
The Potential of Siglecs and Sialic Acids as Biomarkers and Therapeutic Targets in Tumor Immunotherapy
 ,
,
Abstract
Simple Summary
Abstract
1. Introduction
2. Siglecs: Classification, Structure, Function, and Biological Regulation
3. Sialylation Dysregulation and Siglec-Mediated Immune Evasion in Tumorigenesis
3.1. Siglecs and Ligand-Mediated Immune Evasion Mechanisms in Oncogenesis
3.2. Siglecs in Immunosuppression across Cell Types
3.2.1. Siglecs and NK Cells
3.2.2. Siglecs and B Cells
3.2.3. Siglecs and T Cells
3.2.4. Siglecs and Macrophages
3.2.5. Siglecs and Dendritic Cells (DCs)
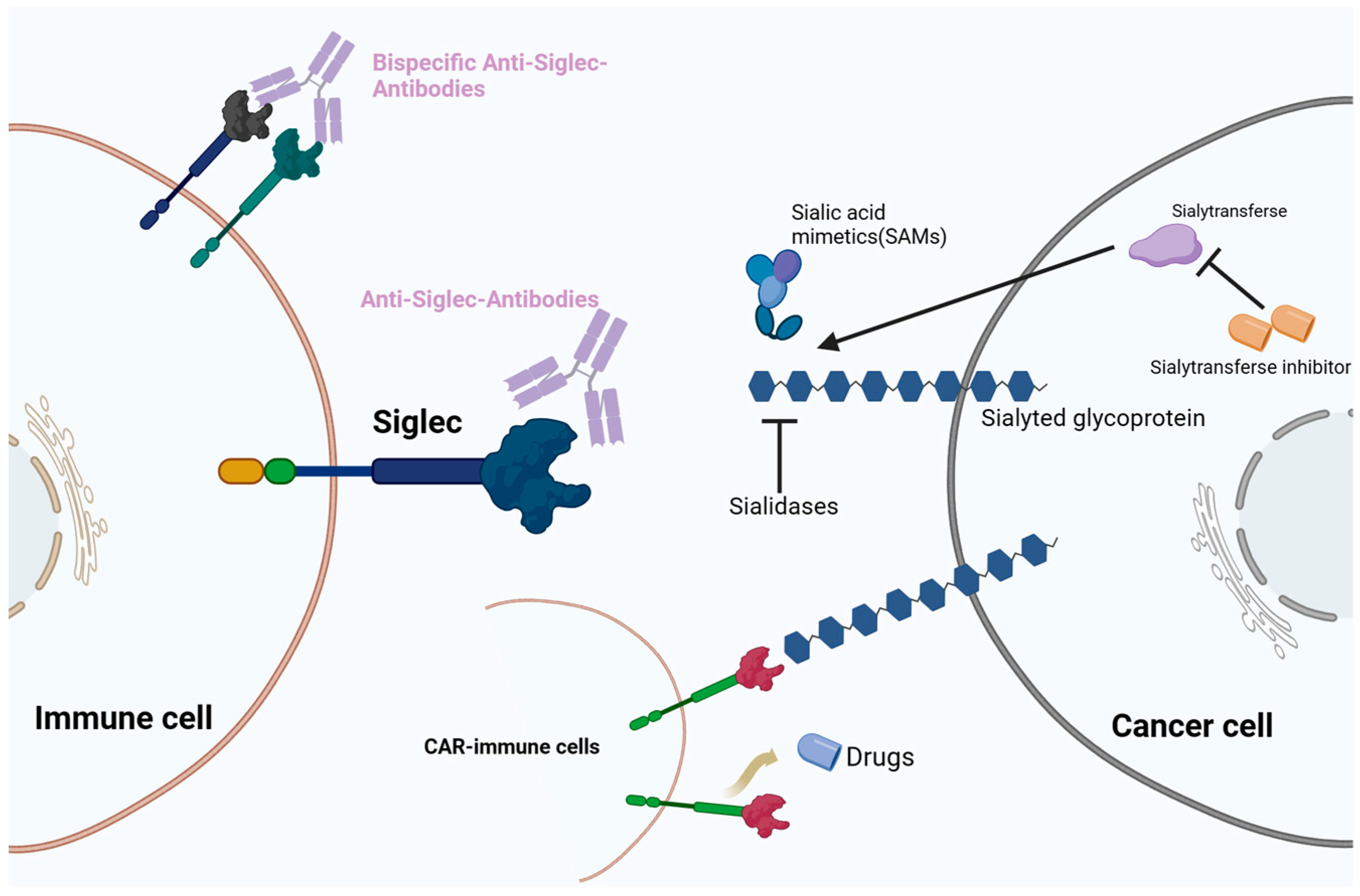
4. Cancer Therapeutic Strategies Targeting the Sialoglycan–Siglec Immune Checkpoint
4.1. Monoclonal Antibodies Targeting Siglecs
4.2. BsAbs
4.3. Antibody-Based Cell Depletion
4.4. CAR-T Therapeutic Approaches
4.5. Sialic Acid Mimetics (SAMs) in Siglec Targeting
5. Siglecs as Disease Biomarkers
6. Discussion and Future Perspectives
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Carlino, M.S.; Larkin, J.; Long, G.V. Immune Checkpoint Inhibitors in Melanoma. Lancet 2021, 398, 1002–1014. [Google Scholar] [CrossRef] [PubMed]
- Granier, C.; De Guillebon, E.; Blanc, C.; Roussel, H.; Badoual, C.; Colin, E.; Saldmann, A.; Gey, A.; Oudard, S.; Tartour, E. Mechanisms of Action and Rationale for the Use of Checkpoint Inhibitors in Cancer. ESMO Open 2017, 2, e000213. [Google Scholar] [CrossRef]
- Kubli, S.P.; Berger, T.; Araujo, D.V.; Siu, L.L.; Mak, T.W. Beyond Immune Checkpoint Blockade: Emerging Immunological Strategies. Nat. Rev. Drug Discov. 2021, 20, 899–919. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Siddiqui, B.A.; Anandhan, S.; Yadav, S.S.; Subudhi, S.K.; Gao, J.; Goswami, S.; Allison, J.P. The Next Decade of Immune Checkpoint Therapy. Cancer Discov. 2021, 11, 838–857. [Google Scholar] [CrossRef] [PubMed]
- Llovet, J.M.; Castet, F.; Heikenwalder, M.; Maini, M.K.; Mazzaferro, V.; Pinato, D.J.; Pikarsky, E.; Zhu, A.X.; Finn, R.S. Immunotherapies for Hepatocellular Carcinoma. Nat. Rev. Clin. Oncol. 2022, 19, 151–172. [Google Scholar] [CrossRef]
- Sharma, P.; Goswami, S.; Raychaudhuri, D.; Siddiqui, B.A.; Singh, P.; Nagarajan, A.; Liu, J.; Subudhi, S.K.; Poon, C.; Gant, K.L.; et al. Immune Checkpoint Therapy-Current Perspectives and Future Directions. Cell 2023, 186, 1652–1669. [Google Scholar] [CrossRef]
- Daly, J.; Carlsten, M.; O’Dwyer, M. Sugar Free: Novel Immunotherapeutic Approaches Targeting Siglecs and Sialic Acids to Enhance Natural Killer Cell Cytotoxicity Against Cancer. Front. Immunol. 2019, 10, 1047. [Google Scholar] [CrossRef]
- Macauley, M.S.; Crocker, P.R.; Paulson, J.C. Siglec-Mediated Regulation of Immune Cell Function in Disease. Nat. Rev. Immunol. 2014, 14, 653–666. [Google Scholar] [CrossRef]
- Crocker, P.R.; Paulson, J.C.; Varki, A. Siglecs and Their Roles in the Immune System. Nat. Rev. Immunol. 2007, 7, 255–266. [Google Scholar] [CrossRef]
- Specific Inactivation of Two Immunomodulatory SIGLEC Genes during Human Evolution—PubMed. Available online: https://pubmed.ncbi.nlm.nih.gov/22665810/ (accessed on 9 December 2023).
- Duan, S.; Paulson, J.C. Siglecs as Immune Cell Checkpoints in Disease. Annu. Rev. Immunol. 2020, 38, 365–395. [Google Scholar] [CrossRef]
- Smith, B.A.H.; Bertozzi, C.R. The Clinical Impact of Glycobiology: Targeting Selectins, Siglecs and Mammalian Glycans. Nat. Rev. Drug Discov. 2021, 20, 217–243. [Google Scholar] [CrossRef] [PubMed]
- Angata, T. Siglecs That Associate with DAP12. Adv. Exp. Med. Biol. 2020, 1204, 215–230. [Google Scholar] [CrossRef] [PubMed]
- McVicar, D.W.; Burshtyn, D.N. Intracellular Signaling by the Killer Immunoglobulin-like Receptors and Ly49. Sci. STKE 2001, 2001, re1. [Google Scholar] [CrossRef] [PubMed]
- Daëron, M.; Jaeger, S.; Du Pasquier, L.; Vivier, E. Immunoreceptor Tyrosine-Based Inhibition Motifs: A Quest in the Past and Future. Immunol. Rev. 2008, 224, 11–43. [Google Scholar] [CrossRef] [PubMed]
- Barrow, A.D.; Trowsdale, J. You Say ITAM and I Say ITIM, Let’s Call the Whole Thing off: The Ambiguity of Immunoreceptor Signalling. Eur. J. Immunol. 2006, 36, 1646–1653. [Google Scholar] [CrossRef] [PubMed]
- Ostroumov, D.; Duong, S.; Wingerath, J.; Woller, N.; Manns, M.P.; Timrott, K.; Kleine, M.; Ramackers, W.; Roessler, S.; Nahnsen, S.; et al. Transcriptome Profiling Identifies TIGIT as a Marker of T-Cell Exhaustion in Liver Cancer. Hepatology 2021, 73, 1399–1418. [Google Scholar] [CrossRef] [PubMed]
- Furumoto, Y.; Nunomura, S.; Terada, T.; Rivera, J.; Ra, C. The FcepsilonRIbeta Immunoreceptor Tyrosine-Based Activation Motif Exerts Inhibitory Control on MAPK and IkappaB Kinase Phosphorylation and Mast Cell Cytokine Production. J. Biol. Chem. 2004, 279, 49177–49187. [Google Scholar] [CrossRef]
- Macauley, M.S.; Pfrengle, F.; Rademacher, C.; Nycholat, C.M.; Gale, A.J.; von Drygalski, A.; Paulson, J.C. Antigenic Liposomes Displaying CD22 Ligands Induce Antigen-Specific B Cell Apoptosis. J. Clin. Investig. 2013, 123, 3074–3083. [Google Scholar] [CrossRef]
- Patsoukis, N.; Duke-Cohan, J.S.; Chaudhri, A.; Aksoylar, H.-I.; Wang, Q.; Council, A.; Berg, A.; Freeman, G.J.; Boussiotis, V.A. Interaction of SHP-2 SH2 Domains with PD-1 ITSM Induces PD-1 Dimerization and SHP-2 Activation. Commun. Biol. 2020, 3, 128. [Google Scholar] [CrossRef]
- Yu, Y.; Peng, W. Recent Progress in Targeting the Sialylated Glycan-SIGLEC Axis in Cancer Immunotherapy. Cancer Biol. Med. 2023, 20, 369–384. [Google Scholar] [CrossRef]
- Zou, W.; Reeve, J.L.; Liu, Y.; Teitelbaum, S.L.; Ross, F.P. DAP12 Couples C-Fms Activation to the Osteoclast Cytoskeleton by Recruitment of Syk. Mol. Cell 2008, 31, 422–431. [Google Scholar] [CrossRef] [PubMed]
- Dalton, R.; Calescibetta, A.; Zhou, J.M.; Maurin, M.; Ward, G.; Trinh, T.L.; Tu, N.; Gilvary, D.; Chen, X.; Cheng, P.; et al. Constitutively Activated DAP12 Induces Functional Anti-Tumor Activation and Maturation of Human Monocyte-Derived DC. Int. J. Mol. Sci. 2021, 22, 1241. [Google Scholar] [CrossRef] [PubMed]
- Karmakar, J.; Mandal, C. Interplay Between Sialic Acids, Siglec-E, and Neu1 Regulates MyD88- and TRIF-Dependent Pathways for TLR4-Activation During Leishmania Donovani Infection. Front. Immunol. 2021, 12, 626110. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Lu, X.; Tao, K.; Shi, L.; Li, W.; Wang, G.; Wu, K. Siglec-10 Is Associated with Survival and Natural Killer Cell Dysfunction in Hepatocellular Carcinoma. J. Surg. Res. 2015, 194, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Bandala-Sanchez, E.; Bediaga, N.G.; Naselli, G.; Neale, A.M.; Harrison, L.C. Siglec-10 Expression Is up-Regulated in Activated Human CD4+ T Cells. Hum. Immunol. 2020, 81, 101–104. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Yu, M.; Guo, L.; Zhang, B.; Liu, S.; Zhang, W.; Zhou, B.; Yan, J.; Ma, Q.; Yang, Z.; et al. Tumor Derived SIGLEC Family Genes May Play Roles in Tumor Genesis, Progression, and Immune Microenvironment Regulation. Front. Oncol. 2020, 10, 586820. [Google Scholar] [CrossRef] [PubMed]
- Kałuża, A.; Szczykutowicz, J.; Ferens-Sieczkowska, M. Glycosylation: Rising Potential for Prostate Cancer Evaluation. Cancers 2021, 13, 3726. [Google Scholar] [CrossRef]
- Munkley, J. The Role of Sialyl-Tn in Cancer. Int. J. Mol. Sci. 2016, 17, 275. [Google Scholar] [CrossRef]
- Diniz, F.; Lamas, S.; Osório, H.; Aguiar, P.; Freitas, D.; Gärtner, F.; Sarmento, B.; Reis, C.A.; Gomes, J. Nanoparticles Targeting Sialyl-Tn for Efficient Tyrosine Kinase Inhibitor Delivery in Gastric Cancer. Acta Biomater. 2023, 170, 142–154. [Google Scholar] [CrossRef]
- Peixoto, A.; Relvas-Santos, M.; Azevedo, R.; Santos, L.L.; Ferreira, J.A. Protein Glycosylation and Tumor Microenvironment Alterations Driving Cancer Hallmarks. Front. Oncol. 2019, 9, 380. [Google Scholar] [CrossRef]
- Dobie, C.; Skropeta, D. Insights into the Role of Sialylation in Cancer Progression and Metastasis. Br. J. Cancer 2021, 124, 76–90. [Google Scholar] [CrossRef]
- Cassetta, L.; Fragkogianni, S.; Sims, A.H.; Swierczak, A.; Forrester, L.M.; Zhang, H.; Soong, D.Y.H.; Cotechini, T.; Anur, P.; Lin, E.Y.; et al. Human Tumor-Associated Macrophage and Monocyte Transcriptional Landscapes Reveal Cancer-Specific Reprogramming, Biomarkers, and Therapeutic Targets. Cancer Cell 2019, 35, 588–602.e10. [Google Scholar] [CrossRef]
- Stanczak, M.A.; Siddiqui, S.S.; Trefny, M.P.; Thommen, D.S.; Boligan, K.F.; von Gunten, S.; Tzankov, A.; Tietze, L.; Lardinois, D.; Heinzelmann-Schwarz, V.; et al. Self-Associated Molecular Patterns Mediate Cancer Immune Evasion by Engaging Siglecs on T Cells. J. Clin. Investig. 2018, 128, 4912–4923. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Li, Z.; Fu, L.; Yan, M.; Wang, Y.; Yu, J.; Wu, J. PD-1 Suppresses the Osteogenic and Odontogenic Differentiation of Stem Cells from Dental Apical Papilla via Targeting SHP2/NF-κB Axis. Stem Cells 2022, 40, 763–777. [Google Scholar] [CrossRef]
- Fu, Q.; Sun, Y.; Tao, Y.; Piao, H.; Wang, X.; Luan, X.; Du, M.; Li, D. Involvement of the JAK-STAT Pathway in Collagen Regulation of Decidual NK Cells. Am. J. Reprod. Immunol. 2017, 78, e12769. [Google Scholar] [CrossRef]
- Singh, V.K.; Khan, A.; Xu, Y.; Mai, S.; Zhang, L.; Mishra, A.; Restrepo, B.I.; Pan, P.-Y.; Chen, S.-H.; Jagannath, C. Antibody-Mediated LILRB2-Receptor Antagonism Induces Human Myeloid-Derived Suppressor Cells to Kill Mycobacterium Tuberculosis. Front. Immunol. 2022, 13, 865503. [Google Scholar] [CrossRef]
- Takagane, K.; Umakoshi, M.; Itoh, G.; Kuriyama, S.; Goto, A.; Tanaka, M. SKAP2 Suppresses Inflammation-Mediated Tumorigenesis by Regulating SHP-1 and SHP-2. Oncogene 2022, 41, 1087–1099. [Google Scholar] [CrossRef]
- He, X.; Xiao, J.; Li, Z.; Ye, M.; Lin, J.; Liu, Z.; Liang, Y.; Dai, H.; Jing, R.; Lin, F. Inhibition of PD-1 Alters the SHP1/2-PI3K/Akt Axis to Decrease M1 Polarization of Alveolar Macrophages in Lung Ischemia-Reperfusion Injury. Inflammation 2023, 46, 639–654. [Google Scholar] [CrossRef]
- Chemnitz, J.M.; Parry, R.V.; Nichols, K.E.; June, C.H.; Riley, J.L. SHP-1 and SHP-2 Associate with Immunoreceptor Tyrosine-Based Switch Motif of Programmed Death 1 upon Primary Human T Cell Stimulation, but Only Receptor Ligation Prevents T Cell Activation. J. Immunol. 2004, 173, 945–954. [Google Scholar] [CrossRef] [PubMed]
- van de Wall, S.; Santegoets, K.C.M.; van Houtum, E.J.H.; Büll, C.; Adema, G.J. Sialoglycans and Siglecs Can Shape the Tumor Immune Microenvironment. Trends Immunol. 2020, 41, 274–285. [Google Scholar] [CrossRef] [PubMed]
- Jandus, C.; Boligan, K.F.; Chijioke, O.; Liu, H.; Dahlhaus, M.; Démoulins, T.; Schneider, C.; Wehrli, M.; Hunger, R.E.; Baerlocher, G.M.; et al. Interactions between Siglec-7/9 Receptors and Ligands Influence NK Cell-Dependent Tumor Immunosurveillance. J. Clin. Investig. 2014, 124, 1810–1820. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, R.; Hintzen, G.; Kemper, A.; Beul, K.; Kempf, S.; Behrens, G.; Sykora, K.W.; Schmidt, R.E. CD56bright Cells Differ in Their KIR Repertoire and Cytotoxic Features from CD56dim NK Cells. Eur. J. Immunol. 2001, 31, 3121–3127. [Google Scholar] [CrossRef] [PubMed]
- Nagler, A.; Lanier, L.L.; Cwirla, S.; Phillips, J.H. Comparative Studies of Human FcRIII-Positive and Negative Natural Killer Cells. J. Immunol. 1989, 143, 3183–3191. [Google Scholar] [CrossRef] [PubMed]
- Nicoll, G.; Ni, J.; Liu, D.; Klenerman, P.; Munday, J.; Dubock, S.; Mattei, M.G.; Crocker, P.R. Identification and Characterization of a Novel Siglec, Siglec-7, Expressed by Human Natural Killer Cells and Monocytes. J. Biol. Chem. 1999, 274, 34089–34095. [Google Scholar] [CrossRef] [PubMed]
- Bordoloi, D.; Kulkarni, A.J.; Adeniji, O.S.; Pampena, M.B.; Bhojnagarwala, P.S.; Zhao, S.; Ionescu, C.; Perales-Puchalt, A.; Parzych, E.M.; Zhu, X.; et al. Siglec-7 Glyco-Immune Binding mAbs or NK Cell Engager Biologics Induce Potent Antitumor Immunity against Ovarian Cancers. Sci. Adv. 2023, 9, eadh4379. [Google Scholar] [CrossRef]
- Belisle, J.A.; Horibata, S.; Jennifer, G.A.A.; Petrie, S.; Kapur, A.; André, S.; Gabius, H.-J.; Rancourt, C.; Connor, J.; Paulson, J.C.; et al. Identification of Siglec-9 as the Receptor for MUC16 on Human NK Cells, B Cells, and Monocytes. Mol. Cancer 2010, 9, 118. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.; Yu, C.; Rodrigues, E.; Shi, Y.; Chen, H.; Wang, P.; Chapla, D.G.; Gao, T.; Zhuang, R.; Moremen, K.W.; et al. Modulation of Siglec-7 Signaling Via In Situ-Created High-Affinity Cis-Ligands. ACS Cent. Sci. 2021, 7, 1338–1346. [Google Scholar] [CrossRef]
- Xiao, H.; Woods, E.C.; Vukojicic, P.; Bertozzi, C.R. Precision Glycocalyx Editing as a Strategy for Cancer Immunotherapy. Proc. Natl. Acad. Sci. USA 2016, 113, 10304–10309. [Google Scholar] [CrossRef]
- Daly, J.; Sarkar, S.; Natoni, A.; Stark, J.C.; Riley, N.M.; Bertozzi, C.R.; Carlsten, M.; O’Dwyer, M.E. Targeting Hypersialylation in Multiple Myeloma Represents a Novel Approach to Enhance NK Cell-Mediated Tumor Responses. Blood Adv. 2022, 6, 3352–3366. [Google Scholar] [CrossRef]
- Fong, J.J.; Tsai, C.-M.; Saha, S.; Nizet, V.; Varki, A.; Bui, J.D. Siglec-7 Engagement by GBS β-Protein Suppresses Pyroptotic Cell Death of Natural Killer Cells. Proc. Natl. Acad. Sci. USA 2018, 115, 10410–10415. [Google Scholar] [CrossRef]
- Meyer, S.J.; Linder, A.T.; Brandl, C.; Nitschke, L. B Cell Siglecs-News on Signaling and Its Interplay With Ligand Binding. Front. Immunol. 2018, 9, 2820. [Google Scholar] [CrossRef] [PubMed]
- Müller, J.; Nitschke, L. The Role of CD22 and Siglec-G in B-Cell Tolerance and Autoimmune Disease. Nat. Rev. Rheumatol. 2014, 10, 422–428. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, A.; Kerr, S.; Jellusova, J.; Zhang, J.; Weisel, F.; Wellmann, U.; Winkler, T.H.; Kneitz, B.; Crocker, P.R.; Nitschke, L. Siglec-G Is a B1 Cell-Inhibitory Receptor That Controls Expansion and Calcium Signaling of the B1 Cell Population. Nat. Immunol. 2007, 8, 695–704. [Google Scholar] [CrossRef] [PubMed]
- Brzezicka, K.A.; Arlian, B.M.; Wang, S.; Olmer, M.; Lotz, M.; Paulson, J.C. Suppression of Autoimmune Rheumatoid Arthritis with Hybrid Nanoparticles That Induce B and T Cell Tolerance to Self-Antigen. ACS Nano 2022, 16, 20206–20221. [Google Scholar] [CrossRef] [PubMed]
- Wong, K.L.; Li, Z.; Ma, F.; Wang, D.; Song, N.; Chong, C.H.; Luk, K.K.; Leung, S.O. SM03, an Anti-CD22 Antibody, Converts Cis-to-Trans Ligand Binding of CD22 against A2,6-Linked Sialic Acid Glycans and Immunomodulates Systemic Autoimmune Diseases. J. Immunol. 2022, 208, 2726–2737. [Google Scholar] [CrossRef]
- Liu, S.; Deng, B.; Yin, Z.; Lin, Y.; An, L.; Liu, D.; Pan, J.; Yu, X.; Chen, B.; Wu, T.; et al. Combination of CD19 and CD22 CAR-T Cell Therapy in Relapsed B-Cell Acute Lymphoblastic Leukemia after Allogeneic Transplantation. Am. J. Hematol. 2021, 96, 671–679. [Google Scholar] [CrossRef]
- Haas, Q.; Boligan, K.F.; Jandus, C.; Schneider, C.; Simillion, C.; Stanczak, M.A.; Haubitz, M.; Seyed Jafari, S.M.; Zippelius, A.; Baerlocher, G.M.; et al. Siglec-9 Regulates an Effector Memory CD8+ T-Cell Subset That Congregates in the Melanoma Tumor Microenvironment. Cancer Immunol. Res. 2019, 7, 707–718. [Google Scholar] [CrossRef]
- Bärenwaldt, A.; Läubli, H. The Sialoglycan-Siglec Glyco-Immune Checkpoint—A Target for Improving Innate and Adaptive Anti-Cancer Immunity. Expert Opin. Ther. Targets 2019, 23, 839–853. [Google Scholar] [CrossRef]
- Haas, Q.; Markov, N.; Muerner, L.; Rubino, V.; Benjak, A.; Haubitz, M.; Baerlocher, G.M.; Ng, C.K.Y.; Münz, C.; Riether, C.; et al. Siglec-7 Represents a Glyco-Immune Checkpoint for Non-Exhausted Effector Memory CD8+ T Cells with High Functional and Metabolic Capacities. Front. Immunol. 2022, 13, 996746. [Google Scholar] [CrossRef]
- Mei, Y.; Wang, X.; Zhang, J.; Liu, D.; He, J.; Huang, C.; Liao, J.; Wang, Y.; Feng, Y.; Li, H.; et al. Siglec-9 Acts as an Immune-Checkpoint Molecule on Macrophages in Glioblastoma, Restricting T-Cell Priming and Immunotherapy Response. Nat. Cancer 2023, 4, 1273–1291. [Google Scholar] [CrossRef]
- Fan, T.; Liao, Q.; Zhao, Y.; Dai, H.; Song, S.; He, T.; Wang, Z.; Huang, J.; Zeng, Z.; Guo, H.; et al. Sialylated IgG in Epithelial Cancers Inhibits Antitumor Function of T Cells via Siglec-7. Cancer Sci. 2023, 114, 370–383. [Google Scholar] [CrossRef] [PubMed]
- Bandala-Sanchez, E.; Zhang, Y.; Reinwald, S.; Dromey, J.A.; Lee, B.-H.; Qian, J.; Böhmer, R.M.; Harrison, L.C. T Cell Regulation Mediated by Interaction of Soluble CD52 with the Inhibitory Receptor Siglec-10. Nat. Immunol. 2013, 14, 741–748. [Google Scholar] [CrossRef] [PubMed]
- Sanmamed, M.F.; Chen, L. A Paradigm Shift in Cancer Immunotherapy: From Enhancement to Normalization. Cell 2019, 176, 677. [Google Scholar] [CrossRef]
- Cyr, M.G.; Mhibik, M.; Qi, J.; Peng, H.; Chang, J.; Gaglione, E.M.; Eik, D.; Herrick, J.; Venables, T.; Novick, S.J.; et al. Patient-Derived Siglec-6-Targeting Antibodies Engineered for T-Cell Recruitment Have Potential Therapeutic Utility in Chronic Lymphocytic Leukemia. J. Immunother. Cancer 2022, 10, e004850. [Google Scholar] [CrossRef]
- Zhang, M.; Angata, T.; Cho, J.Y.; Miller, M.; Broide, D.H.; Varki, A. Defining the in Vivo Function of Siglec-F, a CD33-Related Siglec Expressed on Mouse Eosinophils. Blood 2007, 109, 4280–4287. [Google Scholar] [CrossRef] [PubMed]
- Ding, C.; Liu, Y.; Wang, Y.; Park, B.K.; Wang, C.-Y.; Zheng, P.; Liu, Y. Siglecg Limits the Size of B1a B Cell Lineage by Down-Regulating NFkappaB Activation. PLoS ONE 2007, 2, e997. [Google Scholar] [CrossRef] [PubMed]
- Rempel, H.; Calosing, C.; Sun, B.; Pulliam, L. Sialoadhesin Expressed on IFN-Induced Monocytes Binds HIV-1 and Enhances Infectivity. PLoS ONE 2008, 3, e1967. [Google Scholar] [CrossRef]
- Hartnell, A.; Steel, J.; Turley, H.; Jones, M.; Jackson, D.G.; Crocker, P.R. Characterization of Human Sialoadhesin, a Sialic Acid Binding Receptor Expressed by Resident and Inflammatory Macrophage Populations. Blood 2001, 97, 288–296. [Google Scholar] [CrossRef]
- Angata, T.; Kerr, S.C.; Greaves, D.R.; Varki, N.M.; Crocker, P.R.; Varki, A. Cloning and Characterization of Human Siglec-11. A Recently Evolved Signaling Molecule That Can Interact with SHP-1 and SHP-2 and Is Expressed by Tissue Macrophages, Including Brain Microglia. J. Biol. Chem. 2002, 277, 24466–24474. [Google Scholar] [CrossRef]
- Cao, H.; Lakner, U.; de Bono, B.; Traherne, J.A.; Trowsdale, J.; Barrow, A.D. SIGLEC16 Encodes a DAP12-Associated Receptor Expressed in Macrophages That Evolved from Its Inhibitory Counterpart SIGLEC11 and Has Functional and Non-Functional Alleles in Humans. Eur. J. Immunol. 2008, 38, 2303–2315. [Google Scholar] [CrossRef]
- Eskandari-Sedighi, G.; Jung, J.; Macauley, M.S. CD33 Isoforms in Microglia and Alzheimer’s Disease: Friend and Foe. Mol. Aspects Med. 2023, 90, 101111. [Google Scholar] [CrossRef] [PubMed]
- Rogler, G.; Hausmann, M.; Vogl, D.; Aschenbrenner, E.; Andus, T.; Falk, W.; Andreesen, R.; Schölmerich, J.; Gross, V. Isolation and Phenotypic Characterization of Colonic Macrophages. Clin. Exp. Immunol. 1998, 112, 205–215. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Sun, J.; Liu, L.N.; Flies, D.B.; Nie, X.; Toki, M.; Zhang, J.; Song, C.; Zarr, M.; Zhou, X.; et al. Siglec-15 as an Immune Suppressor and Potential Target for Normalization Cancer Immunotherapy. Nat. Med. 2019, 25, 656–666. [Google Scholar] [CrossRef] [PubMed]
- Xiao, X.; Peng, Y.; Wang, Z.; Zhang, L.; Yang, T.; Sun, Y.; Chen, Y.; Zhang, W.; Chang, X.; Huang, W.; et al. A Novel Immune Checkpoint Siglec-15 Antibody Inhibits LUAD by Modulating Mφ Polarization in TME. Pharmacol. Res. 2022, 181, 106269. [Google Scholar] [CrossRef] [PubMed]
- Stanczak, M.A.; Rodrigues Mantuano, N.; Kirchhammer, N.; Sanin, D.E.; Jacob, F.; Coelho, R.; Everest-Dass, A.V.; Wang, J.; Trefny, M.P.; Monaco, G.; et al. Targeting Cancer Glycosylation Repolarizes Tumor-Associated Macrophages Allowing Effective Immune Checkpoint Blockade. Sci. Transl. Med. 2022, 14, eabj1270. [Google Scholar] [CrossRef]
- Rodriguez, E.; Boelaars, K.; Brown, K.; Eveline Li, R.J.; Kruijssen, L.; Bruijns, S.C.M.; van Ee, T.; Schetters, S.T.T.; Crommentuijn, M.H.W.; van der Horst, J.C.; et al. Sialic Acids in Pancreatic Cancer Cells Drive Tumour-Associated Macrophage Differentiation via the Siglec Receptors Siglec-7 and Siglec-9. Nat. Commun. 2021, 12, 1270. [Google Scholar] [CrossRef]
- Beatson, R.; Tajadura-Ortega, V.; Achkova, D.; Picco, G.; Tsourouktsoglou, T.-D.; Klausing, S.; Hillier, M.; Maher, J.; Noll, T.; Crocker, P.R.; et al. The Mucin MUC1 Modulates the Tumor Immunological Microenvironment through Engagement of the Lectin Siglec-9. Nat. Immunol. 2016, 17, 1273–1281. [Google Scholar] [CrossRef]
- Beatson, R.; Graham, R.; Grundland Freile, F.; Cozzetto, D.; Kannambath, S.; Pfeifer, E.; Woodman, N.; Owen, J.; Nuamah, R.; Mandel, U.; et al. Cancer-Associated Hypersialylated MUC1 Drives the Differentiation of Human Monocytes into Macrophages with a Pathogenic Phenotype. Commun. Biol. 2020, 3, 644. [Google Scholar] [CrossRef]
- Barkal, A.A.; Brewer, R.E.; Markovic, M.; Kowarsky, M.; Barkal, S.A.; Zaro, B.W.; Krishnan, V.; Hatakeyama, J.; Dorigo, O.; Barkal, L.J.; et al. CD24 Signalling through Macrophage Siglec-10 Is a Target for Cancer Immunotherapy. Nature 2019, 572, 392–396. [Google Scholar] [CrossRef]
- Li, W.; Wang, F.; Guo, R.; Bian, Z.; Song, Y. Targeting Macrophages in Hematological Malignancies: Recent Advances and Future Directions. J. Hematol. Oncol. 2022, 15, 110. [Google Scholar] [CrossRef]
- Wculek, S.K.; Cueto, F.J.; Mujal, A.M.; Melero, I.; Krummel, M.F.; Sancho, D. Dendritic Cells in Cancer Immunology and Immunotherapy. Nat. Rev. Immunol. 2020, 20, 7–24. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Guo, Z.; Liu, Y.; Li, X.; Zhang, Q.; Xu, X.; Gu, Y.; Zhang, Y.; Zhao, D.; Cao, X. The Lectin Siglec-G Inhibits Dendritic Cell Cross-Presentation by Impairing MHC Class I-Peptide Complex Formation. Nat. Immunol. 2016, 17, 1167–1175. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Manni, M.; Bärenwaldt, A.; Wieboldt, R.; Kirchhammer, N.; Ivanek, R.; Stanczak, M.; Zippelius, A.; König, D.; Rodrigues Manutano, N.; et al. Siglec Receptors Modulate Dendritic Cell Activation and Antigen Presentation to T Cells in Cancer. Front. Cell Dev. Biol. 2022, 10, 828916. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Zhang, B.; Wang, X.; Zeng, Z.; Huang, Z.; Zhang, L.; Wei, F.; Ren, X.; Yang, L. Expression Signature, Prognosis Value, and Immune Characteristics of Siglec-15 Identified by Pan-Cancer Analysis. Oncoimmunology 2020, 9, 1807291. [Google Scholar] [CrossRef] [PubMed]
- Ruffin, N.; Gea-Mallorquí, E.; Brouiller, F.; Jouve, M.; Silvin, A.; See, P.; Dutertre, C.-A.; Ginhoux, F.; Benaroch, P. Constitutive Siglec-1 Expression Confers Susceptibility to HIV-1 Infection of Human Dendritic Cell Precursors. Proc. Natl. Acad. Sci. USA 2019, 116, 21685–21693. [Google Scholar] [CrossRef] [PubMed]
- Perez-Zsolt, D.; Cantero-Pérez, J.; Erkizia, I.; Benet, S.; Pino, M.; Serra-Peinado, C.; Hernández-Gallego, A.; Castellví, J.; Tapia, G.; Arnau-Saz, V.; et al. Dendritic Cells From the Cervical Mucosa Capture and Transfer HIV-1 via Siglec-1. Front. Immunol. 2019, 10, 825. [Google Scholar] [CrossRef]
- Shafi, S.; Aung, T.N.; Robbins, C.; Zugazagoitia, J.; Vathiotis, I.; Gavrielatou, N.; Yaghoobi, V.; Fernandez, A.; Niu, S.; Liu, L.N.; et al. Development of an Immunohistochemical Assay for Siglec-15. Lab. Investig. 2022, 102, 771–778. [Google Scholar] [CrossRef]
- Clowse, M.E.B.; Wallace, D.J.; Furie, R.A.; Petri, M.A.; Pike, M.C.; Leszczyński, P.; Neuwelt, C.M.; Hobbs, K.; Keiserman, M.; Duca, L.; et al. Efficacy and Safety of Epratuzumab in Moderately to Severely Active Systemic Lupus Erythematosus: Results From Two Phase III Randomized, Double-Blind, Placebo-Controlled Trials. Arthritis Rheumatol. 2017, 69, 362–375. [Google Scholar] [CrossRef]
- Stelljes, M.; Raffel, S.; Alakel, N.; Wäsch, R.; Kondakci, M.; Scholl, S.; Rank, A.; Hänel, M.; Spriewald, B.; Hanoun, M.; et al. Inotuzumab Ozogamicin as Induction Therapy for Patients Older Than 55 Years With Philadelphia Chromosome-Negative B-Precursor ALL. J. Clin. Oncol. 2023, JCO2300546. [Google Scholar] [CrossRef]
- Bachanova, V.; Frankel, A.E.; Cao, Q.; Lewis, D.; Grzywacz, B.; Verneris, M.R.; Ustun, C.; Lazaryan, A.; McClune, B.; Warlick, E.D.; et al. Phase I Study of a Bispecific Ligand-Directed Toxin Targeting CD22 and CD19 (DT2219) for Refractory B-Cell Malignancies. Clin. Cancer Res. 2015, 21, 1267–1272. [Google Scholar] [CrossRef]
- Wang, T.; Tang, Y.; Cai, J.; Wan, X.; Hu, S.; Lu, X.; Xie, Z.; Qiao, X.; Jiang, H.; Shao, J.; et al. Coadministration of CD19- and CD22-Directed Chimeric Antigen Receptor T-Cell Therapy in Childhood B-Cell Acute Lymphoblastic Leukemia: A Single-Arm, Multicenter, Phase II Trial. J. Clin. Oncol. 2023, 41, 1670–1683. [Google Scholar] [CrossRef] [PubMed]
- Siglec-15: An Attractive Immunotherapy Target. Cancer Discov. 2020, 10, 7–8. [CrossRef] [PubMed]
- Qin, H.; Yang, L.; Chukinas, J.A.; Shah, N.; Tarun, S.; Pouzolles, M.; Chien, C.D.; Niswander, L.M.; Welch, A.R.; Taylor, N.; et al. Systematic Preclinical Evaluation of CD33-Directed Chimeric Antigen Receptor T Cell Immunotherapy for Acute Myeloid Leukemia Defines Optimized Construct Design. J. Immunother. Cancer 2021, 9, e003149. [Google Scholar] [CrossRef] [PubMed]
- Borthakur, G.; Ravandi, F.; Patel, K.; Wang, X.; Kadia, T.; DiNardo, C.; Garcia-Manero, G.; Pemmaraju, N.; Jabbour, E.J.; Takahashi, K.; et al. Retrospective Comparison of Survival and Responses to Fludarabine, Cytarabine, GCSF (FLAG) in Combination with Gemtuzumab Ozogamicin (GO) or Idarubicin (IDA) in Patients with Newly Diagnosed Core Binding Factor (CBF) Acute Myelogenous Leukemia: MD Anderson Experience in 174 Patients. Am. J. Hematol. 2022, 97, 1427–1434. [Google Scholar] [CrossRef] [PubMed]
- Aigner, M.; Feulner, J.; Schaffer, S.; Kischel, R.; Kufer, P.; Schneider, K.; Henn, A.; Rattel, B.; Friedrich, M.; Baeuerle, P.A.; et al. T Lymphocytes Can Be Effectively Recruited for Ex Vivo and in Vivo Lysis of AML Blasts by a Novel CD33/CD3-Bispecific BiTE Antibody Construct. Leukemia 2013, 27, 1107–1115. [Google Scholar] [CrossRef]
- Dao, T.; Xiong, G.; Mun, S.S.; Meyerberg, J.; Korontsvit, T.; Xiang, J.; Cui, Z.; Chang, A.Y.; Jarvis, C.A.; Cai, W.; et al. Dual-Receptor T Cell Platform with Ab-TCR and Costimulatory Receptor Achieves Specificity and Potency against AML. Blood 2023, in press. [Google Scholar] [CrossRef]
- Jetani, H.; Navarro-Bailón, A.; Maucher, M.; Frenz, S.; Verbruggen, C.; Yeguas, A.; Vidriales, M.B.; González, M.; Rial Saborido, J.; Kraus, S.; et al. Siglec-6 Is a Novel Target for CAR T-Cell Therapy in Acute Myeloid Leukemia. Blood 2021, 138, 1830–1842. [Google Scholar] [CrossRef]
- Liu, J.; Zheng, X.; Pang, X.; Li, L.; Wang, J.; Yang, C.; Du, G. Ganglioside GD3 Synthase (GD3S), a Novel Cancer Drug Target. Acta Pharm. Sin. B 2018, 8, 713–720. [Google Scholar] [CrossRef]
- Meril, S.; Harush, O.; Reboh, Y.; Matikhina, T.; Barliya, T.; Cohen, C.J. Targeting Glycosylated Antigens on Cancer Cells Using Siglec-7/9-Based CAR T-Cells. Mol. Carcinog. 2020, 59, 713–723. [Google Scholar] [CrossRef]
- Fiedler, W.; DeDosso, S.; Cresta, S.; Weidmann, J.; Tessari, A.; Salzberg, M.; Dietrich, B.; Baumeister, H.; Goletz, S.; Gianni, L.; et al. A Phase I Study of PankoMab-GEX, a Humanised Glyco-Optimised Monoclonal Antibody to a Novel Tumour-Specific MUC1 Glycopeptide Epitope in Patients with Advanced Carcinomas. Eur. J. Cancer 2016, 63, 55–63. [Google Scholar] [CrossRef]
- Aggarwal, N.; Manley, A.L.; Shalhoub, R.; Durrani, J.; Rios, O.; Lotter, J.; Patel, B.A.; Wu, C.O.; Young, N.S.; Groarke, E.M. Alemtuzumab in Relapsed Immune Severe Aplastic Anemia: Long-Term Results of a Phase II Study. Am. J. Hematol. 2023, 98, 932–939. [Google Scholar] [CrossRef] [PubMed]
- Hudak, J.E.; Canham, S.M.; Bertozzi, C.R. Glycocalyx Engineering Reveals a Siglec-Based Mechanism for NK Cell Immunoevasion. Nat. Chem. Biol. 2014, 10, 69–75. [Google Scholar] [CrossRef]
- Ibarlucea-Benitez, I.; Weitzenfeld, P.; Smith, P.; Ravetch, J.V. Siglecs-7/9 Function as Inhibitory Immune Checkpoints in Vivo and Can Be Targeted to Enhance Therapeutic Antitumor Immunity. Proc. Natl. Acad. Sci. USA 2021, 118, e2107424118. [Google Scholar] [CrossRef] [PubMed]
- Fraschilla, I.; Pillai, S. Viewing Siglecs through the Lens of Tumor Immunology. Immunol. Rev. 2017, 276, 178–191. [Google Scholar] [CrossRef] [PubMed]
- Murugesan, G.; Weigle, B.; Crocker, P.R. Siglec and Anti-Siglec Therapies. Curr. Opin. Chem. Biol. 2021, 62, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Forgione, R.E.; Di Carluccio, C.; Guzmán-Caldentey, J.; Gaglione, R.; Battista, F.; Chiodo, F.; Manabe, Y.; Arciello, A.; Del Vecchio, P.; Fukase, K.; et al. Unveiling Molecular Recognition of Sialoglycans by Human Siglec-10. iScience 2020, 23, 101231. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.-T.; Huang, Z.-Z.; Chen, Y.-B.; Yao, H.-Y.; Ke, Z.-H.; He, X.-X.; Qiu, M.-J.; Wang, M.-M.; Xiong, Z.-F.; Yang, S.-L. Integrative Analysis of Siglec-15 mRNA in Human Cancers Based on Data Mining. J. Cancer 2020, 11, 2453–2464. [Google Scholar] [CrossRef]
- Takamiya, R.; Ohtsubo, K.; Takamatsu, S.; Taniguchi, N.; Angata, T. The Interaction between Siglec-15 and Tumor-Associated Sialyl-Tn Antigen Enhances TGF-β Secretion from Monocytes/Macrophages through the DAP12-Syk Pathway. Glycobiology 2013, 23, 178–187. [Google Scholar] [CrossRef]
- Sun, J.; Lu, Q.; Sanmamed, M.F.; Wang, J. Siglec-15 as an Emerging Target for Next-Generation Cancer Immunotherapy. Clin. Cancer Res. 2021, 27, 680–688. [Google Scholar] [CrossRef]
- Landolina, N.; Zaffran, I.; Smiljkovic, D.; Serrano-Candelas, E.; Schmiedel, D.; Friedman, S.; Arock, M.; Hartmann, K.; Pikarsky, E.; Mandelboim, O.; et al. Activation of Siglec-7 Results in Inhibition of in Vitro and in Vivo Growth of Human Mast Cell Leukemia Cells. Pharmacol. Res. 2020, 158, 104682. [Google Scholar] [CrossRef]
- Esfandiari, A.; Cassidy, S.; Webster, R.M. Bispecific Antibodies in Oncology. Nat. Rev. Drug Discov. 2022, 21, 411–412. [Google Scholar] [CrossRef] [PubMed]
- van de Donk, N.W.C.J.; Zweegman, S. T-Cell-Engaging Bispecific Antibodies in Cancer. Lancet 2023, 402, 142–158. [Google Scholar] [CrossRef] [PubMed]
- Klupsch, K.; Baeriswyl, V.; Scholz, R.; Dannenberg, J.; Santimaria, R.; Senn, D.; Kage, E.; Zumsteg, A.; Attinger-Toller, I.; von der Bey, U.; et al. COVA4231, a Potent CD3/CD33 Bispecific FynomAb with IgG-like Pharmacokinetics for the Treatment of Acute Myeloid Leukemia. Leukemia 2019, 33, 805–808. [Google Scholar] [CrossRef] [PubMed]
- Thakkar, D.; Paliwal, S.; Dharmadhikari, B.; Guan, S.; Liu, L.; Kar, S.; Tulsian, N.K.; Gruber, J.J.; DiMascio, L.; Paszkiewicz, K.H.; et al. Rationally Targeted Anti-VISTA Antibody That Blockades the C-C’ Loop Region Can Reverse VISTA Immune Suppression and Remodel the Immune Microenvironment to Potently Inhibit Tumor Growth in an Fc Independent Manner. J. Immunother. Cancer 2022, 10, e003382. [Google Scholar] [CrossRef] [PubMed]
- Yu, B.; Liu, D. Gemtuzumab Ozogamicin and Novel Antibody-Drug Conjugates in Clinical Trials for Acute Myeloid Leukemia. Biomark. Res. 2019, 7, 24. [Google Scholar] [CrossRef] [PubMed]
- Abuhay, M.; Kato, J.; Tuscano, E.; Barisone, G.A.; Sidhu, R.S.; O’Donnell, R.T.; Tuscano, J.M. The HB22.7-vcMMAE Antibody-Drug Conjugate Has Efficacy against Non-Hodgkin Lymphoma Mouse Xenografts with Minimal Systemic Toxicity. Cancer Immunol. Immunother. 2016, 65, 1169–1175. [Google Scholar] [CrossRef] [PubMed]
- Short, N.J.; Kantarjian, H. Using Immunotherapy and Novel Trial Designs to Optimise Front-Line Therapy in Adult Acute Lymphoblastic Leukaemia: Breaking with the Traditions of the Past. Lancet Haematol. 2023, 10, e382–e388. [Google Scholar] [CrossRef]
- O’Sullivan, J.A.; Carroll, D.J.; Cao, Y.; Salicru, A.N.; Bochner, B.S. Leveraging Siglec-8 Endocytic Mechanisms to Kill Human Eosinophils and Malignant Mast Cells. J. Allergy Clin. Immunol. 2018, 141, 1774–1785.e7. [Google Scholar] [CrossRef]
- Li, S.; Tao, Z.; Xu, Y.; Liu, J.; An, N.; Wang, Y.; Xing, H.; Tian, Z.; Tang, K.; Liao, X.; et al. CD33-Specific Chimeric Antigen Receptor T Cells with Different Co-Stimulators Showed Potent Anti-Leukemia Efficacy and Different Phenotype. Hum. Gene Ther. 2018, 29, 626–639. [Google Scholar] [CrossRef]
- Pan, J.; Niu, Q.; Deng, B.; Liu, S.; Wu, T.; Gao, Z.; Liu, Z.; Zhang, Y.; Qu, X.; Zhang, Y.; et al. CD22 CAR T-Cell Therapy in Refractory or Relapsed B Acute Lymphoblastic Leukemia. Leukemia 2019, 33, 2854–2866. [Google Scholar] [CrossRef]
- Maude, S.L.; Frey, N.; Shaw, P.A.; Aplenc, R.; Barrett, D.M.; Bunin, N.J.; Chew, A.; Gonzalez, V.E.; Zheng, Z.; Lacey, S.F.; et al. Chimeric Antigen Receptor T Cells for Sustained Remissions in Leukemia. N. Engl. J. Med. 2014, 371, 1507–1517. [Google Scholar] [CrossRef] [PubMed]
- Schuster, S.J.; Bishop, M.R.; Tam, C.S.; Waller, E.K.; Borchmann, P.; McGuirk, J.P.; Jäger, U.; Jaglowski, S.; Andreadis, C.; Westin, J.R.; et al. Tisagenlecleucel in Adult Relapsed or Refractory Diffuse Large B-Cell Lymphoma. N. Engl. J. Med. 2019, 380, 45–56. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Sun, Q.; Liang, X.; Chen, Z.; Zhang, X.; Zhou, X.; Li, M.; Tu, H.; Liu, Y.; Tu, S.; et al. Mechanisms of Relapse After CD19 CAR T-Cell Therapy for Acute Lymphoblastic Leukemia and Its Prevention and Treatment Strategies. Front. Immunol. 2019, 10, 2664. [Google Scholar] [CrossRef] [PubMed]
- Kovalovsky, D.; Yoon, J.H.; Cyr, M.G.; Simon, S.; Voynova, E.; Rader, C.; Wiestner, A.; Alejo, J.; Pittaluga, S.; Gress, R.E. Siglec-6 Is a Target for Chimeric Antigen Receptor T-Cell Treatment of Chronic Lymphocytic Leukemia. Leukemia 2021, 35, 2581–2591. [Google Scholar] [CrossRef]
- Schweizer, A.; Wöhner, M.; Prescher, H.; Brossmer, R.; Nitschke, L. Targeting of CD22-Positive B-Cell Lymphoma Cells by Synthetic Divalent Sialic Acid Analogues. Eur. J. Immunol. 2012, 42, 2792–2802. [Google Scholar] [CrossRef] [PubMed]
- Duan, S.; Koziol-White, C.J.; Jester, W.F.; Smith, S.A.; Nycholat, C.M.; Macauley, M.S.; Panettieri, R.A.; Paulson, J.C. CD33 Recruitment Inhibits IgE-Mediated Anaphylaxis and Desensitizes Mast Cells to Allergen. J. Clin. Investig. 2019, 129, 1387–1401. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.C.; Completo, G.C.; Sigal, D.S.; Crocker, P.R.; Saven, A.; Paulson, J.C. In Vivo Targeting of B-Cell Lymphoma with Glycan Ligands of CD22. Blood 2010, 115, 4778–4786. [Google Scholar] [CrossRef]
- Rillahan, C.D.; Antonopoulos, A.; Lefort, C.T.; Sonon, R.; Azadi, P.; Ley, K.; Dell, A.; Haslam, S.M.; Paulson, J.C. Global Metabolic Inhibitors of Sialyl- and Fucosyltransferases Remodel the Glycome. Nat. Chem. Biol. 2012, 8, 661–668. [Google Scholar] [CrossRef]
- Balneger, N.; Cornelissen, L.A.M.; Wassink, M.; Moons, S.J.; Boltje, T.J.; Bar-Ephraim, Y.E.; Das, K.K.; Søndergaard, J.N.; Büll, C.; Adema, G.J. Sialic Acid Blockade in Dendritic Cells Enhances CD8+ T Cell Responses by Facilitating High-Avidity Interactions. Cell Mol. Life Sci. 2022, 79, 98. [Google Scholar] [CrossRef]
- Büll, C.; Boltje, T.J.; Balneger, N.; Weischer, S.M.; Wassink, M.; van Gemst, J.J.; Bloemendal, V.R.; Boon, L.; van der Vlag, J.; Heise, T.; et al. Sialic Acid Blockade Suppresses Tumor Growth by Enhancing T-Cell-Mediated Tumor Immunity. Cancer Res. 2018, 78, 3574–3588. [Google Scholar] [CrossRef]
- Rimando, J.C.; Chendamarai, E.; Rettig, M.P.; Jayasinghe, R.; Christopher, M.J.; Ritchey, J.K.; Christ, S.; Kim, M.Y.; Bonvini, E.; DiPersio, J.F. Flotetuzumab and Other T-Cell Immunotherapies Upregulate MHC Class II Expression on Acute Myeloid Leukemia Cells. Blood 2023, 141, 1718–1723. [Google Scholar] [CrossRef] [PubMed]
- Walter, R.B. The Role of CD33 as Therapeutic Target in Acute Myeloid Leukemia. Expert Opin. Ther. Targets 2014, 18, 715–718. [Google Scholar] [CrossRef] [PubMed]
- Tannoury, M.; Garnier, D.; Susin, S.A.; Bauvois, B. Current Status of Novel Agents for the Treatment of B Cell Malignancies: What’s Coming Next? Cancers 2022, 14, 6026. [Google Scholar] [CrossRef] [PubMed]
- Spiegel, J.Y.; Patel, S.; Muffly, L.; Hossain, N.M.; Oak, J.; Baird, J.H.; Frank, M.J.; Shiraz, P.; Sahaf, B.; Craig, J.; et al. CAR T Cells with Dual Targeting of CD19 and CD22 in Adult Patients with Recurrent or Refractory B Cell Malignancies: A Phase 1 Trial. Nat. Med. 2021, 27, 1419–1431. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, Y.; Wang, J.; Li, H.; Chen, Y.; Chen, C.; Wang, D.W. Plasma Siglec-5 and CD163 as Novel Biomarkers for Fulminant Myocarditis. Biomedicines 2022, 10, 2941. [Google Scholar] [CrossRef] [PubMed]
- Höppner, J.; Casteleyn, V.; Biesen, R.; Rose, T.; Windisch, W.; Burmester, G.R.; Siegert, E. SIGLEC-1 in Systemic Sclerosis: A Useful Biomarker for Differential Diagnosis. Pharmaceuticals 2022, 15, 1198. [Google Scholar] [CrossRef]
- Zhou, S.; Wang, Y.; Zhang, R.; Zeng, W.; Liu, S.; Liu, S.; Liu, M.; Yang, H.; Xi, M. Association of Sialic Acid-Binding Immunoglobulin-Like Lectin 15 With Phenotypes in Esophageal Squamous Cell Carcinoma in the Setting of Neoadjuvant Chemoradiotherapy. JAMA Netw. Open 2023, 6, e2250965. [Google Scholar] [CrossRef]
- Islam, M.; Arlian, B.M.; Pfrengle, F.; Duan, S.; Smith, S.A.; Paulson, J.C. Suppressing Immune Responses Using Siglec Ligand-Decorated Anti-Receptor Antibodies. J. Am. Chem. Soc. 2022, 144, 9302–9311. [Google Scholar] [CrossRef]
- Albinger, N.; Pfeifer, R.; Nitsche, M.; Mertlitz, S.; Campe, J.; Stein, K.; Kreyenberg, H.; Schubert, R.; Quadflieg, M.; Schneider, D.; et al. Primary CD33-Targeting CAR-NK Cells for the Treatment of Acute Myeloid Leukemia. Blood Cancer J. 2022, 12, 61. [Google Scholar] [CrossRef]
- Jiang, K.-Y.; Qi, L.-L.; Kang, F.-B.; Wang, L. The Intriguing Roles of Siglec Family Members in the Tumor Microenvironment. Biomark. Res. 2022, 10, 22. [Google Scholar] [CrossRef]
- Wu, Q.; Wei, X.; Chen, F.; Huang, M.; Zhang, S.; Zhu, L.; Zhou, L.; Yang, C.; Song, Y. Aptamer-Assisted Blockade of the Immune Suppressor Sialic Acid-Binding Immunoglobulin-Like Lectin-15 for Cancer Immunotherapy. Angew. Chem. Int. Ed. 2023, 62, e202312609. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhang, Q.; Liang, Y.; Xiong, S.; Cai, Y.; Cao, J.; Xu, Y.; Xu, X.; Wu, Y.; Lu, Q.; et al. Nanoparticles (NPs)-Mediated Siglec15 Silencing and Macrophage Repolarization for Enhanced Cancer Immunotherapy. Acta Pharm. Sin. B 2023, 13, 5048–5059. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, E.; Jung, J.; Park, H.; Loo, C.; Soukhtehzari, S.; Kitova, E.N.; Mozaneh, F.; Daskhan, G.; Schmidt, E.N.; Aghanya, V.; et al. A Versatile Soluble Siglec Scaffold for Sensitive and Quantitative Detection of Glycan Ligands. Nat. Commun. 2020, 11, 5091. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Cell Type | Predominant Siglecs | Associated Functions |
|---|---|---|
| NK Cells | Siglec-7, Siglec-9 | Inhibition of NK cell activity; ligands confer protection |
| B Cells | Siglec-2 (CD22), Siglec-G, Siglec-10 | Modulation of B cell differentiation; immune tolerance |
| T Cells | Siglec-7, Siglec-9, Siglec-5, Siglec-14 | Regulation of T cell activity; diversity in pathology-driven expression |
| Monocytes/Macrophages | Siglec-E, Siglec-1, Siglec-10, Siglec-9, Siglec-11, Siglec-3, Siglec-16, Siglec-15, | Tissue-specific profiles; involvement in T cell differentiation |
| Dendritic Cells | Siglec-E, Siglec-G, Siglec-7, Siglec-9, Siglec-10, Siglec-15 | Impact on DC cross-presentation; association with DC maturation |
| Human | Cell Type | Diseases | Medicine |
|---|---|---|---|
| Siglec-1 (CD169/Sialoadhesion) | Macrophages | Cancer SLE Infectious diseases | |
| Siglec-2 (CD22) | B cells | B-cell lymphoma Type 1 diabetes Sepsis RA SLE | Epratuzumab (mAb, agonistic antibody) [89] Inotuzumab ozogamicin (Antibody-based cell depletion) [90] DT2219 (BsAb) [91] CD22-CAR-T (CAR-T) [92] |
| Siglec-4 (MAG) | Myelin-producing cells (Schwann cells and oligodendrocytes) | Stroke Schizophrenia Neuropathy | |
| Silec-15 (CD33L) | Macrophages Dendritic cells | Cancer Osteoporosis | NC318 (mAb, antagonistic antibody) [93] |
| Siglec-3 (CD33) | Myeloid cells | ML Myelodysplastic syndromes Alzheimer’s disease Mast cell-dependent anaphylaxis Systemic mastocytosis | Lintuzumab (mAb, antagonistic antibody) [94] Gentuzumab ozogamicin (antibody-based cell depletion) [95] AMG330 (BsAb) [96] CD33 CAR-T (CAR-T) [97] |
| Siglec-5 (CD170) | Monocytes Neutrophils B cells Tumor-infiltrating T c | Cancer Neutrophil disorders | |
| Siglec-6 (CD327) | Mast cells B cells Dendritic cells | Allergy Trophoblastic diseases Preeclampsia | Siglec 6 CAR-T (CAR-T) [98] |
| Siglec-7 (CD328) | NK cells Monocytes Mast cells Basophils Neutrophils T cells | Alzheimer’s disease Systemic mastocytosis Mast cell leukemia Cancers | Ganglioside GD3 (BsAb) [99] Siglec 7 CAR-T (CAR-T) [100] |
| Siglec-8 | Eosinophils Mast cells Basophils | Allergic asthma Eosinophilic gastric Chronic urticaria | |
| Siglec-9 (CD329) | Macrophages NK cells Monocytes Dendritic cells Neutrophils T cells | Asthma COPD RA Cancer | Gatipotuzumab (mAb, antagonistic antibody) [101] Siglec 9 CAR-T (CAR-T) [100] |
| Siglec-10 (SLG2) | B cells NK cells Monocytes Dendritic cells CD4+ T cells | Sepsis Cancer | Alemtuzumab (mAb, antagonistic antibody) [102] |
| Siglec-11 | Macrophages Microglia B cells | Alzheimer’s disease Acute tissue injury | |
| Siglec-12 | Macrophages | Cancer | |
| Siglec-14 | Monocytes Neutrophils | SLE COPD GBS infection | |
| Siglec-15 | Microglia Macrophages | Schizophrenia |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, H.; Feng, J.; Han, X.; Ying, Y.; Lou, W.; Liu, L.; Zhang, L. The Potential of Siglecs and Sialic Acids as Biomarkers and Therapeutic Targets in Tumor Immunotherapy. Cancers 2024, 16, 289. https://doi.org/10.3390/cancers16020289
Feng H, Feng J, Han X, Ying Y, Lou W, Liu L, Zhang L. The Potential of Siglecs and Sialic Acids as Biomarkers and Therapeutic Targets in Tumor Immunotherapy. Cancers. 2024; 16(2):289. https://doi.org/10.3390/cancers16020289
Chicago/Turabian StyleFeng, Haokang, Jiale Feng, Xu Han, Ying Ying, Wenhui Lou, Liang Liu, and Lei Zhang. 2024. "The Potential of Siglecs and Sialic Acids as Biomarkers and Therapeutic Targets in Tumor Immunotherapy" Cancers 16, no. 2: 289. https://doi.org/10.3390/cancers16020289
APA StyleFeng, H., Feng, J., Han, X., Ying, Y., Lou, W., Liu, L., & Zhang, L. (2024). The Potential of Siglecs and Sialic Acids as Biomarkers and Therapeutic Targets in Tumor Immunotherapy. Cancers, 16(2), 289. https://doi.org/10.3390/cancers16020289