
Investigating the Radiobiological Response to Peptide Receptor Radionuclide Therapy Using Patient-Derived Meningioma Spheroids

 , ,
, ,  and
and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Patient Samples
2.2. Isolation of Tissue Fragments
2.3. Culturing of Meningioma Spheroids
2.4. Radiolabeling
2.5. EBRT and PRRT
2.6. Uptake Assays
2.7. Hematoxylin and Eosin, Immunohistochemical and Immunofluorescent Stainings
2.8. Imaging of DNA Damage Repair Foci
2.9. Antibodies
2.10. Statistical Analyses
3. Results
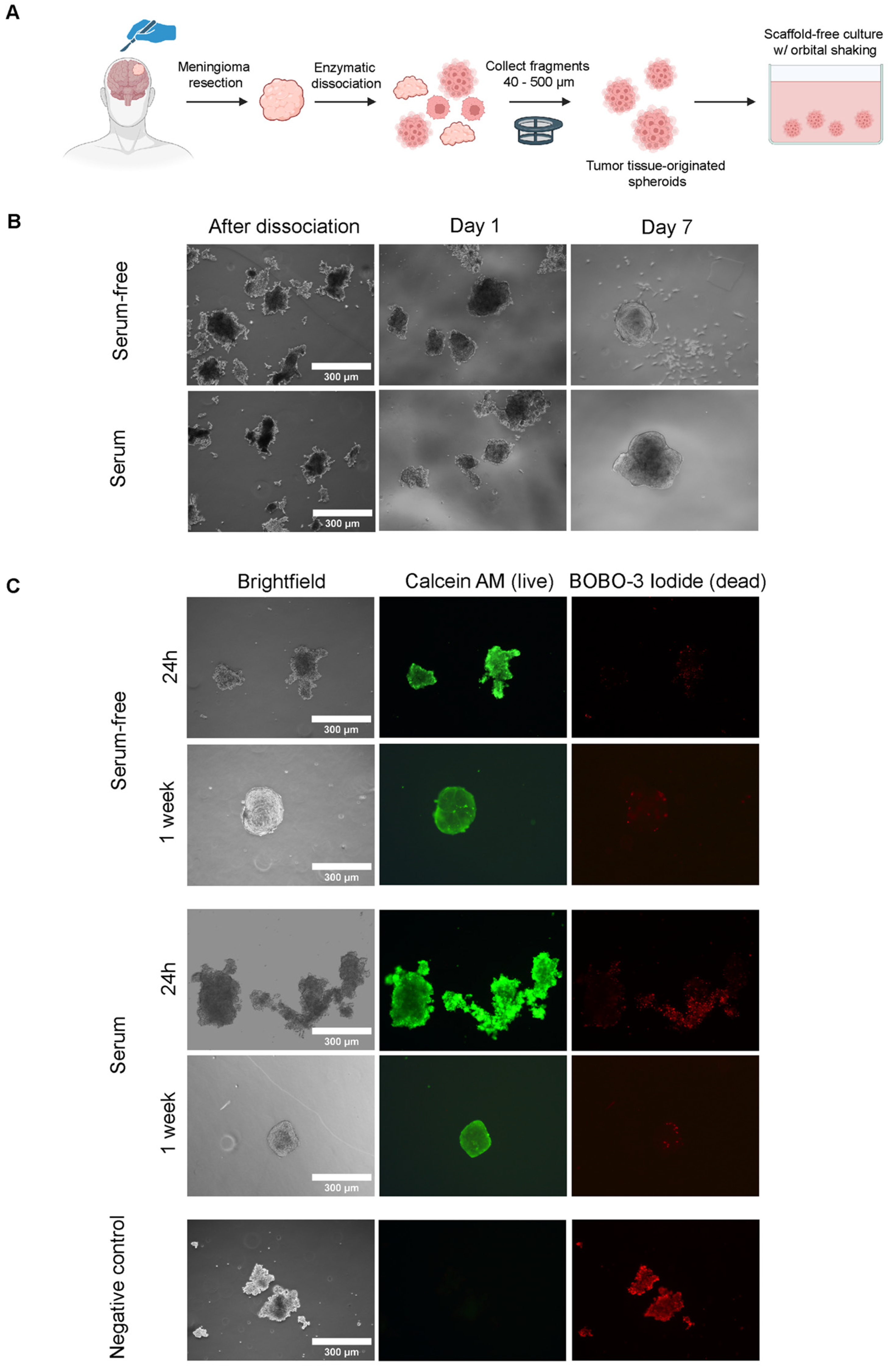
3.1. Establishment of Meningioma Spheroid Cultures
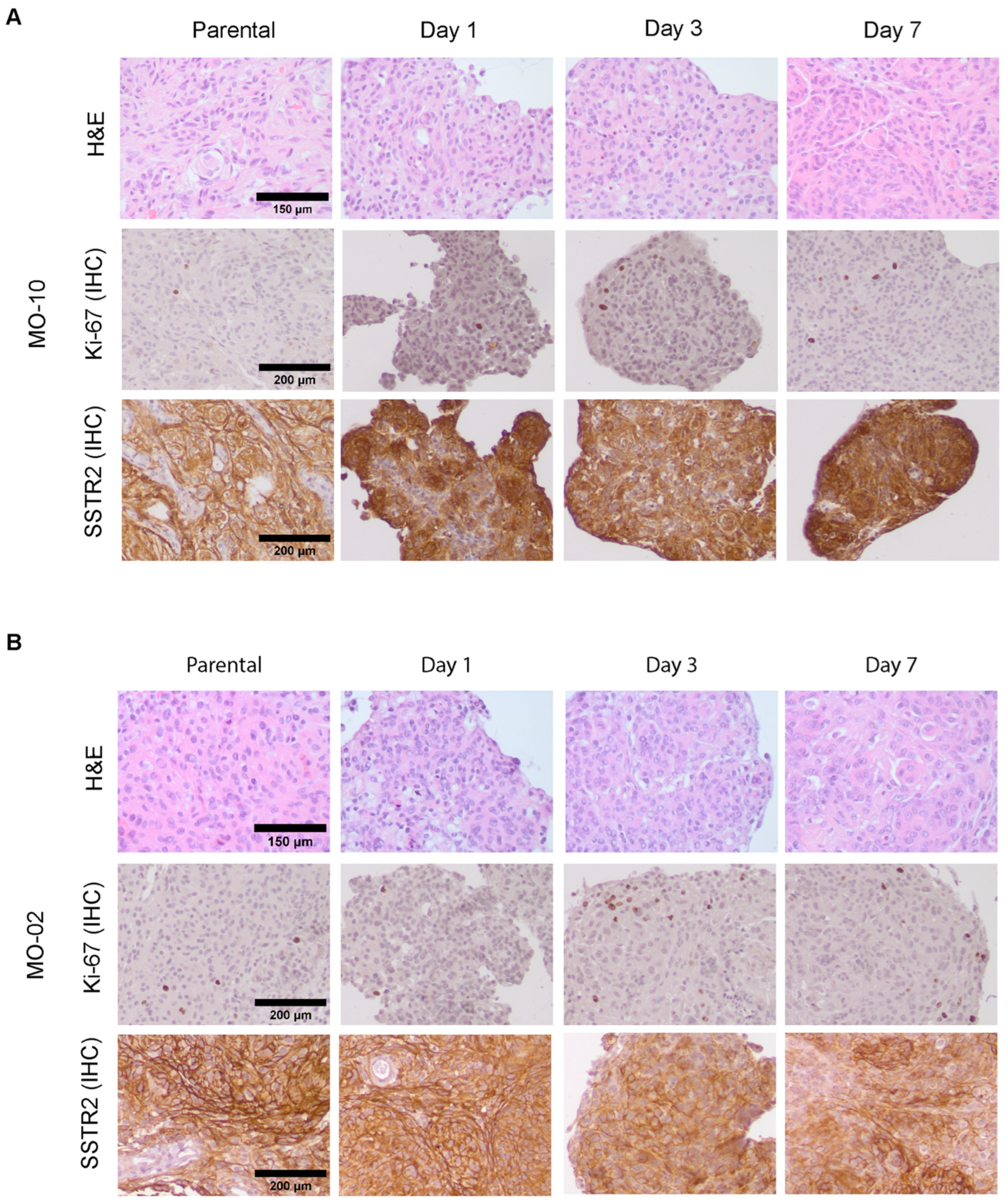
3.2. Meningioma Spheroids Recapitulate Tissue Architecture and Meningioma Characteristics but Show Phenotypic Drift over Time
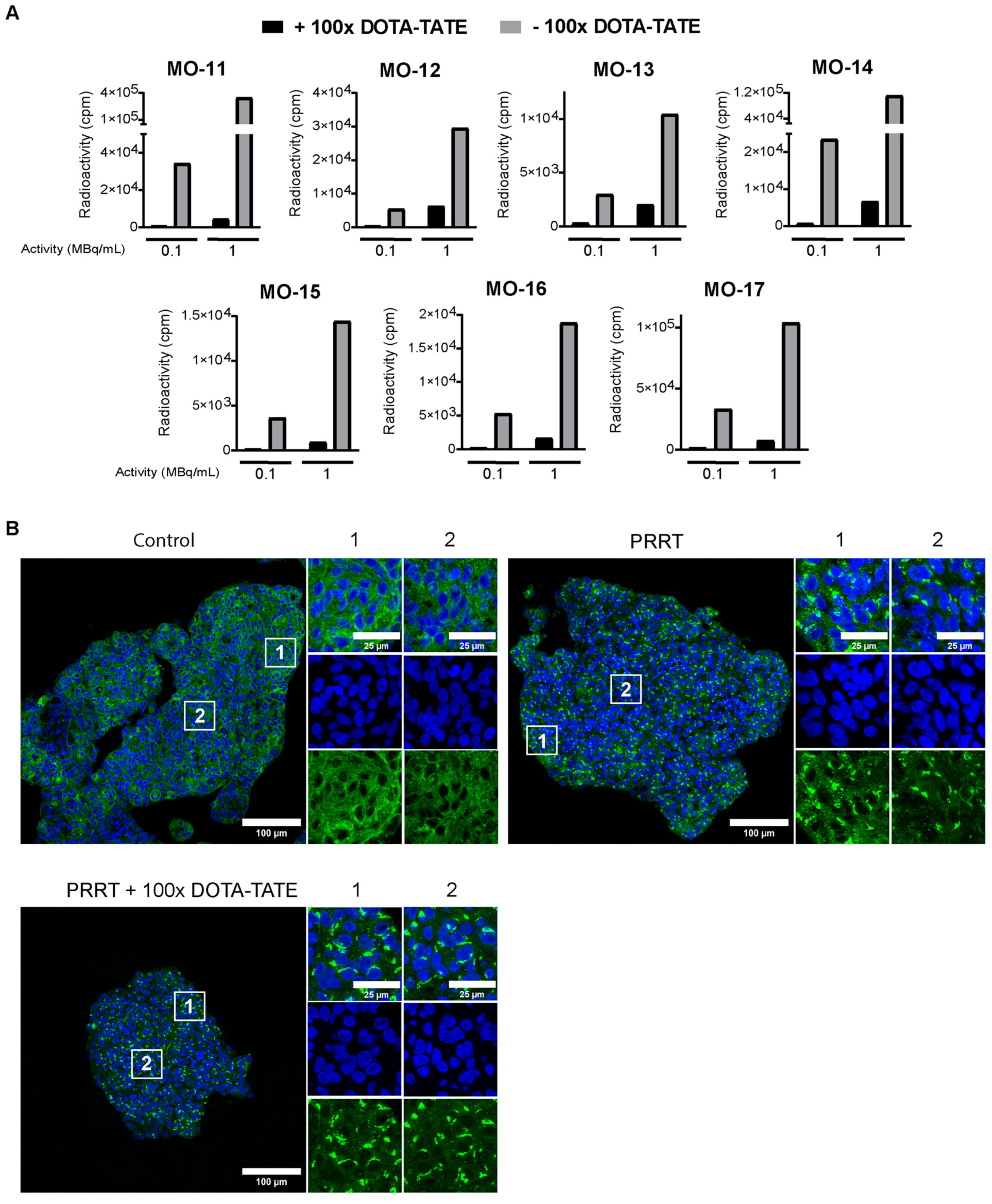
3.3. 177Lu-DOTA-TATE Binds to Meningioma Spheroids via SSTR2
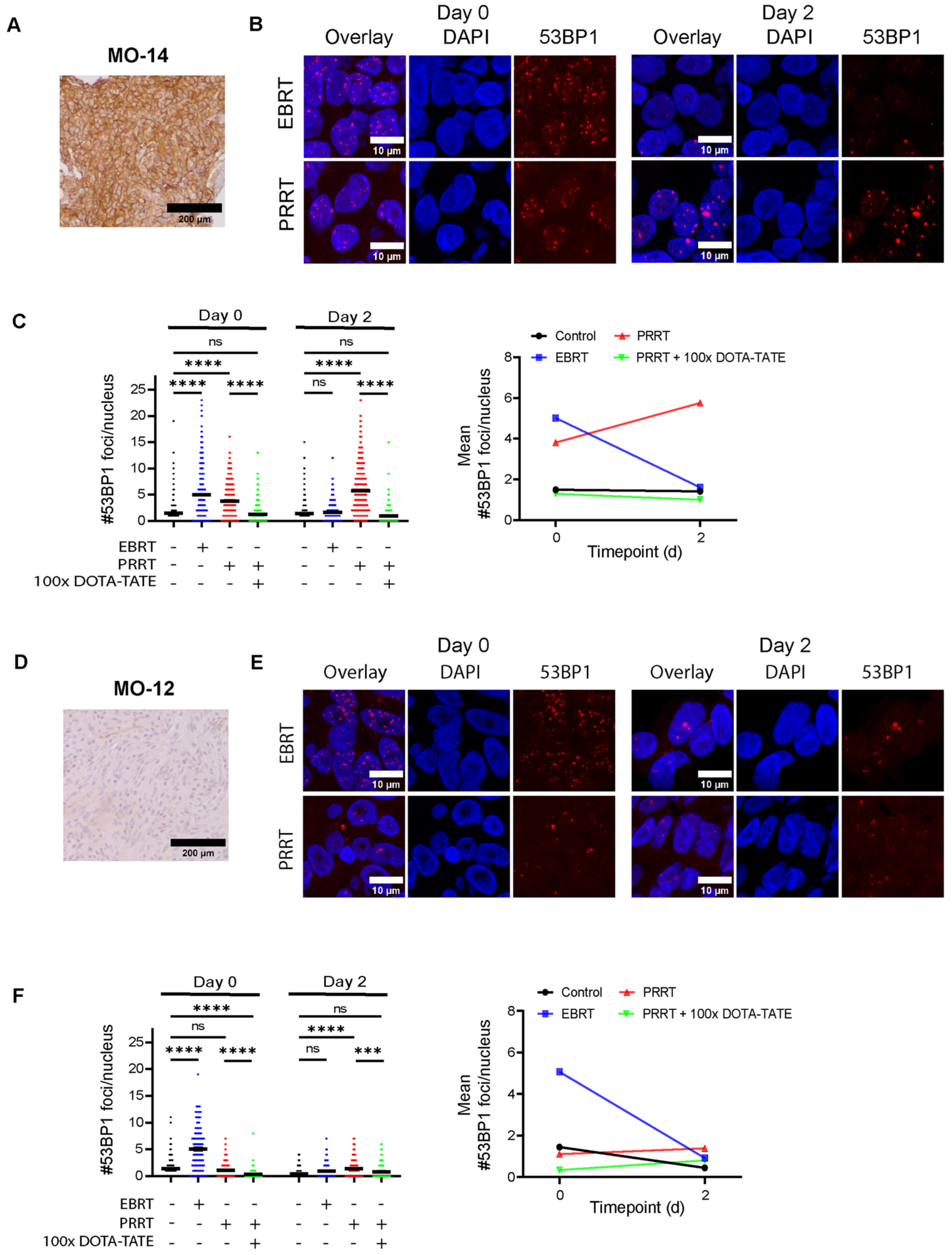
3.4. 177Lu-DOTA-TATE Induces DNA Damage in Meningioma Spheroids Correlating with SSTR2 Expression Levels
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fathi, A.R.; Roelcke, U. Meningioma. Curr. Neurol. Neurosci. Rep. 2013, 13, 337. [Google Scholar] [CrossRef]
- Goldbrunner, R.; Minniti, G.; Preusser, M.; Jenkinson, M.D.; Sallabanda, K.; Houdart, E.; von Deimling, A.; Stavrinou, P.; Lefranc, F.; Lund-Johansen, M.; et al. EANO guidelines for the diagnosis and treatment of meningiomas. Lancet Oncol. 2016, 17, e383–e391. [Google Scholar] [CrossRef]
- Ildan, F.; Erman, T.; Gocer, A.I.; Tuna, M.; Bagdatoglu, H.; Cetinalp, E.; Burgut, R. Predicting the probability of meningioma recurrence in the preoperative and early postoperative period: A multivariate analysis in the midterm follow-up. Skull Base 2007, 17, 157–171. [Google Scholar] [CrossRef] [PubMed]
- Bergner, A.; Maier, A.D.; Mirian, C.; Mathiesen, T.I. Adjuvant radiotherapy and stereotactic radiosurgery in grade 3 meningiomas—A systematic review and meta-analysis. Neurosurg. Rev. 2022, 45, 2639–2658. [Google Scholar] [CrossRef] [PubMed]
- Maclean, J.; Fersht, N.; Short, S. Controversies in radiotherapy for meningioma. Clin. Oncol. (R. Coll. Radiol.) 2014, 26, 51–64. [Google Scholar] [CrossRef] [PubMed]
- Pinzi, V.; Bisogno, I.; Prada, F.; Ciusani, E.; Fariselli, L. Radiotherapy of meningioma: A treatment in need of radiobiological research. Int. J. Radiat. Biol. 2018, 94, 621–627. [Google Scholar] [CrossRef] [PubMed]
- Silva, C.B.; Ongaratti, B.R.; Trott, G.; Haag, T.; Ferreira, N.P.; Leaes, C.G.; Pereira-Lima, J.F.; Oliveira Mda, C. Expression of somatostatin receptors (SSTR1-SSTR5) in meningiomas and its clinicopathological significance. Int. J. Clin. Exp. Pathol. 2015, 8, 13185–13192. [Google Scholar]
- Wu, W.; Zhou, Y.; Wang, Y.; Liu, L.; Lou, J.; Deng, Y.; Zhao, P.; Shao, A. Clinical Significance of Somatostatin Receptor (SSTR) 2 in Meningioma. Front. Oncol. 2020, 10, 1633. [Google Scholar] [CrossRef] [PubMed]
- Mirian, C.; Duun-Henriksen, A.K.; Maier, A.; Pedersen, M.M.; Jensen, L.R.; Bashir, A.; Graillon, T.; Hrachova, M.; Bota, D.; van Essen, M.; et al. Somatostatin Receptor-Targeted Radiopeptide Therapy in Treatment-Refractory Meningioma: Individual Patient Data Meta-analysis. J. Nucl. Med. 2021, 62, 507–513. [Google Scholar] [CrossRef]
- Minczeles, N.S.; Bos, E.M.; de Leeuw, R.C.; Kros, J.M.; Konijnenberg, M.W.; Bromberg, J.E.C.; de Herder, W.W.; Dirven, C.M.F.; Hofland, J.; Brabander, T. Efficacy and safety of peptide receptor radionuclide therapy with [(177)Lu]Lu-DOTA-TATE in 15 patients with progressive treatment-refractory meningioma. Eur. J. Nucl. Med. Mol. Imaging 2023, 50, 1195–1204. [Google Scholar] [CrossRef] [PubMed]
- Feijtel, D.; de Jong, M.; Nonnekens, J. Peptide Receptor Radionuclide Therapy: Looking Back, Looking Forward. Curr. Top. Med. Chem. 2020, 20, 2959–2969. [Google Scholar] [CrossRef] [PubMed]
- Strosberg, J.R.; Caplin, M.E.; Kunz, P.L.; Ruszniewski, P.B.; Bodei, L.; Hendifar, A.; Mittra, E.; Wolin, E.M.; Yao, J.C.; Pavel, M.E.; et al. (177)Lu-Dotatate plus long-acting octreotide versus high-dose long-acting octreotide in patients with midgut neuroendocrine tumours (NETTER-1): Final overall survival and long-term safety results from an open-label, randomised, controlled, phase 3 trial. Lancet Oncol. 2021, 22, 1752–1763. [Google Scholar] [CrossRef]
- Choudhury, A.; Raleigh, D.R. Preclinical models of meningioma: Cell culture and animal systems. Handb. Clin. Neurol. 2020, 169, 131–136. [Google Scholar] [PubMed]
- Boetto, J.; Peyre, M.; Kalamarides, M. Mouse Models in Meningioma Research: A Systematic Review. Cancers 2021, 13, 3712. [Google Scholar] [CrossRef] [PubMed]
- Doctor, A.; Seifert, V.; Ullrich, M.; Hauser, S.; Pietzsch, J. Three-Dimensional Cell Culture Systems in Radiopharmaceutical Cancer Research. Cancers 2020, 12, 2765. [Google Scholar] [CrossRef] [PubMed]
- Eke, I.; Cordes, N. Radiobiology goes 3D: How ECM and cell morphology impact on cell survival after irradiation. Radiother. Oncol. 2011, 99, 271–278. [Google Scholar] [CrossRef] [PubMed]
- Nagle, P.W.; Coppes, R.P. Current and Future Perspectives of the Use of Organoids in Radiobiology. Cells 2020, 9, 2649. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.; Xu, S.; Li, Y.; Shang, L.; Zhan, X.; Qin, C.; Su, J.; Zhao, Z.; He, Y.; Qin, L.; et al. Novel Human Meningioma Organoids Recapitulate the Aggressiveness of the Initiating Cell Subpopulations Identified by ScRNA-Seq. Adv. Sci. 2023, 10, e2205525. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, S.; Ohka, F.; Hirano, M.; Shiraki, Y.; Motomura, K.; Tanahashi, K.; Tsujiuchi, T.; Motomura, A.; Aoki, K.; Shinjo, K.; et al. Newly established patient-derived organoid model of intracranial meningioma. Neuro-Oncology 2021, 23, 1936–1948. [Google Scholar] [CrossRef] [PubMed]
- Feijtel, D.; Doeswijk, G.N.; Verkaik, N.S.; Haeck, J.C.; Chicco, D.; Angotti, C.; Konijnenberg, M.W.; de Jong, M.; Nonnekens, J. Inter and intra-tumor somatostatin receptor 2 heterogeneity influences peptide receptor radionuclide therapy response. Theranostics 2021, 11, 491–505. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, U.; Weigert, M.; Broaddus, C.; Myers, G. Cell Detection with Star-Convex Polygons. In Medical Image Computing and Computer Assisted Intervention–MICCAI 2018: Proceedings of the 21st International Conference, Granada, Spain, 16–20 September 2018; Springer International Publishing: Cham, Switzerland, 2018; pp. 265–273. [Google Scholar]
- Tang, H.; Gong, Y.; Mao, Y.; Xie, Q.; Zheng, M.; Wang, D.; Zhu, H.; Wang, X.; Chen, H.; Chen, X.; et al. CD133-positive cells might be responsible for efficient proliferation of human meningioma cells. Int. J. Mol. Sci. 2012, 13, 6424–6439. [Google Scholar] [CrossRef] [PubMed]
- Menke, J.R.; Raleigh, D.R.; Gown, A.M.; Thomas, S.; Perry, A.; Tihan, T. Somatostatin receptor 2a is a more sensitive diagnostic marker of meningioma than epithelial membrane antigen. Acta Neuropathol. 2015, 130, 441–443. [Google Scholar] [CrossRef] [PubMed]
- Panier, S.; Boulton, S.J. Double-strand break repair: 53BP1 comes into focus. Nat. Rev. Mol. Cell Biol. 2014, 15, 7–18. [Google Scholar] [CrossRef] [PubMed]
- Fodi, C.K.; Schittenhelm, J.; Honegger, J.; Castaneda-Vega, S.G.; Behling, F. The Current Role of Peptide Receptor Radionuclide Therapy in Meningiomas. J. Clin. Med. 2022, 11, 2364. [Google Scholar] [CrossRef] [PubMed]
- van de Weijer, L.L.; Ercolano, E.; Zhang, T.; Shah, M.; Banton, M.C.; Na, J.; Adams, C.L.; Hilton, D.; Kurian, K.M.; Hanemann, C.O. A novel patient-derived meningioma spheroid model as a tool to study and treat epithelial-to-mesenchymal transition (EMT) in meningiomas. Acta Neuropathol. Commun. 2023, 11, 198. [Google Scholar] [CrossRef] [PubMed]
- Endo, H.; Okami, J.; Okuyama, H.; Kumagai, T.; Uchida, J.; Kondo, J.; Takehara, T.; Nishizawa, Y.; Imamura, F.; Higashiyama, M.; et al. Spheroid culture of primary lung cancer cells with neuregulin 1/HER3 pathway activation. J. Thorac. Oncol. 2013, 8, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Kondo, J.; Endo, H.; Okuyama, H.; Ishikawa, O.; Iishi, H.; Tsujii, M.; Ohue, M.; Inoue, M. Retaining cell-cell contact enables preparation and culture of spheroids composed of pure primary cancer cells from colorectal cancer. Proc. Natl. Acad. Sci. USA 2011, 108, 6235–6240. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, T.; Okuyama, H.; Nakayama, M.; Endo, H.; Nonomura, N.; Nishimura, K.; Inoue, M. High-dose chemotherapeutics of intravesical chemotherapy rapidly induce mitochondrial dysfunction in bladder cancer-derived spheroids. Cancer Sci. 2015, 106, 69–77. [Google Scholar] [CrossRef]
- Lim, C.Y.; Chang, J.H.; Lee, W.S.; Lee, K.M.; Yoon, Y.C.; Kim, J.; Park, I.Y. Organotypic slice cultures of pancreatic ductal adenocarcinoma preserve the tumor microenvironment and provide a platform for drug response. Pancreatology 2018, 18, 913–927. [Google Scholar] [CrossRef] [PubMed]
- Meijer, T.G.; Naipal, K.A.; Jager, A.; van Gent, D.C. Ex vivo tumor culture systems for functional drug testing and therapy response prediction. Future Sci. OA 2017, 3, FSO190. [Google Scholar] [CrossRef] [PubMed]
- Naipal, K.A.; Verkaik, N.S.; Sanchez, H.; van Deurzen, C.H.; den Bakker, M.A.; Hoeijmakers, J.H.; Kanaar, R.; Vreeswijk, M.P.; Jager, A.; van Gent, D.C. Tumor slice culture system to assess drug response of primary breast cancer. BMC Cancer 2016, 16, 78. [Google Scholar] [CrossRef]
- Weitz, J.; Hurtado de Mendoza, T.; Tiriac, H.; Lee, J.; Sun, S.; Garg, B.; Patel, J.; Li, K.; Baumgartner, J.; Kelly, K.J.; et al. An Ex Vivo Organotypic Culture Platform for Functional Interrogation of Human Appendiceal Cancer Reveals a Prominent and Heterogenous Immunological Landscape. Clin. Cancer Res. 2022, 28, 4793–4806. [Google Scholar] [CrossRef] [PubMed]
- Antonelli, F. 3D Cell Models in Radiobiology: Improving the Predictive Value of In Vitro Research. Int. J. Mol. Sci. 2023, 24, 10620. [Google Scholar] [CrossRef] [PubMed]
- Storch, K.; Eke, I.; Borgmann, K.; Krause, M.; Richter, C.; Becker, K.; Schrock, E.; Cordes, N. Three-dimensional cell growth confers radioresistance by chromatin density modification. Cancer Res. 2010, 70, 3925–3934. [Google Scholar] [CrossRef] [PubMed]
- Akolawala, Q.; Rovituso, M.; Versteeg, H.H.; Rondon, A.M.R.; Accardo, A. Evaluation of Proton-Induced DNA Damage in 3D-Engineered Glioblastoma Microenvironments. ACS Appl. Mater. Interfaces 2022, 14, 20778–20789. [Google Scholar] [CrossRef] [PubMed]
- Raitanen, J.; Barta, B.; Fuchs, H.; Hacker, M.; Balber, T.; Georg, D.; Mitterhauser, M. Radiobiological Assessment of Targeted Radionuclide Therapy with [177Lu]Lu-PSMA-I&T in 2D vs. 3D Cell Culture Models. Int. J. Mol. Sci. 2023, 24, 17015. [Google Scholar] [CrossRef] [PubMed]
- Dijkstra, B.M.; Motekallemi, A.; den Dunnen, W.F.A.; Jeltema, J.R.; van Dam, G.M.; Kruyt, F.A.E.; Groen, R.J.M. SSTR-2 as a potential tumour-specific marker for fluorescence-guided meningioma surgery. Acta Neurochir. 2018, 160, 1539–1546. [Google Scholar] [CrossRef] [PubMed]
- Koehorst, S.G.; Spapens, M.E.; Van Der Kallen, C.J.; Van, T.V.J.W.; Blaauw, G.; Thijssen, J.H.; Blankenstein, M.A. Progesterone receptor synthesis in human meningiomas: Relation to the estrogen-induced proteins pS2 and cathepsin-D and influence of epidermal growth factor, Forskolin and phorbol ester in vitro. Int. J. Biol. Markers 1998, 13, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Zech, H.B.; Berger, J.; Mansour, W.Y.; Nordquist, L.; von Bargen, C.M.; Bussmann, L.; Oetting, A.; Christiansen, S.; Mockelmann, N.; Bottcher, A.; et al. Patient derived ex vivo tissue slice cultures demonstrate a profound DNA double-strand break repair defect in HPV-positive oropharyngeal head and neck cancer. Radiother. Oncol. 2022, 168, 138–146. [Google Scholar] [CrossRef]
- Willers, H.; Gheorghiu, L.; Liu, Q.; Efstathiou, J.A.; Wirth, L.J.; Krause, M.; von Neubeck, C. DNA Damage Response Assessments in Human Tumor Samples Provide Functional Biomarkers of Radiosensitivity. Semin. Radiat. Oncol. 2015, 25, 237–250. [Google Scholar] [CrossRef] [PubMed]
- Menegakis, A.; von Neubeck, C.; Yaromina, A.; Thames, H.; Hering, S.; Hennenlotter, J.; Scharpf, M.; Noell, S.; Krause, M.; Zips, D.; et al. gammaH2AX assay in ex vivo irradiated tumour specimens: A novel method to determine tumour radiation sensitivity in patient-derived material. Radiother. Oncol. 2015, 116, 473–479. [Google Scholar] [CrossRef] [PubMed]
- Capala, M.E.; Pachler, K.S.; Lauwers, I.; de Korte, M.A.; Verkaik, N.S.; Mast, H.; Jonker, B.P.; Sewnaik, A.; Hardillo, J.A.; Keereweer, S.; et al. Ex Vivo Functional Assay for Evaluating Treatment Response in Tumor Tissue of Head and Neck Squamous Cell Carcinoma. Cancers 2023, 15, 478. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; van Weerden, W.M.; de Ridder, C.M.A.; Erkens-Schulze, S.; Schonfeld, E.; Meijer, T.G.; Kanaar, R.; van Gent, D.C.; Nonnekens, J. Ex vivo treatment of prostate tumor tissue recapitulates in vivo therapy response. Prostate 2019, 79, 390–402. [Google Scholar] [CrossRef] [PubMed]
- Schotzig, U.; Schrader, H.; Schonfeld, E.; Gunther, E.; Klein, R. Standardisation and decay data of 177Lu and 188Re. Appl. Radiat. Isot. 2001, 55, 89–96. [Google Scholar] [CrossRef]
- O’Neill, E.; Mosley, M.; Cornelissen, B. Imaging DNA damage response by gammaH2AX in vivo predicts treatment response to Lutetium-177 radioligand therapy and suggests senescence as a therapeutically desirable outcome. Theranostics 2023, 13, 1302–1310. [Google Scholar] [CrossRef] [PubMed]
- Louis, D.N.; Perry, A.; Wesseling, P.; Brat, D.J.; Cree, I.A.; Figarella-Branger, D.; Hawkins, C.; Ng, H.K.; Pfister, S.M.; Reifenberger, G.; et al. The 2021 WHO Classification of Tumors of the Central Nervous System: A summary. Neuro-Oncology 2021, 23, 1231–1251. [Google Scholar] [CrossRef] [PubMed]
- Patel, A.J.; Wan, Y.W.; Al-Ouran, R.; Revelli, J.P.; Cardenas, M.F.; Oneissi, M.; Xi, L.; Jalali, A.; Magnotti, J.F.; Muzny, D.M.; et al. Molecular profiling predicts meningioma recurrence and reveals loss of DREAM complex repression in aggressive tumors. Proc. Natl. Acad. Sci. USA 2019, 116, 21715–21726. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, A.; Magill, S.T.; Eaton, C.D.; Prager, B.C.; Chen, W.C.; Cady, M.A.; Seo, K.; Lucas, C.G.; Casey-Clyde, T.J.; Vasudevan, H.N.; et al. Meningioma DNA methylation groups identify biological drivers and therapeutic vulnerabilities. Nat. Genet. 2022, 54, 649–659. [Google Scholar] [CrossRef] [PubMed]
- Maas, S.L.N.; Stichel, D.; Hielscher, T.; Sievers, P.; Berghoff, A.S.; Schrimpf, D.; Sill, M.; Euskirchen, P.; Blume, C.; Patel, A.; et al. Integrated Molecular-Morphologic Meningioma Classification: A Multicenter Retrospective Analysis, Retrospectively and Prospectively Validated. J. Clin. Oncol. 2021, 39, 3839–3852. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.C.; Choudhury, A.; Youngblood, M.W.; Polley, M.C.; Lucas, C.G.; Mirchia, K.; Maas, S.L.N.; Suwala, A.K.; Won, M.; Bayley, J.C.; et al. Targeted gene expression profiling predicts meningioma outcomes and radiotherapy responses. Nat. Med. 2023, 29, 3067–3076. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tumor Code | Grade | Tumor Location | Experimental Use | Patient Info (Sex, Age) |
|---|---|---|---|---|
| MO-1 | 1 | Right parietal | Culture conditions | F, 56 |
| MO-2 | 1 | Left frontal, parasagittal | Culture conditions | F, 70 |
| MO-3 | 1 | Left frontal parasagittal | Culture conditions | F, 63 |
| MO-4 | 1 | Right frontal | Culture conditions | F, 61 |
| MO-5 | 1 | Right parasagittal | Culture conditions | F, 51 |
| MO-6 | 1 | Skull base | Culture conditions | F, 74 |
| MO-7 | 1 | Right frontal | Culture conditions | M, 57 |
| MO-8 | 1 | Tuberculum sellae | Culture conditions | M, 71 |
| MO-9 | 1 | Cerebellar | Culture conditions | F, 54 |
| MO-10 | 1 | Right frontal | Culture conditions + Therapy | F, 55 |
| MO-11 | 2 | Skull base | Therapy | M, 60 |
| MO-12 | 1 | Left occipital | Therapy | M, 54 |
| MO-13 | 1 | Right anterior clinoid process | Therapy | M, 72 |
| MO-14 | 3 | Right frontotemporal | Therapy | F, 57 |
| MO-15 | 1 | Falx | Therapy | F, 42 |
| MO-16 | 1 | Right tentorial | Therapy | M, 30 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reuvers, T.G.A.; Grandia, V.; Brandt, R.M.C.; Arab, M.; Maas, S.L.N.; Bos, E.M.; Nonnekens, J. Investigating the Radiobiological Response to Peptide Receptor Radionuclide Therapy Using Patient-Derived Meningioma Spheroids. Cancers 2024, 16, 2515. https://doi.org/10.3390/cancers16142515
Reuvers TGA, Grandia V, Brandt RMC, Arab M, Maas SLN, Bos EM, Nonnekens J. Investigating the Radiobiological Response to Peptide Receptor Radionuclide Therapy Using Patient-Derived Meningioma Spheroids. Cancers. 2024; 16(14):2515. https://doi.org/10.3390/cancers16142515
Chicago/Turabian StyleReuvers, Thom G. A., Vivian Grandia, Renata M. C. Brandt, Majd Arab, Sybren L. N. Maas, Eelke M. Bos, and Julie Nonnekens. 2024. "Investigating the Radiobiological Response to Peptide Receptor Radionuclide Therapy Using Patient-Derived Meningioma Spheroids" Cancers 16, no. 14: 2515. https://doi.org/10.3390/cancers16142515
APA StyleReuvers, T. G. A., Grandia, V., Brandt, R. M. C., Arab, M., Maas, S. L. N., Bos, E. M., & Nonnekens, J. (2024). Investigating the Radiobiological Response to Peptide Receptor Radionuclide Therapy Using Patient-Derived Meningioma Spheroids. Cancers, 16(14), 2515. https://doi.org/10.3390/cancers16142515