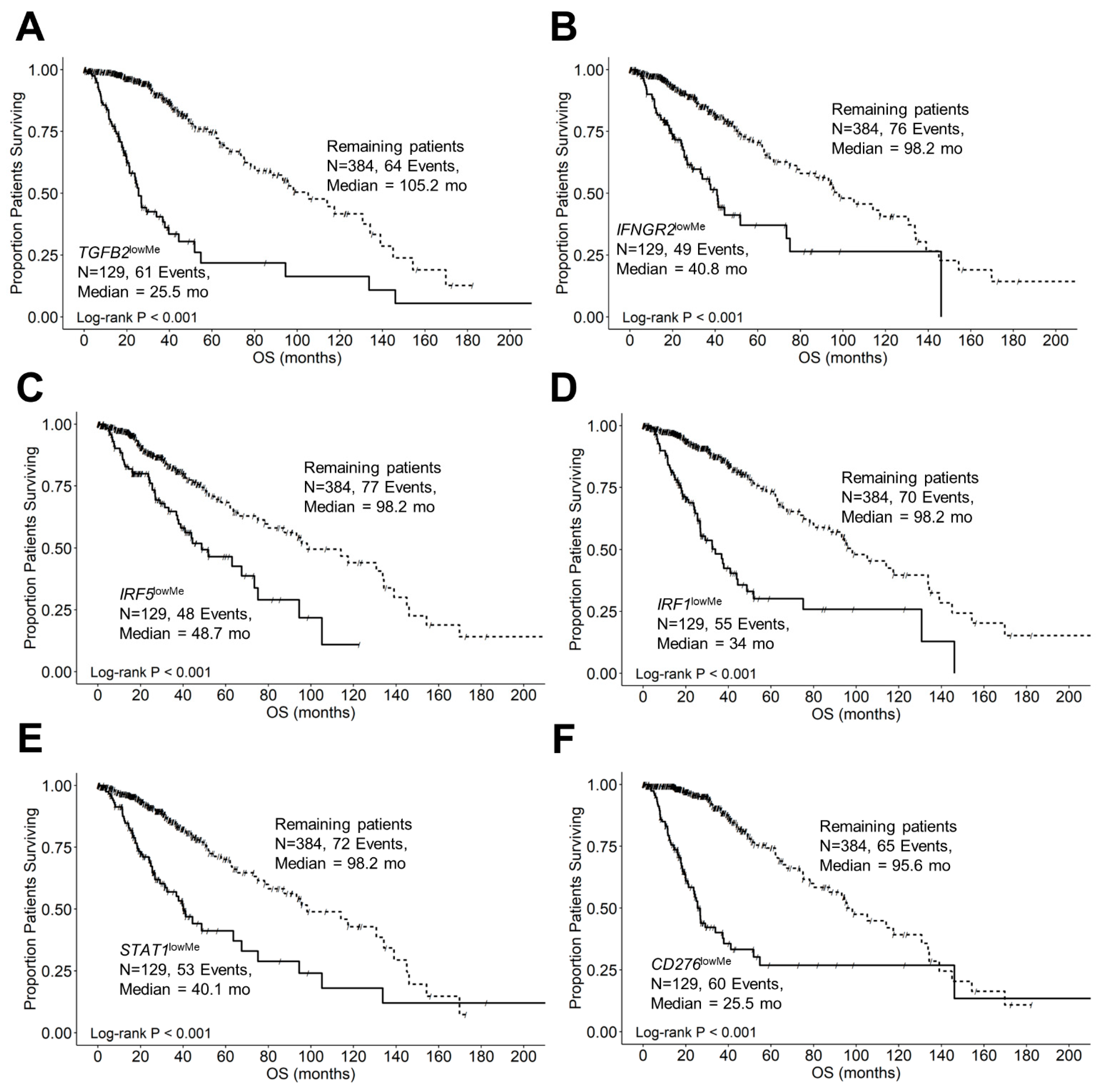
3.2. Expression Levels of TGFB2, IFNGR2, CD276, and MSR1/CD204 mRNA Were Treatment-Independent Negative Prognostic Markers in LGG Tumors
3.3. TGFB2 Is a Significant Negative Prognostic Indicator for OS in LGG Patients Independent of IDH Mutational Status
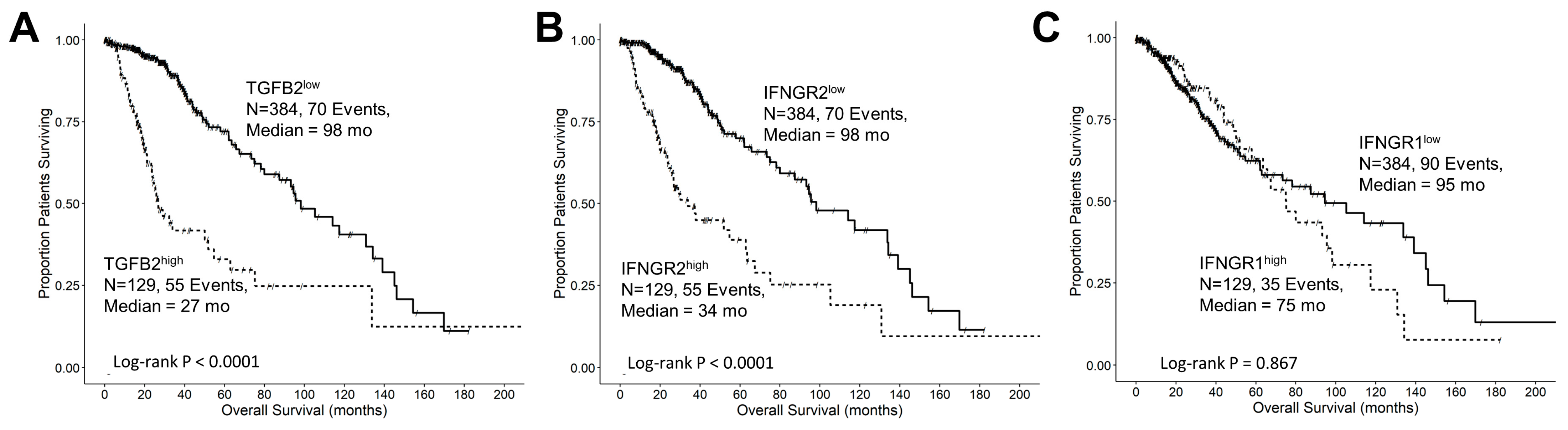
3.4. TGFB2 and IFNGR2 Are Significant Negative Prognostic Indicators for OS in LGG Patients
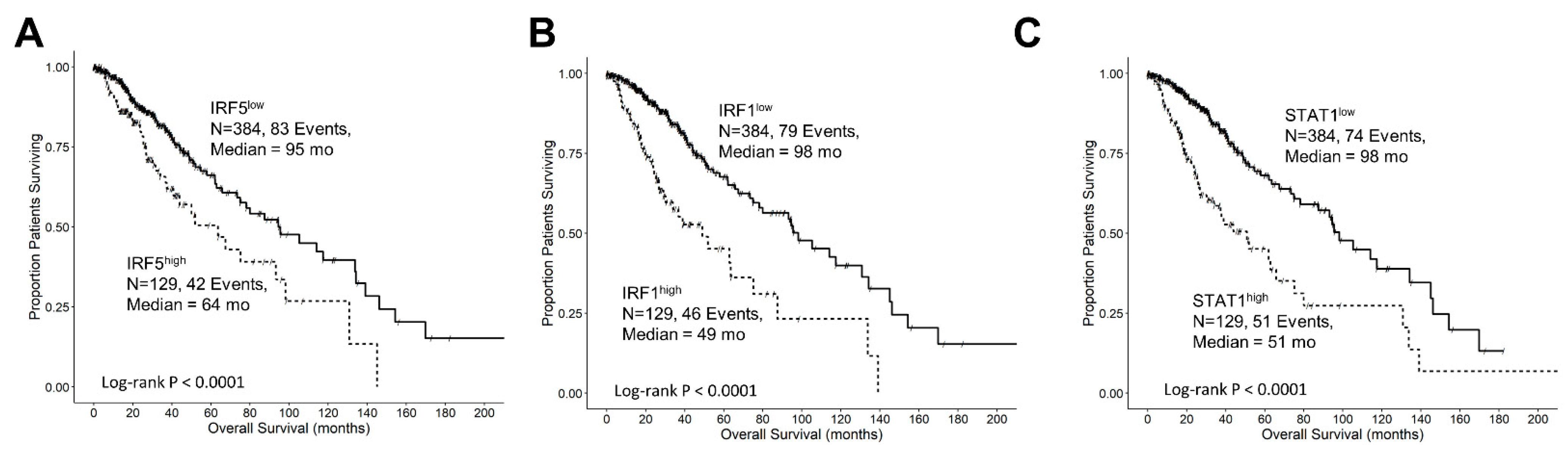
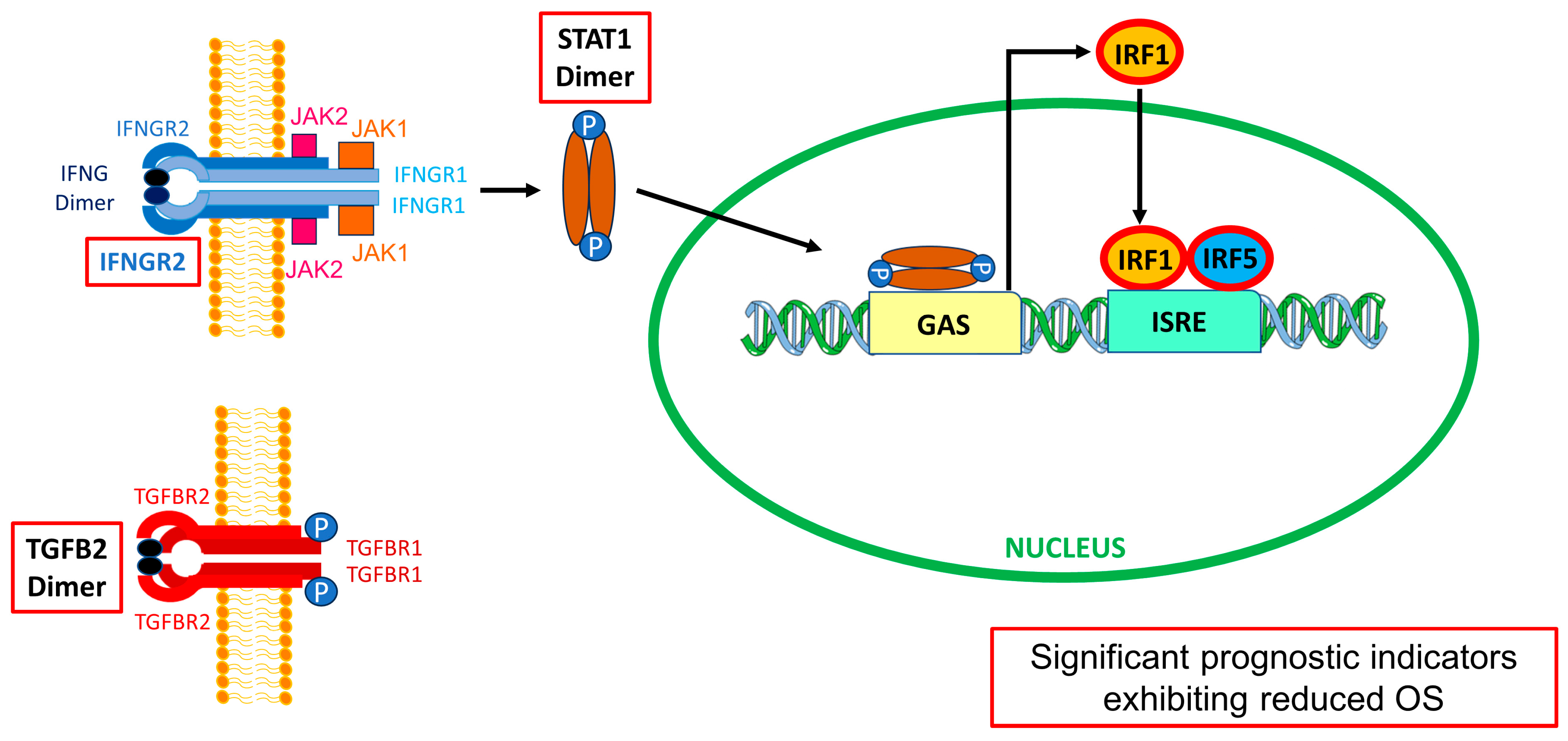
3.5. Interferon-Gamma Receptor-Activated Downstream Signaling Molecules STAT1, IRF1, IRF5 Displayed Significant Negative Prognosis for OS in LGG Patients
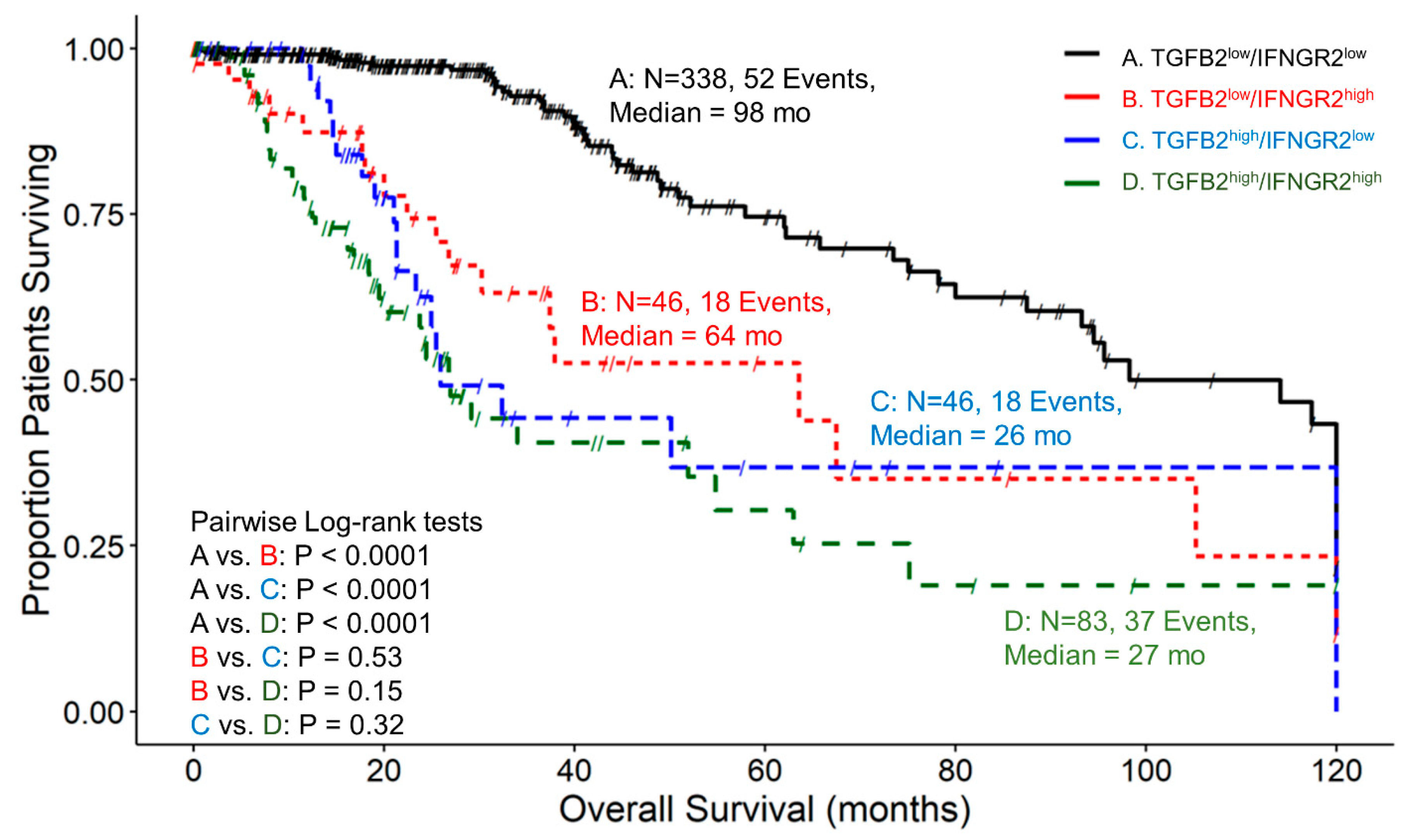
3.6. The Prognostic Impact of TGFB2 mRNA Levels in Tumors of LGG Patients Is Dependent on IFNGR2 mRNA Expression
3.7. The Prognostic Effect of TGFB2 mRNA Levels in Tumors of LGG Patients Is Independent of the Levels of IRF5
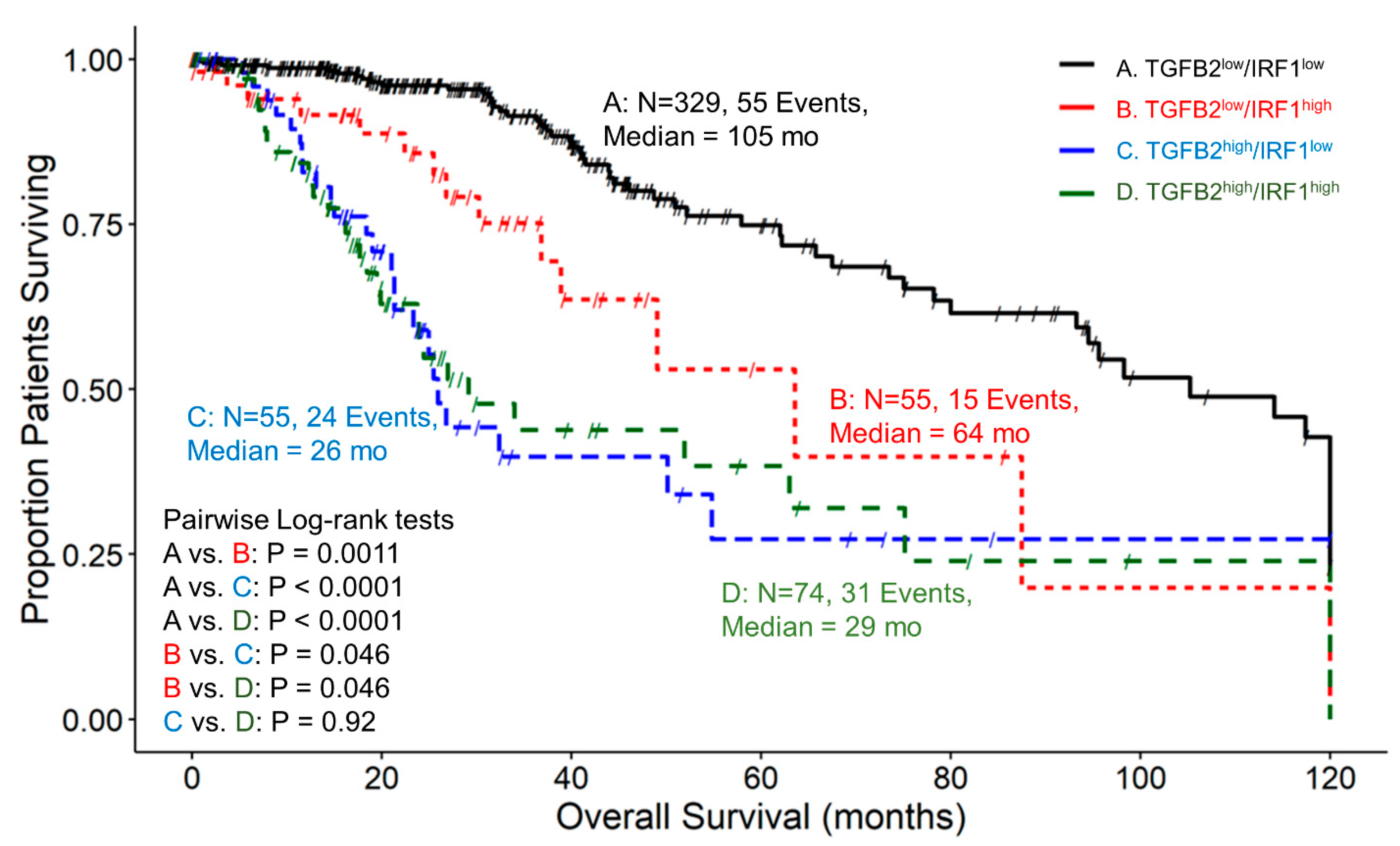
3.8. The Prognostic Impact of TGFB2 mRNA Levels in Tumors of LGG Patients Is Independent of the Levels of IRF1, While at TGFB2low, IRF1 Exhibits Worse OS
3.9. At Low Levels of STAT1, High TGFB2 mRNA Levels Exhibit Worse Survival Outcomes, and at Low Levels of TGFB2, STAT1 Is a Significant Negative Prognostic Indicator for OS in LGG Patients
3.10. Independent Impact on OS of High TGFB2, IFNGR2 mRNA, and Downstream Signaling Molecules IRF5, IRF1, and STAT1 Determined from Multivariate Cox Proportional Hazards Models
3.11. TGFB2 mRNA Levels Were Positively Correlated with M1 and M2 Macrophage Infiltration into LGG Tumors
3.12. Demethylation of Target Genes TGFB2, IFNGR2, IRF5, IRF1, STAT1, and CD276 Exhibited Significantly Shorter OS Times and Overrepresentation in IDH Wildtype for LGG Patients
4. Discussion
4.1. Differential Expression of Macrophage Markers, TGFB Ligands, Interferon-Gamma Receptor, and Signaling Molecules in LGG Tumors Compared to Normal Brain Tissue
4.2. TGFB2high and IFNGR2high Subsets of Patients Are Over-Represented in IDHwt LGG Patients
4.3. High Levels of TGFB2, IFNGR2, and Downstream Signaling Molecules Exhibited Worse OS Outcomes than Those of the Remaining Patients
4.4. Estimating the OS Impact of the Interaction of TGFB2 Levels and the Molecules Involved in Interferon-Gamma Receptor Activation Shows That High TGFB2 Levels Are Independent Prognostic Indicators
4.5. Combinations of High Levels of TGFB2 and IFNGR2 or the Downstream IRF1/IRF5 Signaling Molecules as Targets for LGG Patients
4.6. CD276/B7-H3 in Combination with TGFB2 Levels as Targets for Immune Therapy in Gliomas
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dixit, K.; Raizer, J. Newer Strategies for the Management of Low-Grade Gliomas. Oncology 2017, 31, 680–682, 684–685. [Google Scholar]
- Forst, D.A.; Nahed, B.V.; Loeffler, J.S.; Batchelor, T.T. Low-Grade Gliomas. Oncologist 2014, 19, 403–413. [Google Scholar] [CrossRef]
- Oberheim Bush, N.A.; Chang, S. Treatment Strategies for Low-Grade Glioma in Adults. J. Oncol. Pract. 2016, 12, 1235–1241. [Google Scholar] [CrossRef]
- Smith, J.S.; Chang, E.F.; Lamborn, K.R.; Chang, S.M.; Prados, M.D.; Cha, S.; Tihan, T.; Vandenberg, S.; McDermott, M.W.; Berger, M.S. Role of Extent of Resection in the Long-Term Outcome of Low-Grade Hemispheric Gliomas. J. Clin. Oncol. 2008, 26, 1338–1345. [Google Scholar] [CrossRef]
- Yan, Z.; Wang, J.; Dong, Q.; Zhu, L.; Lin, W.; Jiang, X. Predictors of Tumor Progression of Low-Grade Glioma in Adult Patients within 5 Years Follow-up after Surgery. Front. Surg. 2022, 9, 937556. [Google Scholar] [CrossRef]
- Yan, H.; Parsons, D.W.; Jin, G.; McLendon, R.; Rasheed, B.A.; Yuan, W.; Kos, I.; Batinic-Haberle, I.; Jones, S.; Riggins, G.J.; et al. IDH1 and IDH2 Mutations in Gliomas. N. Engl. J. Med. 2009, 360, 765–773. [Google Scholar] [CrossRef]
- Hartmann, C.; Hentschel, B.; Wick, W.; Capper, D.; Felsberg, J.; Simon, M.; Westphal, M.; Schackert, G.; Meyermann, R.; Pietsch, T.; et al. Patients with IDH1 Wild Type Anaplastic Astrocytomas Exhibit Worse Prognosis than IDH1-Mutated Glioblastomas, and IDH1 Mutation Status Accounts for the Unfavorable Prognostic Effect of Higher Age: Implications for Classification of Gliomas. Acta Neuropathol. 2010, 120, 707–718. [Google Scholar] [CrossRef]
- Murugan, A.K.; Alzahrani, A.S. Isocitrate Dehydrogenase IDH1 and IDH2 Mutations in Human Cancer: Prognostic Implications for Gliomas. Br. J. Biomed. Sci. 2022, 79, 10208. [Google Scholar] [CrossRef]
- Xia, L.; Wu, B.; Fu, Z.; Feng, F.; Qiao, E.; Li, Q.; Sun, C.; Ge, M. Prognostic Role of IDH Mutations in Gliomas: A Meta-Analysis of 55 Observational Studies. Oncotarget 2015, 6, 17354–17365. [Google Scholar] [CrossRef]
- Mellinghoff, I.K.; van den Bent, M.J.; Blumenthal, D.T.; Touat, M.; Peters, K.B.; Clarke, J.; Mendez, J.; Yust-Katz, S.; Welsh, L.; Mason, W.P.; et al. Vorasidenib in IDH1- or IDH2-Mutant Low-Grade Glioma. N. Engl. J. Med. 2023, 389, 589–601. [Google Scholar] [CrossRef]
- Tesileanu, C.M.S.; Dirven, L.; Wijnenga, M.M.J.; Koekkoek, J.A.F.; Vincent, A.J.P.E.; Dubbink, H.J.; Atmodimedjo, P.N.; Kros, J.M.; van Duinen, S.G.; Smits, M.; et al. Survival of Diffuse Astrocytic Glioma, IDH1/2 Wildtype, with Molecular Features of Glioblastoma, WHO Grade IV: A Confirmation of the CIMPACT-NOW Criteria. Neuro Oncol. 2020, 22, 515–523. [Google Scholar] [CrossRef]
- Brat, D.J.; Aldape, K.; Colman, H.; Holland, E.C.; Louis, D.N.; Jenkins, R.B.; Kleinschmidt-DeMasters, B.K.; Perry, A.; Reifenberger, G.; Stupp, R.; et al. cIMPACT-NOW Update 3: Recommended Diagnostic Criteria for “Diffuse Astrocytic Glioma, IDH-Wildtype, with Molecular Features of Glioblastoma, WHO Grade IV”. Acta Neuropathol. 2018, 136, 805–810. [Google Scholar] [CrossRef]
- Stichel, D.; Ebrahimi, A.; Reuss, D.; Schrimpf, D.; Ono, T.; Shirahata, M.; Reifenberger, G.; Weller, M.; Hänggi, D.; Wick, W.; et al. Distribution of EGFR Amplification, Combined Chromosome 7 Gain and Chromosome 10 Loss, and TERT Promoter Mutation in Brain Tumors and Their Potential for the Reclassification of IDHwt Astrocytoma to Glioblastoma. Acta Neuropathol. 2018, 136, 793–803. [Google Scholar] [CrossRef]
- Aoki, K.; Nakamura, H.; Suzuki, H.; Matsuo, K.; Kataoka, K.; Shimamura, T.; Motomura, K.; Ohka, F.; Shiina, S.; Yamamoto, T.; et al. Prognostic Relevance of Genetic Alterations in Diffuse Lower-Grade Gliomas. Neuro Oncol. 2018, 20, 66–77. [Google Scholar] [CrossRef]
- Keric, N.; Krenzlin, H.; Kalasauskas, D.; Freyschlag, C.F.; Schnell, O.; Misch, M.; von der Brelie, C.; Gempt, J.; Krigers, A.; Wagner, A.; et al. Treatment Outcome of IDH1/2 Wildtype CNS WHO Grade 4 Glioma Histologically Diagnosed as WHO Grade II or III Astrocytomas. J. Neurooncol. 2024. [Google Scholar] [CrossRef] [PubMed]
- Pellerino, A.; Marchesani, E.M.; Bertero, L.; Ricci, A.A.; Mangherini, L.; Bruno, F.; Soffietti, R.; Rudà, R. CNSC-16. Molecular Profiling and Actionable Mutations in Adult Patients with IDH Wild Type Gliomas and Glioneuronal and Neuronal Tumors. Neuro Oncol. 2023, 25, v25–v26. [Google Scholar] [CrossRef]
- Toader, C.; Eva, L.; Costea, D.; Corlatescu, A.D.; Covache-Busuioc, R.-A.; Bratu, B.-G.; Glavan, L.A.; Costin, H.P.; Popa, A.A.; Ciurea, A.V. Low-Grade Gliomas: Histological Subtypes, Molecular Mechanisms, and Treatment Strategies. Brain Sci. 2023, 13, 1700. [Google Scholar] [CrossRef]
- Wu, D.; Liu, X.; Mu, J.; Yang, J.; Wu, F.; Zhou, H. Therapeutic Approaches Targeting Proteins in Tumor-Associated Macrophages and Their Applications in Cancers. Biomolecules 2022, 12, 392. [Google Scholar] [CrossRef]
- Franson, A.; McClellan, B.L.; Varela, M.L.; Comba, A.; Syed, M.F.; Banerjee, K.; Zhu, Z.; Gonzalez, N.; Candolfi, M.; Lowenstein, P.; et al. Development of Immunotherapy for High-Grade Gliomas: Overcoming the Immunosuppressive Tumor Microenvironment. Front. Med. 2022, 9, 966458. [Google Scholar] [CrossRef]
- Richard, Q.; Laurenge, A.; Mallat, M.; Sanson, M.; Castro-Vega, L.J. New Insights into the Immune TME of Adult-Type Diffuse Gliomas. Curr. Opin. Neurol. 2022, 35, 794–802. [Google Scholar] [CrossRef]
- Haddad, A.F.; Young, J.S.; Oh, J.Y.; Okada, H.; Aghi, M.K. The Immunology of Low-Grade Gliomas. Neurosurg. Focus 2022, 52, E2. [Google Scholar] [CrossRef] [PubMed]
- Bunse, L.; Pusch, S.; Bunse, T.; Sahm, F.; Sanghvi, K.; Friedrich, M.; Alansary, D.; Sonner, J.K.; Green, E.; Deumelandt, K.; et al. Suppression of Antitumor T Cell Immunity by the Oncometabolite (R)-2-Hydroxyglutarate. Nat. Med. 2018, 24, 1192–1203. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Sioud, M. Tumor-Associated Macrophage Subsets: Shaping Polarization and Targeting. Int. J. Mol. Sci. 2023, 24, 7493. [Google Scholar] [CrossRef]
- Gao, J.; Liang, Y.; Wang, L. Shaping Polarization Of Tumor-Associated Macrophages In Cancer Immunotherapy. Front. Immunol. 2022, 13, 888713. [Google Scholar] [CrossRef]
- Tzavlaki, K.; Moustakas, A. TGF-β Signaling. Biomolecules 2020, 10, 487. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; He, L.; Lugano, R.; Roodakker, K.; Bergqvist, M.; Smits, A.; Dimberg, A. IDH Mutation Status Is Associated with Distinct Vascular Gene Expression Signatures in Lower-Grade Gliomas. Neuro Oncol. 2018, 20, 1505–1516. [Google Scholar] [CrossRef]
- Li, L.; Wen, Q.; Ding, R. Therapeutic Targeting of VEGF and/or TGF-β to Enhance Anti-PD-(L)1 Therapy: The Evidence from Clinical Trials. Front. Oncol. 2022, 12. [Google Scholar] [CrossRef]
- Wick, W.; Platten, M.; Weller, M. Glioma Cell Invasion: Regulation of Metalloproteinase Activity by TGF-Beta. J. Neurooncol. 2001, 53, 177–185. [Google Scholar] [CrossRef]
- Ye, X.; Xu, S.; Xin, Y.; Yu, S.; Ping, Y.; Chen, L.; Xiao, H.; Wang, B.; Yi, L.; Wang, Q.; et al. Tumor-Associated Microglia/Macrophages Enhance the Invasion of Glioma Stem-like Cells via TGF-Β1 Signaling Pathway. J. Immunol. 2012, 189, 444–453. [Google Scholar] [CrossRef]
- Moon, J.W.; Kong, S.-K.; Kim, B.S.; Kim, H.J.; Lim, H.; Noh, K.; Kim, Y.; Choi, J.-W.; Lee, J.-H.; Kim, Y.-S. IFNγ Induces PD-L1 Overexpression by JAK2/STAT1/IRF-1 Signaling in EBV-Positive Gastric Carcinoma. Sci. Rep. 2017, 7, 17810. [Google Scholar] [CrossRef]
- Smithy, J.W.; Moore, L.M.; Pelekanou, V.; Rehman, J.; Gaule, P.; Wong, P.F.; Neumeister, V.M.; Sznol, M.; Kluger, H.M.; Rimm, D.L. Nuclear IRF-1 Expression as a Mechanism to Assess “Capability” to Express PD-L1 and Response to PD-1 Therapy in Metastatic Melanoma. J. Immunother. Cancer 2017, 5, 25. [Google Scholar] [CrossRef]
- Zhang, X.; Chu, H.; Cheng, Y.; Ren, J.; Wang, W.; Liu, X.; Yan, X. Identification of RUNX1 and IFNGR2 as Prognostic-Related Biomarkers Correlated with Immune Infiltration and Subtype Differentiation of Low-Grade Glioma. Biomol. Biomed. 2023, 23, 405–425. [Google Scholar] [CrossRef] [PubMed]
- Goldman, M.J.; Craft, B.; Hastie, M.; Repečka, K.; McDade, F.; Kamath, A.; Banerjee, A.; Luo, Y.; Rogers, D.; Brooks, A.N.; et al. Visualizing and Interpreting Cancer Genomics Data via the Xena Platform. Nat. Biotechnol. 2020, 38, 675–678. [Google Scholar] [CrossRef] [PubMed]
- Vivian, J.; Rao, A.A.; Nothaft, F.A.; Ketchum, C.; Armstrong, J.; Novak, A.; Pfeil, J.; Narkizian, J.; Deran, A.D.; Musselman-Brown, A.; et al. Toil Enables Reproducible, Open Source, Big Biomedical Data Analyses. Nat. Biotechnol. 2017, 35, 314–316. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Fan, J.; Wang, B.; Traugh, N.; Chen, Q.; Liu, J.S.; Li, B.; Liu, X.S. TIMER: A Web Server for Comprehensive Analysis of Tumor-Infiltrating Immune Cells. Cancer Res. 2017, 77, e108–e110. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Fu, J.; Zeng, Z.; Cohen, D.; Li, J.; Chen, Q.; Li, B.; Liu, X.S. TIMER2.0 for Analysis of Tumor-Infiltrating Immune Cells. Nucleic Acids Res. 2020, 48, W509–W514. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.; Lin, K.; Li, X.; Yuan, X.; Xu, P.; Ni, P.; Xu, D. Redefining Tumor-Associated Macrophage Subpopulations and Functions in the Tumor Microenvironment. Front. Immunol. 2020, 11, 1731. [Google Scholar] [CrossRef] [PubMed]
- Martinez, F.O.; Gordon, S. The M1 and M2 Paradigm of Macrophage Activation: Time for Reassessment. F1000Prime Rep. 2014, 6, 13. [Google Scholar] [CrossRef] [PubMed]
- Müller, S.; Kohanbash, G.; Liu, S.J.; Alvarado, B.; Carrera, D.; Bhaduri, A.; Watchmaker, P.B.; Yagnik, G.; Di Lullo, E.; Malatesta, M.; et al. Single-Cell Profiling of Human Gliomas Reveals Macrophage Ontogeny as a Basis for Regional Differences in Macrophage Activation in the Tumor Microenvironment. Genome Biol. 2017, 18, 234. [Google Scholar] [CrossRef]
- Zeiner, P.S.; Preusse, C.; Golebiewska, A.; Zinke, J.; Iriondo, A.; Muller, A.; Kaoma, T.; Filipski, K.; Müller-Eschner, M.; Bernatz, S.; et al. Distribution and Prognostic Impact of Microglia/Macrophage Subpopulations in Gliomas. Brain Pathol. 2019, 29, 513–529. [Google Scholar] [CrossRef]
- Ji, Q.; Huang, K.; Jiang, Y.; Lei, K.; Tu, Z.; Luo, H.; Zhu, X. Comprehensive Analysis of the Prognostic and Role in Immune Cell Infiltration of MSR1 Expression in lower-grade Gliomas. Cancer Med. 2022, 11, 2020–2035. [Google Scholar] [CrossRef] [PubMed]
- Halliday, N.; Williams, C.; Kennedy, A.; Waters, E.; Pesenacker, A.M.; Soskic, B.; Hinze, C.; Hou, T.Z.; Rowshanravan, B.; Janman, D.; et al. CD86 Is a Selective CD28 Ligand Supporting FoxP3+ Regulatory T Cell Homeostasis in the Presence of High Levels of CTLA-4. Front. Immunol. 2020, 11, 600000. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Li, S.; Liu, F.; Yang, K. Role of CD68 in Tumor Immunity and Prognosis Prediction in Pan-Cancer. Sci. Rep. 2022, 12, 7844. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhang, C.; Zhang, Z.; Han, B.; Shen, Z.; Li, L.; Liu, S.; Zhao, X.; Ye, F.; Zhang, Y. Specific Clinical and Immune Features of CD68 in Glioma via 1024 Samples. Cancer Manag. Res. 2018, 10, 6409–6419. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.-T.; Jin, W.-L. B7-H3/CD276: An Emerging Cancer Immunotherapy. Front. Immunol. 2021, 12, 701006. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.; Li, H.; Xia, Y.; Wang, Y.; Wang, Y.; Shi, Y.; Xing, H.; Qu, T.; Wang, Y.; Ma, W. Immune Checkpoint of B7-H3 in Cancer: From Immunology to Clinical Immunotherapy. J. Hematol. Oncol. 2022, 15, 153. [Google Scholar] [CrossRef]
- Dapash, M.; Hou, D.; Castro, B.; Rashidi, A.; Zhang, P.; Stupp, R.; Miska, J.; Lesniak, M.; Chang, C. EXTH-29. Dual TGFB and PD1 Blockade Promotes Germinal-Center B-Cell Immune responses against Glioblastoma. Neuro Oncol. 2021, 23, vi169. [Google Scholar] [CrossRef]
- Kaminska, B.; Kocyk, M.; Kijewska, M. TGF Beta Signaling and Its Role in Glioma Pathogenesis. Adv. Exp. Med. Biol. 2013, 986, 171–187. [Google Scholar] [CrossRef]
- Han, J.; Alvarez-Breckenridge, C.A.; Wang, Q.-E.; Yu, J. TGF-β Signaling and Its Targeting for Glioma Treatment. Am. J. Cancer Res. 2015, 5, 945–955. [Google Scholar]
- Yamada, N.; Kato, M.; Yamashita, H.; Nistér, M.; Miyazono, K.; Heldin, C.H.; Funa, K. Enhanced Expression of Transforming Growth Factor-Beta and Its Type-I and Type-II Receptors in Human Glioblastoma. Int. J. Cancer 1995, 62, 386–392. [Google Scholar] [CrossRef]
- Samuels, V.; Barrett, J.M.; Bockman, S.; Pantazis, C.G.; Allen, M.B. Immunocytochemical Study of Transforming Growth Factor Expression in Benign and Malignant Gliomas. Am. J. Pathol. 1989, 134, 894–902. [Google Scholar]
- Maxwell, M.; Galanopoulos, T.; Neville-Golden, J.; Antoniades, H.N. Effect of the Expression of Transforming Growth Factor-Β2 in Primary Human Glioblastomas on Immunosuppression and Loss of Immune Surveillance. J. Neurosurg. 1992, 76, 799–804. [Google Scholar] [CrossRef] [PubMed]
- Toedebusch, R.; Grodzki, A.C.; Dickinson, P.J.; Woolard, K.; Vinson, N.; Sturges, B.; Snyder, J.; Li, C.-F.; Nagasaka, O.; Consales, B.; et al. Glioma-Associated Microglia/Macrophages Augment Tumorigenicity in Canine Astrocytoma, a Naturally Occurring Model of Human Glioma. Neurooncol. Adv. 2021, 3. [Google Scholar] [CrossRef] [PubMed]
- Wesolowska, A.; Kwiatkowska, A.; Slomnicki, L.; Dembinski, M.; Master, A.; Sliwa, M.; Franciszkiewicz, K.; Chouaib, S.; Kaminska, B. Microglia-Derived TGF-Beta as an Important Regulator of Glioblastoma Invasion—An Inhibition of TGF-Beta-Dependent Effects by ShRNA against Human TGF-Beta Type II Receptor. Oncogene 2008, 27, 918–930. [Google Scholar] [CrossRef]
- Platanitis, E.; Decker, T. Regulatory Networks Involving STATs, IRFs, and NFκB in Inflammation. Front. Immunol. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Hertzog, P.J.; Weerd, N.A. A Structural “Star” in Interferon Gamma Signaling. Immunol. Cell Biol. 2019, 97, 442–444. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Massagué, J. Mechanisms of TGF-β Signaling from Cell Membrane to the Nucleus. Cell 2003, 113, 685–700. [Google Scholar] [CrossRef] [PubMed]
- De Benedetti, F.; Prencipe, G.; Bracaglia, C.; Marasco, E.; Grom, A.A. Targeting Interferon-γ in Hyperinflammation: Opportunities and Challenges. Nat. Rev. Rheumatol. 2021, 17, 678–691. [Google Scholar] [CrossRef]
- Qazi, S.; Talebi, Z.; Trieu, V. Transforming Growth Factor Beta 2 (TGFB2) and Interferon Gamma Receptor 2 (IFNGR2) MRNA Levels in the Brainstem Tumor Microenvironment (TME) Significantly Impact Overall Survival in Pediatric DMG Patients. Biomedicines 2024, 12, 191. [Google Scholar] [CrossRef]
- Tian, W.; Zhang, W.; Wang, Y.; Jin, R.; Wang, Y.; Guo, H.; Tang, Y.; Yao, X. Recent Advances of IDH1 Mutant Inhibitor in Cancer Therapy. Front. Pharmacol. 2022, 13. [Google Scholar] [CrossRef]
- Unruh, D.; Zewde, M.; Buss, A.; Drumm, M.R.; Tran, A.N.; Scholtens, D.M.; Horbinski, C. Methylation and Transcription Patterns Are Distinct in IDH Mutant Gliomas Compared to Other IDH Mutant Cancers. Sci. Rep. 2019, 9, 8946. [Google Scholar] [CrossRef]
- Ruiz-Rodado, V.; Seki, T.; Dowdy, T.; Lita, A.; Zhang, M.; Han, S.; Yang, C.; Cherukuri, M.K.; Gilbert, M.R.; Larion, M. Metabolic Landscape of a Genetically Engineered Mouse Model of IDH1 Mutant Glioma. Cancers 2020, 12, 1633. [Google Scholar] [CrossRef]
- Philip, B.; Yu, D.X.; Silvis, M.R.; Shin, C.H.; Robinson, J.P.; Robinson, G.L.; Welker, A.E.; Angel, S.N.; Tripp, S.R.; Sonnen, J.A.; et al. Mutant IDH1 Promotes Glioma Formation In Vivo. Cell Rep. 2018, 23, 1553–1564. [Google Scholar] [CrossRef]
- Ban, T.; Kikuchi, M.; Sato, G.R.; Manabe, A.; Tagata, N.; Harita, K.; Nishiyama, A.; Nishimura, K.; Yoshimi, R.; Kirino, Y.; et al. Genetic and Chemical Inhibition of IRF5 Suppresses Pre-Existing Mouse Lupus-like Disease. Nat. Commun. 2021, 12, 4379. [Google Scholar] [CrossRef]
- Song, S.; De, S.; Nelson, V.; Chopra, S.; LaPan, M.; Kampta, K.; Sun, S.; He, M.; Thompson, C.D.; Li, D.; et al. Inhibition of IRF5 Hyperactivation Protects from Lupus Onset and Severity. J. Clin. Investig. 2020, 130, 6700–6717. [Google Scholar] [CrossRef]
- Getu, A.A.; Tigabu, A.; Zhou, M.; Lu, J.; Fodstad, Ø.; Tan, M. New Frontiers in Immune Checkpoint B7-H3 (CD276) Research and Drug Development. Mol. Cancer 2023, 22, 43. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, C.; Prawira, A.; Antonia, S.; Rahma, O.; Tolcher, A.; Cohen, R.B.; Lou, Y.; Hauke, R.; Vogelzang, N.; Zandberg, D.P.; et al. Dual Checkpoint Targeting of B7-H3 and PD-1 with Enoblituzumab and Pembrolizumab in Advanced Solid Tumors: Interim Results from a Multicenter Phase I/II Trial. J. Immunother. Cancer 2022, 10, e004424. [Google Scholar] [CrossRef]
- Bottino, C.; Vitale, C.; Dondero, A.; Castriconi, R. B7-H3 in Pediatric Tumors: Far beyond Neuroblastoma. Cancers 2023, 15, 3279. [Google Scholar] [CrossRef]
- Pulido, R.; Nunes-Xavier, C.E. Hopes on Immunotherapy Targeting B7-H3 in Neuroblastoma. Transl. Oncol. 2023, 27, 101580. [Google Scholar] [CrossRef]
- Anderson, J.; Majzner, R.G.; Sondel, P.M. Immunotherapy of Neuroblastoma: Facts and Hopes. Clin. Cancer Res. 2022, 28, 3196–3206. [Google Scholar] [CrossRef]
- Rasic, P.; Jeremic, M.; Jeremic, R.; Dusanovic Pjevic, M.; Rasic, M.; Djuricic, S.M.; Milickovic, M.; Vukadin, M.; Mijovic, T.; Savic, D. Targeting B7-H3—A Novel Strategy for the Design of Anticancer Agents for Extracranial Pediatric Solid Tumors Treatment. Molecules 2023, 28, 3356. [Google Scholar] [CrossRef]
- Vitanza, N.A.; Wilson, A.L.; Huang, W.; Seidel, K.; Brown, C.; Gustafson, J.A.; Yokoyama, J.K.; Johnson, A.J.; Baxter, B.A.; Koning, R.W.; et al. Intraventricular B7-H3 CAR T Cells for Diffuse Intrinsic Pontine Glioma: Preliminary First-in-Human Bioactivity and Safety. Cancer Discov. 2023, 13, 114–131. [Google Scholar] [CrossRef]
- Kjellman, C.; Olofsson, S.P.; Hansson, O.; Von Schantz, T.; Lindvall, M.; Nilsson, I.; Salford, L.G.; Sjögren, H.O.; Widegren, B. Expression of TGF-Beta Isoforms, TGF-Beta Receptors, and SMAD Molecules at Different Stages of Human Glioma. Int. J. Cancer 2000, 89, 251–258. [Google Scholar] [CrossRef]
- Uckun, F.M.; Qazi, S.; Trieu, V.N. Proof of Concept for the Clinical Utility of a Convection Enhanced Delivery Platform for Immuno-Oncology Drugs against Brain Tumors. Cancer Rep. Rev. 2019, 3, 1–8. [Google Scholar] [CrossRef]
- Uckun, F.M.; Qazi, S.; Hwang, L.; Trieu, V.N. Recurrent or Refractory High-Grade Gliomas Treated by Convection-Enhanced Delivery of a TGFβ2-Targeting RNA Therapeutic: A Post-Hoc Analysis with Long-Term Follow-Up. Cancers 2019, 11, 1892. [Google Scholar] [CrossRef] [PubMed]
- Tong, S.; Cinelli, M.A.; El-Sayed, N.S.; Huang, H.; Patel, A.; Silverman, R.B.; Yang, S. Inhibition of Interferon-Gamma-Stimulated Melanoma Progression by Targeting Neuronal Nitric Oxide Synthase (NNOS). Sci. Rep. 2022, 12, 1701. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trieu, V.; Maida, A.E.; Qazi, S. Transforming Growth Factor Beta 2 (TGFB2) mRNA Levels, in Conjunction with Interferon-Gamma Receptor Activation of Interferon Regulatory Factor 5 (IRF5) and Expression of CD276/B7-H3, Are Therapeutically Targetable Negative Prognostic Markers in Low-Grade Gliomas. Cancers 2024, 16, 1202. https://doi.org/10.3390/cancers16061202
Trieu V, Maida AE, Qazi S. Transforming Growth Factor Beta 2 (TGFB2) mRNA Levels, in Conjunction with Interferon-Gamma Receptor Activation of Interferon Regulatory Factor 5 (IRF5) and Expression of CD276/B7-H3, Are Therapeutically Targetable Negative Prognostic Markers in Low-Grade Gliomas. Cancers. 2024; 16(6):1202. https://doi.org/10.3390/cancers16061202
Chicago/Turabian StyleTrieu, Vuong, Anthony E. Maida, and Sanjive Qazi. 2024. "Transforming Growth Factor Beta 2 (TGFB2) mRNA Levels, in Conjunction with Interferon-Gamma Receptor Activation of Interferon Regulatory Factor 5 (IRF5) and Expression of CD276/B7-H3, Are Therapeutically Targetable Negative Prognostic Markers in Low-Grade Gliomas" Cancers 16, no. 6: 1202. https://doi.org/10.3390/cancers16061202
APA StyleTrieu, V., Maida, A. E., & Qazi, S. (2024). Transforming Growth Factor Beta 2 (TGFB2) mRNA Levels, in Conjunction with Interferon-Gamma Receptor Activation of Interferon Regulatory Factor 5 (IRF5) and Expression of CD276/B7-H3, Are Therapeutically Targetable Negative Prognostic Markers in Low-Grade Gliomas. Cancers, 16(6), 1202. https://doi.org/10.3390/cancers16061202