

Expression of Interleukin-13 Receptor Alpha 2 in Brainstem Gliomas
 ,
,  , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Tumor Sample
2.2. TMA Construction and Multiplex Immunofluorescence Analysis
2.3. Bioinformatic Confirmation and Analysis
2.4. Western Blot
2.5. qPCR
2.6. Cell Culture
2.7. Statistical Analysis
3. Results
3.1. Brainstem Glioma Cohorts Included in This Study
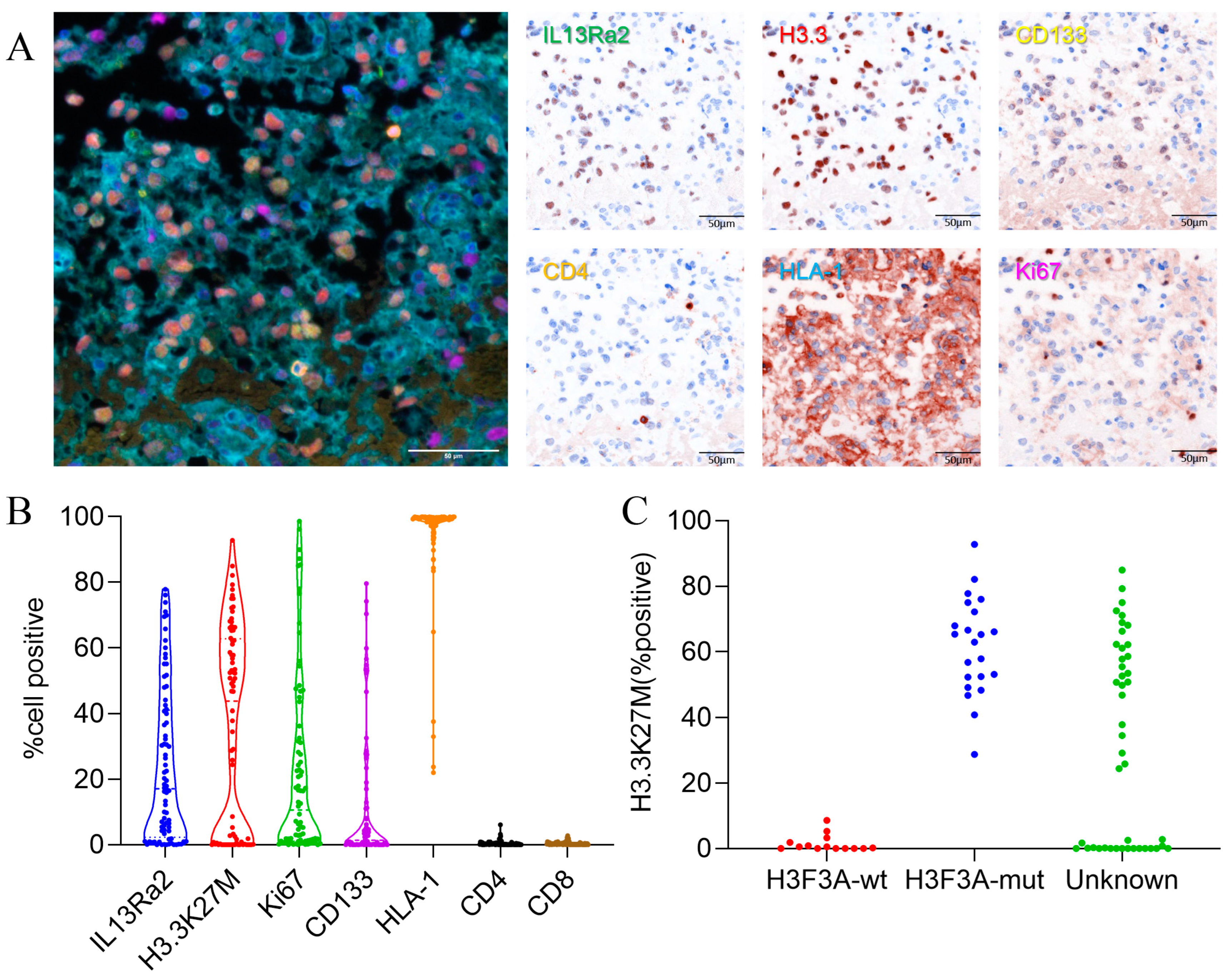
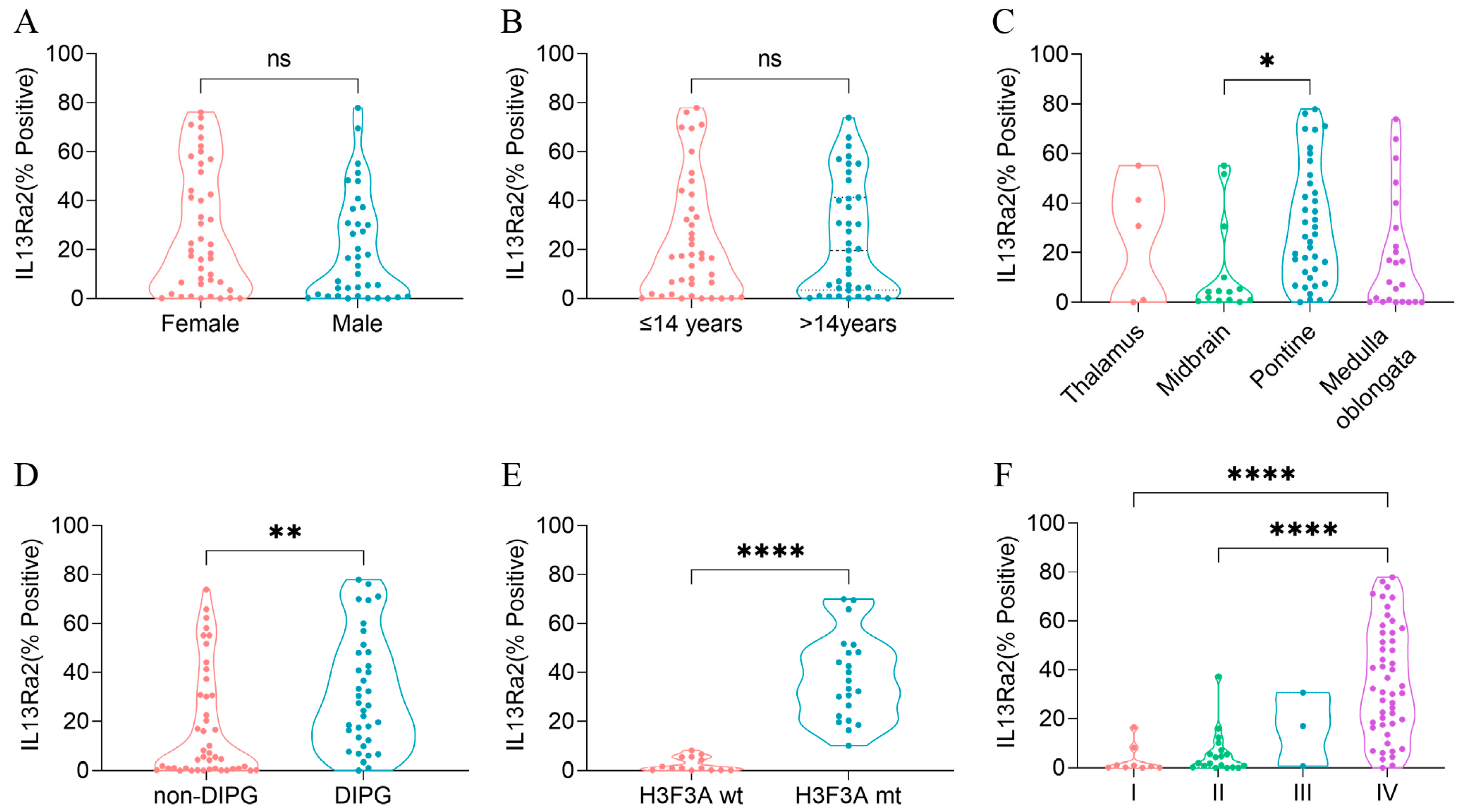
3.2. IL13Ra2 Staining in BSGs Tissues
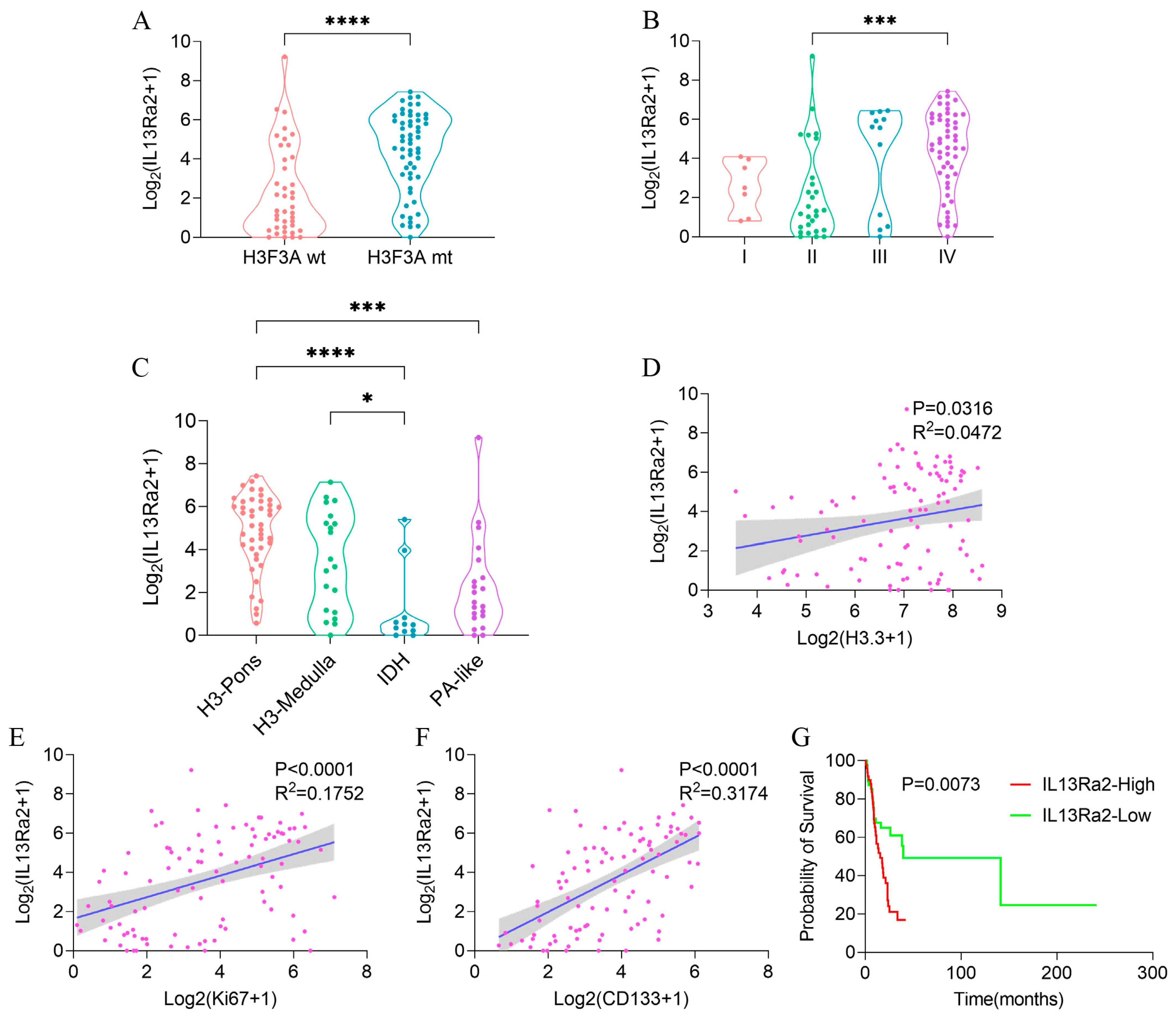
3.3. High IL13Ra2 Expression Was Enriched in H3.3-Mutant BSGs
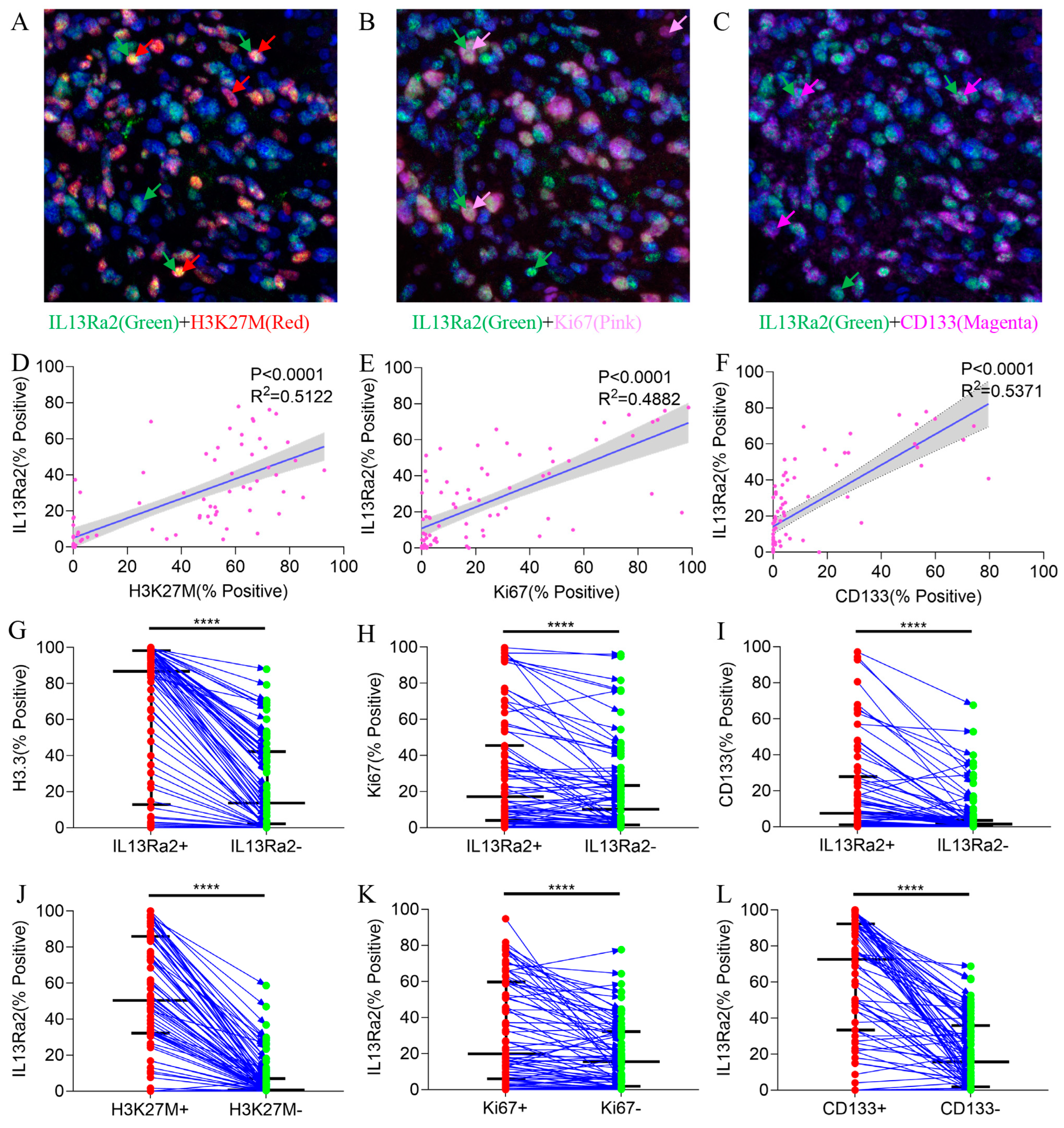
3.4. IL13Ra2 Expression Was Enriched in Glioma Cells Harboring the H3F3A Mutation, Proliferation Features, and Stem Cell Properties
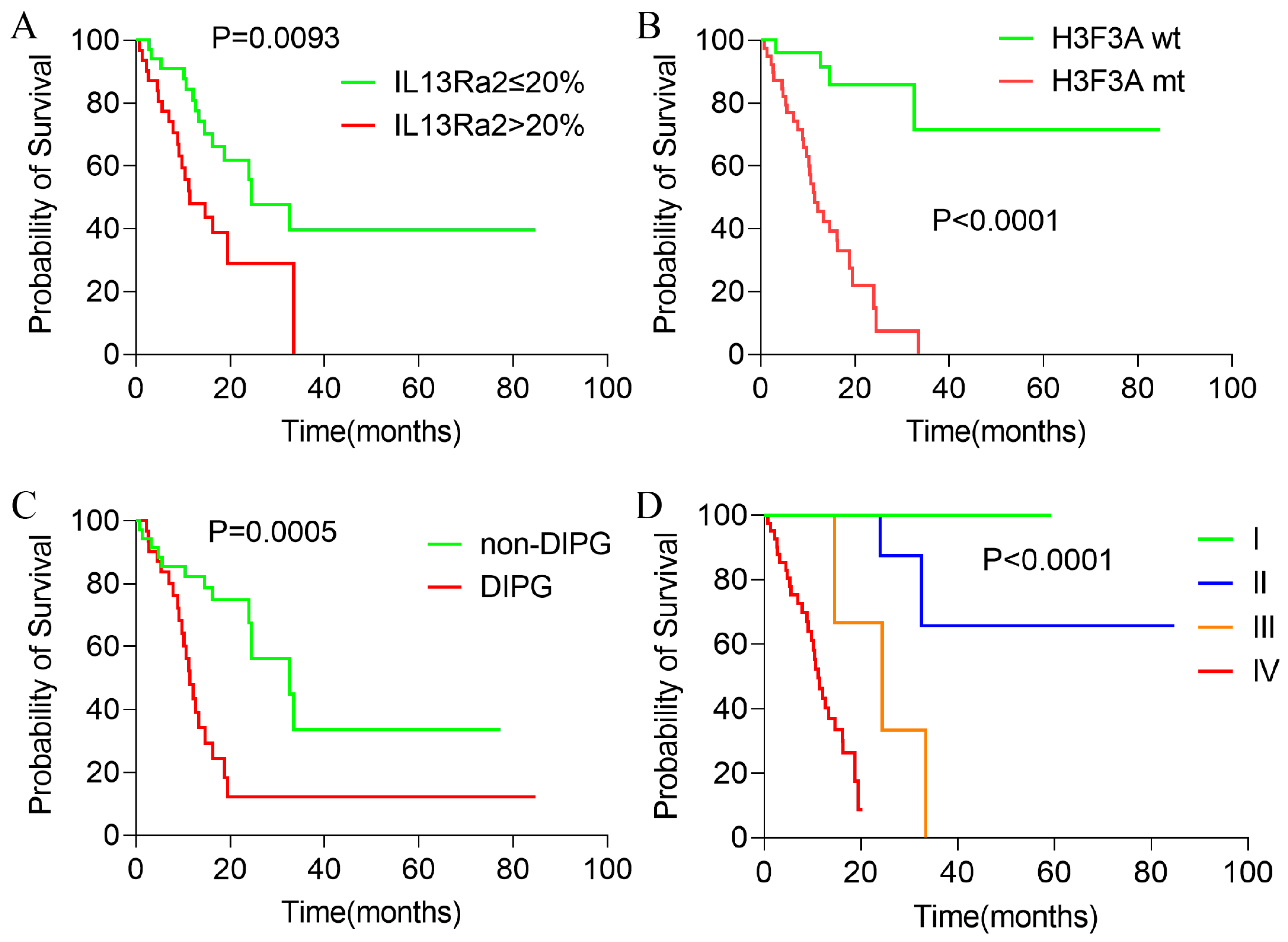
3.5. Higher IL13Ra2 Expression Associated with Shorten Overall Survival
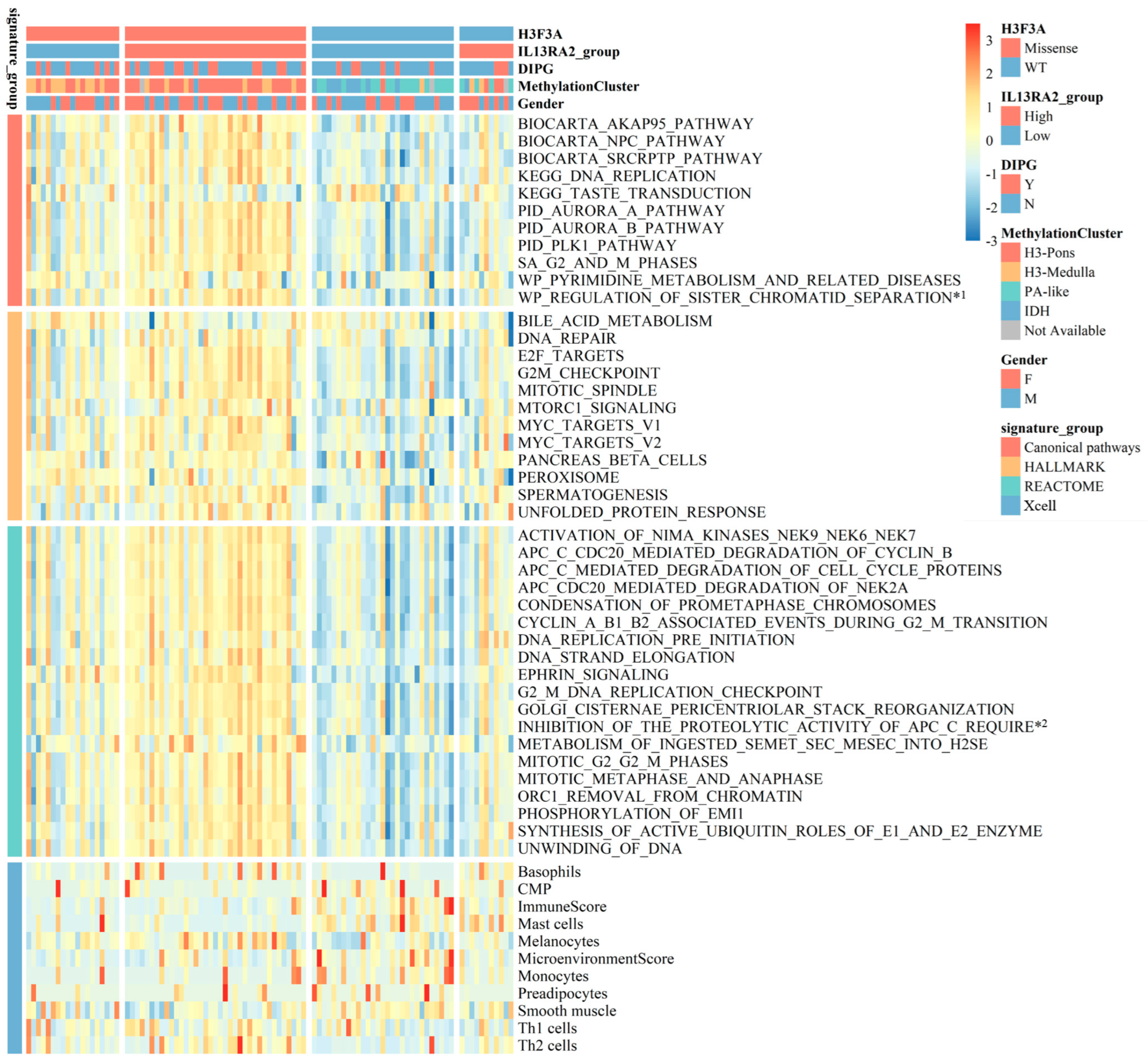
3.6. IL13Ra2 and Pathways Promoting BSG Progression
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Grimm, S.A.; Chamberlain, M.C. Brainstem glioma: A review. Curr. Neurol. Neurosci. Rep. 2013, 13, 346. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Western, S.; Kesari, S. Brainstem glioma in adults. Front. Oncol. 2016, 6, 180. [Google Scholar] [CrossRef] [PubMed]
- Wummer, B.; Woodworth, D.; Flores, C. Brain stem gliomas and current landscape. J. Neuro-Oncol. 2021, 151, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Ostrom, Q.T.; Gittleman, H.; Fulop, J.; Liu, M.; Blanda, R.; Kromer, C.; Wolinsky, Y.; Kruchko, C.; Barnholtz-Sloan, J.S. Cbtrus statistical report: Primary brain and central nervous system tumors diagnosed in the united states in 2008-2012. Neuro-Oncology 2015, 17, v1–v62. [Google Scholar] [CrossRef] [PubMed]
- Khuong-Quang, D.; Buczkowicz, P.; Rakopoulos, P.; Liu, X.; Fontebasso, A.M.; Bouffet, E.; Bartels, U.; Albrecht, S.; Schwartzentruber, J.; Letourneau, L.; et al. K27m mutation in histone h3.3 defines clinically and biologically distinct subgroups of pediatric diffuse intrinsic pontine gliomas. Acta Neuropathol. 2012, 124, 439–447. [Google Scholar] [CrossRef] [PubMed]
- Schwartzentruber, J.; Korshunov, A.; Liu, X.; Jones, D.T.W.; Pfaff, E.; Jacob, K.; Sturm, D.; Fontebasso, A.M.; Quang, D.K.; Tönjes, M.; et al. Driver mutations in histone h3.3 and chromatin remodelling genes in paediatric glioblastoma. Nature 2012, 482, 226–231. [Google Scholar] [CrossRef] [PubMed]
- Ius, T.; Lombardi, G.; Baiano, C.; Berardinelli, J.; Romano, A.; Montemurro, N.; Cavallo, L.M.; Pasqualetti, F.; Feletti, A. Surgical management of adult brainstem gliomas: A systematic review and meta-analysis. Curr. Oncol. 2023, 30, 9772–9785. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Xiao, X.; Gu, G.; Zhang, P.; Wu, W.; Wang, Y.; Pan, C.; Wang, L.; Li, H.; Wu, Z.; et al. Role of neuronavigation in the surgical management of brainstem gliomas. Front. Oncol. 2023, 13, 1159230. [Google Scholar] [CrossRef]
- Pan, C.; Zhang, M.; Xiao, X.; Kong, L.; Wu, Y.; Zhao, X.; Sun, T.; Zhang, P.; Geng, Y.; Zuo, P.; et al. A multimodal imaging-based classification for pediatric diffuse intrinsic pontine gliomas. Neurosurg. Rev. 2023, 46, 151. [Google Scholar] [CrossRef]
- Rashed, W.M.; Maher, E.; Adel, M.; Saber, O.; Zaghloul, M.S. Pediatric diffuse intrinsic pontine glioma: Where do we stand? Cancer Metastasis Rev. 2019, 38, 759–770. [Google Scholar] [CrossRef]
- Gwak, H.S.; Park, H.J. Developing chemotherapy for diffuse pontine intrinsic gliomas (dipg). Crit. Rev. Oncol./Hematol. 2017, 120, 111–119. [Google Scholar] [CrossRef]
- Johung, T.B.; Monje, M. Diffuse intrinsic pontine glioma: New pathophysiological insights and emerging therapeutic targets. Curr. Neuropharmacol. 2017, 15, 88–97. [Google Scholar] [CrossRef] [PubMed]
- Grosser, R.; Cherkassky, L.; Chintala, N.; Adusumilli, P.S. Combination immunotherapy with car t cells and checkpoint blockade for the treatment of solid tumors. Cancer Cell 2019, 36, 471–482. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.K.; Mcguirk, J.P. Car t cells: Continuation in a revolution of immunotherapy. Lancet Oncol. 2020, 21, e168–e178. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Zhang, Y.; Ji, N. Challenges in the treatment of glioblastoma by chimeric antigen receptor t-cell immunotherapy and possible solutions. Front. Immunol. 2022, 13, 927132. [Google Scholar] [CrossRef] [PubMed]
- Akhavan, D.; Alizadeh, D.; Wang, D.; Weist, M.R.; Shepphird, J.K.; Brown, C.E. Car t cells for brain tumors: Lessons learned and road ahead. Immunol. Rev. 2019, 290, 60–84. [Google Scholar] [CrossRef] [PubMed]
- Brown, C.E.; Alizadeh, D.; Starr, R.; Weng, L.; Wagner, J.R.; Naranjo, A.; Ostberg, J.R.; Blanchard, M.S.; Kilpatrick, J.; Simpson, J.; et al. Regression of glioblastoma after chimeric antigen receptor t-cell therapy. N. Engl. J. Med. 2016, 375, 2561–2569. [Google Scholar] [CrossRef]
- Thaci, B.; Brown, C.E.; Binello, E.; Werbaneth, K.; Sampath, P.; Sengupta, S. Significance of interleukin-13 receptor alpha 2-targeted glioblastoma therapy. Neuro-Oncology 2014, 16, 1304–1312. [Google Scholar] [CrossRef]
- Rahaman, S.O.; Sharma, P.; Harbor, P.C.; Aman, M.J.; Vogelbaum, M.A.; Haque, S.J. Il-13r(alpha)2, a decoy receptor for il-13 acts as an inhibitor of il-4-dependent signal transduction in glioblastoma cells. Cancer Res. 2002, 62, 1103–1109. [Google Scholar]
- Arima, K.; Sato, K.; Tanaka, G.; Kanaji, S.; Terada, T.; Honjo, E.; Kuroki, R.; Matsuo, Y.; Izuhara, K. Characterization of the interaction between interleukin-13 and interleukin-13 receptors. J. Biol. Chem. 2005, 280, 24915–24922. [Google Scholar] [CrossRef]
- Zeng, J.; Zhang, J.; Yang, Y.Z.; Wang, F.; Jiang, H.; Chen, H.D.; Wu, H.Y.; Sai, K.; Hu, W.M. Il13ra2 is overexpressed in malignant gliomas and related to clinical outcome of patients. Am. J. Transl. Res. 2020, 12, 4702–4714. [Google Scholar] [PubMed]
- Brown, C.E.; Warden, C.D.; Starr, R.; Deng, X.; Badie, B.; Yuan, Y.; Forman, S.J.; Barish, M.E. Glioma il13rα2 is associated with mesenchymal signature gene expression and poor patient prognosis. PLoS ONE 2013, 8, e77769. [Google Scholar] [CrossRef] [PubMed]
- Bhardwaj, R.; Suzuki, A.; Leland, P.; Joshi, B.H.; Puri, R.K. Identification of a novel role of il-13rα2 in human glioblastoma multiforme: Interleukin-13 mediates signal transduction through ap-1 pathway. J. Transl. Med. 2018, 16, 369. [Google Scholar] [CrossRef]
- Han, J.; Puri, R.K. Analysis of the cancer genome atlas (tcga) database identifies an inverse relationship between interleukin-13 receptor α1 and α2 gene expression and poor prognosis and drug resistance in subjects with glioblastoma multiforme. J. Neuro-Oncol. 2018, 136, 463–474. [Google Scholar] [CrossRef]
- Joshi, B.H.; Puri, R.A.; Leland, P.; Varricchio, F.; Gupta, G.; Kocak, M.; Gilbertson, R.J.; Puri, R.K. Identification of interleukin-13 receptor α2 chain overexpression in situ in high-grade diffusely infiltrative pediatric brainstem glioma. Neuro-Oncology 2008, 10, 265–274. [Google Scholar] [CrossRef] [PubMed]
- Okada, H.; Low, K.L.; Kohanbash, G.; Mcdonald, H.A.; Hamilton, R.L.; Pollack, I.F. Expression of glioma-associated antigens in pediatric brain stem and non-brain stem gliomas. J. Neuro-Oncol. 2008, 88, 245–250. [Google Scholar] [CrossRef]
- Berlow, N.E.; Svalina, M.N.; Quist, M.J.; Settelmeyer, T.P.; Zherebitskiy, V.; Kogiso, M.; Qi, L.; Du, Y.; Hawkins, C.E.; Hulleman, E.; et al. Il-13 receptors as possible therapeutic targets in diffuse intrinsic pontine glioma. PLoS ONE 2018, 13, e193565. [Google Scholar] [CrossRef] [PubMed]
- Jaén, M.; Martín-Regalado, Á.; Bartolomé, R.A.; Robles, J.; Casal, J.I. Interleukin 13 receptor alpha 2 (il13rα2): Expression, signaling pathways and therapeutic applications in cancer. Biochim. Et Biophys. Acta (BBA)-Rev. Cancer 2022, 1877, 188802. [Google Scholar] [CrossRef]
- Chen, L.H.; Pan, C.; Diplas, B.H.; Xu, C.; Hansen, L.J.; Wu, Y.; Chen, X.; Geng, Y.; Sun, T.; Sun, Y.; et al. The integrated genomic and epigenomic landscape of brainstem glioma. Nat. Commun. 2020, 11, 3077. [Google Scholar] [CrossRef]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Genome Biol. 2010, 11, R106. [Google Scholar] [CrossRef]
- Gene Ontology Consortium. Gene ontology consortium: Going forward. Nucleic Acids Res. 2015, 43, D1049–D1056. [Google Scholar] [CrossRef] [PubMed]
- Hanzelmann, S.; Castelo, R.; Guinney, J. Gsva: Gene set variation analysis for microarray and rna-seq data. Bmc Bioinformatics 2013, 14, 7. [Google Scholar] [CrossRef] [PubMed]
- Louis, D.N.; Perry, A.; Wesseling, P.; Brat, D.J.; Cree, I.A.; Figarella-Branger, D.; Hawkins, C.; Ng, H.K.; Pfister, S.M.; Reifenberger, G.; et al. The 2021 who classification of tumors of the central nervous system: A summary. Neuro-Oncology 2021, 23, 1231–1251. [Google Scholar] [CrossRef] [PubMed]
- Louis, D.N.; Ohgaki, H.; Wiestler, O.D.; Cavenee, W.K. WHO Classification of Tumours of the Central Nervous System; World Health Organization (WHO): Geneva, Switzerland, 2016; pp. 10, 57–59. [Google Scholar]
- Pawlowski, K.D.; Duffy, J.T.; Tiwari, A.; Zannikou, M.; Balyasnikova, I.V. Bi-specific killer cell engager enhances nk cell activity against interleukin-13 receptor alpha-2 positive gliomas. Cells 2023, 12, 1716. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.; Rodriguez, J.L.; Li, N.; Thokala, R.; Nasrallah, M.P.; Hu, L.; Zhang, L.; Zhang, J.V.; Logun, M.T.; Kainth, D.; et al. Locally secreted bites complement car t cells by enhancing killing of antigen heterogeneous solid tumors. Mol. Ther. 2022, 30, 2537–2553. [Google Scholar] [CrossRef] [PubMed]
- Pandya, H.; Gibo, D.M.; Garg, S.; Kridel, S.; Debinski, W. An interleukin 13 receptor α 2–specific peptide homes to human glioblastoma multiforme xenografts. Neuro-Oncology 2012, 14, 6–18. [Google Scholar] [CrossRef]
- Balyasnikova, I.V.; Wainwright, D.A.; Solomaha, E.; Lee, G.; Han, Y.; Thaci, B.; Lesniak, M.S. Characterization and immunotherapeutic implications for a novel antibody targeting interleukin (il)-13 receptor α2. J. Biol. Chem. 2012, 287, 30215–30227. [Google Scholar] [CrossRef]
- Shimato, S.; Natsume, A.; Wakabayashi, T.; Tsujimura, K.; Nakahara, N.; Ishii, J.; Ito, M.; Akatsuka, Y.; Kuzushima, K.; Yoshida, J. Identification of a human leukocyte antigen-a24–restricted t-cell epitope derived from interleukin-13 receptor α2 chain, a glioma-associated antigen. J. Neurosurg. 2008, 109, 117–122. [Google Scholar] [CrossRef]
- Yan, X.; Su, Z.; Zhang, J.; Wu, Z.; Ye, S.; Lu, X.; Wu, J.; Zeng, Y.; Zheng, W. Killing effect of interleukin-13 receptor alpha 2 (il-13ralpha2) sensitized dc-ctl cells on human glioblastoma u251 cells. Cell. Immunol. 2010, 263, 172–175. [Google Scholar] [CrossRef]
- Iwami, K.; Shimato, S.; Ohno, M.; Okada, H.; Nakahara, N.; Sato, Y.; Yoshida, J.; Suzuki, S.; Nishikawa, H.; Shiku, H.; et al. Peptide-pulsed dendritic cell vaccination targeting interleukin-13 receptor α2 chain in recurrent malignant glioma patients with hla-a*24/a*02 allele. Cytotherapy 2012, 14, 733–742. [Google Scholar] [CrossRef]
- Okada, H.; Butterfield, L.H.; Hamilton, R.L.; Hoji, A.; Sakaki, M.; Ahn, B.J.; Kohanbash, G.; Drappatz, J.; Engh, J.; Amankulor, N.; et al. Induction of robust type-i cd8+ t-cell responses in who grade 2 low-grade glioma patients receiving peptide-based vaccines in combination with poly-iclc. Clin. Cancer Res. 2015, 21, 286–294. [Google Scholar] [CrossRef] [PubMed]
- Okada, H.; Kalinski, P.; Ueda, R.; Hoji, A.; Kohanbash, G.; Donegan, T.E.; Mintz, A.H.; Engh, J.A.; Bartlett, D.L.; Brown, C.K.; et al. Induction of cd8+ t-cell responses against novel glioma–associated antigen peptides and clinical activity by vaccinations with α-type 1 polarized dendritic cells and polyinosinic-polycytidylic acid stabilized by lysine and carboxymethylcellulose in patients with recurrent malignant glioma. J. Clin. Oncol. 2011, 29, 330–336. [Google Scholar] [PubMed]
- Debinski, W.; Obiri, N.I.; Powers, S.K.; Pastan, I.; Puri, R.K. Human glioma cells overexpress receptors for interleukin 13 and are extremely sensitive to a novel chimeric protein composed of interleukin 13 and pseudomonas exotoxin. Clin. Cancer Res. 1995, 1, 1253–1258. [Google Scholar]
- Rustamzadeh, E.; Vallera, D.A.; Todhunter, D.A.; Low, W.C.; Panoskaltsis-Mortari, A.; Hall, W.A. Immunotoxin pharmacokinetics: A comparison of the anti-glioblastoma bi-specific fusion protein (dtat13) to dtat and dtil13. J. Neuro-Oncol. 2006, 77, 257–266. [Google Scholar] [CrossRef]
- Kioi, M.; Seetharam, S.; Puri, R.K. Targeting il-13rα2-positive cancer with a novel recombinant immunotoxin composed of a single-chain antibody and mutatedpseudomonas exotoxin. Mol. Cancer Ther. 2008, 7, 1579–1587. [Google Scholar] [CrossRef]
- Candolfi, M.; Xiong, W.; Yagiz, K.; Liu, C.; Muhammad, A.K.; Puntel, M.; Foulad, D.; Zadmehr, A.; Ahlzadeh, G.E.; Kroeger, K.M.; et al. Gene therapy-mediated delivery of targeted cytotoxins for glioma therapeutics. Proc. Natl. Acad. Sci. USA 2010, 107, 20021–20026. [Google Scholar] [CrossRef]
- Rossmeisl, J.H.; Herpai, D.; Quigley, M.; Cecere, T.E.; Robertson, J.L.; D’Agostino, R.B.; Hinckley, J.; Tatter, S.B.; Dickinson, P.J.; Debinski, W. Phase i trial of convection-enhanced delivery of il13ra2 and epha2 receptor targeted cytotoxins in dogs with spontaneous intracranial gliomas. Neuro-Oncology 2021, 23, 422–434. [Google Scholar] [CrossRef]
- Ulasov, I.V.; Tyler, M.A.; Han, Y.; Glasgow, J.N.; Lesniak, M.S. Novel recombinant adenoviral vector that targets the interleukin-13 receptor alpha2 chain permits effective gene transfer to malignant glioma. Hum. Gene Ther. 2007, 18, 118–129. [Google Scholar] [CrossRef]
- Ou, W.; Marino, M.P.; Suzuki, A.; Joshi, B.; Husain, S.R.; Maisner, A.; Galanis, E.; Puri, R.K.; Reiser, J. Specific targeting of human interleukin (il)-13 receptor α2-positive cells with lentiviral vectors displaying il-13. Hum. Gene Ther. Methods 2012, 23, 137–147. [Google Scholar] [CrossRef] [PubMed]
- Allen, C.; Paraskevakou, G.; Iankov, I.; Giannini, C.; Schroeder, M.; Sarkaria, J.; Puri, R.K.; Russell, S.J.; Galanis, E. Interleukin-13 displaying retargeted oncolytic measles virus strains have significant activity against gliomas with improved specificity. Mol. Ther. 2008, 16, 1556–1564. [Google Scholar] [CrossRef]
- Madhankumar, A.B.; Slagle-Webb, B.; Mintz, A.; Sheehan, J.M.; Connor, J.R. Interleukin-13 receptor-targeted nanovesicles are a potential therapy for glioblastoma multiforme. Mol. Cancer Ther. 2006, 5, 3162–3169. [Google Scholar] [CrossRef] [PubMed]
- Madhankumar, A.B.; Slagle-Webb, B.; Wang, X.; Yang, Q.X.; Antonetti, D.A.; Miller, P.A.; Sheehan, J.M.; Connor, J.R. Efficacy of interleukin-13 receptor-targeted liposomal doxorubicin in the intracranial brain tumor model. Mol. Cancer Ther. 2009, 8, 648–654. [Google Scholar] [CrossRef] [PubMed]
- Rozhkova, E.A.; Ulasov, I.; Lai, B.; Dimitrijevic, N.M.; Lesniak, M.S.; Rajh, T. A high-performance nanobio photocatalyst for targeted brain cancer therapy. Nano Lett. 2009, 9, 3337–3342. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Rozhkova, E.A.; Ulasov, I.V.; Bader, S.D.; Rajh, T.; Lesniak, M.S.; Novosad, V. Biofunctionalized magnetic-vortex microdiscs for targeted cancer-cell destruction. Nat. Mater. 2010, 9, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Bartolomé, R.A.; Martín-Regalado, Á.; Jaén, M.; Zannikou, M.; Zhang, P.; de Los Ríos, V.; Balyasnikova, I.V.; Casal, J.I. Protein tyrosine phosphatase-1b inhibition disrupts il13rα2-promoted invasion and metastasis in cancer cells. Cancers 2020, 12, 500. [Google Scholar] [CrossRef] [PubMed]
- Vora, P.; Venugopal, C.; Salim, S.K.; Tatari, N.; Bakhshinyan, D.; Singh, M.; Seyfrid, M.; Upreti, D.; Rentas, S.; Wong, N.; et al. The rational development of cd133-targeting immunotherapies for glioblastoma. Cell Stem Cell 2020, 26, 832–844. [Google Scholar] [CrossRef] [PubMed]
- Bukur, J.; Jasinski, S.; Seliger, B. The role of classical and non-classical hla class i antigens in human tumors. Semin. Cancer Biol. 2012, 22, 350–358. [Google Scholar] [CrossRef]
- Montemurro, N.; Pahwa, B.; Tayal, A.; Shukla, A.; De Jesus Encarnacion, M.; Ramirez, I.; Nurmukhametov, R.; Chavda, V.; De Carlo, A. Macrophages in recurrent glioblastoma as a prognostic factor in the synergistic system of the tumor microenvironment. Neurol. Int. 2023, 15, 595–608. [Google Scholar] [CrossRef]
- Lv, Y.; Zhang, S.; Liu, Z.; Tian, Y.; Liang, N.; Zhang, J. Prognostic value of preoperative neutrophil to lymphocyte ratio is superior to systemic immune inflammation index for survival in patients with glioblastoma. Clin. Neurol. Neurosurg. 2019, 181, 24–27. [Google Scholar] [CrossRef]
- Lin, G.L.; Nagaraja, S.; Filbin, M.G.; Suvà, M.L.; Vogel, H.; Monje, M. Non-inflammatory tumor microenvironment of diffuse intrinsic pontine glioma. Acta Neuropathol. Commun. 2018, 6, 51. [Google Scholar] [CrossRef]
- Chheda, Z.S.; Kohanbash, G.; Okada, K.; Jahan, N.; Sidney, J.; Pecoraro, M.; Yang, X.; Carrera, D.A.; Downey, K.M.; Shrivastav, S.; et al. Novel and shared neoantigen derived from histone 3 variant h3.3k27m mutation for glioma t cell therapy. J. Exp. Med. 2018, 215, 141–157. [Google Scholar] [CrossRef] [PubMed]
- Mueller, S.; Taitt, J.M.; Villanueva-Meyer, J.E.; Bonner, E.R.; Nejo, T.; Lulla, R.R.; Goldman, S.; Banerjee, A.; Chi, S.N.; Whipple, N.S.; et al. Mass cytometry detects h3.3k27m-specific vaccine responses in diffuse midline glioma. J. Clin. Investig. 2020, 130, 6325–6337. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Xiao, X.; Wang, Y.; Gu, G.; Li, T.; Wang, Y.; Li, C.; Zhang, P.; Ji, N.; Zhang, Y.; et al. Expression of Interleukin-13 Receptor Alpha 2 in Brainstem Gliomas. Cancers 2024, 16, 228. https://doi.org/10.3390/cancers16010228
Li X, Xiao X, Wang Y, Gu G, Li T, Wang Y, Li C, Zhang P, Ji N, Zhang Y, et al. Expression of Interleukin-13 Receptor Alpha 2 in Brainstem Gliomas. Cancers. 2024; 16(1):228. https://doi.org/10.3390/cancers16010228
Chicago/Turabian StyleLi, Xiaoou, Xiong Xiao, Yi Wang, Guocan Gu, Tian Li, Yi Wang, Chunzhao Li, Peng Zhang, Nan Ji, Yang Zhang, and et al. 2024. "Expression of Interleukin-13 Receptor Alpha 2 in Brainstem Gliomas" Cancers 16, no. 1: 228. https://doi.org/10.3390/cancers16010228
APA StyleLi, X., Xiao, X., Wang, Y., Gu, G., Li, T., Wang, Y., Li, C., Zhang, P., Ji, N., Zhang, Y., & Zhang, L. (2024). Expression of Interleukin-13 Receptor Alpha 2 in Brainstem Gliomas. Cancers, 16(1), 228. https://doi.org/10.3390/cancers16010228