
Lactococcus lactis subsp. cremoris C60 Upregulates Macrophage Function by Modifying Metabolic Preference in Enhanced Anti-Tumor Immunity



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Lactic Acid Bacteria Culture
2.2. Isolation of Bacterial Cell Wall Extract
2.3. Cell Culture
2.4. Mice and Tumor Model
2.5. Flow Cytometry
2.6. Real-Time Polymerase Chain Reaction (Real-Time PCR)
2.7. Tumor Cell Isolation
2.8. Tumor Killing Assay
2.9. Antigen Uptake Assay
2.10. Antigen Presentation Assay
2.11. Adenosine Triphosphate (ATP) Assay
2.12. Glucose Uptake Assay
2.13. Lipid Uptake Assay
2.14. Cytokine Production Assay
2.15. Real-Time Metabolic Assay
2.16. Metabolomics
2.17. Statistics
3. Results
3.1. C60 Suppresses Tumor Growth by Inducing a Predominantly Inflammatory Phenotype in Macrophages
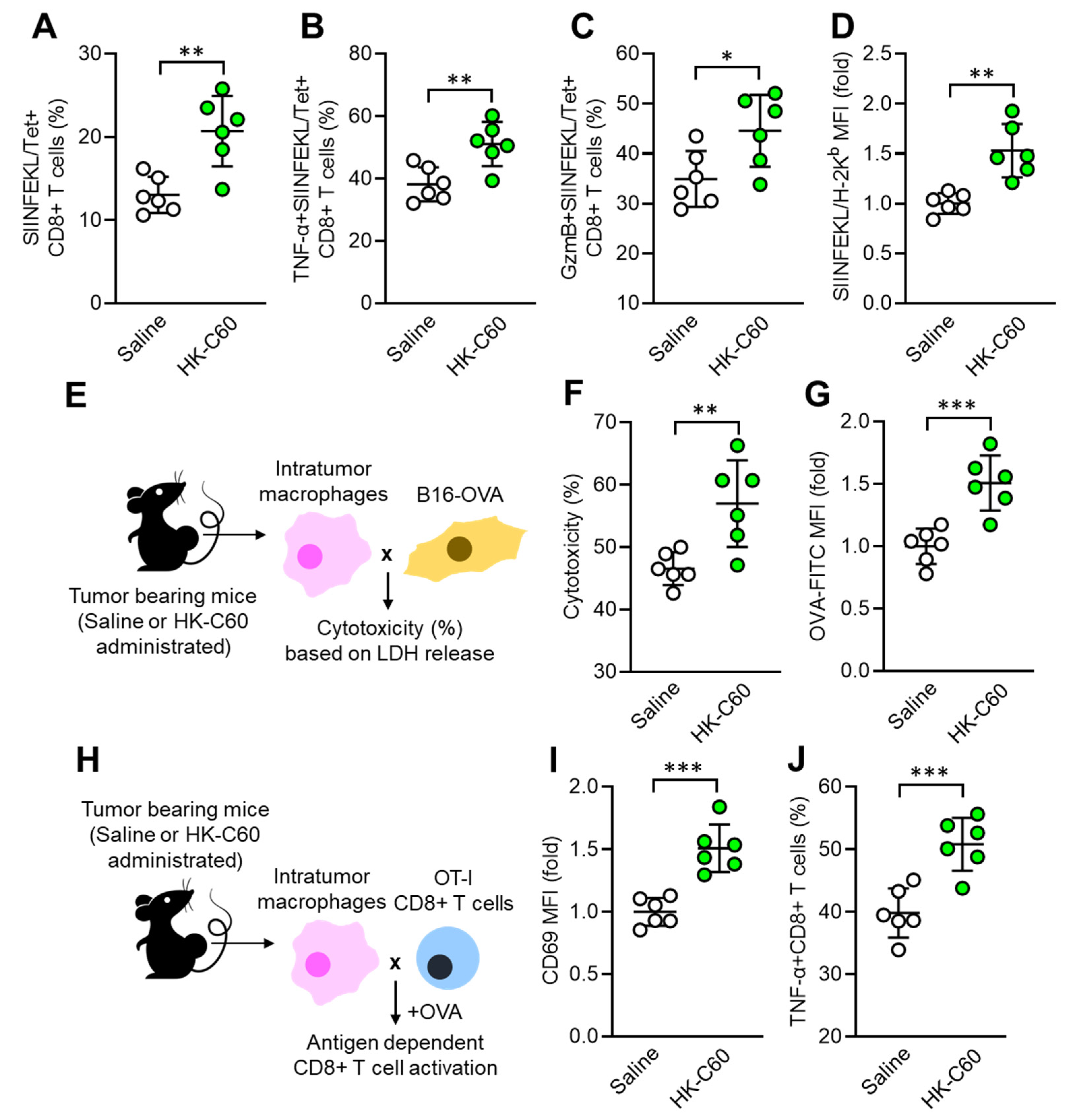
3.2. C60 Enhances Antigen Presentation Function of Macrophages Activating CD8+ T Cells
3.3. C60 Enhances Mitochondrial Oxidative Metabolism to Increase ATP Production in Macrophages
3.4. C60 Modifies Metabolic Demand as Glycolysis Preferable Manner in Macrophages
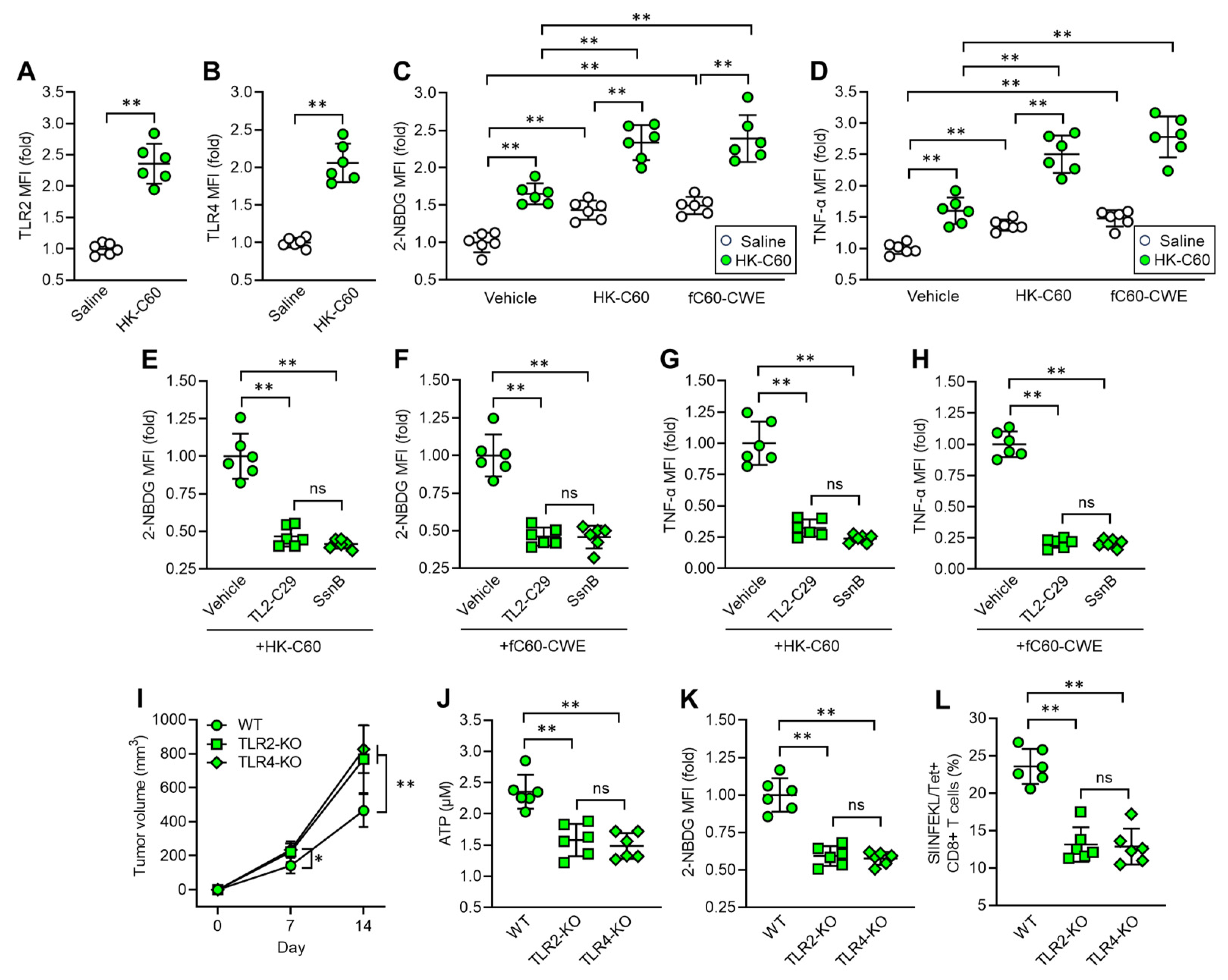
3.5. C60 Modifies Macrophage Activity and Metabolism via TLR Signaling in Enhanced Anti-Tumor Immunity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Klaenhammer, T.R.; Kleerebezem, M.; Kopp, M.V.; Rescigno, M. The impact of probiotics and prebiotics on the immune system. Nat. Rev. Immunol. 2012, 12, 728–734. [Google Scholar] [CrossRef] [PubMed]
- Cristofori, F.; Dargenio, V.N.; Dargenio, C.; Miniello, V.L.; Barone, M.; Francavilla, R. Anti-Inflammatory and Immunomodulatory Effects of Probiotics in Gut Inflammation: A Door to the Body. Front. Immunol. 2021, 12, 578386. [Google Scholar] [CrossRef] [PubMed]
- Christensen, H.R.; Frøkiær, H.; Pestka, J.J. Lactobacilli Differentially Modulate Expression of Cytokines and Maturation Surface Markers in Murine Dendritic Cells. J. Immunol. 2002, 168, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Hart, A.L.; Lammers, K.; Brigidi, P.; Vitali, B.; Rizzello, F.; Gionchetti, P.; Campieri, M.; Kamm, M.A.; Knight, S.C.; Stagg, A.J. Modulation of human dendritic cell phenotype and function by probiotic bacteria. Gut 2004, 53, 1602–1609. [Google Scholar] [CrossRef] [PubMed]
- Fong, F.L.Y.; Kirjavainen, P.V.; El-Nezami, H. Immunomodulation of Lactobacillus rhamnosus GG (LGG)-derived soluble factors on antigen-presenting cells of healthy blood donors. Sci. Rep. 2016, 6, 22845. [Google Scholar] [CrossRef] [PubMed]
- Ren, C.; Zhang, Q.; de Haan, B.J.; Zhang, H.; Faas, M.M.; de Vos, P. Identification of TLR2/TLR6 signalling lactic acid bacteria for supporting immune regulation. Sci. Rep. 2016, 6, 34561. [Google Scholar] [CrossRef] [PubMed]
- Kawashima, T.; Kosaka, A.; Yan, H.; Guo, Z.; Uchiyama, R.; Fukui, R.; Kaneko, D.; Kumagai, Y.; You, D.-J.; Carreras, J.; et al. Double-Stranded RNA of Intestinal Commensal but Not Pathogenic Bacteria Triggers Production of Protective Interferon-β. Immunity 2013, 38, 1187–1197. [Google Scholar] [CrossRef]
- Mohamadzadeh, M.; Olson, S.; Kalina, W.V.; Ruthel, G.; Demmin, G.L.; Warfield, K.L.; Bavari, S.; Klaenhammer, T.R. Lactobacilli activate human dendritic cells that skew T cells toward T helper 1 polarization. Proc. Natl. Acad. Sci. USA 2005, 102, 2880–2885. [Google Scholar] [CrossRef] [PubMed]
- Corthésy, B.; Gaskins, H.R.; Mercenier, A. Cross-Talk between Probiotic Bacteria and the Host Immune System. J. Nutr. 2007, 137 (Suppl. 2), 781S–790S. [Google Scholar] [CrossRef]
- Jeon, S.G.; Kayama, H.; Ueda, Y.; Takahashi, T.; Asahara, T.; Tsuji, H.; Tsuji, N.M.; Kiyono, H.; Ma, J.S.; Kusu, T.; et al. Probiotic Bifidobacterium breve Induces IL-10-Producing Tr1 Cells in the Colon. PLoS Pathog. 2012, 8, e1002714. [Google Scholar] [CrossRef]
- Kawashima, T.; Ikari, N.; Kouchi, T.; Kowatari, Y.; Kubota, Y.; Shimojo, N.; Tsuji, N.M. The molecular mechanism for activating IgA production by Pediococcus acidilactici K15 and the clinical impact in a randomized trial. Sci. Rep. 2018, 8, 5065. [Google Scholar] [CrossRef]
- Kawashima, T.; Ikari, N.; Watanabe, Y.; Kubota, Y.; Yoshio, S.; Kanto, T.; Motohashi, S.; Shimojo, N.; Tsuji, N.M. Double-Stranded RNA Derived from Lactic Acid Bacteria Augments Th1 Immunity via Interferon-β from Human Dendritic Cells. Front. Immunol. 2018, 9, 27. [Google Scholar] [CrossRef]
- Kamiya, T.; Watanabe, Y.; Makino, S.; Kano, H.; Tsuji, N.M. Improvement of Intestinal Immune Cell Function by Lactic Acid Bacteria for Dairy Products. Microorganisms 2016, 5, 1. [Google Scholar] [CrossRef]
- Saito, S.; Kakizaki, N.; Okuno, A.; Maekawa, T.; Tsuji, N.M. Lactococcus lactis subsp. cremoris C60 restores T Cell Population in Small Intestinal Lamina Propria in Aged Interleukin-18 Deficient Mice. Nutrients 2020, 12, 3287. [Google Scholar]
- Saito, S.; Okuno, A.; Kakizaki, N.; Maekawa, T.; Tsuji, N.M. Lactococcus lactis subsp. cremoris C60 induces macrophages activation that enhances CD4+ T cell-based adaptive immunity. Biosci. Microbiota Food Health 2022, 41, 130–136. [Google Scholar]
- Jang, S.O.; Kim, H.J.; Kim, Y.J.; Kang, M.J.; Kwon, J.W.; Seo, J.H.; Kim, H.Y.; Kim, B.J.; Yu, J.; Hong, S.J. Asthma Prevention by Lactobacillus rhamnosus in a Mouse Model is Associated with CD4+CD25+Foxp3+ T Cells. Allergy Asthma Immunol. Res. 2012, 4, 150–156. [Google Scholar] [CrossRef]
- Wang, X.; Hui, Y.; Zhao, L.; Hao, Y.; Guo, H.; Ren, F. Oral administration of Lactobacillus paracasei L9 attenuates PM2.5-induced enhancement of airway hyperresponsiveness and allergic airway response in murine model of asthma. PLoS ONE 2017, 12, e0171721. [Google Scholar] [CrossRef]
- Fu, L.; Peng, J.; Zhao, S.; Zhang, Y.; Su, X.; Wang, Y. Lactic acid bacteria-specific induction of CD4+Foxp3+T cells ameliorates shrimp tropomyosin-induced allergic response in mice via suppression of mTOR signaling. Sci. Rep. 2017, 7, 1987. [Google Scholar] [CrossRef]
- Saito, S.; Okuno, A.; Peng, Z.; Cao, D.-Y.; Tsuji, N.M. Probiotic lactic acid bacteria promote anti-tumor immunity through enhanced major histocompatibility complex class I-restricted antigen presentation machinery in dendritic cells. Front. Immunol. 2024, 15, 1335975. [Google Scholar] [CrossRef]
- Saito, S.; Quadery, A.F. Staphylococcus aureus Lipoprotein Induces Skin Inflammation, Accompanied with IFN-γ-Producing T Cell Accumulation through Dermal Dendritic Cells. Pathogens 2018, 7, 64. [Google Scholar] [CrossRef]
- Peng, Z.; Saito, S. Creatine supplementation enhances anti-tumor immunity by promoting adenosine triphosphate production in macrophages. Front. Immunol. 2023, 14, 1176956. [Google Scholar] [CrossRef]
- Cao, D.Y.; Spivia, W.R.; Veiras, L.C.; Khan, Z.; Peng, Z.; Jones, A.E.; Bernstein, E.A.; Saito, S.; Okwan-Duodu, D.; Parker, S.J.; et al. ACE overexpression in myeloid cells increases oxidative metabolism and cellular ATP. J. Biol. Chem. 2020, 295, 1369–1384. [Google Scholar] [CrossRef]
- Saito, S.; Shahbaz, S.; Luo, X.; Osman, M.; Redmond, D.; Tervaert, J.W.C.; Li, L.; Elahi, S. Metabolomic and immune alterations in long COVID patients with chronic fatigue syndrome. Front. Immunol. 2024, 15, 1341843. [Google Scholar] [CrossRef]
- Raskov, H.; Orhan, A.; Christensen, J.P.; Gögenur, I. Cytotoxic CD8+ T cells in cancer and cancer immunotherapy. Br. J. Cancer 2021, 124, 359–367. [Google Scholar] [CrossRef]
- Liu, S.; Liu, S.; He, B.; Li, L.; Li, L.; Wang, J.; Cai, T.; Chen, S.; Jiang, H. OXPHOS deficiency activates global adaptation pathways to maintain mitochondrial membrane potential. EMBO Rep. 2021, 22, e51606. [Google Scholar] [CrossRef]
- Matsubara, V.H.; Ishikawa, K.H.; Ando-Suguimoto, E.S.; Bueno-Silva, B.; Nakamae, A.E.M.; Mayer, M.P.A. Probiotic Bacteria Alter Pattern-Recognition Receptor Expression and Cytokine Profile in a Human Macrophage Model Challenged with Candida albicans and Lipopolysaccharide. Front. Microbiol. 2017, 8, 2280. [Google Scholar] [CrossRef]
- Hishiki, H.; Kawashima, T.; Tsuji, N.M.; Ikari, N.; Takemura, R.; Kido, H.; Shimojo, N. A Double-Blind, Randomized, Placebo-Controlled Trial of Heat-Killed Pediococcus acidilactici K15 for Prevention of Respiratory Tract Infections among Preschool Children. Nutrients 2020, 12, 1989. [Google Scholar] [CrossRef]
- Noverr, M.C.; Huffnagle, G.B. Does the microbiota regulate immune responses outside the gut? Trends Microbiol. 2004, 12, 562–568. [Google Scholar] [CrossRef]
- Bogunovic, M.; Ginhoux, F.; Helft, J.; Shang, L.; Hashimoto, D.; Greter, M.; Liu, K.; Jakubzick, C.; Ingersoll, M.A.; Leboeuf, M.; et al. Origin of the Lamina Propria Dendritic Cell Network. Immunity 2009, 31, 513–525. [Google Scholar] [CrossRef]
- Wells, J.M.; Mercenier, A. Mucosal delivery of therapeutic and prophylactic molecules using lactic acid bacteria. Nat. Rev. Microbiol. 2008, 6, 349–362. [Google Scholar] [CrossRef]
- Chapot-Chartier, M.P.; Kulakauskas, S. Cell wall structure and function in lactic acid bacteria. Microb. Cell Factories 2014, 13 (Suppl. 1), S9. [Google Scholar] [CrossRef]
- Teame, T.; Wang, A.; Xie, M.; Zhang, Z.; Yang, Y.; Ding, Q.; Gao, C.; Olsen, R.E.; Ran, C.; Zhou, Z. Paraprobiotics and Postbiotics of Probiotic Lactobacilli, Their Positive Effects on the Host and Action Mechanisms: A Review. Front Nutr. 2020, 7, 570344. [Google Scholar] [CrossRef]
- Chen, L.; Ozato, K. Innate Immune Memory in Hematopoietic Stem/Progenitor Cells: Myeloid-Biased Differentiation and the Role of Interferon. Front. Immunol. 2021, 12, 621333. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, J.; Lv, M.; Shao, Z.; Hungwe, M.; Wang, J.; Bai, X.; Xie, J.; Wang, Y.; Geng, W. Metabolism Characteristics of Lactic Acid Bacteria and the Expanding Applications in Food Industry. Front. Bioeng. Biotechnol. 2021, 9, 612285. [Google Scholar] [CrossRef]
- Nes, I.F.; Holo, H. Class II antimicrobial peptides from lactic acid bacteria. Biopolymers 2000, 55, 50–61. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saito, S.; Cao, D.-Y.; Maekawa, T.; Tsuji, N.M.; Okuno, A. Lactococcus lactis subsp. cremoris C60 Upregulates Macrophage Function by Modifying Metabolic Preference in Enhanced Anti-Tumor Immunity. Cancers 2024, 16, 1928. https://doi.org/10.3390/cancers16101928
Saito S, Cao D-Y, Maekawa T, Tsuji NM, Okuno A. Lactococcus lactis subsp. cremoris C60 Upregulates Macrophage Function by Modifying Metabolic Preference in Enhanced Anti-Tumor Immunity. Cancers. 2024; 16(10):1928. https://doi.org/10.3390/cancers16101928
Chicago/Turabian StyleSaito, Suguru, Duo-Yao Cao, Toshio Maekawa, Noriko M. Tsuji, and Alato Okuno. 2024. "Lactococcus lactis subsp. cremoris C60 Upregulates Macrophage Function by Modifying Metabolic Preference in Enhanced Anti-Tumor Immunity" Cancers 16, no. 10: 1928. https://doi.org/10.3390/cancers16101928
APA StyleSaito, S., Cao, D.-Y., Maekawa, T., Tsuji, N. M., & Okuno, A. (2024). Lactococcus lactis subsp. cremoris C60 Upregulates Macrophage Function by Modifying Metabolic Preference in Enhanced Anti-Tumor Immunity. Cancers, 16(10), 1928. https://doi.org/10.3390/cancers16101928