
Circular STAG2 RNA Modulates Bladder Cancer Progression via miR-145-5p/TAGLN2 and Is Considered as a Biomarker for Recurrence
 ,
,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Patient Specimens
2.3. RNA Extraction and RT-qPCR
2.4. RNase R Treatment
2.5. Molecular Cloning and Cell Transfection
2.6. Wound-Healing Assay
2.7. Transwell Invasion Assay
2.8. Dual-Luciferase Reporter Assay
2.9. Statistical Analysis
3. Results
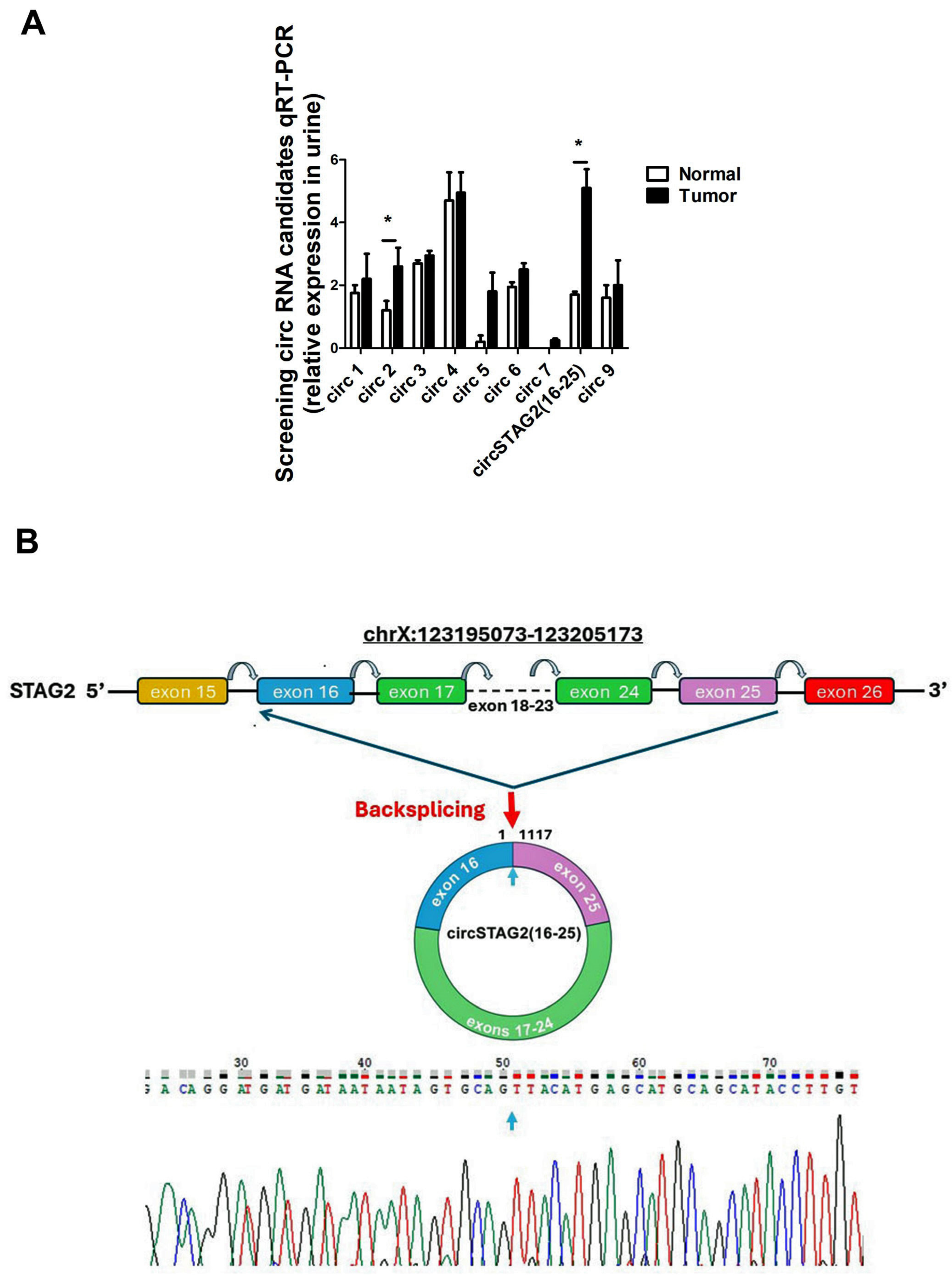
3.1. CircSTAG2(16–25) Is Up-Regulated in BCa and in More Aggressive Urothelial Carcinoma Cell Lines
3.2. CircSTAG2(16–25) Is Expressed at Higher Levels in Urine in Recurrent Samples
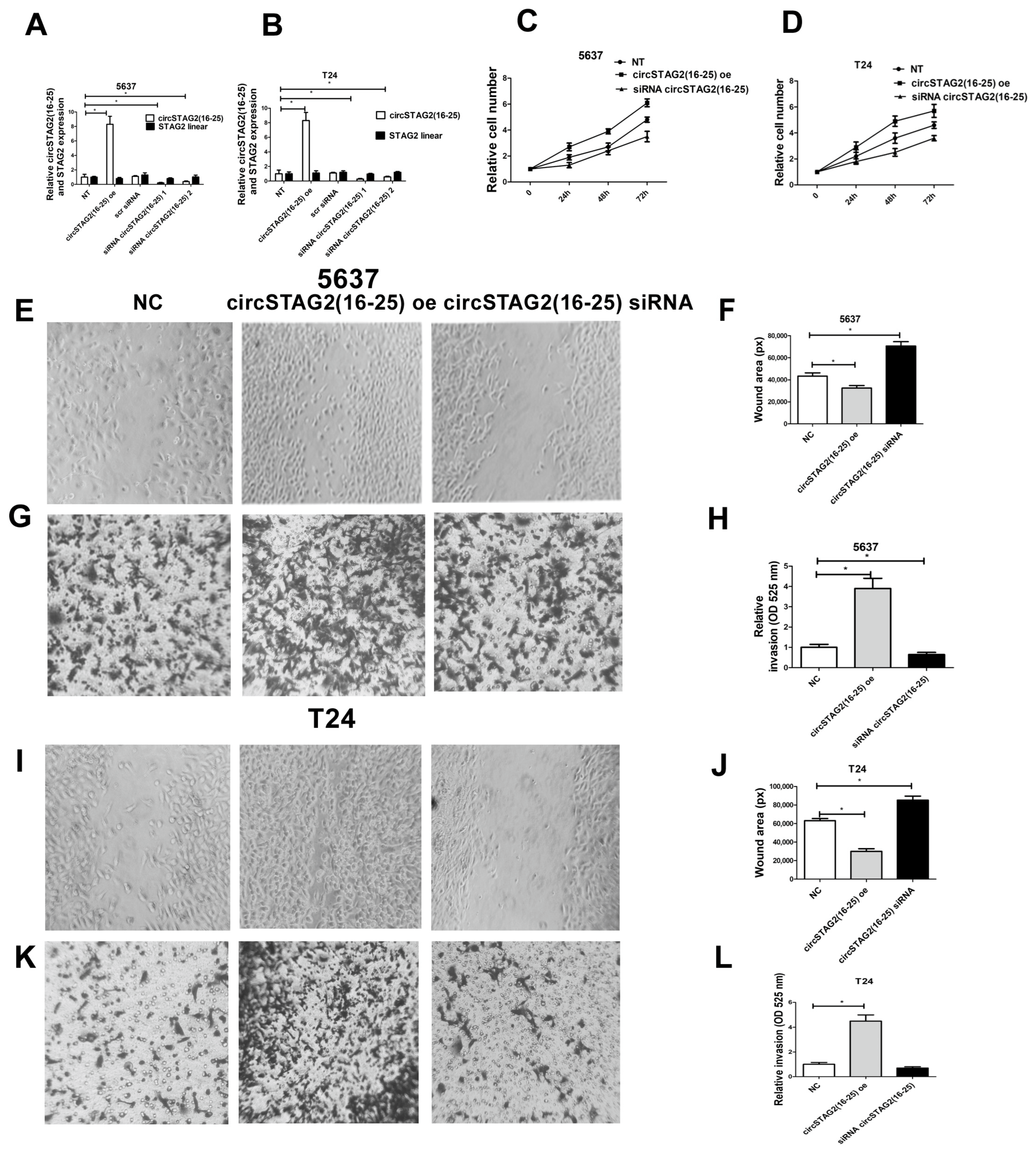
3.3. Knockdown of circSTAG2(16–25) Inhibited BCa Cell Proliferation, While circSTAG2(16–25) Overexpression Promoted It
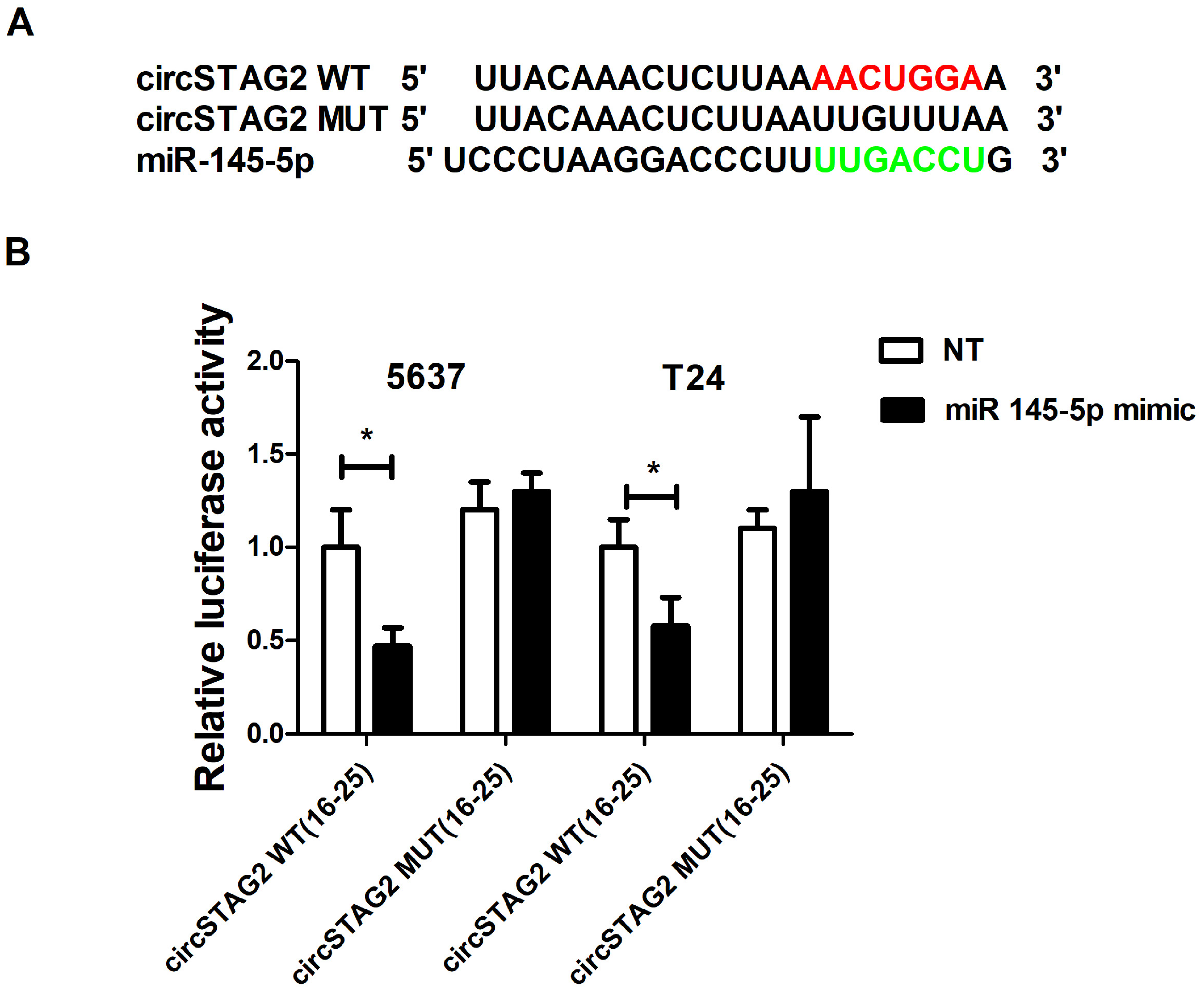
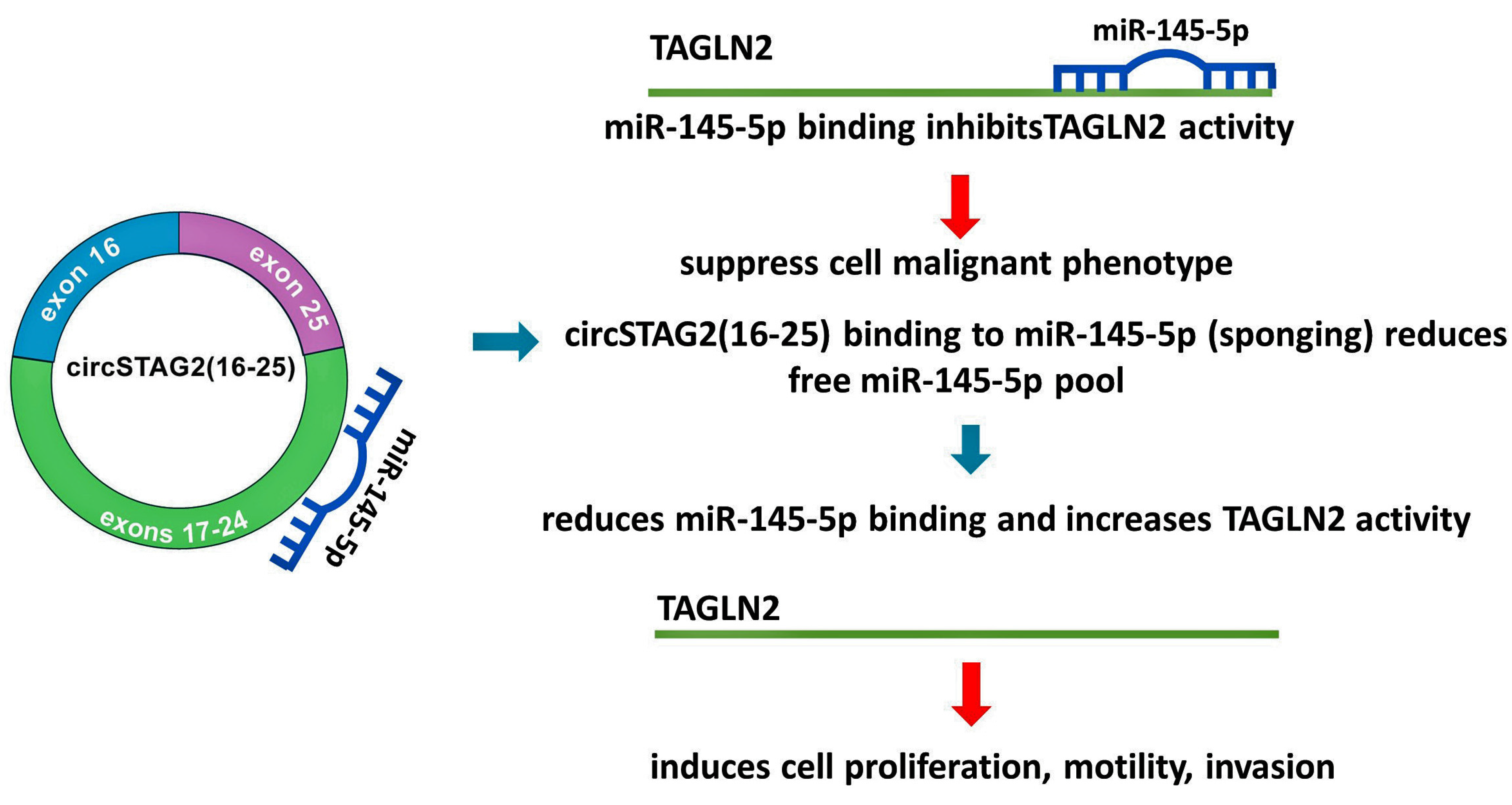
3.4. CircSTAG2 May Act as a Sponge for miR-145-5p in BCa Cells
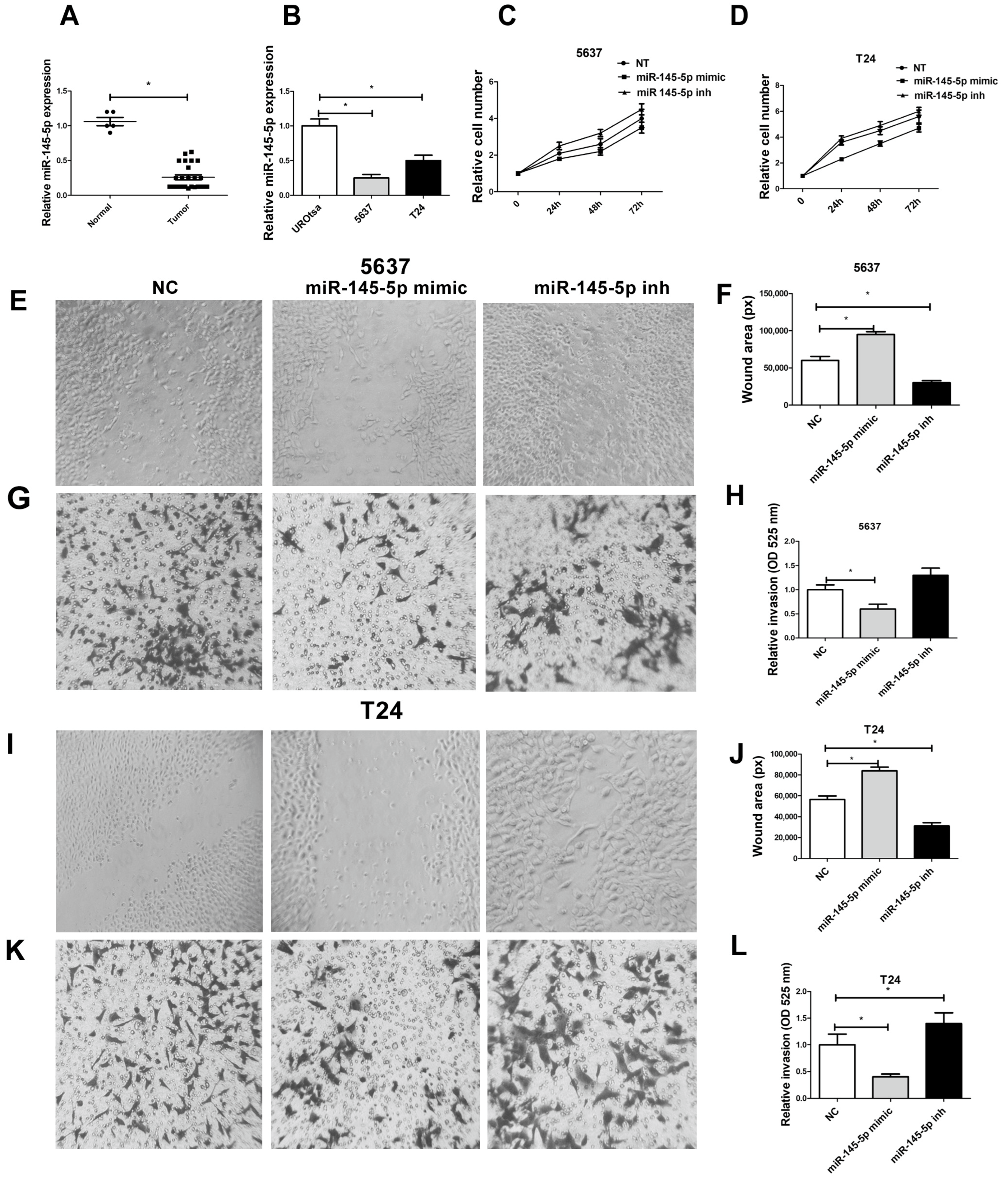
3.5. miR-145-5p Expression Is Correlated with BCa Progression and It Suppresses Proliferation, Migration, and Invasion of BCa Cells
3.6. miR-145-5p Suppressed BCa Cell Proliferation, Migration, and Invasion by Targeting TAGLN2
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dyrskjot, L.; Hansel, D.E.; Efstathiou, J.A.; Knowles, M.A.; Galsky, M.D.; Teoh, J.; Theodorescu, D. Bladder cancer. Nat. Rev. Dis. Prim. 2023, 9, 58. [Google Scholar] [CrossRef]
- Varchulova Novakova, Z.; Kuniakova, M.; Ziaran, S.; Harsanyi, S. Molecular Biomarkers of Bladder Cancer: A Mini-Review. Physiol. Res. 2023, 72, S247–S256. [Google Scholar] [CrossRef]
- Lee, H.H.; Kim, S.H. Review of non-invasive urinary biomarkers in bladder cancer. Transl. Cancer Res. 2020, 9, 6554–6564. [Google Scholar] [CrossRef] [PubMed]
- Papatsirou, M.; Artemaki, P.I.; Karousi, P.; Scorilas, A.; Kontos, C.K. Circular RNAs: Emerging Regulators of the Major Signaling Pathways Involved in Cancer Progression. Cancers 2021, 13, 2744. [Google Scholar] [CrossRef]
- Vo, J.N.; Cieslik, M.; Zhang, Y.; Shukla, S.; Xiao, L.; Zhang, Y.; Wu, Y.M.; Dhanasekaran, S.M.; Engelke, C.G.; Cao, X.; et al. The Landscape of Circular RNA in Cancer. Cell 2019, 176, 869–881.E13. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.H.; Wang, Y.; Zhang, Y.; Zheng, S.F.; Feng, T.; Tian, X.; Abudurexiti, M.; Wang, Z.D.; Zhu, W.K.; Su, J.Q.; et al. The function and mechanisms of action of circular RNAs in Urologic Cancer. Mol. Cancer 2023, 22, 61. [Google Scholar] [CrossRef]
- Ebbesen, K.K.; Hansen, T.B.; Kjems, J. Insights into circular RNA biology. RNA Biol. 2017, 14, 1035–1045. [Google Scholar] [CrossRef] [PubMed]
- Salzman, J. Circular RNA Expression: Its Potential Regulation and Function. Trends Genet. 2016, 32, 309–316. [Google Scholar] [CrossRef]
- Wilusz, J. Circular RNA and Splicing: Skip Happens. J. Mol. Biol. 2015, 427, 2411–2413. [Google Scholar] [CrossRef]
- Huang, Y.; Zhu, Q. Mechanisms Regulating Abnormal Circular RNA Biogenesis in Cancer. Cancers 2021, 13, 4185. [Google Scholar] [CrossRef]
- Sun, K.; Wang, D.; Yang, B.B.; Ma, J. The Emerging Functions of Circular RNAs in Bladder Cancer. Cancers 2021, 13, 4618. [Google Scholar] [CrossRef]
- Gordon, N.S.; Humayun-Zakaria, N.; Goel, A.; Abbotts, B.; Zeegers, M.P.; Cheng, K.K.; James, N.D.; Arnold, R.; Bryan, R.T.; Ward, D.G. STAG2 Protein Expression in Non-muscle-invasive Bladder Cancer: Associations with Sex, Genomic and Transcriptomic Changes, and Clinical Outcomes. Eur. Urol. Open Sci. 2022, 38, 88–95. [Google Scholar] [CrossRef]
- Taber, A.; Park, Y.; Lelo, A.; Prip, F.; Xiao, J.; Berry, D.L.; Chaldekas, K.; Jensen, J.B.; Philips, G.; Kim, J.S.; et al. STAG2 as a prognostic biomarker in low-grade non-muscle invasive bladder cancer. Urol. Oncol. 2021, 39, 438.e1–438.e9. [Google Scholar] [CrossRef]
- Lelo, A.; Prip, F.; Harris, B.T.; Solomon, D.; Berry, D.L.; Chaldekas, K.; Kumar, A.; Simko, J.; Jensen, J.B.; Bhattacharyya, P.; et al. STAG2 Is a Biomarker for Prediction of Recurrence and Progression in Papillary Non-Muscle-Invasive Bladder Cancer. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2018, 24, 4145–4153. [Google Scholar] [CrossRef]
- Aquila, L.; Ohm, J.; Woloszynska-Read, A. The role of STAG2 in bladder cancer. Pharmacol. Res. 2018, 131, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Qiao, Y.; Zhu, X.; Li, A.; Yang, S.; Zhang, J. Complete loss of STAG2 expression is an indicator of good prognosis in patients with bladder cancer. Tumor Biol. 2016, 37, 10279–10286. [Google Scholar] [CrossRef]
- Taylor, C.F.; Platt, F.M.; Hurst, C.D.; Thygesen, H.H.; Knowles, M.A. Frequent inactivating mutations of STAG2 in bladder cancer are associated with low tumour grade and stage and inversely related to chromosomal copy number changes. Hum. Mol. Genet. 2014, 23, 1964–1974. [Google Scholar] [CrossRef]
- Black, P. Frequent truncating mutations of STAG2 in bladder cancer. Urology 2014, 83, 691–692. [Google Scholar] [CrossRef] [PubMed]
- Mamdouh, S.; Sherif, H.; Romeih, M.; Elesaily, K. Urine micro-RNA signature as a potential non-invasive diagnostic biomarker in bladder cancer. Asian Pac. J. Cancer Prev. APJCP 2023, 24, 121–131. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Jiang, M.; Liu, Q.; Han, Z.; Zhao, Y.; Ji, S. miR-145-5p inhibits the proliferation and migration of bladder cancer cells by targeting TAGLN2. Oncol. Lett. 2018, 16, 6355–6360. [Google Scholar] [CrossRef]
- Minami, K.; Taniguchi, K.; Sugito, N.; Kuranaga, Y.; Inamoto, T.; Takahara, K.; Takai, T.; Yoshikawa, Y.; Kiyama, S.; Akao, Y.; et al. MiR-145 negatively regulates Warburg effect by silencing KLF4 and PTBP1 in bladder cancer cells. Oncotarget 2017, 8, 33064–33077. [Google Scholar] [CrossRef] [PubMed]
- Fujii, T.; Shimada, K.; Tatsumi, Y.; Hatakeyama, K.; Obayashi, C.; Fujimoto, K.; Konishi, N. microRNA-145 promotes differentiation in human urothelial carcinoma through down-regulation of syndecan-1. BMC Cancer 2015, 15, 818. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Xu, T.; Wang, L.; Wang, X.; Zhong, S.; Xu, C.; Shen, Z. MicroRNA-145 directly targets the insulin-like growth factor receptor I in human bladder cancer cells. FEBS Lett. 2014, 588, 3180–3185. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Li, M.; Wang, L.; Zhou, Y.; Zhou, J.; Pan, H.; Qu, P. microRNA-145 inhibits cell proliferation, migration and invasion by targeting matrix metallopeptidase-11 in renal cell carcinoma. Mol. Med. Rep. 2014, 10, 393–398. [Google Scholar] [CrossRef] [PubMed]
- Kou, B.; Gao, Y.; Du, C.; Shi, Q.; Xu, S.; Wang, C.Q.; Wang, X.; He, D.; Guo, P. miR-145 inhibits invasion of bladder cancer cells by targeting PAK1. Urol. Oncol. 2014, 32, 846–854. [Google Scholar] [CrossRef] [PubMed]
- Xiao, M.S.; Wilusz, J.E. An improved method for circular RNA purification using RNase R that efficiently removes linear RNAs containing G-quadruplexes or structured 3′ ends. Nucleic Acids Res. 2019, 47, 8755–8769. [Google Scholar] [CrossRef]
- Kramer, M.C.; Liang, D.; Tatomer, D.C.; Gold, B.; March, Z.M.; Cherry, S.; Wilusz, J.E. Combinatorial control of Drosophila circular RNA expression by intronic repeats, hnRNPs, and SR proteins. Genes Dev. 2015, 29, 2168–2182. [Google Scholar] [CrossRef]
- Lara-Reyes, J.A.; Jimenez-Buendia, M.G.; Aranda-Abreu, G.E.; Herrera-Covarrubias, D.; Sampieri, C.L.; Aquino-Galvez, A.; Manzo-Denes, J.; Hernandez-Aguilar, M.E.; Rojas-Duran, F. Razor scrape assay, an alternative variation to wound and healing assays. MethodsX 2020, 7, 101135. [Google Scholar] [CrossRef]
- Sheng, N.; Tan, G.; You, W.; Chen, H.; Gong, J.; Chen, D.; Zhang, H.; Wang, Z. MiR-145 inhibits human colorectal cancer cell migration and invasion via PAK4-dependent pathway. Cancer Med. 2017, 6, 1331–1340. [Google Scholar] [CrossRef]
- Eichelser, C.; Stuckrath, I.; Muller, V.; Milde-Langosch, K.; Wikman, H.; Pantel, K.; Schwarzenbach, H. Increased serum levels of circulating exosomal microRNA-373 in receptor-negative breast cancer patients. Oncotarget 2014, 5, 9650–9663. [Google Scholar] [CrossRef]
- Park, Y.; Lelo, A.; Harris, B.; Berry, D.L.; Chaldekas, K.; Kim, J.S.; Waldman, T. Identification of STAG2-Mutant Bladder Cancers by Immunohistochemistry. Methods Mol. Biol. 2023, 2684, 145–151. [Google Scholar] [CrossRef]
- Dudekula, D.B.; Panda, A.C.; Grammatikakis, I.; De, S.; Abdelmohsen, K.; Gorospe, M. CircInteractome: A web tool for exploring circular RNAs and their interacting proteins and microRNAs. RNA Biol. 2016, 13, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Zhao, W.; Chen, Z.; Li, M.; Li, S.; Wu, B.; Bu, R. Circ_0058063 regulates CDK6 to promote bladder cancer progression by sponging miR-145-5p. J. Cell. Physiol. 2019, 234, 4812–4824. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Tan, D.; Xie, Y.; Wu, L.; Wu, S.; Tang, Y.; Luo, Y.; Xiao, X.; Li, X. CircEPSTI1 Promotes the Proliferation of HER2-Positive Breast Cancer Cells via circEPSTI1/miR-145/ERBB3 Axis. J. Oncol. 2022, 2022, 1028851. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.; Zhou, L.H.; Ge, Y.Z.; Ping, W.W.; Wu, X.; Xu, Z.L.; Wang, M.; Sha, Z.L.; Jia, R.P. MicroRNA-133b suppresses bladder cancer malignancy by targeting TAGLN2-mediated cell cycle. J. Cell. Physiol. 2019, 234, 4910–4923. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.L.; Chung, T.; Wu, C.C.; Ng, K.F.; Yu, J.S.; Tsai, C.H.; Chang, Y.S.; Liang, Y.; Tsui, K.H.; Chen, Y.T. Comparative Tissue Proteomics of Microdissected Specimens Reveals Novel Candidate Biomarkers of Bladder Cancer. Mol. Cell. Proteom. 2015, 14, 2466–2478. [Google Scholar] [CrossRef] [PubMed]
- Yoshino, H.; Chiyomaru, T.; Enokida, H.; Kawakami, K.; Tatarano, S.; Nishiyama, K.; Nohata, N.; Seki, N.; Nakagawa, M. The tumour-suppressive function of miR-1 and miR-133a targeting TAGLN2 in bladder cancer. Br. J. Cancer 2011, 104, 808–818. [Google Scholar] [CrossRef] [PubMed]
- Rybak-Wolf, A.; Stottmeister, C.; Glazar, P.; Jens, M.; Pino, N.; Giusti, S.; Hanan, M.; Behm, M.; Bartok, O.; Ashwal-Fluss, R.; et al. Circular RNAs in the Mammalian Brain Are Highly Abundant, Conserved, and Dynamically Expressed. Mol. Cell 2015, 58, 870–885. [Google Scholar] [CrossRef]
- Athans, S.; Krishnan, N.; Ramakrishnan, S.; Cortes Gomez, E.; Lage-Vickers, S.; Rak, M.; Kazmierczak, Z.; Ohm, J.; Attwood, K.; Wang, J.; et al. STAG2 expression is associated with adverse survival outcomes and regulates cell phenotype in muscle-invasive bladder cancer. Cancer Res. Commun. 2022, 2, 1129–1143. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Li, D.; Shao, J.; Wang, X. Animal models for bladder cancer: The model establishment and evaluation (Review). Oncol. Lett. 2015, 9, 1515–1519. [Google Scholar] [CrossRef]
- Jager, W.; Moskalev, I.; Raven, P.; Goriki, A.; Bidnur, S.; Black, P.C. Orthotopic Mouse Models of Urothelial Cancer. Methods Mol. Biol. 2018, 1655, 177–197. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Template | Primer 5′–3′ | |
|---|---|---|
| hsa_circ_0091459 circ 1 | Forward Reverse | GTCCAAACCGAATGAATGG TGGTAAAATGGTGGCTTGC |
| hsa_circ_0091460 circ 2 | Forward Reverse | CAGTCGGTGGTAGATGATTGGA GTGGCTTGCCTTCTTTCTTCT |
| hsa_circ_0139698 circ 3 | Forward Reverse | TCATTGCAGTCTGAGTTGCTC CACGTTGGTCAGGTAATGTTG |
| hsa_circ_0091464 circ 4 | Forward Reverse |
AGAACTTGCTCGACGTTTTGC
TGGCCGGTGAGCTGAATAAA |
| hsa_circ_0139699 circ 5 | Forward Reverse |
GCCATTGCCATGCTACACAA
TGAAGGCCTGTTCCTTAACAGTAG |
| hsa_circ_0139701 circ 6 | Forward Reverse | GGGGGTCAACAGTACGGAGT CACGCCCTCCTGACATAATC |
| hsa_circ_0139700 circ 7 | Forward Reverse | CTAATCCGCAAGGGGAGAG TCTTTAAACGCAGCAAGTCC CAGCTCAGTAGCACAGTCCC |
| circSTAG2(16–25) | Forward Reverse |
TCATTGCAGTCTGAGTTGCT
CAGCTCAGTAGCACAGTCCC |
| hsa_circ_0139702 circ 9 | Forward Reverse | CTAATCCGCAAGGGGAGAG AAGCAAAACGTCGAGCAAGT |
| miR-145-5p | Forward Reverse |
CACGCGGTCCAGTTTTCC
CCAGTGCAGGGTCCGAGGTA |
| U6 | Forward Reverse |
GCTTCGGCAGCACATATACTAAAAT
CGCTTCACGAATTTGCGTGTCAT |
| TAGLN2 | Forward Reverse |
AGAACCCTGAAGTCCTCCCT
ATATGCAGGTCCCCTGTTGG |
| β-ACT | Forward Reverse | CACCATTGGCAATGAGCGGTTC AGGTCTTTGCGGATGTCCACGT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Du, C.; Waltzer, W.C.; Wilusz, J.E.; Spaliviero, M.; Darras, F.; Romanov, V. Circular STAG2 RNA Modulates Bladder Cancer Progression via miR-145-5p/TAGLN2 and Is Considered as a Biomarker for Recurrence. Cancers 2024, 16, 978. https://doi.org/10.3390/cancers16050978
Du C, Waltzer WC, Wilusz JE, Spaliviero M, Darras F, Romanov V. Circular STAG2 RNA Modulates Bladder Cancer Progression via miR-145-5p/TAGLN2 and Is Considered as a Biomarker for Recurrence. Cancers. 2024; 16(5):978. https://doi.org/10.3390/cancers16050978
Chicago/Turabian StyleDu, Chris, Wayne C. Waltzer, Jeremy E. Wilusz, Massimiliano Spaliviero, Frank Darras, and Victor Romanov. 2024. "Circular STAG2 RNA Modulates Bladder Cancer Progression via miR-145-5p/TAGLN2 and Is Considered as a Biomarker for Recurrence" Cancers 16, no. 5: 978. https://doi.org/10.3390/cancers16050978
APA StyleDu, C., Waltzer, W. C., Wilusz, J. E., Spaliviero, M., Darras, F., & Romanov, V. (2024). Circular STAG2 RNA Modulates Bladder Cancer Progression via miR-145-5p/TAGLN2 and Is Considered as a Biomarker for Recurrence. Cancers, 16(5), 978. https://doi.org/10.3390/cancers16050978