
Experimental Murine Models for Colorectal Cancer Research



Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
3. Carcinogen-Induced Models (CIMs)
3.1. 1,2-Dimethylhydrazine (DMH)
DMH and Colitis-Associated CRC (CAC) Models
3.2. Azoxymethane (AOM)
AOM and Colitis-Associated CRC (CAC) Models
3.3. Heterocyclic Amines (HCAs)
3.4. Aromatic Amines
3.5. Alkylating Substances

{kind=link}
{kind=link}
| Carcinogen | Animal Strain and Gender | Dose/Route | Latency Period | Tumor Characteristics | Ref. |
|---|---|---|---|---|---|
| DMH | Swiss albino mice | 10 mg/kg b.w./wk, s.c. | 17 wks | Hyperplasia with irregular-shaped mucosa, distorted crypts and laminar cellular infiltration (CD31 and Vegf) | [69] |
| Female Wistar rats | 20 mg/kg b.w./wk, s.c. | 30 wks | Adenocarcinoma; ACF, MDF and disintegration of goblet cells (NF-κB, iNOS, β-catenin, PCNA, COX-2, Bax, cleavedPARP, Bcl-2, Apc) | [70] | |
| Male Wistar albino rats | 20 mg/kg b.w./wk, s.c. or i.r. | 15 wks | Tumor cells indicative of anaplasia, dysplasia and hyperchromasia in the lumen (Krt20, SOD, CAT, Bax Bcl-2, caspase-3, cytochrome C, iNOS, TNF-α/β, IL-1β and COX-2) | [71] | |
| Male Balb/C mice | 20 mg/kg, b.w./wk, s.c. | 30 wks | Adenomas and adenocarcinomas | [72] | |
| Male Wistar rats | 40 mg/kg b.w./2 times a wk, s.c. | 20 wks | Signet-ring cell carcinoma (p53, PI3K-Akt, IKK/NF-κB, MAPK and intrinsic apoptotic signaling pathways bioinformatics analysis) | [73] | |
| Female CD1 Swiss albino mice | 20 mg/kg b.w./wk, s.c. | 20 wks | Tubular adenoma, dysplasia and anal squamous cell carcinoma (inflammation markers (IL-17, IL-10, TGF-β)) | [74] | |
| Male albino Balb/c mice | 20 mg/kg, b.w./wk, i.p. | 24 wks | Adenoma and adenocarcinoma (Wnt pathway, COX-2, iNOS) | [75] | |
| Male Fisher rats | 35 mg/Kg, b.w., o.g. | 78 wks | Adenocarcinoma | [76] | |
| C57B1/6J and mice | 10, 20 and 50 mg/kg b.w., i.p. or o.g. | 24 h | Nuclear aberration (NA) | [77] | |
| Male Wistar rats | 40 mg/kg b.w., i.p./wk | 10 wks | Wnt signalling pathway (e.g., β-catenin and p53), cell regulation (e.g., c-Myc and cyclin D1), inflammation (e.g., IL-6, ROS and COX-2) and alterations of bacterial enzymes (e.g., β-glucuronidase and β-glucosidase) | [78] | |
| DMH/TNBS | Male Wistar rats | DMH (40 mg/kg b.w./ 2 times a wk, s.c., 2 wks); TNBS (10 mg in 0.25 mL of 50% ethanol (v/v), i.r.) | 25 wks | Adenocarcinoma (Ki-67, β-catenin, Cx43, Msh6, Ppara, Akt3, Dlc1 and Vegfd) | [39] |
| DMH/DSS | Male Wistar rats | DMH (30 mg/kg b.w./single dose, i.p.; 1 week after 2% (w/v); DSS in drinking water for 7 days | 18 wks | Adenoma (apoptosis-associated p53/Bcl-2/Bax signaling) | [79,80] |
| Male BALB/c mice | DMH (20 mg/kg b.w./wk, i.p., wks 0, 3 and 6); DSS (3% w/v, 3 cycles) for 7 days (2 wks gaps) | 10 wks | Aberrant crypts, loss of goblet cells and increased cell infiltration (SOD, Nrf2, NF-κB, Caspase-1, STAT-3 and IL-6 expression) | [81] | |
| Male F344 rats | DMH (40 mg/kg b.w./3 times a wk, i.p.); DSS (2% in drinking water) for 1 wk | 10 wks | Preneoplastic ACF and MDF (SOD, Bcl-2, p53, Bax and caspase-3 expression) | [82] | |
| AOM | Female A/J mice | 10 mg/kg b.w./wk, s.c. | 16 wks | (Hif-1a, Aldoa, Pgk1, Raptor, Dek and Vegf expression) | [83] |
| Male C57BL/6 mice | 10 mg/kg b.w./single dose, i.p. | 9 wks | Adenoma (Ki-67 and PCNA protein expression; IFN-γ, IL-6, TNF-α, Th1 and Th17) | [84] | |
| Balb/c mice | 15 mg/kg b.w./single dose, i.p. | 8–9 wks | Adenoma and adenocarcinoma (pro-apoptotic (cytochrome C, DR4, DR5, TNFRSF1A, Bax and BAD) and anti-apoptotic proteins (Hsp70, Hsp32, and XIAP)) | [85] | |
| Male Sprague Dawley rats | 7 mg/kg b.w./wk, s.c. | 8 wks | ACF dysplastic and hyperplastic | [86] | |
| A/J mice | 8 mg/kg b.w./wk, i.p. | 12 wks | Adenoma–carcinoma sequence | [87] | |
| Male Balb/c mice | 10 mg/kg b.w./wk, i.p. | 25 wks | Adenocarcinoma (PI3K/Akt/mTOR pathway) | [88] | |
| Male Wistar rats | 15 mg/kg b.w./wk, s.c.. | 37 wks | Adenoma and adenocarcinoma (metastases-associated in colon cancer 1 (MACC1)) | [89] | |
| C57BL/6J and KKAy | (10 mg/kg b.w./wk, i.p. | 6 wks | Polyps, adenocarcinomas and ACF | [90] | |
| Male Wistar rats | 15 mg/kg b.w./wk, s.c. | 2 wks | Numerous large ACF with hyperplastic and dysplastic features, precancerous mucin-depleted foci (MDF) and multiple tubular adenomas | [44] | |
| A/J mice | 10 mg/kg b.w./wk, i.p. | 6 wks | Multiple tubular adenoma (overexpression of Hif-1a, Aldoa, Pgk1 and Vegf genes) | [83] | |
| AOM/DSS | Male C57BL/6 mice | AOM (12.5 mg/kg b.w./single dose, i.p.); DSS (2.5% in drinking water) for 5 days at wks 2, 6 and 9 | 12 wks | Adenoma (inflammation markers (IL-1β, IL-8, IL-10, TNF-α), claudin-1, β-actin, NF-κB and p38 MAPK pathways) | [91] |
| Male C57BL/6 mice | AOM (10 mg/kg b.w./single dose, i.p.); DSS (2.5% in drinking water) for 1 wk at wks 2, 5 and 6 | 10 wks | Adenoma (Inflammation markers (IL-6, IL-1β, COX-2 and TNF-α), cell-proliferation marker Ki67, tight junction proteins (ZO-1 and occludin) and Wnt/β-catenin pathway) | [92] | |
| Female Balb/C and C57/Bl6 mice | AOM (12.5 mg/kg b.w./single dose, i.p.); DSS (1, 2, or 3% (w/v) in drinking water) for 5 days at wks 2, 5 and 8 | 12 wks | Carcinomas (3% DSS) (cell-proliferation marker Ki67) | [93] | |
| Female FVB/NJ mice | AOM (10 mg/kg b.w./single dose, i.p.); DSS (3% in drinking water, 2 cycles) for 7 days | 8 wks | Adenoma (cell-proliferation marker Ki67; inflammation markers (IL-6, IL-10, IL-22, IL-1β, IL-17α and TNF-α) | [94] | |
| Male F344 rats | AOM (15 mg/kg b.w./1 time a wk, i.p., 3 wks); DSS (3% in drinking water, 2 cycles) for 7 days | 21 wks | Adenocarcinoma (microbiome-community phylogenetic analysis) | [95] | |
| Male Wistar rats | AOM (10 mg/kg b.w./1 time a wk, s.c., 2 wks); DSS (4% in drinking water, 2 cycles) for 7 days | 10 wks | Adenoma and adenocarcinoma (inflammation markers (IL-6, IL-10, COX-2, NF-κB) and Wnt/β-catenin signaling pathway) | [96] | |
| Lgr5 eGFP-IRES-CreERT2 mice | AOM (10 mg/kg b.w., i.p.); DSS (2% in drinking water; 3 cycles) for 5 days | 11 wks | Adenoma (Ly6a (Sca-1), Tacstd2 (Trop2) and Sox9 gene expression | [97] | |
| AOM/TNBS | C57BL/6 mice | AOM (10 mg/kg b.w./single dose, i.p.); 2.5 mg of TNBS (150 µL 50% EtOH) i.r. | NR | Extensive inflammatory, dysplasia or carcinoma lesions all over entire mucosa with numerous ulcers (TNF-α, IFN-γ, IL-1β and anti-inflammatory cytokines IL-10 and IL-12) | [47] |
| IFN-γ−/− and IL-4−/− mice | AOM (10 mg/kg b.w./1 time a wk, i.p., 3–6 wks); TNBS (2% PBS:ethanol (1:1), i.r., 3–10 wks) | 33 wks | Adenocarcinomas (p53, β-catenin, Th1 and Th2) | [35] | |
| PhIP | hCYP1A mice | PhIP (0.01–200 mg/kg b.w., o.g. DSS (1.5% (w/v) in drinking water for 5 days) | 8 wks | Adenoma (p53 signaling network and regulatory pathways) | [98] |
| hCYP1A mice | PhIP (100 mg/kg b.w./2 doses, i.g. with 3 days apart); DSS (1.5% (w/v) in drinking water for 4 days) | 10 wks | Adenocarcinoma (oxidative and nitrosative stress markers (8-oxo-dG and nitrotyrosine) and inflammation markers (NF-κB and p-STAT3) | [99] | |
| MNNG | Female C57BL6 mice | 100 mg/kg b.w., i.r. | 12 wks | Adenoma–carcinoma sequence (endoscopic evaluation) | [25] |
| Male BALB/c mice | 4 successive dosages (5 mg/mL; i.r. deposits of 100 µL, twice a wk for 2 wks | 10 wks | Adenocarcinoma (PCNA, COX-2, IL-12, IL-10, TNF-α and INF-γ) | [100] | |
| Male C57/BL6 mice | |||||
| Male IL-10−/− mice | |||||
| Female C57BL/6 mice | 4 successive dosages (5 mg/mL; i.r. deposits of 100 µL, twice a wk for 2 wks | 8 wks | Adenoma and adenocarcinoma (PCNA, Ki67, c-Myc, Vegf, CD133, CD34 and CD31) | [101] | |
| MNU | Male albino Wistar rats | 1.2% in 1.9% citric acid, i.r. | 12 wks | Adenoma (MLH-1 and SOD) | [102] |
| Male Wistar rats | 8 mg/kg b.w., 3 times a wk, 4 wks, i.r. | 25 wks | Adenocarcinoma and signet ring cell carcinoma (Kras, Ki67 and caspase-3 expression; IFN-γ, IL-1β, IL-8, TGF-β, TNF-α and IL-6; Wnt-Apc-β-catenin pathway) | [103] | |
| Female Sprague Dawley rats | 10 mg/kg b.w., 3 times a wk, 4 wks, i.r. | NR | Adenoma (PI3K/AKT/Bcl-2 pathway) | [104] | |
| Male Wistar rats | 8 mg/kg b.w., 5 times a wk, 6 wks, i.r. | 8 wks | FRZ-8, GAPDH, Apc gene expression, Wnt-Apc-β-catenin pathway | [105] | |
| Male Sprague-Dawley rats | 8 mg/kg b.w., 3 times a wk, 5 wks, i.r. | 16 and 24 wks | 16 wks—Adenoma; 24 wks—Adenocarcinoma (Wnt/β-catenin and Notch pathways) | [106,107] | |
| Male F344/DuCrj rats | 8 mg/kg b.w., 3 times a wk, 4 wks, i.r. | 20 wks | ACF (PCNA) | [108] |
4. Genetically Engineered Murine Models (GEMMs)
4.1. Adenomatous Polyposis Mouse Models (APMM)
4.2. Hereditary Nonpolyposis Colon Cancer Mouse Models (HNPCC)
| GEMM | Outcome(s) | Advantages | Disadvantages | Ref. |
|---|---|---|---|---|
| All | Evaluate the role of genes involved in carcinogenesis; Studies of chemoprevention and therapeutic agents; Assessing the influence of carcinogens; Lifestyle/dietary influence on tumor formation. | Genetic event is known; In situ tumor development; Reproduces early stages of oncogenesis; Modified gene is expressed on physiologic level; Tumor cells and stroma are from the same specie; Intact immune system. | Limited options for non-invasive imaging (would need CT/MRI capability); Expensive and time consuming to develop; Only partial replication of the human tumoral morphology and physiology; Secondary mutations are different from the human tumors; Low metastases rate. | [50,136] |
| Apc580S | Adenoma formation in the distal rectum in most of the Apc 580S homozygotes within 4 weeks after infection by rectal infusion with recombinant adenoviruses encoding the Cre recombinase. In total, 50% of animals show invasive adenocarcinoma after 1 year without lymphatic or distant metastases. | Useful for studying the mechanism of CRC development and to test therapeutic or chemopreventative agents. | Only effective in Apc 580S/580S mice and not Apc 580S/+, an outcome that reflects the poor ability of the approach to influence the proliferating cells at the crypt base. | [137] |
| CAC; APC580S/+ | Adenomatous lesions in the distal colon; DSS treatment increased the incidence and number of tumors, and this occurred predominantly in distal colon. | Mimics the tissue and cellular environment of heritable cancers such as FAP and LS. | Early CRC development may limit the ability to test therapeutic or chemopreventative agents; increased animal numbers for CRC studies. | [138] |
| ApcMin/+ Mom1R/R P53−/− | p53 deficiency increases intestinal adenoma multiplicity and malignancy. | p53-deficient tumors studies | Short lifespan (122 days). | [139] |
| ApcMin/+ Mom1R/s P53−/− | ||||
| K-RasG12D | Adenocarcinomas expressing invariably exhibit uniform high-grade dysplasia | KRAS signaling pathway studies | Do not develop metastases. | [140] |
| Pik3caH1047R | Develop invasive adenocarcinomas strikingly similar to invasive adenocarcinomas found in human CRC. | PI3K/AKT/mTOR pathway therapeutic studies | Late CRC | [141] |
| Msh2−/− | Development of colorectal tumors with defects in DNA mismatch repair. | Model of LS (3% of all CRCs) | Msh2 mutation in all cells of body and mice are predisposed to lymphomas. | [142] |
| Smad4TKO | Development of colorectal tumors with loss of function mutations in the tumor-suppressor gene Smad4. | IFN-γ expression correlates with the onset of spontaneous CAC by 6 months of age. | Do not develop metastases. | [143] |
| ApcCKO/LSL-Kras | Cre-mediated knockout of Apc and KrasG12D activation by surgical application of AdenoCre to the colonic epithelium leads to tumor formation after 3 weeks and adenocarcinomas with 20% liver metastases after 20 weeks. | FAP and LS genetic mutations are present in the germline; mTOR Pathway and metastatic model. | 20–24 weeks for metastases development. | [144] |
| Villin-Cre/K-rasG12Dint/Ink4a/Arf−/− | Most invasive adenocarcinomas (79%) progress within 12 weeks, and 60% of these tumors metastasize to the lungs. | Use tissue specific promoters in intestinal mucosa to target gene knockout; Some invasive adenocarcinomas seen can be used to target specific tumor-suppressor or oncogenes. | Requires rectal instillation of recombinant adenovirus expressing Cre. | [145] |
| Villin-Cre; LSL-KrasG12D/+ | ||||
| Villin-Cre; KrasG12Dint | ||||
| Lgr5CreERT2 | Hyperproliferating intestinal adenomas were formed 4 weeks after tamoxifen injection. | CDX models (HCT-116 or SW480 cells); Wnt/β- catenin pathway | Do not develop metastases. | [146] |
| β-cateninexon3 | ||||
| Rosa26LSL-rtta-ires-EGFP | ||||
| TRE-Spdef | ||||
| Apc1638N/++ AOM | A 6-fold increase in colonic tumor formation compared to Apc Min/+ mice; higher incidence of colonic adenocarcinomas. | Increased the tumor burden in the colon; Suitable and straightforward model to study the influence of immune cells and chemokines on colon carcinogenesis. | Do not develop metastases. | [120] |
| Apc Min/+ + PhIP | Increased tumor development by 2- to 3-fold compared to Apc Min/+ mice | Ideal gene expression for FAP studies | Do not develop metastases; Most of the tumors are in the small intestine. | [147] |
| ApcΔ716 Tgfbr2flox/flox; villin-CreER + DSS (2%) | TGF-signaling disruption include the development of adenocarcinomas with a local invasion pattern | Ideal for CAC CRC studies | No metastases reported. | [148] |
| ApcΔ716 KrasG12D | Increased multiplicity of intestinal tumors | Metastatic model; PDOX model; Efficient metastases by Wnt activation, Kras activation, and TGFβ suppression combination. | No spontaneous metastases. | [149] |
| ApcΔ716 Trp53R270H | Developed adenocarcinomas with invasion to submucosa or deeper | |||
| ApcΔ716KrasG12DFbxw7−/− | Distinct histologic type and accelerated tumorigenesis | |||
| ApcΔ716KrasG12DTgfbr−/− | Efficient liver metastases | |||
| Dpc4+/−: Apc+/Δ716 | Submucosal infiltration and a progression from adenoma to carcinoma can be seen in the small intestine and colon of Dpc4 and ApcΔ716cis-compound heterozygote mice | Ideal for FAP CRC studies (Histological features of tumors are identical) | Do not develop metastases. | [150] |
| Fen1null/Apc1638N | Increased intestinal tumor malignancy via MSI comparatively to Apc1638N mice | FAP and LS studies with Fen1 gene | Do not develop metastases. | [151] |
| Fbw7flox/flox; P53flox/flox; Villin-Cre | Allografts derived from tumors with a double deletion of Fbw7 and p53 develop into highly malignant adenocarcinomas with a high rate of metastases | Important tool for future studies of the pathogenesis and treatment of metastatic and chromosomally unstable CRC. | Long latency period (up to 101 weeks) | [152] |
| AhCre+/T; Kras+/LSLV12, Apc+/fl | Although KrasV12 mutation does not affect the intestinal epithelium, it accelerates tumorigenesis when combined with Apc loss. Invasive adenocarcinomas make up 17% of all tumors | Suitable for Raf-MEK-ERK pathway studies | Do not develop metastases. | [153] |
| Pms2 ki/ki | A ∼4.5-fold increase in intestinal polyp formation compared to Apc+/− or Pms2ki/+; Apc+/− mice | LS studies with MMR genes; Suppression of de novo splice site. | Do not develop metastases. | [135] |
| BRAF-V600E | Promotes rapid serrated tumor development and progression and assesses the role of Smad4 in early-stage serrated tumorigenesis | Oncogenic β-catenin mutations (combinations of Ctnnb1, Braf, and Smad4) drive rapid serrated dysplasia formation. | Do not develop metastases. | [154] |
5. Transplant and Metastatic Murine Models
5.1. Transplant Murine Models (TMMs)
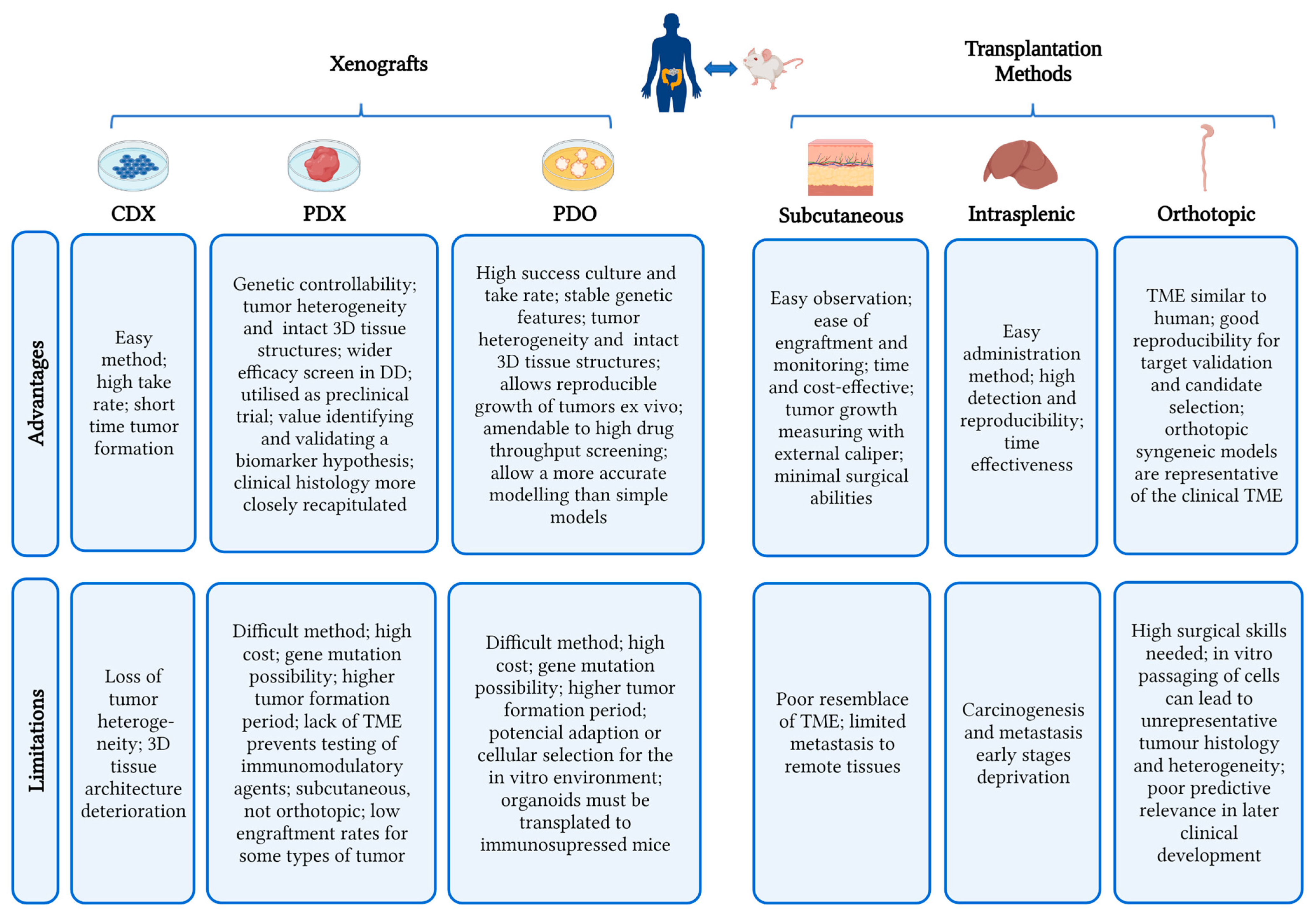
5.1.1. Cell-Derived Xenografts (CDX) Models
5.1.2. Patient-Derived Xenograft (PDX) Models
5.1.3. Patient-Derived Organoid Xenograft (PDOX) Models
5.2. Metastases Models
| Model | Predominant Histopathology | Metastases and Main Location | Ref. |
|---|---|---|---|
| Carcinogen-induced Models(CIM) | |||
| Tp53ΔIEC + AOM | Adenocarcinoma | Lymph nodes | [241] |
| LSL-KrasG12D/+; p53 flox/flox+sgApc-Cas9-Cre | Adenocarcinoma | Lymph nodes and liver | [242] |
| Genetically Engineered Models (GEMMs) | |||
| ApcCKO/CKOLSL-G12D; Kras tm4tyj/+ | Adenocarcinoma | Lymph nodes and liver | [144] |
| ApcLox/Lox; p53Lox/Lox; Tet-O-LSL-KrasG12D; VillinCreERT2 | Adenocarcinoma | Lymph nodes, liver, and lungs | [243] |
| Villin-CreERT2 Apc fl/fl | Adenocarcinoma | Lymph nodes | [179] |
| LSL-KRASG12V/APCflox/flox | Adenocarcinoma | Lymph nodes and liver | [244] |
| Cell-derived Xenografts (CDXs) | |||
| NSG mice + HT29p53-mut/LUC cells | Adenocarcinoma | Lymph nodes, liver, lungs, and bone marrow. | [3] |
| Balb/c (i.c.) + CT-26 cells | Carcinoma | No metastases | [245] |
| NOD/SCID (i.c.) + HCT-116 cells | Adenocarcinoma | Liver | [234] |
| Balb/c nude mice (s.c.) + HCT15 cells | Adenocarcinoma | NR | [246] |
| Balb/c nude mice (s.c.) + HCT-116 cells | Adenocarcinoma | NR | [247] |
| Balb/c nude mice (i.v.) +HCT-116-Luc cells | Adenocarcinoma | Lungs | [248] |
| C57BL/6J mice (s.c.) + MC38 cells | NR | ||
| Traj18−/− (s.c., i.c.) + MC38 cells | Adenocarcinoma | NR | [249] |
| CD1d−/− (s.c., i.c.) + MC38 cells | Adenocarcinoma | NR | |
| NSG mice (i.c.; i.s.) + SW480 cells | Adenocarcinoma | Liver | [250] |
| NSG mice (i.c.; i.s.) + SW620 cells | Adenocarcinoma | Liver | [250] |
| Balb/c nude mice (i.c.) + SW620 cells | NR | Liver | [187] |
| Patient-derived Xenografts (PDX) | |||
| NSG mice (i.s.) | Adenocarcinoma and carcinoma | Lymph nodes, liver and lungs. | [251] |
| Balb/c mice (i.s.) | Adenocarcinoma | Liver | [240] |
| Balb/c nude mice (i.c.) | NR | Liver | [187] |
| Balb/c nude mice (s.c.) | Adenocarcinoma | NR | [248] |
| NCG mice (i.v.) | Adenomas and carcinomas | Liver and lungs. | [252] |
| NOD/SCID mice (i.c.) | Adenocarcinoma | Lungs | [253] |
| NOD/SCID mice (s.c.) | Carcinoma | NR | [254] |
| NCG mice (s.c.) | Adenocarcinoma | NR | [255] |
| Patient-derived Organoids Xenografts (PDOXs) | |||
| Balb/c-nu mice (i.s.) | Macrometastatic colonies | Liver and lungs. | [236] |
| NOG mice (i.s.) | Micro- and macrometastatic colonies | Liver | [256] |
| NSG mice (i.s.) | Macrometastatic colonies | Liver | [257] |
| NSG mice (s.c.; i.c.; i.s.) | Micro- and macrometastatic colonies | Liver | [250] |
6. Meeting the Criteria for a Successful Murine Model for Colorectal Cancer Investigation
7. Future Perspectives
8. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ACF | Aberrant crypt foci |
| AOM | Azoxymethane |
| APMM | Adenomatous Polyposis Mouse Models |
| b.w. | Body weight |
| CAC | Colitis-associated CRC |
| CD | Crohn’s disease |
| CDX | Cell-derived xenografts |
| CIMs | Carcinogen-induced models |
| CRC | Colorectal cancer |
| CTCs | Circulating tumor cells |
| DMAB | 3,2-dimethyl-4-aminobiphenyl |
| DMH | 1,2-dimethylhydrazine |
| DSS | dextran sulphate sodium |
| FAP | Familial adenomatous polyposis |
| GEMMs | Genetically Engineered Murine Models |
| H | Hours |
| HCAs | Heterocyclic amines |
| HNPCC | Hereditary Nonpolyposis Colon Cancer Mouse Models |
| IBD | Inflammatory bowel disease |
| i.c. | Intra-caecal |
| i.g. | Intragastric gavage |
| i.m. | Intramuscular |
| i.p. | Intraperitoneal |
| i.r. | Intrarectal |
| i.v. | Intravenous |
| IQ | 2-amino-3-methylimidazo[4,5-f]quinoline |
| LS | Lynch syndrome |
| MAM | Methylazoxymethanol |
| MM | Metastatic Models |
| MNNG | N-methyl-N-nitro-N-nitrosoguanidine |
| MNU | Methylnitrosourea |
| mos | Months |
| MSI | Microsatellite instability |
| NA | Nuclear aberration |
| NF-κB | nuclear factor kappa B |
| NR | Not reported |
| OoC | Organs-on-chips |
| o.g. | Oral gavage |
| PDO | Patient-derived organoids |
| PDOX | Patient-derived organoid xenografts |
| PDX | Patient-derived xenografts |
| PhIP | 2-amino-1-methyl-6-phenylimidazo [4,5-b] pyridine |
| s.c. | Subcutaneous |
| sCRC | Sporadic colorectal cancer |
| TME | Tumor microenvironment |
| TMMs | Transplant metastatic models |
| TNBS | 2,4,6-Trinitrobenzenesulfonic acid |
| UC | Ulcerative colitis |
| wk | Week |
| wks | Weeks |
References
- GLOBOCAN. Colorectal Cancer Incidence in The World. Glob. Cancer Obs. 2020, 419, 1–2. [Google Scholar]
- Rabeneck, L.; Chiu, H.M.; Senore, C. International Perspective on the Burden of Colorectal Cancer and Public Health Effects. Gastroenterology 2020, 158, 447–452. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, S.; Behring, M.; Kim, H.-G.; Bajpai, P.; Chakravarthi, B.V.S.K.; Gupta, N.; Elkholy, A.; Al Diffalha, S.; Varambally, S.; Manne, U. Targeting P4HA1 with a Small Molecule Inhibitor in a Colorectal Cancer PDX Model. Transl. Oncol. 2020, 13, 100754. Available online: https://www.sciencedirect.com/science/article/pii/S1936523320301030 (accessed on 23 March 2023). [CrossRef] [PubMed]
- American Cancer Society. Colorectal Cancer Early Detection, Diagnosis, and Staging. Cancer.org. Available online: https://www.cancer.org/content/dam/CRC/PDF/Public/8606.00.pdf (accessed on 1 March 2023).
- Percario, R.; Panaccio, P.; Francesco, F.; Grottola, T.; Sebastiano, P.D.I. The Complex Network between Inflammation and Colorectal Cancer: A Systematic Review of the Literature. Cancers 2021, 13, 6237. [Google Scholar] [CrossRef] [PubMed]
- Ahmad Kendong, S.M.; Raja Ali, R.A.; Nawawi, K.N.M.; Ahmad, H.F.; Mokhtar, N.M. Gut Dysbiosis and Intestinal Barrier Dysfunction: Potential Explanation for Early-Onset Colorectal Cancer. Front. Cell. Infect. Microbiol. 2021, 11, 744606. Available online: https://www.frontiersin.org/articles/10.3389/fcimb.2021.744606/full (accessed on 22 January 2022). [CrossRef]
- Alzahrani Mohammad, S.; Al Doghaither Abdulaziz, H.; Al-Ghafari Badr, A. General insight into cancer: An overview of colorectal cancer (Review). Mol. Clin. Oncol. 2021, 15, 271. [Google Scholar] [CrossRef]
- Nguyen, L.H.; Goel, A.; Chung, D.C. Pathways of Colorectal Carcinogenesis. Gastroenterology 2020, 158, 291–302. [Google Scholar] [CrossRef]
- Taleban, S.; Elquza, E.; Gower-Rousseau, C.; Peyrin-Biroulet, L. Cancer and inflammatory bowel disease in the elderly. Dig. Liver Dis. 2016, 48, 1105–1111. [Google Scholar] [CrossRef]
- Direito, R.; Lima, A.; Rocha, J.; Ferreira, R.B.; Mota, J.; Rebelo, P.; Fernandes, A.; Pinto, R.; Alves, P.; Bronze, R.; et al. Dyospiros kaki phenolics inhibit colitis and colon cancer cell proliferation, but not gelatinase activities. J. Nutr. Biochem. 2017, 46, 100–108. [Google Scholar] [CrossRef]
- Frigerio, S.; Lartey, D.A.; D’Haens, G.R.; Grootjans, J. The Role of the Immune System in IBD-Associated Colorectal Cancer: From Pro to Anti-Tumorigenic Mechanisms. Int. J. Mol. Sci. 2021, 22, 12739. [Google Scholar] [CrossRef]
- Marabotto, E.; Kayali, S.; Buccilli, S.; Levo, F.; Bodini, G.; Giannini, E.G.; Savarino, V.; Savarino, E.V. Colorectal Cancer in Inflammatory Bowel Diseases: Epidemiology and Prevention: A Review. Cancers 2022, 14, 4254. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.S.; Liu, C.Y.; Wen, D.; Gao, D.Z.; Lin, S.; He, H.F.; Zhao, X.F. Recent advances in the development of transplanted colorectal cancer mouse models. Transl. Res. 2022, 249, 128–143. [Google Scholar] [CrossRef] [PubMed]
- Lotfollahzadeh, S.; Recio-Boiles, A.; Cagir, B. Colon Cancer; StatPearls Publishing: Tampa, FL, USA, 2022. Available online: https://www.ncbi.nlm.nih.gov/books/NBK470380/ (accessed on 1 March 2023).
- Lannagan, T.R.; Jackstadt, R.; Leedham, S.J.; Sansom, O.J. Advances in colon cancer research: In vitro and animal models. Curr. Opin. Genet. Dev. 2021, 66, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Tsitskari, M.; Filippiadis, D.; Kostantos, C.; Palialexis, K.; Zavridis, P.; Kelekis, N.; Brountzos, E. The role of interventional oncology in the treatment of colorectal cancer liver metastases. Ann. Gastroenterol. 2019, 32, 147–155. [Google Scholar] [CrossRef]
- McQuade, R.M.; Stojanovska, V.; Bornstein, J.C.; Nurgali, K. Colorectal Cancer Chemotherapy: The Evolution of Treatment and New Approaches. Curr. Med. Chem. 2017, 24, 1537–1557. [Google Scholar] [CrossRef]
- Tam, S.Y.; Wu, V.W.C. A review on the special radiotherapy techniques of colorectal cancer. Front. Oncol. 2019, 9, 208. [Google Scholar] [CrossRef]
- Xie, Y.H.; Chen, Y.X.; Fang, J.Y. Comprehensive review of targeted therapy for colorectal cancer. Signal Transduct. Target. Ther. 2020, 5, 22. [Google Scholar] [CrossRef]
- Cappell, M.S. Pathophysiology, Clinical Presentation, and Management of Colon Cancer. Gastroenterol. Clin. N. Am. 2008, 37, 1–24. [Google Scholar] [CrossRef]
- ACS. Colorectal Cancer Facts and Figures 2020–2022. Am. Cancer Soc. 2020, 66, 1–41. Available online: https://www.cancer.org/content/dam/cancer-org/research/cancer-facts-and-statistics/colorectal-cancer-facts-and-figures/colorectal-cancer-facts-and-figures-2020-2022.pdf (accessed on 20 January 2022).
- Song, M.; Chan, A.T.; Sun, J. Influence of the Gut Microbiome, Diet, and Environment on Risk of Colorectal Cancer. Gastroenterology 2020, 158, 322–340. [Google Scholar] [CrossRef]
- Rubio, C.A. Three pathways of colonic carcinogenesis in rats. Anticancer Res. 2017, 37, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Oh, B.Y.; Hong, H.K.; Lee, W.Y.; Cho, Y.B. Animal models of colorectal cancer with liver metastasis. Cancer Lett. 2017, 387, 114–120. [Google Scholar] [CrossRef] [PubMed]
- Machado, V.F.; Parra, R.S.; Leite, C.A.; Minto, S.B.; Cunha, T.M.; Cunha, F.Q.; Garcia, S.B.; Feitosa, M.R.; Da Rocha, J.J.R.; Feres, O. Experimental model of rectal carcinogenesis induced by n-methyl-n-nitrosoguanidine in mice with endoscopic evaluation. Int. J. Med. Sci. 2020, 17, 2505–2510. [Google Scholar] [CrossRef] [PubMed]
- Mittal, V.K.; Singh Bhullar, J.; Kumar, J. Animal models of human colorectal cancer: Current status, uses and limitations. World J. Gastroenterol. 2015, 21, 11854–11861. [Google Scholar] [CrossRef]
- Rosenberg, D.W.; Giardina, C.; Tanaka, T. Mouse models for the study of colon carcinogenesis. Carcinogenesis 2009, 30, 183–196. [Google Scholar] [CrossRef]
- Yuan, C.; Zhao, X.; Wangmo, D.; Alshareef, D.; Gates, T.J.; Subramanian, S. Tumor models to assess immune response and tumor-microbiome interactions in colorectal cancer. Pharmacol. Ther. 2022, 231, 107981. [Google Scholar] [CrossRef]
- Nascimento-Gonçalves, E.; Mendes, B.A.L.; Silva-Reis, R.; Faustino-Rocha, A.I.; Gama, A.; Oliveira, P.A. Animal models of colorectal cancer: From spontaneous to genetically engineered models and their applications. Vet. Sci. 2021, 8, 59. [Google Scholar] [CrossRef]
- Jackstadt, R.; Sansom, O.J. Mouse models of intestinal cancer. J. Pathol. 2016, 238, 141–151. [Google Scholar] [CrossRef]
- Venkatachalam, K.; Vinayagam, R.; Anand, M.A.V.; Isa, N.M.; Ponnaiyan, R. Biochemical and molecular aspects of 1,2-dimethylhydrazine (DMH)-induced colon carcinogenesis: A review. Toxicol. Res. 2020, 9, 2–18. [Google Scholar] [CrossRef]
- Gurley, K.E.; Moser, R.D.; Kemp, C.J. Induction of colon cancer in mice with 1,2-dimethylhydrazine. Cold Spring Harb. Protoc. 2015, 2015, 814–815. [Google Scholar] [CrossRef]
- Meng, S.; Li, Y.; Zang, X.; Jiang, Z.; Ning, H.; Li, J. Effect of TLR2 on the proliferation of inflammation-related colorectal cancer and sporadic colorectal cancer. Cancer Cell. Int. 2020, 20, 95. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Kwon, J.E.; Cho, M.L. Immunological pathogenesis of inflammatory bowel disease. Intest. Res. 2018, 16, 26–42. [Google Scholar] [CrossRef] [PubMed]
- Osawa, E.; Nakajima, A.; Fujisawa, T.; Kawamura, Y.I.; Toyama-Sorimachi, N.; Nakagama, H.; Dohi, T. Predominant T helper type 2-inflammatory responses promote murine colon cancers. Int. J. Cancer 2006, 118, 2232–2236. [Google Scholar] [CrossRef] [PubMed]
- Modesto, R.; Estarreja, J.; Silva, I.; Rocha, J.; Pinto, R.; Mateus, V. Chemically Induced Colitis-Associated Cancer Models in Rodents for Pharmacological Modulation: A Systematic Review. J. Clin. Med. 2022, 11, 2739. [Google Scholar] [CrossRef]
- Gadaleta, R.M.; Garcia-Irigoyen, O.; Moschetta, A. Exploration of Inflammatory Bowel Disease in Mice: Chemically Induced Murine Models of Inflammatory Bowel Disease (IBD). Curr. Protoc. Mouse Biol. 2017, 7, 13–28. [Google Scholar] [CrossRef]
- Kumar, S.; Agnihotri, N. Piperlongumine targets NF-κB and its downstream signaling pathways to suppress tumor growth and metastatic potential in experimental colon cancer. Mol. Cell. Biochem. 2021, 476, 1765–1781. [Google Scholar] [CrossRef]
- Fragoso, M.F.; Romualdo, G.R.; Vanderveer, L.A.; Franco-Barraza, J.; Cukierman, E.; Clapper, M.L.; Carvalho, R.F.; Barbisan, L.F. Lyophilized açaí pulp (Euterpe oleracea Mart) attenuates colitis-associated colon carcinogenesis while its main anthocyanin has the potential to affect the motility of colon cancer cells. Food Chem. Toxicol. 2018, 121, 237–245. [Google Scholar] [CrossRef]
- Megaraj, V.; Ding, X.; Fang, C.; Kovalchuk, N.; Zhu, Y.; Zhang, Q.Y. Role of hepatic and intestinal P450 enzymes in the metabolic activation of the colon carcinogen azoxymethane in mice. Chem. Res. Toxicol. 2014, 27, 656–662. [Google Scholar] [CrossRef]
- Machado, V.F.; Feitosa, M.R.; da Rocha, J.J.R.; Féres, O. A review of experimental models in colorectal carcinogenesis. J. Coloproctol. 2016, 36, 53–57. [Google Scholar] [CrossRef]
- Stastna, M.; Janeckova, L.; Hrckulak, D.; Kriz, V.; Korinek, V. Human colorectal cancer from the perspective of mouse models. Genes 2019, 10, 788. [Google Scholar] [CrossRef]
- Li, C.; Lau, H.C.H.; Zhang, X.; Yu, J. Mouse Models for Application in Colorectal Cancer: Understanding the Pathogenesis and Relevance to the Human Condition. Biomedicines 2022, 10, 1710. [Google Scholar] [CrossRef] [PubMed]
- Kensara, O.A.; El-Shemi, A.G.; Mohamed, A.M.; Refaat, B.; Idris, S.; Ahmad, J. Thymoquinone subdues tumor growth and potentiates the chemopreventive effect of 5-fluorouracil on the early stages of colorectal carcinogenesis in rats. Drug Des. Dev. Ther. 2016, 10, 2239–2253. [Google Scholar]
- Amos-Landgraf, J.M.; Kwong, L.N.; Kendziorski, C.M.; Reichelderfer, M.; Torrealba, J.; Weichert, J.; Haag, J.D.; Chen, K.S.; Waller, J.L.; Gould, M.N.; et al. A target-selected Apc-mutant rat kindred enhances the modeling of familial human colon cancer. Proc. Natl. Acad. Sci. USA 2007, 104, 4036–4041. [Google Scholar] [CrossRef] [PubMed]
- Tanimura, Y.; Fukui, T.; Horitani, S.; Matsumoto, Y.; Miyamoto, S.; Suzuki, R.; Tanaka, T.; Tomiyama, T.; Ikeura, T.; Ando, Y.; et al. Long-term model of colitis-associated colorectal cancer suggests tumor spread mechanism and nature of cancer stem cells. Oncol. Lett. 2021, 21, 7. [Google Scholar] [CrossRef]
- Xiao, Y.; Dai, X.; Li, K.; Gui, G.; Liu, J.; Yang, H. Clostridium butyricum partially regulates the development of colitis-associated cancer through miR-200c. Cell Mol. Biol. 2017, 63, 59–66. [Google Scholar] [CrossRef]
- Clapper, M.L.; Cooper, H.S.; Chang, W.C.L. Dextran sulfate sodium-induced colitis-associated neoplasia: A promising model for the development of chemopreventive interventions. Acta Pharmacol. Sin. 2007, 28, 1450–1459. [Google Scholar] [CrossRef]
- Tong, Y.; Yang, W.; Koeffler, H.P. Mouse models of colorectal cancer. Chin. J. Cancer 2011, 30, 450–462. [Google Scholar] [CrossRef]
- Evans, J.P.; Sutton, P.A.; Winiarski, B.K.; Fenwick, S.W.; Malik, H.Z.; Vimalachandran, D.; Tweedle, E.M.; Costello, E.; Palmer, D.H.; Park, B.K.; et al. From mice to men: Murine models of colorectal cancer for use in translational research. Crit. Rev. Oncol. Hematol. 2016, 98, 94–105. [Google Scholar] [CrossRef]
- National Comprehensive Cancer Network. NCCN Guidelines for Patients; National Comprehensive Cancer Network: Plymouth Meeting, PA, USA, 2022. [Google Scholar]
- De Robertis, M.; Massi, E.; Poeta, M.L.; Carotti, S.; Morini, S.; Cecchetelli, L.; Signori, E.; Fazio, V.M. The AOM/DSS murine model for the study of colon carcinogenesis: From pathways to diagnosis and therapy studies. J. Carcinog. 2011, 10, 9. [Google Scholar]
- Durmus, S.; van der Valk, M.; Teunissen, S.F.; Song, J.Y.; Wagenaar, E.; Beijnen, J.H.; Schinkel, A.H. ABC transporters Mdr1a/1b, Bcrp1, Mrp2 and Mrp3 determine the sensitivity to PhIP/DSS-induced colon carcinogenesis and inflammation. Arch. Toxicol. 2019, 93, 775–790. [Google Scholar] [CrossRef]
- Chen, J.X.; Wang, H.; Liu, A.; Zhang, L.; Reuhl, K.; Yang, C.S. PhIP/DSS-induced colon carcinogenesis in CYP1A-humanized mice and the possible role of Lgr5+ stem cells. Toxicol. Sci. 2017, 155, 224–233. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhou, H.; Liu, A.; Guo, X.; Yang, C.S. Genetic analysis of colon tumors induced by a dietary carcinogen PhIP in CYP1A humanized mice: Identification of mutation of β-catenin/Ctnnb1 as the driver gene for the carcinogenesis. Mol. Carcinog. 2016, 23, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Lorenz, E.; Stewart, H.L. Intestinal carcinoma and other lesions in mice following oral administration of 1,2,5,6-dibenzanthracene and 20- methyleholanthrene. J. Natl. Cancer Inst. 1941, 1, 17–40. [Google Scholar]
- Walpole, A.L.; Williams, M.H.; Roberts, D.C. The carcinogenic action of 4-aminodiphenyl and 3:2’-dimethyl-4-amino-diphenyl. Br. J. Ind. Med. 1952, 9, 255–263. [Google Scholar] [PubMed]
- Reddy, B.S.; Mori, H. Effect of dietary wheat bran and dehydrated citrus fiber on 3,2′-dimethyl-4-aminobiphenyl-induced intestinal carcinogenesis in F344 rats. Carcinogenesis 1981, 2, 21–25. [Google Scholar] [CrossRef]
- Ravoori, S.; Feng, Y.; Neale, J.R.; Jeyabalan, J.; Srinivasan, C.; Hein, D.W.; Gupta, R.C. Dose-dependent reduction of 3,2’-dimethyl-4-aminobiphenylderived DNA adducts in colon and liver of rats administered celecoxib. Mutat. Res. 2008, 638, 103–109. [Google Scholar] [CrossRef]
- Komala, M.; Radhika, M.N.; Anbu, J.; Pyngrope, K.R. A review on chemical models of colorectal cancer: Criteria with mechanism of carcinogenesis. Int. J. Pharm. Sci. Rev. Res. 2019, 56, 66–72. Available online: https://www.embase.com/search/results?subaction=viewrecord&id=L2002112743&from=export (accessed on 30 November 2022).
- Feng, Y.; Fretland, A.J.; Rustan, T.D.; Jiang, W.; Becker, W.K.; Hein, D.W. Higher frequency of aberrant crypt foci in rapid than slow acetylator inbred rats administered the colon carcinogen 3,2’-dimethyl-4-aminobiphenyl. Toxicol. Appl. Pharmacol. 1997, 147, 56–62. [Google Scholar] [CrossRef]
- Narisawa, T.; Weisburger, J.H. Colon cancer induction in mice by intrarectal instillation of N-methylnitosorurea (38498). Biol. Med. Proc. Soc. Exp. 1975, 148, 166–169. [Google Scholar] [CrossRef]
- Narisawa, T.; Sato, T.; Hayakawa, M.; Sakuma, A.; Nakano, H. Carcinoma of the colon and rectum of rats by rectal infusion of N-methyl-N’-nitro-N-nitrosoguanidine. Gann 1971, 62, 231–234. [Google Scholar]
- Narisawa, T.; Wong, C.Q.; Maronpot, R.R.; Weisburger, J.H. Large bowel carcinogenesis in mice and rats by several intrarectal doses of methylnitrosourea and negative effect of nitrite plus methylurea. Cancer Res. 1976, 36, 505–510. [Google Scholar] [PubMed]
- Qin, X.; Zarkovic, M.; Nakatsuru, Y.; Arai, M.; Oda, H.; Ishikawa, T. DNA adduct formation and assessment of aberrant crypt foci in vivo in the rat colon mucosa after treatment with N-methyl-N-nitrosourea. Carcinogenesis 1994, 15, 851–855. [Google Scholar] [CrossRef] [PubMed]
- Endo, T.; Ookawa, K.; Tanaka, M.; Nakaji, S.; Tsuchida, S.; Sugawara, K. Differences in carcinogenesis by the length of carcinogen exposure period in rat colon. Dig. Dis. Sci. 2001, 46, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R.L.; Fleet, J.C. Animal models of colorectal cancer. Cancer Metastasis Rev. 2013, 32, 39–61. [Google Scholar] [CrossRef] [PubMed]
- Karakurt, S.; Durmus, I.M.; Erturk, S. Handbook of Animal Models and Its Uses in Cancer Research; Springer: Cham, Switzerland, 2020. [Google Scholar]
- Alshaman, R.; Alattar, A.; El-Sayed, R.M.; Gardouh, A.R.; Elshaer, R.E.; Elkazaz, A.Y.; Eladl, M.A.; El-Sherbiny, M.; Farag, N.E.; Hamdan, A.M.; et al. Formulation and Characterization of Doxycycline-Loaded Polymeric Nanoparticles for Testing Antitumor/Antiangiogenic Action in Experimental Colon Cancer in Mice. Nanomaterials 2022, 12, 857. [Google Scholar] [CrossRef]
- Shree, A.; Islam, J.; Sultana, S. Quercetin ameliorates reactive oxygen species generation, inflammation, mucus depletion, goblet disintegration, and tumor multiplicity in colon cancer: Probable role of adenomatous polyposis coli, β-catenin. Phyther Res. 2021, 35, 2171–2184. [Google Scholar] [CrossRef]
- Alkhuriji, A.F.; Alsaiari, S.G.; Alomar, S.Y.; Alnafjan, A.A.; Alobaid, H.; El-Khadragy, M.F. Effect of mesenchymal stem cells on cytochrome-c release and inflammation in colon cancer induced by 1,2-dimethylhydrazine in Wistar albino rats. Biosci. Rep. 2021, 41, BSR20204356. [Google Scholar] [CrossRef]
- Ertekin, T.; Ekinci, N.; Karaca, O.; Nisari, M.; Canoz, O.; Ulger, H. Effect of angiostatin on 1,2-dimethylhydrazine-induced colon cancer in mice. Toxicol. Ind. Health 2013, 29, 490–497. [Google Scholar] [CrossRef]
- Wang, Y.; Jin, H.-Y.; Fang, M.-Z.; Wang, X.-F.; Chen, H.; Huang, S.-L.; Kong, D.-S.; Li, M.; Zhang, X.; Sun, Y.; et al. Epigallocatechin gallate inhibits dimethylhydrazine-induced colorectal cancer in rats. World J. Gastroenterol. 2020, 26, 2064–2081. [Google Scholar] [CrossRef]
- Eissa, M.M.; Ismail, C.A.; El-Azzouni, M.Z.; Ghazy, A.A.; Hadi, M.A. Immuno-therapeutic potential of Schistosoma mansoni and Trichinella spiralis antigens in a murine model of colon cancer. Investig. New Drugs 2019, 37, 47–56. [Google Scholar] [CrossRef]
- El Joumaa, M.M.; Taleb, R.I.; Rizk, S.; Borjac, J.M. Protective effect of matricaria chamomilla extract against 1,2-dimethylhydrazine-induced colorectal cancer in mice. J. Complement. Integr. Med. 2020, 17, 20190143. [Google Scholar] [CrossRef] [PubMed]
- Schiller, C.M.; Curley, W.H.; McConnell, E.E. Induction of colon tumors by a single oral dose of 1,2-dimethylhydrazine. Cancer Lett. 1980, 11, 75–79. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, M.T.; Schop, R.N.; Reidy, J.A. Assessment of 1,2-dimethylhydrazine in bone marrow micronucleus assay: Variations in protocol and response. Environ. Mol. Mutagen. 1991, 17, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; Jin, Z.; Wu, W.; Gao, R.; Guo, B.; Gao, Z.; Yang, Y.; Qin, H. Analysis of the intestinal lumen microbiota in an animal model of colorectal cancer. PLoS ONE 2014, 9, e90849. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Qiao, X.; Wang, J.; Yang, J.; Yang, C.; Qiao, Y.; Guan, Y.; Wen, A.; Jiang, L. Amelioration of DMH-induced colon cancer by eupafolin through the reprogramming of apoptosis-associated p53/Bcl2/Bax signaling in rats. Eur. J. Inflamm. 2022, 20, 20587392211069771. [Google Scholar] [CrossRef]
- Zhao, C.; Ghosh, B.; Chakraborty, T.; Roy, S. Bavachinin mitigates DMH induced colon cancer in rats by altering p53/Bcl2/BAX signaling associated with apoptosis. Biotech. Histochem. 2020, 96, 179–190. [Google Scholar] [CrossRef]
- Babu, S.S.N.; Singla, S.; Jena, G. Role of Combination Treatment of Aspirin and Zinc in DMH-DSS-induced Colon Inflammation, Oxidative Stress and Tumour Progression in Male BALB/c Mice. Biol. Trace Elem. Res. 2022, 201, 1327–1343. [Google Scholar] [CrossRef]
- Lin, P.Y.; Li, S.C.; Lin, H.P.; Shih, C.K. Germinated brown rice combined with Lactobacillus acidophilus and Bifidobacterium animalis subsp. lactis inhibits colorectal carcinogenesis in rats. Food Sci. Nutr. 2019, 7, 216–224. [Google Scholar] [CrossRef]
- Mundo, A.I.; Muhammad, A.; Balza, K.; Nelson, C.E.; Muldoon, T.J. Longitudinal examination of perfusion and angiogenesis markers in primary colorectal tumors shows distinct signatures for metronomic and maximum-tolerated dose strategies R. Neoplasia 2022, 32, 100825. [Google Scholar] [CrossRef]
- Tian, Y.; Zuo, L.; Guan, B.; Wu, H.; He, Y.; Xu, Z.; Shen, M.; Hu, J.; Qian, J. Microbiota from patients with ulcerative colitis promote colorectal carcinogenesis in mice. Nutrition 2022, 102, 111712. [Google Scholar] [CrossRef]
- Arango-Varela, S.S.; Luzardo-Ocampo, I.; Maldonado-Celis, M.E. Andean berry (Vaccinium meridionale Swartz) juice, in combination with Aspirin, displayed antiproliferative and pro-apoptotic mechanisms in vitro while exhibiting protective effects against AOM-induced colorectal cancer in vivo. Food Res. Int. 2022, 157, 111244. [Google Scholar] [CrossRef] [PubMed]
- Dikeocha, I.J.; Al-Kabsi, A.M.; Chiu, H.T.; Alshawsh, M.A. Faecalibacterium prausnitzii Ameliorates Colorectal Tumorigenesis and Suppresses Proliferation of HCT116 Colorectal Cancer Cells. Biomedicines 2022, 10, 1128. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, F.; Zheng, H.; Kalasabail, S.; Hicks, C.; Fung, K.Y.; Preaudet, A.; Putoczki, T.; Beretov, J.; Millar EK, A.; et al. Fecal DNA Virome Is Associated with the Development of Colorectal Neoplasia in a Murine Model of Colorectal Cancer. Pathogens 2022, 11, 457. [Google Scholar] [CrossRef] [PubMed]
- Almaimani, R.A.; Aslam, A.; Ahmad, J.; El-Readi, M.Z.; El-Boshy, M.E.; Abdelghany, A.H.; Idris, S.; Alhadrami, M.; Althubiti, M.; Almasmoum, H.A.; et al. In Vivo and In Vitro Enhanced Tumoricidal Effects of of Metformin, Active Vitamin D3, and 5-Fluorouracil Triple Therapy against Colon Cancer by Modulating the PI3K/Akt/PTEN/mTOR Network. Cancers 2022, 14, 1538. [Google Scholar] [CrossRef]
- Bähr, I.; Jaeschke, L.; Nimptsch, K.; Janke, J.; Herrmann, P.; Kobelt, D.; Kielstein, H.; Pischon, T.; Stein, U. Obesity, colorectal cancer and MACC1 expression: A possible novel molecular association. Int. J. Oncol. 2022, 60, 17. [Google Scholar] [CrossRef]
- Iwama, N.; Matsuda, M.; Tsuruta, M.; Okabayashi, K.; Shigeta, K.; Kanai, T.; Kitagawa, Y. Relationship between obesity-related colorectal tumors and the intestinal microbiome: An animal-based trial. J. Cancer Res. Clin. Oncol. 2022. [Google Scholar] [CrossRef]
- Ma, F.; Song, Y.; Sun, M.; Wang, A.; Jiang, S.; Mu, G.; Tuo, Y. Exopolysaccharide produced by lactiplantibacillus plantarum-12 alleviates intestinal inflammation and colon cancer symptoms by modulating the gut microbiome and metabolites of C57BL/6 mice treated by azoxymethane/dextran sulfate sodium salt. Foods 2021, 10, 3060. [Google Scholar] [CrossRef]
- Deng, J.; Zhao, L.; Yuan, X.; Li, Y.; Shi, J.; Zhang, H.; Zhao, Y.; Han, L.; Wang, H.; Yan, Y.; et al. Pre-Administration of Berberine Exerts Chemopreventive Effects in AOM/DSS-Induced Colitis-Associated Carcinogenesis Mice via Modulating Inflammation and Intestinal Microbiota. Nutrients 2022, 14, 726. [Google Scholar] [CrossRef]
- Schepelmann, M.; Kupper, N.; Gushchina, V.; Mesteri, I.; Manhardt, T.; Moritsch, S.; Müller, C.; Piatek, K.; Salzmann, M.; Vlasaty, A.; et al. AOM/DSS Induced Colitis-Associated Colorectal Cancer in 14-Month-Old Female Balb/C and C57/Bl6 Mice—A Pilot Study. Int. J. Mol. Sci. 2022, 23, 5278. [Google Scholar] [CrossRef]
- Yang, M.; Zhang, F.; Yang, C.; Wang, L.; Sung, J.; Garg, P.; Zhang, M.; Merlin, D. Oral targeted delivery by nanoparticles enhances efficacy of an Hsp90 inhibitor by reducing systemic exposure in murine models of colitis and colitis-associated cancer. J. Crohns Colitis 2020, 14, 130–141. [Google Scholar] [CrossRef]
- Ferreira-Lazarte, A.; Fernández, J.; Gallego-Lobillo, P.; Villar, C.J.; Lombó, F.; Moreno, F.J.; Villamiel, M. Behaviour of citrus pectin and modified citrus pectin in an azoxymethane/dextran sodium sulfate (AOM/DSS)-induced rat colorectal carcinogenesis model. Int. J. Biol. Macromol. 2021, 167, 1349–1360. [Google Scholar] [CrossRef] [PubMed]
- Tajasuwan, L.; Kettawan, A.; Rungruang, T.; Wunjuntuk, K.; Prombutara, P.; Muangnoi, C.; Kettawan, A. Inhibitory Effect of Dietary Defatted Rice Bran in an AOM/DSS-Induced Colitis-Associated Colorectal Cancer Experimental Animal Model. Foods 2022, 11, 3488. [Google Scholar] [CrossRef] [PubMed]
- Bala, P.; Rennhack, J.P.; Aitymbayev, D.; Morris, C.; Moyer, S.M.; Duronio, G.N.; Doan, P.; LI, Z.; Liang, X.; Hornick, J.L.; et al. Aberrant cell state plasticity mediated by developmental reprogramming precedes colorectal cancer initiation. Sci. Adv. 2023, 9, eadf0927. Available online: https://www.science.org/doi/10.1126/sciadv.adf0927 (accessed on 24 April 2023). [CrossRef]
- Yang, X.; Peng, H.; Luo, Z.; Luo, A.; Cai, M.; Xu, L.; Wang, H. The dietary carcinogen PhIP activates p53-dependent DNA damage response in the colon of CYP1A-humanized mice. BioFactors 2021, 47, 612–626. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.X.; Liu, A.; Lee, M.J.; Wang, H.; Yu, S.; Chi, E.; Reuhl, K.; Suh, N.; Yang, C. δ- and γ-tocopherols inhibit phIP/DSS-induced colon carcinogenesis by protection against early cellular and DNA damages. Mol. Carcinog. 2017, 56, 172–183. [Google Scholar] [CrossRef] [PubMed]
- Frajacomo, F.T.; Kannen, V.; Deminice, R.; Geraldino, T.H.; Pereira-Da-Silva, G.; Uyemura, S.A.; Jordão-Jr, A.A.; Garcia, S.B. Aerobic Training Activates Interleukin 10 for Colon Anticarcinogenic Effects. Med. Sci. Sport Exerc. 2015, 47, 1806–1813. [Google Scholar] [CrossRef] [PubMed]
- Kannen, V.; Hintzsche, H.; Zanette, D.L.; Silva, W.A.; Garcia, S.B.; Waaga-Gasser, A.M.; Stopper, H. Antiproliferative Effects of Fluoxetine on Colon Cancer Cells and in a Colonic Carcinogen Mouse Model. PLoS ONE 2012, 7, e50043. [Google Scholar]
- Yusuf, A.; Odeh, O.E.; Alhassan, S.O.; Atawodi, S.E. Evaluation of the preventive potential of graded dietary inclusion of Hyphaene thebaica (Linn) fruit in rat model of colon carcinogenesis. J. Food Biochem. 2022, 46, e14446. [Google Scholar] [CrossRef]
- Qayum, A.; Singh, J.; Kumar, A.; Shah, S.M.; Srivastava, S.; Kushwaha, M.; Magotra, A.; Nandi, U.; Malik, R.; Sha, B.A.; et al. 2-Pyridin-4-yl-methylene-beta-boswellic Acid-A Potential Candidate for Targeting O6-Methylguanine-DNA Methyltransferase Epi-transcriptional Reprogramming in KRAS G13D-Microsatellite Stable, G12V-Microsatellite Instable Mutant Colon Cancer. ACS Pharmacol. Transl. Sci. 2022, 5, 306–320. [Google Scholar] [CrossRef]
- Huang, Z.; Liu, C.A.; Cai, P.Z.; Xu, F.P.; Zhu, W.J.; Wang, W.W.; Jiang, H.P. Omega-3PUFA attenuates mnu-induced colorectal cancer in rats by blocking PI3K/AKT/ BCL-2 signaling. Oncol. Targets Ther. 2020, 13, 1953–1965. [Google Scholar] [CrossRef]
- Attia, N.A.; Sayed, A.H.; Mahmoud, N.S.; Ahmed, H.H. Phytochemical remedies: A key strategy towards reversing the aggressive murine colon cancer. Med. Chem. Res. 2017, 26, 2614–2623. [Google Scholar] [CrossRef]
- Ahmed, H.H.; El-Abhar, H.S.; Hassanin, E.A.K.; Abdelkader, N.F.; Shalaby, M.B. Punica granatum suppresses colon cancer through downregulation of Wnt/β-catenin in rat model. Rev. Bras. Farmacogn. 2017, 27, 627–635. [Google Scholar] [CrossRef]
- Ahmed, H.H.; El-Abhar, H.S.; Hassanin, E.A.K.; Abdelkader, N.F.; Shalaby, M.B.; Ginkgo Biloba, L. Leaf extract offers multiple mechanisms in bridling N-methylnitrosourea—Mediated experimental colorectal cancer. Biomed. Pharmacother. 2017, 95, 387–393. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, Y.; Inoue, Y.; Minagawa, N.; Onitsuka, K.; Nagata, J.; Shibao, K.; Hirata, K.; Sako, T.; Nagata, N.; Yamaguchi, K. Chemopreventive effect of 4-[3,5-bis(trimethylsilyl) benzamido] benzoic acid (TAC-101) on MNU-induced colon carcinogenesis in a rat model. Anticancer Res. 2009, 29, 2059–2065. [Google Scholar] [PubMed]
- De-Souza, A.S.C.; Costa-Casagrande, T.A. Animal model for colorectal cancer. ABCD Arq. Bras. Cir. Dig. 2018, 31, e1369. [Google Scholar] [CrossRef] [PubMed]
- Cefalì, M.; Epistolio, S.; Palmarocchi, M.C.; Frattini, M.; De Dosso, S. Research progress on KRAS mutations in colorectal cancer. J. Cancer Metastasis Treat. 2021, 7, 26. [Google Scholar] [CrossRef]
- Peehl, D.M.; Badea, C.T.; Chenevert, T.L.; Daldrup-Link, H.E.; Ding, L.; Dobrolecki, L.E.; Houghton, A.M.; Kinahan, P.E.; Kurhanewicz, J.; Lewis, M.T.; et al. Animal Models and Their Role in Imaging-Assisted Co-Clinical Trials. Tomography 2023, 9, 657–680. [Google Scholar] [CrossRef]
- Chandra, R.; Karalis, J.D.; Liu, C.; Murimwa, G.Z.; Park, J.V.; Heid, C.A.; Resnik, S.I.; Huang, E.; Minna, J.D.; Brekken, R.A. The Colorectal Cancer Tumor Microenvironment and Its Impact on Liver and Lung Metastasis. Cancers 2021, 13, 6206. [Google Scholar] [CrossRef]
- Hassin, O.; Nataraj, N.B.; Shreberk-Shaked, M.; Aylon, Y.; Yaeger, R.; Fontemaggi, G.; Mukherjee, S.; Maddalena, M.; Avioz, A.; Iancu, O.; et al. Different hotspot p53 mutants exert distinct phenotypes and predict outcome of colorectal cancer patients. Nat. Commun. 2022, 13, 2800. [Google Scholar] [CrossRef]
- Hankey, W.; Frankel, W.L.; Groden, J. Functions of the APC tumor suppressor protein dependent and independent of canonical WNT signaling: Implications for therapeutic targeting. Physiol. Behav. 2018, 37, 159–172. [Google Scholar] [CrossRef]
- Ren, J.; Sui, H.; Fang, F.; Li, Q.; Li, B. The application of Apc Min/+ mouse model in colorectal tumor researches. J. Cancer Res. Clin. Oncol. 2019, 145, 1111–1122. [Google Scholar] [CrossRef] [PubMed]
- Mcintyre, R.E.; Buczacki, S.J.A.; Arends, M.J.; Adams, D.J. Mouse models of colorectal cancer as preclinical models. BioEssays 2015, 37, 909–920. [Google Scholar] [CrossRef] [PubMed]
- Bürtin, F.; Mullins, C.S.; Linnebacher, M. Mouse models of colorectal cancer: Past, present and future perspectives. World J. Gastroenterol. 2020, 26, 1394–1426. [Google Scholar] [CrossRef] [PubMed]
- Karim, B.O.; Huso, D.L. Mouse models for colorectal cancer. Am. J. Cancer Res. 2013, 3, 240–250. [Google Scholar]
- Liu, X.M.; Zhu, W.T.; Jia, M.L.; Li, Y.T.; Hong, Y.; Liu, Z.Q.; Yan, P.K. Rapamycin Liposomes Combined with 5-Fluorouracil Inhibits Angiogenesis and Tumor Growth of APC(Min/+) Mice and AOM/ DSS-Induced Colorectal Cancer Mice. Int. J. Nanomed. 2022, 17, 5049–5061. [Google Scholar] [CrossRef]
- Metzger, R.; Maruskova, M.; Krebs, S.; Janssen, K.P.; Krug, A.B. Increased Incidence of Colon Tumors in AOM-Treated Apc1638N/+ Mice Reveals Higher Frequency of Tumor Associated Neutrophils in Colon Than Small Intestine. Front. Oncol. 2019, 9, 1001. [Google Scholar] [CrossRef]
- Clark, C.R.; Starr, T.K. Mouse models for the discovery of colorectal cancer driver genes. World J. Gastroenterol. 2016, 22, 815–822. [Google Scholar] [CrossRef]
- Muller, P.A.J.; Caswell, P.T.; Doyle, B.; Iwanicki, M.P.; Tan, E.H.; Karim, S.; Lukashchuk, N.; Gillespie, D.A.; Ludwig, R.L.; Gosselin, P.; et al. Mutant p53 Drives Invasion by Promoting Integrin Recycling. Cell. 2009, 139, 1327–1341. [Google Scholar] [CrossRef]
- Sei, S.; Ahadova, A.; Keskin, D.B.; Bohaumilitzky, L.; Gebert, J.; von Knebel Doeberitz, M.; Lipkin, S.M.; Kloor, M. Lynch syndrome cancer vaccines: A roadmap for the development of precision immunoprevention strategies. Front. Oncol. 2023, 13, 1147590. [Google Scholar] [CrossRef]
- Telang, N. Isolation and characterization of chemo-resistant stem cells from a mouse model of hereditary non-polyposis colon cancer. Stem Cells Cloning Adv. Appl. 2021, 14, 19–25. [Google Scholar] [CrossRef]
- Pećina-Šlaus, N.; Kafka, A.; Salamon, I.; Bukovac, A. Mismatch Repair Pathway, Genome Stability and Cancer. Front. Mol. Biosci. 2020, 7, 122. [Google Scholar] [CrossRef] [PubMed]
- Valle, L.; Vilar, E.; Tavtigian, S.V.; Stoffel, E.M. Genetic predisposition to colorectal cancer: Syndromes, genes, classification of genetic variants and implications for precision medicine. J. Pathol. 2019, 247, 574–588. Available online: https://onlinelibrary.wiley.com/doi/10.1002/path.5229 (accessed on 7 February 2023). [CrossRef] [PubMed]
- Hryhorowicz, S.; Kaczmarek-Ryś, M.; Lis-Tanaś, E.; Porowski, J.; Szuman, M.; Grot, N.; Kryszczyńska, A.; Paszkowski, J.; Banasiewicz, T.; Pławski, A. Strong Hereditary Predispositions to Colorectal Cancer. Genes 2022, 13, 2326. [Google Scholar] [CrossRef] [PubMed]
- Taketo, M.M.; Edelmann, W. Mouse Models of Colon Cancer. Gastroenterology 2009, 136, 780–798. [Google Scholar] [CrossRef]
- Heijstek, M.W.; Kranenburg, O.; Rinkes, I.H.M.B. Mouse Models of Colorectal Cancer and Liver Metastases. Dig. Surg. 2005, 22, 26–33. Available online: http://www.ncbi.nlm.nih.gov/pubmed/15838167 (accessed on 30 November 2022). [CrossRef]
- Uronis, J.M.; Threadgill, D.W. Murine models of colorectal cancer. Mamm. Genome 2009, 20, 261–268. [Google Scholar] [CrossRef]
- Schnell, U.; Cirulli, V.; Giepmans, B.N.G. Biochimica et Biophysica Acta EpCAM: Structure and function in health and disease. BBA Biomembr. 2013, 1828, 1989–2001. [Google Scholar] [CrossRef]
- Gires, O. Expression and function of epithelial cell adhesion molecule EpCAM: Where are we after 40 years? Cancer Metastasis Rev. 2020, 39, 969–987. [Google Scholar] [CrossRef]
- Hosono, H.; Ohishi, T.; Takei, J.; Asano, T.; Sayama, Y.; Kawada, M.; Kaneko, M.; Kato, Y. The anti-epithelial cell adhesion molecule (EpCAM) monoclonal antibody EpMab-16 exerts antitumor activity in a mouse model of colorectal adenocarcinoma. Oncol. Lett. 2020, 20, 383. [Google Scholar] [CrossRef]
- Alamo, P.; Gallardo, A.; Di Nicolantonio, F.; Pavón, M.A.; Casanova, I.; Trias, M.; Mangues, M.A.; Lopez-Pousa, A.; Villaverde, A.; Vázquez, E.; et al. Higher metastatic efficiency of KRas G12V than KRas G13D in a colorectal cancer model. FASEB J. 2015, 29, 464–476. [Google Scholar] [CrossRef]
- Biswas, K.; Couillard, M.; Cavallone, L.; Burkett, S.; Stauffer, S.; Martin, B.K.; Southon, E.; Reid, S.; Plona, T.M.; Baugher, R.N.; et al. A novel mouse model of PMS2 founder mutation that causes mismatch repair defect due to aberrant splicing. Cell Death Dis. 2021, 12, 838. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, R.C.; Abrantes, A.M.; Tralhão, J.G.; Botelho, M.F. The role of mouse models in colorectal cancer research—The need and the importance of the orthotopic models. Anim. Model. Exp. Med. 2020, 3, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Shibata, H.; Toyama, K.; Shioya, H.; Ito, M.; Hirota, M.; Hasegawa, S.; Matsumoto, H.; Takano, H.; Akiyama, T.; Toyoshima, K.; et al. Rapid Colorectal Adenoma Formation Initiated by Conditional Targeting of the Apc Gene. Science 1997, 278, 120–123. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.; Johnson, R.; DeSmet, M.; Snyder, P.W.; Fleet, J.C. Generation of a Transgenic Mouse for Colorectal Cancer Research with Intestinal Cre-Expression Limited to the Large Intestine. Mol. Cancer Res. 2010, 8, 1095–1104. [Google Scholar] [CrossRef] [PubMed]
- Halberg, R.B.; Katzung, D.S.; Hoff, P.D.; Moser, A.R.; Cole, C.E.; Lubet, R.A.; Donehower, L.A.; Jacoby, R.F.; Dove, W.F. Tumorigenesis in the multiple intestinal neoplasia mouse: Redundancy of negative regulators and specificity of modifiers. Proc. Natl. Acad. Sci. USA 2000, 97, 3461–3466. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, K.; Lin, B.; Nunomura, K.; Izawa, T.; Nakagawa, S. The K-Ras(G12D)-inhibitory peptide KS-58 suppresses growth of murine CT26 colorectal cancer cell-derived tumors. Sci. Rep. 2022, 12, 8121. [Google Scholar] [CrossRef]
- Yueh, A.E.; Payne, S.N.; Leystra, A.A.; Van De Hey, D.R.; Foley, T.M.; Pasch, C.A.; Clipson, L.; Matkowskyj, K.A.; Deming, D.A. Colon cancer tumorigenesis initiated by the H1047R mutant PI3K. PLoS ONE 2016, 11, e0148730. [Google Scholar] [CrossRef]
- Herberg, M.; Siebert, S.; Quaas, M.; Thalheim, T.; Rother, K.; Hussong, M.; Altmüller, J.; Kerner, C.; Galle, J.; Aust, G.; et al. Loss of Msh2 and a single-radiation hit induce common, genome-wide, and persistent epigenetic changes in the intestine. Clin. Epigenet. 2019, 11, 65. [Google Scholar] [CrossRef]
- Choi, S.H.; Huang, A.Y.; Letterio, J.J.; Kim, B.G. Smad4-deficient T cells promote colitis-associated colon cancer via an IFN-γ-dependent suppression of 15-hydroxyprostaglandin dehydrogenase. Front. Immunol. 2022, 13, 932412. [Google Scholar] [CrossRef]
- Hung, K.E.; Maricevich, M.A.; Richard, L.G.; Chen, W.Y.; Richardson, M.P.; Kunin, A.; Bronson, R.T.; Mahmood, U.; Kucherlapati, R. Development of a mouse model for sporadic and metastatic colon tumors and its use in assessing drug treatment. Proc. Natl. Acad. Sci. USA. 2010, 107, 1565–1570. [Google Scholar] [CrossRef]
- Bennecke, M.; Kriegl, L.; Bajbouj, M.; Retzlaff, K.; Robine, S.; Jung, A.; Arkan, M.C.; Kirchner, T.; Greten, F.R. Ink4a/Arf and oncogene-induced senescence prevent tumor progression during alternative colorectal tumorigenesis. Cancer Cell 2010, 18, 135–146. [Google Scholar] [CrossRef] [PubMed]
- Lo, Y.H.; Noah, T.K.; Chen, M.S.; Zou, W.; Borras, E.; Vilar, E.; Shroyer, N.F. SPDEF Induces Quiescence of Colorectal Cancer Cells by Changing the Transcriptional Targets of β-catenin. Gastroenterology 2017, 153, 205–218.e8. [Google Scholar] [CrossRef] [PubMed]
- Steffensen, I.L.; Alexander, J. Impact of genetic background on spontaneous or 2-amino-1-methyl-6-phenylimidazo[4,5-b]pyridine (PhIP)-induced intestinal tumorigenesis in Min/+ mice. Cancer Lett. 2006, 240, 289–296. [Google Scholar] [CrossRef] [PubMed]
- Oshima, H.; Nakayama, M.; Han, T.S.; Naoi, K.; Ju, X.; Maeda, Y.; Robine, S.; Tsuchiya, K.; Sato, T.; Sato, H.; et al. Suppressing TGFβ signaling in regenerating epithelia in an inflammatory microenvironment is sufficient to cause invasive intestinal cancer. Cancer Res. 2015, 75, 766–776. [Google Scholar] [CrossRef] [PubMed]
- Sakai, E.; Nakayama, M.; Oshima, H.; Kouyama, Y.; Niida, A.; Fujii, S.; Ochiai, A.; Nakayama, K.I.; Mimori, K.; Suzuki, Y.; et al. Combined mutation of Apc, Kras, and Tgfbr2 effectively drives metastasis of intestinal cancer. Cancer Res. 2018, 78, 1334–1346. [Google Scholar] [CrossRef]
- Takaku, K.; Oshima, M.; Miyoshi, H.; Matsui, M.; Seldin, M.F.; Taketo, M.M. Intestinal tumorigenesis in compound mutant mice of both Dpc4 (Smad4) and Apc genes. Cell 1998, 92, 645–656. [Google Scholar] [CrossRef]
- Kucherlapati, M.; Yang, K.; Kuraguchi, M.; Zhao, J.; Lia, M.; Heyer, J.; Kane, M.F.; Fan, K.; Russell, R.; Brown, A.M.C.; et al. Haploinsufficiency of Flap endonuclease (Fen1) leads to rapid tumor progression. Proc. Natl. Acad. Sci. USA 2002, 99, 9924–9929. [Google Scholar] [CrossRef]
- Grim, J.E.; Knoblaugh, S.E.; Guthrie, K.A.; Hagar, A.; Swanger, J.; Hespelt, J.; Delrow, J.J.; Small, T.; Grady, W.M.; Nakayama, K.I.; et al. Fbw7 and p53 Cooperatively Suppress Advanced and Chromosomally Unstable Intestinal Cancer. Mol. Cell Biol. 2012, 32, 2160–2167. [Google Scholar] [CrossRef]
- Sansom, O.J.; Meniel, V.; Wilkins, J.A.; Cole, A.M.; Oien, K.A.; Marsh, V.; Jamieson, T.J.; Guerra, C.; Ashton, G.H.; Barbacid, M.; et al. Loss of Apc allows phenotypic manifestation of the transforming properties of an endogenous K-ras oncogene in vivo. Proc. Natl. Acad. Sci. USA 2006, 103, 14122–14127. [Google Scholar] [CrossRef]
- Tong, K.; Kothari, O.A.; Haro, K.S.; Panda, A.; Bandari, M.M.; Carrick, J.N.; Hur, J.J.; Zhang, L.; Chan, C.S.; Xing, J.; et al. SMAD4 is critical in suppression of BRAF-V600E serrated tumorigenesis. Oncogene 2021, 40, 6034–6048. [Google Scholar] [CrossRef]
- Zhou, H.; Liu, Z.; Wang, Y.; Wen, X.; Amador, E.H.; Yuan, L.; Ran, X.; Xiong, L.; Ran, Y.; Chen, W.; et al. Colorectal liver metastasis: Molecular mechanism and interventional therapy. Signal Transduct. Target. Ther. Vol. 2022, 7, 70. [Google Scholar] [CrossRef] [PubMed]
- Abdolahi, S.; Ghazvinian, Z.; Muhammadnejad, S.; Saleh, M.; Asadzadeh Aghdaei, H.; Baghaei, K. Patient-derived xenograft (PDX) models, applications and challenges in cancer research. J. Transl. Med. 2022, 20, 206. [Google Scholar] [CrossRef] [PubMed]
- Talmadge, J.E.; Singh, R.K.; Fidler, I.J.; Raz, A. Murine models to evaluate novel and conventional therapeutic strategies for cancer. Am. J. Pathol. 2007, 170, 793–804. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Xin, Z.; Wang, K. Patient-derived xenograft model in colorectal cancer basic and translational research. Anim. Model. Exp. Med. 2022, 6, 26–40. [Google Scholar] [CrossRef]
- Chen, J.; Liao, S.; Xiao, Z.; Pan, Q.; Wang, X.; Shen, K.; Wang, S.; Yang, L.; Guo, F.; Liu, H.; et al. The development and improvement of immunodeficient mice and humanized immune system mouse models. Front. Immunol. 2022, 13, 1007579. [Google Scholar] [CrossRef]
- Golovko, D.; Kedrin, D.; Yilmaz, O.H.; Roper, J. Review: US Spelling Colorectal cancer models for novel drug discovery. Expert Opin. Drug. Discov. 2015, 10, 1217–1229. [Google Scholar] [CrossRef]
- Medico, E.; Russo, M.; Picco, G.; Cancelliere, C.; Valtorta, E.; Corti, G.; Buscarino, M.; Isella, C.; Lamba, S.; Martinoglio, B.; et al. The molecular landscape of colorectal cancer cell lines unveils clinically actionable kinase targets. Nat. Commun. 2015, 6, 7002. [Google Scholar] [CrossRef]
- Ben-David, U.; Siranosian, B.; Ha, G.; Tang, H.; Oren, Y.; Hinohara, K.; Strathdee, C.A.; Dempster, J.; Lyons, N.J.; Burns, B.R.; et al. Genetic and transcriptional evolution alters cancer cell line drug response. Nature 2018, 560, 325–330. [Google Scholar] [CrossRef]
- Greenlee, J.D.; King, M.R. A syngeneic MC38 orthotopic mouse model of colorectal cancer metastasis. Biol. Methods Protoc. 2022, 7. Available online: https://watermark.silverchair.com/bpac024.pdf?token=AQECAHi208BE49Ooan9kkhW_Ercy7Dm3ZL_9Cf3qfKAc485ysgAAAvAwggLsBgkqhkiG9w0BBwagggLdMIIC2QIBADCCAtIGCSqGSIb3DQEHATAeBglghkgBZQMEAS4wEQQMW7ifi2ah6wSp2kfKAgEQgIICowvrwbE8DwMIaRs0BJ56xOKaLEpvyVYhYFKunlHZT9dhBMX (accessed on 13 March 2023). [CrossRef]
- Kishimoto, H.; Momiyama, M.; Aki, R.; Kimura, H.; Suetsugu, A.; Bouvet, M.; Fujiwara, T.; Hoffman, R.M. Development of a clinically-precise mouse model of rectal cancer. PLoS ONE 2013, 8, e79453. [Google Scholar] [CrossRef]
- Gulbake, A.; Jain, A.; Jain, A.; Jain, A.; Jain, S.K. Insight to drug delivery aspects for colorectal cancer. World J. Gastroenterol. 2016, 22, 582–599. [Google Scholar] [CrossRef] [PubMed]
- Dinger, T.F.; Chen, O.; Dittfeld, C.; Hetze, L.; Hüther, M.; Wondrak, M.; Löck, S.; Eicheler, W.; Breier, G.; Kunz-Schughart, L.A. Microenvironmentally-driven plasticity of CD44 isoform expression determines engraftment and stem-like phenotype in CRC cell lines. Theranostics 2020, 10, 7599–7621. [Google Scholar] [CrossRef] [PubMed]
- Almajali, B.; Al-Jamal, H.A.N.; Taib, W.R.W.; Ismail, I.; Johan, M.F.; Doolaanea, A.A.; Ibrahim, W.N. Thymoquinone, as a novel therapeutic candidate of cancers. Pharmaceuticals 2021, 14, 369. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.R.; Kozalak, G.; Di Bari, I.; Babar, Q.; Niknam, Z.; Rasmi, Y.; Yong, K.W. In Vitro Human Cancer Models for Biomedical Applications. Cancers 2022, 14, 2284. [Google Scholar] [CrossRef]
- Rajput, A.; Agarwal, E.; Leiphrakpam, P.; Brattain, M.G.; Chowdhury, S. Establishment and Validation of an Orthotopic Metastatic Mouse Model of Colorectal Cancer. ISRN Hepatol. 2013, 2013, 206875. [Google Scholar] [CrossRef]
- Idrisova, K.F.; Simon, H.U.; Gomzikova, M.O. Role of Patient-Derived Models of Cancer in Translational Oncology. Cancers 2023, 15, 139. [Google Scholar] [CrossRef]
- Hon, K.W.; Zainal Abidin, S.A.; Othman, I.; Naidu, R. The Crosstalk Between Signaling Pathways and Cancer Metabolism in Colorectal Cancer. Front. Pharmacol. 2021, 12, 768861. [Google Scholar] [CrossRef]
- Zhang, B.; Liu, Q.; Wen, W.; Gao, H.; Wei, W.; Tang, A.; Qin, B.; Lyu, H.; Meng, X.; Li, K.; et al. The chromatin remodeler CHD6 promotes colorectal cancer development by regulating TMEM65-mediated mitochondrial dynamics via EGF and Wnt signaling. Cell Discov. 2022, 8, 130. [Google Scholar] [CrossRef]
- Khan, F.A.; Albalawi, R.; Pottoo, F.H. Trends in targeted delivery of nanomaterials in colon cancer diagnosis and treatment. Med. Res. Rev. 2022, 42, 227–258. [Google Scholar] [CrossRef]
- Páez-Franco, J.C.; Zermeño-Ortega, M.R.; de la O-Contreras, C.M.; Canseco-González, D.; Parra-Unda, J.R.; Avila-Sorrosa, A.; Enríquez, R.G.; Germán-Acacio, J.M.; Morales-Morales, D. Relevance of Fluorinated Ligands to the Design of Metallodrugs for Their Potential Use in Cancer Treatment. Pharmaceutics 2022, 14, 402. [Google Scholar] [CrossRef]
- Dsouza, V.L.; Kuthethur, R.; Kabekkodu, S.P.; Chakrabarty, S. Organ-on-Chip platforms to study tumor evolution and chemosensitivity. Biochim. Biophys. Acta Rev. Cancer 2022, 1877, 188717. [Google Scholar] [CrossRef] [PubMed]
- Klose, J.; Trefz, S.; Wagner, T.; Steffen, L.; Charrier, A.P.; Radhakrishnan, P.; Volz, C.; Schmidt, T.; Ulrich, A.; Dieter, S.M.; et al. Salinomycin: Anti-tumor activity in a preclinical colorectal cancer model. PLoS ONE 2019, 14, e0211916. [Google Scholar] [CrossRef]
- Xu, C.; Li, X.; Liu, P.; Li, M.; Luo, F. Patient-derived xenograft mouse models: A high fidelity tool for individualized medicine (review). Oncol. Lett. 2019, 17, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Calcagno, S.R.; Li, S.; Colon, M.; Kreinest, P.A.; Thompson, E.A.; Fields, A.P.; Murray, N.R. Oncogenic K-ras promotes early carcinogenesis in the mouse proximal colon. Int. J. Cancer 2008, 122, 2462–2470. [Google Scholar] [CrossRef] [PubMed]
- Cook, D.R.; Kang, M.; Martin, T.D.; Galanko, J.A.; Loeza, G.H.; Trembath, D.G.; Justilien, V.; Pickering, K.A.; Vincent, D.F.; Jarosch, A.; et al. Aberrant Expression and Subcellular Localization of ECT2 Drives Colorectal Cancer Progression and Growth. Cancer Res. 2021, 82, 90–104. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Huang, L.; Qi, S.; Liu, S.; Xie, W.; Du, L.; Cui, J.; Zhang, X.; Zhang, B.; Liu, L.; et al. Ferroptosis: The Entanglement between Traditional Drugs and Nanodrugs in Tumor Therapy. Adv. Healthc. Mater. 2023. [Google Scholar] [CrossRef]
- Wani, W.A.; Prashar, S.; Shreaz, S.; Gomez-Ruiz, S. Nanostructured materials functionalized with metal complexes: In search of alternatives for administering anticancer metallodrugs. Coord. Chem. Rev. 2016, 312, 67–98. [Google Scholar] [CrossRef]
- Wu, S.; Wang, J.; Fu, Z.; Familiari, G.; Relucenti, M.; Aschner, M.; Li, X.; Chen, H.; Chen, R. Matairesinol Nanoparticles Restore Chemosensitivity and Suppress Colorectal Cancer Progression in Preclinical Models: Role of Lipid Metabolism Reprogramming. Nano Lett. 2023, 23, 1970–1980. Available online: https://pubs.acs.org/doi/10.1021/acs.nanolett.3c00035 (accessed on 30 November 2022). [CrossRef]
- Sun, X.; Ng, T.T.H.; Sham, K.W.Y.; Zhang, L.; Chan, M.T.V.; Wu, W.K.K.; Cheng, C.H.K. Bufalin, a traditional Chinese medicine compound, prevents tumor formation in two murine models of colorectal cancer. Cancer Prev. Res. 2019, 12, 653–666. [Google Scholar] [CrossRef]
- Xu, Y.; Zhang, L.; Wang, Q.; Zheng, M. Comparison of Different Colorectal Cancer With Liver Metastases Models Using Six Colorectal Cancer Cell Lines. Pathol. Oncol. Res. 2020, 26, 2177–2183. [Google Scholar] [CrossRef]
- Kim, H.D.; Park, E.J.; Choi, E.K.; Song, S.Y.; Hoe, K.L.; Kim, D.U. G-749 Promotes Receptor Tyrosine Kinase TYRO3 Degradation and Induces Apoptosis in Both Colon Cancer Cell Lines and Xenograft Mouse Models. Front. Pharmacol. 2021, 12, 730241. [Google Scholar] [CrossRef]
- Salama, A.A.A.; Allam, R.M. Promising targets of chrysin and daidzein in colorectal cancer: Amphiregulin, CXCL1, and MMP-9. Eur. J. Pharmacol. 2021, 892, 173763. [Google Scholar] [CrossRef]
- Teng, S.; Li, Y.E.; Yang, M.; Qi, R.; Huang, Y.; Wang, Q.; Zhang, Y.; Chen, S.; Li, S.; Lin, K.; et al. Tissue-specific transcription reprogramming promotes liver metastasis of colorectal cancer. Cell Res. 2020, 30, 34–49. [Google Scholar] [CrossRef] [PubMed]
- Shimura, T.; Toden, S.; Komarova, N.L.; Boland, C.R.; Wodarz, D.; Goel, A. A comprehensive in vivo and mathematic modelingbased kinetic characterization for aspirin-induced chemoprevention in colorectal cancer. Carcinogenesis 2020, 41, 751–760. [Google Scholar] [CrossRef]
- Li, C.; Wang, Y.; Liu, D.; Wong, C.C.; Coker, O.O.; Zhang, X.; Liu, C.; Zhou, Y.; Liu, Y.; Kang, W.; et al. Squalene epoxidase drives cancer cell proliferation and promotes gut dysbiosis to accelerate colorectal carcinogenesis. Gut 2022, 71, 2253–2265. [Google Scholar] [CrossRef] [PubMed]
- Hare, J.I.; Neijzen, R.W.; Anantha, M.; Dos Santos, N.; Harasym, N.; Webb, M.S.; Allen, T.M.; Bally, M.B.; Waterhouse, D.N. Treatment of Colorectal Cancer Using a Combination of Liposomal Irinotecan (Irinophore CTM) and 5-Fluorouracil. PLoS ONE 2013, 8, e62349. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, T.; Morotomi, M.; Nomoto, K. A novel mouse model of rectal cancer established by orthotopic implantation of colon cancer cells. Cancer Sci. 2004, 95, 514–519. [Google Scholar] [CrossRef]
- Hollandsworth, H.M.; Amirfakhri, S.; Filemoni, F.; Hoffman, R.M.; Molnar, J.; Yazaki, P.J.; Bouvet, M. Humanized Anti–Tumor-Associated Glycoprotein–72 for Submillimeter Near-Infrared Detection of Colon Cancer in Metastatic Mouse Models. J. Surg. Res. 2020, 252, 16–21. [Google Scholar] [CrossRef]
- Hollandsworth, H.M.; Amirfakhri, S.; Filemoni, F.; Molnar, J.; Hoffman, R.M.; Yazaki, P.; Bouvet, M. Near-infrared photoimmunotherapy is effective treatment for colorectal cancer in orthotopic nude-mouse models. PLoS ONE 2020, 15, e0234643. [Google Scholar] [CrossRef]
- Thangaiyan, R.; Aljahdali, I.A.M.; Lent-Moore, K.Y.; Liao, J.; Ling, X.; Li, F. Kras mutation subtypes distinctly affect colorectal cancer cell sensitivity to FL118, a novel inhibitor of survivin, Mcl-1, XIAP, cIAP2 and MdmX. Am. J. Transl. Res. 2021, 13, 7458–7474. [Google Scholar]
- Murdocca, M.; Capuano, R.; Pucci, S.; Cicconi, R.; Polidoro, C.; Catini, A.; Martinelli, E.; Paolesse, R.; Orlandi, A.; Mango, R.; et al. Targeting LOX-1 inhibits colorectal cancer metastasis in an animal model. Front. Oncol. 2019, 9, 927. [Google Scholar] [CrossRef]
- Ko, E.J.; Ock, M.S.; Choi, Y.H.; Iovanna, J.L.; Mun, S.; Han, K.; Kim, H.; Cha, H. Human Endogenous Retrovirus (HERV)-K env Gene Knockout Affects Tumorigenic Characteristics of nupr1 Gene in DLD-1 Colorectal Cancer Cells. Int. J. Mol. Sci. 2021, 22, 3941. [Google Scholar] [CrossRef]
- Céspedes, M.V.; Espina, C.; García-Cabezas, M.A.; Trias, M.; Boluda, A.; Gómez Del Pulgar, M.T.; Sancho, F.J.; Nistal, M.; Lacal, J.C.; Mangues, R. Orthotopic Microinjection of Human Colon Cancer Cells in Nude Mice Induces Tumor Foci in All Clinically Relevant Metastatic Sites. Am. J. Pathol. 2007, 170, 1077–1085. [Google Scholar] [CrossRef] [PubMed]
- Rivera, M.; Fichtner, I.; Wulf-Goldenberg, A.; Sers, C.; Merk, J.; Patone, G.; Alp, K.M.; Kanashova, T.; Mertins, P.; Hoffmann, J.; et al. Patient-derived xenograft (PDX) models of colorectal carcinoma (CRC) as a platform for chemosensitivity and biomarker analysis in personalized medicine. Neoplasia US 2021, 23, 21–35. [Google Scholar] [CrossRef] [PubMed]
- Relucenti, M.; Francescangeli, F.; De Angelis, M.L.; D’Andrea, V.; Miglietta, S.; Donfrancesco, O.; Li, X.; Chen, R.; Zeuner, A.; Familiari, G. A Different Exosome Secretion Pattern Characterizes Patient-Derived Colorectal Cancer Multicellular Spheroids and Their Mouse Xenografts. Biology 2022, 11, 1427. Available online: https://www.mdpi.com/2079-7737/11/10/1427 (accessed on 30 November 2022).
- Hua, L.; Chen, L.; Huang, J.; Chen, X.; Guo, S.; Wang, J. Establishment of RET inhibitor-induced resistant patient-derived colorectal cancer xenograft models. Eur. J. Cancer 2022, 174, S44. [Google Scholar] [CrossRef]
- Drury, J.; Young, L.E.A.; Scott, T.L.; Kelson, C.O.; He, D.; Liu, J.; Wu, Y.; Wang, C.; Weiss, H.L.; Fan, T.; et al. Tissue-Specific Downregulation of Fatty Acid Synthase Suppresses Intestinal Adenoma Formation via Coordinated Reprograming of Transcriptome and Metabolism in the Mouse Model of Apc-Driven Colorectal Cancer. Int. J. Mol. Sci. 2022, 23, 6510. [Google Scholar] [CrossRef]
- Liu, Y.; Wu, W.; Cai, C.; Zhang, H.; Shen, H.; Han, Y. Patient-derived xenograft models in cancer therapy: Technologies and applications. Signal. Transduct. Target. Ther. 2023, 8, 160. [Google Scholar] [CrossRef]
- Janakiraman, H.; Zhu, Y.; Becker, S.A.; Wang, C.; Cross, A.; Curl, E.; Lewin, D.; Hoffman, B.J.; Warren, G.W.; Hill, E.G.; et al. Modeling rectal cancer to advance neoadjuvant precision therapy. Int. J. Cancer 2020, 147, 1405–1418. [Google Scholar] [CrossRef]
- De Angelis, M.L.; Francescangeli, F.; Nicolazzo, C.; Xhelili, E.; La Torre, F.; Colace, L.; Bruselles, A.; Macchia, D.; Vitale, S.; Gazzaniga, P.; et al. An Orthotopic Patient-Derived Xenograft (PDX) Model Allows the Analysis of Metastasis-Associated Features in Colorectal Cancer. Front. Oncol. 2022, 12, 869485. [Google Scholar] [CrossRef]
- Cassidy, J.W.; Caldas, C.; Bruna, A. Maintaining tumor heterogeneity in patient-derived tumor xenografts. Cancer Res. 2015, 75, 2963–2968. [Google Scholar] [CrossRef]
- Cho, S.-Y. Patient-derived xenografts as compatible models for precision oncology. Lab. Anim. Res. 2020, 36, 14. [Google Scholar] [CrossRef] [PubMed]
- Ramzy, G.M.; Koessler, T.; Ducrey, E.; McKee, T.; Ris, F.; Buchs, N.; Rubbia-Brandt, L.; Dietrich, P.-Y.; Nowak-Sliwinska, P. Patient-derived in vitro models for drug discovery in colorectal carcinoma. Cancers 2020, 12, 1423. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, N.; Weinberg, E.M.; Nguyen, A.; Liberti, M.V.; Goodarzi, H.; Janjigian, Y.Y.; Paty, P.B.; Saltz, L.B.; Kingham, T.P.; Loo, J.M.; et al. PCK1 and DHODH drive colorectal cancer liver metastatic colonization and hypoxic growth by promoting nucleotide synthesis. Elife 2019, 8, e52135. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Lin, W.; Huang, Y.; Chen, X.; Wang, H.; Teng, L. The essential factors of establishing patient-derived tumor model. J. Cancer 2021, 12, 28–37. [Google Scholar] [CrossRef]
- Wang, E.; Xiang, K.; Zhang, Y.; Wang, X.F. Patient-derived organoids (PDOs) and PDO-derived xenografts (PDOXs): New opportunities in establishing faithful pre-clinical cancer models. J. Natl. Cancer Cent. 2022, 2, 263–276. [Google Scholar] [CrossRef]
- Hassani, I.; Anbiah, B.; Kuhlers, P.; Habbit, N.L.; Ahmed, B.; Heslin, M.J.; Mobley, J.A.; Greene, M.W.; Lipke, E.A. Engineered colorectal cancer tissue recapitulates key attributes of a patient-derived xenograft tumor line. Biofabrication 2022, 14, 045001. [Google Scholar] [CrossRef]
- Fondevila, F.; Méndez-Blanco, C.; Fernández-Palanca, P.; González-Gallego, J.; Mauriz, J.L. Anti-tumoral activity of single and combined regorafenib treatments in preclinical models of liver and gastrointestinal cancers. Exp. Mol. Med. 2019, 51, 1–15. [Google Scholar] [CrossRef]
- Vaghi, C.; Mauri, G.; Giuseppe Agostara, A.; Patelli, G.; Gregory Pizzutilo, E.; Nakamura, Y.; Yoshino, T.; Siena, S.; Sartore-Bianchi, A. The predictive role of ERBB2 point mutations in metastatic colorectal cancer: A systematic review. Cancer Treat. Rev. 2022, 112, 102488. [Google Scholar] [CrossRef]
- La Salvia, A.; Lopez-Gomez, V.; Garcia-Carbonero, R. HER2-targeted therapy: An emerging strategy in advanced colorectal cancer. Expert. Opin. Investig. Drugs 2019, 28, 29–38. [Google Scholar] [CrossRef]
- Lazzari, L.; Corti, G.; Picco, G.; Isella, C.; Montone, M.; Arcela, P.; Durinikova, E.; Zanella, E.R.; Novara, L.; Barbosa, F.; et al. Patient-Derived Xenografts and Matched Cell Lines Identify Pharmacogenomic Vulnerabilities in Colorectal Cancer. Clin. Cancer Res. 2019, 25, 6243–6259. [Google Scholar] [CrossRef]
- Wagner, S.; Beger, N.T.; Matschos, S.; Szymanski, A.; Przybylla, R.; Bürtin, F.; Prall, F.; Linnebacher, M.; Mullins, C.S. Tumour-Derived Cell Lines and Their Potential for Therapy Prediction in Patients with Metastatic Colorectal Cancer. Cancers 2021, 13, 4717. [Google Scholar] [CrossRef] [PubMed]
- Verginelli, F.; Pisacane, A.; Gambardella, G.; D’Ambrosio, A.; Candiello, E.; Ferrio, M.; Panero, M.; Casorzo, L.; Benvenuti, S.; Cascardi, E.; et al. Cancer of unknown primary stem-like cells model multi-organ metastasis and unveil liability to MEK inhibition. Nat. Commun. 2021, 12, 2498. [Google Scholar] [CrossRef] [PubMed]
- Madan, B.; Harmston, N.; Nallan, G.; Montoya, A.; Faull, P.; Petretto, E.; Virshup, D.M. Temporal dynamics of Wnt-dependent transcriptome reveal an oncogenic Wnt/MYC/ribosome axis. J. Clin. Investig. 2018, 128, 5620–5633. [Google Scholar] [CrossRef] [PubMed]
- Hou, X.; Du, C.; Lu, L.; Yuan, S.; Zhan, M.; You, P.; Du, H. Opportunities and challenges of patient-derived models in cancer research: Patient-derived xenografts, patient-derived organoid and patient-derived cells. World J. Surg. Oncol. 2022, 20, 37. [Google Scholar] [CrossRef]
- Ivanics, T.; Bergquist, J.R.; Liu, G.; Kim, M.P.; Kang, Y.; Katz, M.H.; Perez, M.V.R.; Thomas, R.M.; Truty, M.J. Patient-derived xenograft cryopreservation and reanimation outcomes are dependent on cryoprotectant type. Lab. Investig. 2018, 98, 947–956. [Google Scholar] [CrossRef]
- Pyo, D.H.; Hong, H.K.; Lee, W.Y.; Cho, Y.B. Patient-derived cancer modeling for precision medicine in colorectal cancer: Beyond the cancer cell line. Cancer Biol. Ther. 2020, 21, 495–502. [Google Scholar] [CrossRef]
- Kanikarla Marie, P.; Sorokin, A.V.; Bitner, L.A.; Aden, R.; Lam, M.; Manyam, G.; Woods, M.N.; Anderson, A.; Capasso, A.; Fowlkes, N.; et al. Autologous humanized mouse models to study combination and single-agent immunotherapy for colorectal cancer patient-derived xenografts. Front. Oncol. 2022, 12, 994333. [Google Scholar] [CrossRef]
- Choi, Y.; Lee, S.; Kim, K.; Kim, S.H.; Chung, Y.J.; Lee, C. Studying cancer immunotherapy using patient-derived xenografts (PDXs) in humanized mice. Exp. Mol. Med. 2018, 50, 1–9. [Google Scholar] [CrossRef]
- Kwisda, K.; White, L.; Hübner, D. Ethical arguments concerning human-animal chimera research: A systematic review. BMC Med. Ethics 2020, 21, 24. [Google Scholar] [CrossRef]
- Inoue, A.; Deem, A.K.; Kopetz, S.; Heffernan, T.P.; Draetta, G.F.; Carugo, A. Current and future horizons of patient-derived xenograft models in colorectal cancer translational research. Cancers 2019, 11, 1321. [Google Scholar] [CrossRef]
- Shi, J.; Li, Y.; Jia, R.; Fan, X. The fidelity of cancer cells in PDX models: Characteristics, mechanism and clinical significance. Int. J. Cancer 2020, 146, 2078–2088. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.Q.; Ijiri, M.; Rodriguez, R.; Gandour-Edwards, R.; Lee, J.; Tepper, C.G.; Li, Y.; Beckett, L.; Lam, K.; Goodwin, N.; et al. Novel Patient Metastatic Pleural Effusion-Derived Xenograft Model of Renal Medullary Carcinoma Demonstrates Therapeutic Efficacy of Sunitinib. Front. Oncol. 2021, 11, 648097. [Google Scholar] [CrossRef] [PubMed]
- Tayoun, T.; Faugeroux, V.; Oulhen, M.; Aberlenc, A.; Pawlikowska, P.; Farace, F. CTC-Derived Models: A Window into the Seeding Capacity of Circulating Tumor Cells (CTCs). Cells 2019, 8, 1145. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Yu, Y. Patient-derived organoids in translational oncology and drug screening. Cancer Lett. 2023, 562, 216180. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Cong, L.; Cong, X. Patient-Derived Organoids in Precision Medicine: Drug Screening, Organoid-on-a-Chip and Living Organoid Biobank. Front. Oncol. 2021, 11, 762184. [Google Scholar] [CrossRef]
- Bae, J.; Choi, Y.S.; Cho, G.; Jang, S.J. The Patient-Derived Cancer Organoids: Promises and Challenges as Platforms for Cancer Discovery. Cancers 2022, 14, 2144. [Google Scholar] [CrossRef]
- Liu, Y.C.; Chen, Q.; Yang, X.L.; Tang, Q.S.; Yao, K.T.; Xu, Y. Generation of a new strain of NOD/SCID/IL2Rγ-/- mice with targeted disruption of Prkdc and IL2Rγ genes using CRISPR/Cas9 system. Nan Fang Yi Ke Da Xue Xue Bao 2018, 38, 639–646. [Google Scholar]
- Zhang, H.; Qi, L.; Du, Y.; Huang, L.F.; Braun, F.K.; Kogiso, M.; Zhao, Y.; Li, C.; Lindsay, H.; Zhao, S.; et al. Patient-derived orthotopic xenograft (PDOX) mouse models of primary and recurrent meningioma. Cancers 2020, 12, 1478. [Google Scholar] [CrossRef]
- Liu, C.; Wang, T.; Yang, J.; Zhang, J.; Wei, S.; Guo, Y.; Yu, R.; Tan, Z.; Wang, S.; Dong, W. Distant Metastasis Pattern and Prognostic Prediction Model of Colorectal Cancer Patients Based on Big Data Mining. Front. Oncol. 2022, 12, 878805. [Google Scholar] [CrossRef]
- Hsu, P.-L.; Chien, C.-W.; Tang, Y.-A.; Lin, B.-W.; Lin, S.-C.; Lin, Y.-S.; Chen, S.-Y.; Sun, H.S.; Tsai, S.-J. Targeting BRD3 eradicates nuclear TYRO3-induced colorectal cancer metastasis. Sci. Adv. 2023, 9, eade3422. Available online: https://www.science.org/doi/10.1126/sciadv.ade3422 (accessed on 30 November 2022). [CrossRef]
- Jian, M.; Ren, L.; Ren, L.; He, G.; He, G.; Lin, Q.; Chen, J.; Liu, T.; Ji, M.; Wei, Y.; et al. A novel patient-derived organoids-based xenografts model for preclinical drug response testing in patients with colorectal liver metastases. J. Transl. Med. 2020, 18, 234. [Google Scholar] [CrossRef] [PubMed]
- Andres, S.F.; Williams, K.N.; Rustgi, A.K. The Molecular Basis of Metastatic Colorectal Cancer. Curr. Color. Cancer Rep. 2018, 14, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Yu, Z.; Das, M.; Huang, L. Nano Codelivery of Oxaliplatin and Folinic Acid Achieves Synergistic Chemo-Immunotherapy with 5-Fluorouracil for Colorectal Cancer and Liver Metastasis. ACS Nano 2020, 14, 5075–5089. [Google Scholar] [CrossRef] [PubMed]
- Kasashima, H.; Duran, A.; Cid-Diaz, T.; Muta, Y.; Kinoshita, H.; Batlle, E.; Diaz-Meco, M.T.; Moscat, J. Mouse model of colorectal cancer: Orthotopic co-implantation of tumor and stroma cells in cecum and rectum. STAR Protoc. 2021, 2, 100297. [Google Scholar] [CrossRef] [PubMed]
- Rashidi, B.; Gamagami, R.; Sasson, A.; Sun, F.X.; Geller, J.; Moossa, A.R.; Hoffman, R.M. An orthotopic mouse model of remetastasis of human colon cancer liver metastasis. Clin. Cancer Res. 2000, 6, 2556–2561. [Google Scholar]
- Schwitalla, S.; Ziegler, P.K.; Horst, D.; Becker, V.; Kerle, I.; Begus-Nahrmann, Y.; Lechel, A.; Rudolph, K.L.; Langer, R.; Slotta-Huspenina, J.; et al. Loss of p53 in Enterocytes Generates an Inflammatory Microenvironment Enabling Invasion and Lymph Node Metastasis of Carcinogen-Induced Colorectal Tumors. Cancer Cell 2013, 23, 93–106. [Google Scholar] [CrossRef]
- Dow, L.E.; O’Rourke, K.P.; Simon, J.; Tschaharganeh, D.F.; Van Es, J.H.; Clevers, H.; Lowe, S.W. Apc restoration promotes cellular differentiation and reestablishes crypt homeostasis in colorectal cancer. Cell 2015, 161, 1539–1552. [Google Scholar] [CrossRef]
- Boutin, A.T.; Liao, W.T.; Wang, M.; Hwang, S.S.; Karpinets, T.V.; Cheung, H.; Chu, G.C.; Jiang, S.; Hu, J.; Chang, K.; et al. Oncogenic Kras drives invasion and maintains metastases in colorectal cancer. Genes Dev. 2017, 31, 370–382. [Google Scholar] [CrossRef]
- Rampetsreiter, P.; Casanova, E.; Eferl, R. Genetically modified mouse models of cancer invasion and metastasis. Drug. Discov. Today Dis. Model. 2011, 8, 67–74. [Google Scholar] [CrossRef]
- Takasago, T.; Hayashi, R.; Ueno, Y.; Ariyoshi, M.; Onishi, K.; Yamashita, K.; Hiyama, Y.; Takigawa, H.; Yuge, R.; Urabe, Y.; et al. Anti-tumor necrosis factor-alpha monoclonal antibody suppresses colorectal cancer growth in an orthotopic transplant mouse model. PLoS ONE 2023, 18, e0283822. [Google Scholar] [CrossRef]
- Zhang, Q.; Yang, X.; Wu, J.; Ye, S.; Gong, J.; Cheng, W.M.; Luo, Z.; Yu, J.; Liu, Y.; Zeng, W.; et al. Reprogramming of palmitic acid induced by dephosphorylation of ACOX1 promotes β-catenin palmitoylation to drive colorectal cancer progression. Cell. Discov. 2023, 9, 26. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.; Yao, B.; Liu, J.; Gong, G.-W.; Liu, M.; Li, J.; Pan, H.-F.; Li, Q.; Yang, D.; Lu, P.; et al. The SMARCA4R1157W mutation facilitates chromatin remodeling and confers PRMT1/SMARCA4 inhibitors sensitivity in colorectal cancer. npj Precis. Oncol. 2023, 7, 28. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Luo, H.; Zhang, J.; Li, K.; Lee, M.H. Therapeutic potential of Clostridium butyricum anticancer effects in colorectal cancer. Gut Microbes 2023, 15, 2186114. [Google Scholar] [CrossRef] [PubMed]
- Lattanzi, G.; Strati, F.; Díaz-Basabe, A.; Perillo, F.; Amoroso, C.; Protti, G.; Rita Giuffrè, M.; Iachini, L.; Baeri, A.; Baldari, L.; et al. iNKT cell-neutrophil crosstalk promotes colorectal cancer pathogenesis. Mucosal Immunol. 2023, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Cave, D.D.; Hernando-Momblona, X.; Sevillano, M.; Minchiotti, G.; Lonardo, E. Nodal-induced L1CAM/CXCR4 subpopulation sustains tumor growth and metastasis in colorectal cancer derived organoids. Theranostics 2021, 11, 5686–5699. [Google Scholar] [CrossRef]
- Zhang, Y.; Lee, S.H.; Wang, C.; Gao, Y.; Li, J.; Xu, W. Establishing metastatic patient-derived xenograft model for colorectal cancer. Jpn. J. Clin. Oncol. 2020, 50, 1108–1116. [Google Scholar] [CrossRef]
- Yang, H.; Li, X.; Meng, Q.; Sun, H.; Wu, S.; Hu, W.; Liu, G.; Li, X.; Yang, Y.; Chen, R. CircPTK2 (hsa_circ_0005273) as a novel therapeutic target for metastatic colorectal cancer. Mol. Cancer 2020, 19, 13. [Google Scholar] [CrossRef]
- Torchiaro, E.; Petti, C.; Arena, S.; Sassi, F.; Migliardi, G.; Mellano, A.; Porporato, R.; Basiricò, M.; Gammaitoni, L.; Berrino, E.; et al. Case report: Preclinical efficacy of NEDD8 and proteasome inhibitors in patient-derived models of signet ring high-grade mucinous colorectal cancer from a Lynch syndrome patient. Front. Oncol. 2023, 13, 1–8. Available online: https://www.frontiersin.org/articles/10.3389/fonc.2023.1130852/full (accessed on 30 November 2022). [CrossRef]
- Sun, H.; Liu, C.; Han, F.; Lin, X.; Cao, L.; Liu, C.; Ji, Q.; Cui, J.; Yao, Y.; Wang, B.; et al. The regulation loop of MARVELD1 interacting with PARP1 in DNA damage response maintains genome stability and promotes therapy resistance of cancer cells. Cell Death Differ. 2023, 30, 922–937. Available online: https://www.nature.com/articles/s41418-023-01118-z (accessed on 30 November 2022). [CrossRef]
- Shang, Y.; Zhu, Z.; Zhang, Y.; Ji, F.; Zhu, L.; Liu, M.; Deng, Y.; Lv, G.; Li, D.; Zhou, Z.; et al. MiR-7-5p/KLF4 signaling inhibits stemness and radioresistance in colorectal cancer. Cell Death Discov. 2023, 9, 42. Available online: https://www.nature.com/articles/s41420-023-01339-8 (accessed on 30 November 2022). [CrossRef]
- Fujii, M.; Shimokawa, M.; Date, S.; Takano, A.; Matano, M.; Nanki, K.; Ohta, Y.; Toshimitsu, K.; Nakazato, Y.; Kawasaki, K.; et al. A Colorectal Tumor Organoid Library Demonstrates Progressive Loss of Niche Factor Requirements during Tumorigenesis. Cell Stem Cell 2016, 18, 827–838. [Google Scholar] [CrossRef] [PubMed]
- Vlachogiannis, G.; Hedayat, S.; Vatsiou, A.; Jamin, Y.; Fernández-mateos, J.; Khan, K.; Lampis, A.; Eason, K.; Huntingford, I.; Burke, R.; et al. Patient-derived organoids model treatment response of metastatic gastrointestinal cancers. Science 2018, 359, 920–926. Available online: https://www.science.org/doi/10.1126/science.aao2774 (accessed on 30 November 2022). [CrossRef]
- Kersten, K.; Visser, K.E.; Miltenburg, M.H.; Jonkers, J. Genetically engineered mouse models in oncology research and cancer medicine. EMBO Mol. Med. 2017, 9, 137–153. [Google Scholar] [CrossRef] [PubMed]
- Avolio, M.; Trusolino, L. Rational treatment of metastatic colorectal cancer: A reverse tale of men, mice, and culture dishes. Cancer Discov. 2021, 11, 1644–1660. [Google Scholar] [CrossRef] [PubMed]
- Kwon, J.; Oh, S.; Park, M.; Kong, J.S.; Lee, S.; Lee, H.; Kim, Y.; Kang, K.-T.; Shin, U.S.; Jung, J. Advanced Xenograft Model with Cotransplantation of Patient-Derived Organoids and Endothelial Colony-Forming Cells for Precision Medicine. J. Oncol. 2021, 2021, 9994535. [Google Scholar] [CrossRef]
- Kang, K.T.; Lin, R.Z.; Kuppermann, D.; Melero-Martin, J.M.; Bischoff, J. Endothelial colony forming cells and mesenchymal progenitor cells form blood vessels and increase blood flow in ischemic muscle. Sci. Rep. 2017, 7, 770. [Google Scholar] [CrossRef]
- Matano, M.; Date, S.; Shimokawa, M.; Takano, A.; Fujii, M.; Ohta, Y.; Watanabe, T.; Kanai, T.; Sato, T. Modeling colorectal cancer using CRISPR-Cas9-mediated engineering of human intestinal organoids. Nat. Med. 2015, 21, 256–262. [Google Scholar] [CrossRef]
- Liu, X.; Su, Q.; Zhang, X.; Yang, W.; Ning, J.; Jia, K.; Xin, J.; Li, H.; Yu, L.; Liao, Y.; et al. Recent Advances of Organ-on-a-Chip in Cancer Modeling Research. Biosensors 2022, 12, 1045. Available online: http://www.ncbi.nlm.nih.gov/pubmed/36421163%0Ahttp://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=PMC9688857 (accessed on 20 December 2022). [CrossRef]
- Guinney, J.; Dienstmann, R.; Wang, X.; De Reyniès, A.; Schlicker, A.; Soneson, C.; Marisa, L.; Roepman, P.; Nyamundanda, G.; Angelino, P.; et al. The consensus molecular subtypes of colorectal cancer. Nat. Med. 2015, 21, 1350–1356. [Google Scholar] [CrossRef]
- Borelli, B.; Fontana, E.; Giordano, M.; Antoniotti, C.; Lonardi, S.; Bergamo, F.; Pietrantonio, F.; Morano, F.; Tamburini, E.; Boccaccino, A.; et al. Prognostic and predictive impact of consensus molecular subtypes and CRCAssigner classifications in metastatic colorectal cancer: A translational analysis of the TRIBE2 study. ESMO Open 2021, 6, 100073. [Google Scholar] [CrossRef]
- Stillman, N.R.; Kovacevic, M.; Balaz, I.; Hauert, S. In silico modelling of cancer nanomedicine, across scales and transport barriers. npj Comput. Mater. 2020, 6, 92. [Google Scholar] [CrossRef]
- Jean-Quartier, C.; Jeanquartier, F.; Jurisica, I.; Holzinger, A. In silico cancer research towards 3R. BMC Cancer 2018, 18, 408. [Google Scholar] [CrossRef]
- Cruz, S.; Gomes, S.E.; Borralho, P.M.; Rodrigues, C.M.P.; Gaudêncio, S.P.; Pereira, F. In silico HCT116 human colon cancer cell-based models en route to the discovery of lead-like anticancer drugs. Biomolecules 2018, 8, 56. [Google Scholar] [CrossRef] [PubMed]
- Gold, A.; Choueiry, F.; Jin, N.; Mo, X.; Zhu, J. The Application of Metabolomics in Recent Colorectal Cancer Studies: A State-of-the-Art Review. Cancers 2022, 14, 725. [Google Scholar] [CrossRef] [PubMed]
- Gulfidan, G.; Turanli, B.; Beklen, H.; Sinha, R.; Arga, K.Y. Pan-cancer mapping of differential protein-protein interactions. Sci. Rep. 2020, 10, 3272. [Google Scholar] [CrossRef] [PubMed]
- Uhlen, M.; Zhang, C.; Lee, S.; Sjöstedt, E.; Fagerberg, L.; Bidkhori, G.; Benfeitas, R.; Arif, M.; Liu, Z.; Edfors, F.; et al. A pathology atlas of the human cancer transcriptome. Science 2017, 357, 6352. [Google Scholar] [CrossRef]
- Subramanian, A.; Narayan, R.; Corsello, S.M.; Peck, D.D.; Natoli, T.E.; Lu, X.; Gould, J.; Davis, J.F.; Tubelli, A.A.; Asiedu, J.K.; et al. A Next Generation Connectivity Map: L1000 Platform and the First 1,000,000 Profiles. Cell 2017, 171, 1437–1452.e17. [Google Scholar] [CrossRef]
- Rintala, T.J.; Ghosh, A.; Fortino, V. Network approaches for modeling the effect of drugs and diseases. Brief Bioinform. 2022, 23, bbac229. [Google Scholar] [CrossRef]


| Models | CIMs | GEMMs | TMMs | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Features | DMH | AOM | HCA | DMAB | AS | APMM | HNPCC | CDX | PDX | PDOX | MM | |
| Complexity | + | + | + | + | + | +++ | +++ | ++ a | ++ | +++ | +++ | |
| Time consuming | ++ | ++ | ++ | ++ | ++ | +++ | +++ | ++ a | +++ | +++ | ++ | |
| Cost | + | + | + | + | + | +++ | +++ | ++ | +++ | +++ | +++ | |
| Surgical skills | + | + | + | + | + | ++ | ++ | ++ a | +++ | +++ | +++ | |
| Translational models | ++ | ++ | + | + | + | +++ | +++ | ++ | ++ | +++ | ++ | |
| Tumor heterogeneity | +++ | +++ | ++ | ++ | ++ | ++ | ++ | ++ | +++ | +++ | ++ | |
| Tumor microenvironment | +++ | +++ | + | + | + | +++ | +++ | + | + c | ++ | ++ | |
| Engraftment rate | +++ | +++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | +++ | ++ | |
| Metastases | + | + | + | + | + | ++ | ++ | ++ a | ++ | ++ | +++ | |
| Precision medicine | ++ | ++ | + | + | + | ++ | ++ | + | ++ | +++ | ++ | |
| Chemotherapy studies | +++ | +++ | + | + | + | ++ | ++ | +++ | ++ | +++ | +++ | |
| Immuno-oncology | ++ | ++ | + | + | + | +++ | +++ | ++ d | + b | ++ | + | |
| High-throughput omics | + | + | + | + | + | ++ | ++ | + | ++ | +++ | + | |
| Drug discovery | +++ | +++ | + | + | ++ | ++ | ++ | +++ | ++ | ++ | ++ | |
| Biomarker discovery | ++ | ++ | + | + | + | +++ | +++ | ++ | +++ | +++ | ++ | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Neto, Í.; Rocha, J.; Gaspar, M.M.; Reis, C.P. Experimental Murine Models for Colorectal Cancer Research. Cancers 2023, 15, 2570. https://doi.org/10.3390/cancers15092570
Neto Í, Rocha J, Gaspar MM, Reis CP. Experimental Murine Models for Colorectal Cancer Research. Cancers. 2023; 15(9):2570. https://doi.org/10.3390/cancers15092570
Chicago/Turabian StyleNeto, Íris, João Rocha, Maria Manuela Gaspar, and Catarina P. Reis. 2023. "Experimental Murine Models for Colorectal Cancer Research" Cancers 15, no. 9: 2570. https://doi.org/10.3390/cancers15092570
APA StyleNeto, Í., Rocha, J., Gaspar, M. M., & Reis, C. P. (2023). Experimental Murine Models for Colorectal Cancer Research. Cancers, 15(9), 2570. https://doi.org/10.3390/cancers15092570