
Identification of F-Box/SPRY Domain-Containing Protein 1 (FBXO45) as a Prognostic Biomarker for TMPRSS2–ERG-Positive Primary Prostate Cancers

 , , , , , ,
, , , , , ,  and
and
Abstract
Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Transcriptome Data Assembly
2.2. Immunohistochemistry
2.3. Cell Lines and Culture Conditions
2.4. Antisense LNA GapmeR-Mediated Knockdown
2.5. RNA Isolation
2.6. Real-Time PCR
2.7. Western Blot
2.8. Cell Proliferation Assays
2.9. Migration Assays
2.10. Statistical Analysis
2.11. Ethical Approval and Consent to Participate
3. Results
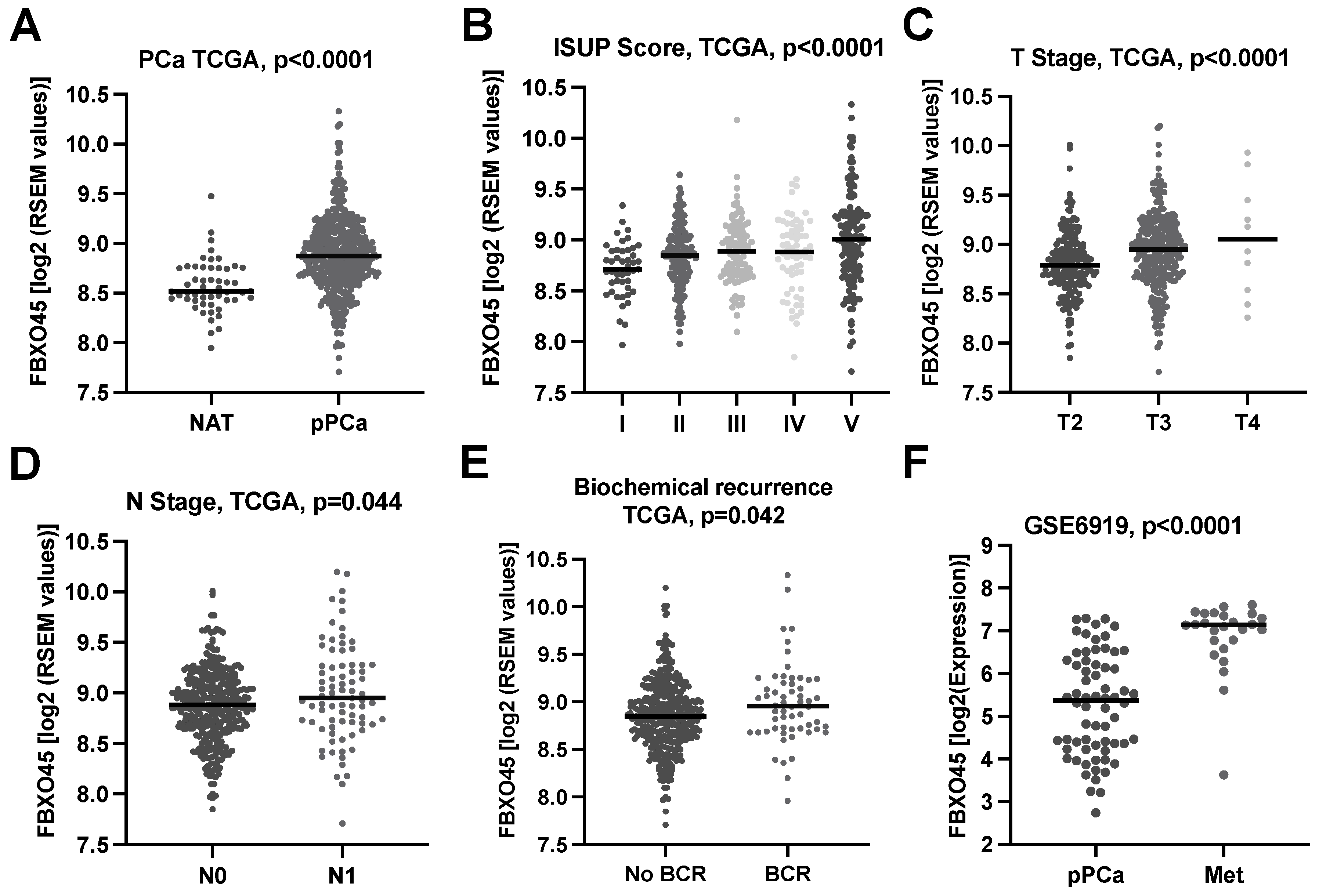
3.1. F-Box/SPRY Domain-Containing Protein 1 (FBXO45) In Silico Expression Analysis
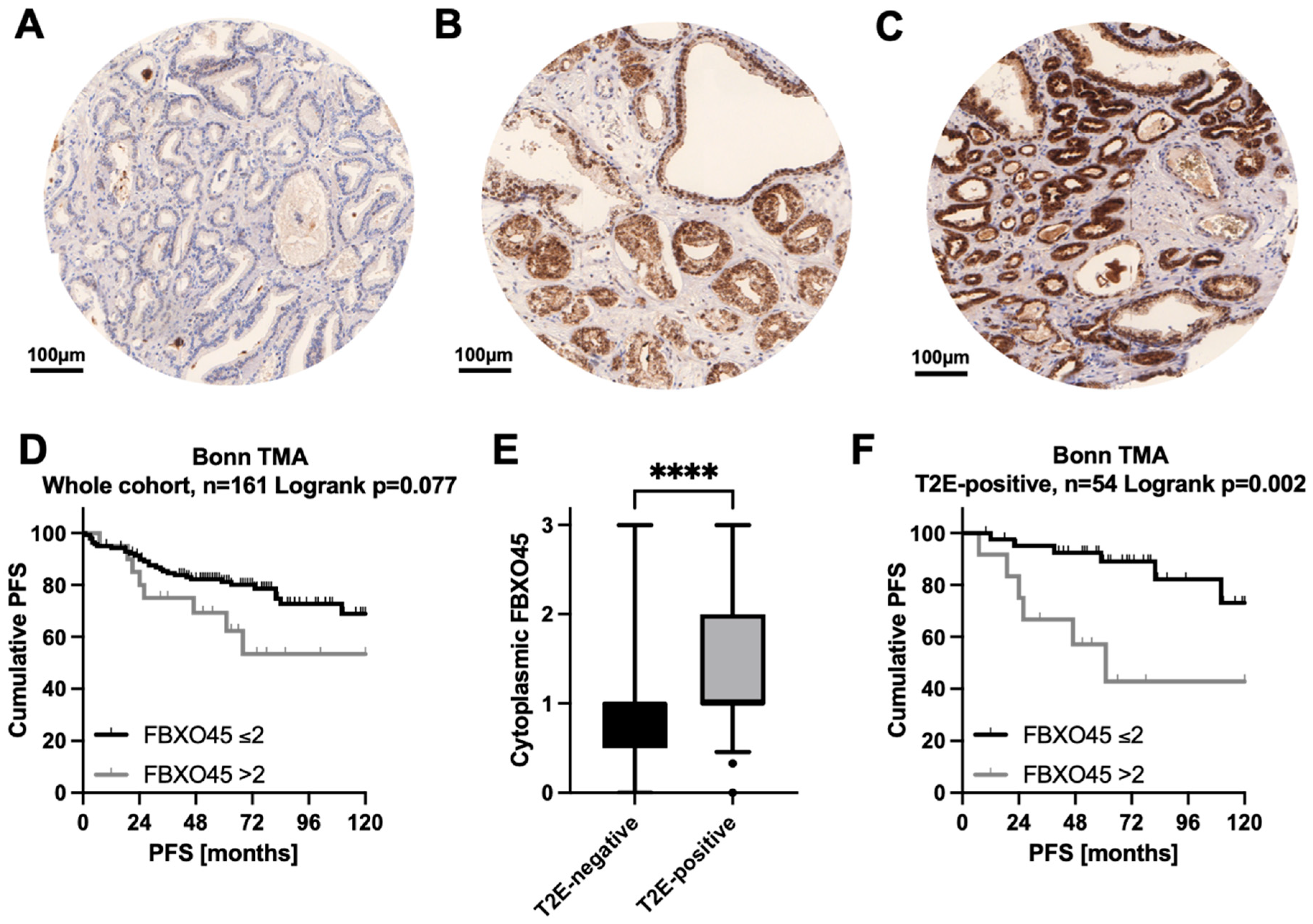
3.2. FBXO45 Protein Expression on a PCA Tissue Microarray (TMA)
3.3. Functional Characterization of FBXO45 In Vitro
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Schröder, F.H.; Hugosson, J.; Roobol, M.J.; Tammela, T.L.J.; Ciatto, S.; Nelen, V.; Kwiatkowski, M.; Lujan, M.; Lilja, H.; Zappa, M.; et al. Screening and Prostate-Cancer Mortality in a Randomized European Study. N. Engl. J. Med. 2009, 360, 1320–1328. [Google Scholar] [CrossRef] [PubMed]
- Dellis, A.; Zagouri, F.; Liontos, M.; Mitropoulos, D.; Bamias, A.; Papatsoris, A.G. Management of Advanced Prostate Cancer: A Systematic Review of Existing Guidelines and Recommendations. Cancer Treat. Rev. 2019, 73, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Gillessen, S.; Attard, G.; Beer, T.M.; Beltran, H.; Bjartell, A.; Bossi, A.; Briganti, A.; Bristow, R.G.; Chi, K.N.; Clarke, N.; et al. Management of Patients with Advanced Prostate Cancer: Report of the Advanced Prostate Cancer Consensus Conference 2019. Eur. Urol. 2020, 77, 508–547. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Tang, F.; Lu, Z.; Huang, Y.; Lei, H.; Li, Z.; Zeng, G. Analysis of Differentially Expressed Genes, Clinical Value and Biological Pathways in Prostate Cancer. Am. J. Transl. Res. 2018, 10, 1444–1456. [Google Scholar]
- Gerke, J.S.; Orth, M.F.; Tolkach, Y.; Romero-Pérez, L.; Wehweck, F.S.; Stein, S.; Musa, J.; Knott, M.M.L.; Hölting, T.L.B.; Li, J.; et al. Integrative Clinical Transcriptome Analysis Reveals TMPRSS2-ERG Dependency of Prognostic Biomarkers in Prostate Adenocarcinoma. Int. J. Cancer 2020, 146, 2036–2046. [Google Scholar] [CrossRef]
- Cenciarelli, C.; Chiaur, D.S.; Guardavaccaro, D.; Parks, W.; Vidal, M.; Pagano, M. Identification of a Family of Human F-Box Proteins. Curr. Biol. 1999, 9, 1177–1179. [Google Scholar] [CrossRef]
- Jin, J.; Cardozo, T.; Lovering, R.C.; Elledge, S.J.; Pagano, M.; Harper, J.W. Systematic Analysis and Nomenclature of Mammalian F-Box Proteins. Genes Dev. 2004, 18, 2573–2580. [Google Scholar] [CrossRef]
- Skowyra, D.; Craig, K.L.; Tyers, M.; Elledge, S.J.; Harper, J.W. F-Box Proteins Are Receptors That Recruit Phosphorylated Substrates to the SCF Ubiquitin-Ligase Complex. Cell 1997, 91, 209–219. [Google Scholar] [CrossRef]
- Liao, E.H.; Hung, W.; Abrams, B.; Zhen, M. An SCF-like Ubiquitin Ligase Complex That Controls Presynaptic Differentiation. Nature 2004, 430, 345–350. [Google Scholar] [CrossRef]
- Chen, X.; Sahasrabuddhe, A.A.; Szankasi, P.; Chung, F.; Basrur, V.; Rangnekar, V.M.; Pagano, M.; Lim, M.S.; Elenitoba-Johnson, K.S.J. Fbxo45-Mediated Degradation of the Tumor-Suppressor Par-4 Regulates Cancer Cell Survival. Cell Death Differ. 2014, 21, 1535–1545. [Google Scholar] [CrossRef] [PubMed]
- Peschiaroli, A.; Scialpi, F.; Bernassola, F.; Pagano, M.; Melino, G. The F-Box Protein FBXO45 Promotes the Proteasome-Dependent Degradation of P73. Oncogene 2009, 28, 3157–3166. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Wei, W. Fbxo45 Joins the ‘Par-4’ty in Controlling Apoptosis of Cancer Cells. Cell Death Differ. 2014, 21, 1508–1510. [Google Scholar] [CrossRef] [PubMed]
- Abshire, C.F.; Carroll, J.L.; Dragoi, A.-M. FLASH Protects ZEB1 from Degradation and Supports Cancer Cells’ Epithelial-to-Mesenchymal Transition. Oncogenesis 2016, 5, e254. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, M.-C.; Ho, Y.-C.; Lai, C.-Y.; Chou, D.; Chen, G.-D.; Lin, T.-B.; Peng, H.-Y. Spinal TNF-α Impedes Fbxo45-Dependent Munc13-1 Ubiquitination to Mediate Neuropathic Allodynia in Rats. Cell Death Dis. 2018, 9, 811. [Google Scholar] [CrossRef]
- Kogure, N.; Yokobori, T.; Ogata, K.; Altan, B.; Mochiki, E.; Ohno, T.; Toyomasu, Y.; Yanai, M.; Kimura, A.; Yanoma, T.; et al. Low Expression of FBXO45 Is Associated with Gastric Cancer Progression and Poor Prognosis. Anticancer Res. 2017, 37, 191–196. [Google Scholar] [CrossRef]
- Wang, K.; Qu, X.; Liu, S.; Yang, X.; Bie, F.; Wang, Y.; Huang, C.; Du, J. Identification of Aberrantly Expressed F-Box Proteins in Squamous-Cell Lung Carcinoma. J. Cancer Res. Clin. Oncol. 2018, 144, 1509–1521. [Google Scholar] [CrossRef]
- Lin, M.; Wang, Z.; Zhu, X. FBXO45 Is a Potential Therapeutic Target for Cancer Therapy. Cell Death Discov. 2020, 6, 55. [Google Scholar] [CrossRef]
- Chandran, U.R.; Ma, C.; Dhir, R.; Bisceglia, M.; Lyons-Weiler, M.; Liang, W.; Michalopoulos, G.; Becich, M.; Monzon, F.A. Gene Expression Profiles of Prostate Cancer Reveal Involvement of Multiple Molecular Pathways in the Metastatic Process. BMC Cancer 2007, 7, 64. [Google Scholar] [CrossRef]
- Stein, J.; Majores, M.; Rohde, M.; Lim, S.; Schneider, S.; Krappe, E.; Ellinger, J.; Dietel, M.; Stephan, C.; Jung, K.; et al. KDM5C Is Overexpressed in Prostate Cancer and Is a Prognostic Marker for Prostate-Specific Antigen-Relapse Following Radical Prostatectomy. Am. J. Pathol. 2014, 184, 2430–2437. [Google Scholar] [CrossRef]
- Mang, J.; Korzeniewski, N.; Dietrich, D.; Sailer, V.; Tolstov, Y.; Searcy, S.; von Hardenberg, J.; Perner, S.; Kristiansen, G.; Marx, A.; et al. Prognostic Significance and Functional Role of CEP57 in Prostate Cancer. Transl. Oncol. 2015, 8, 487–496. [Google Scholar] [CrossRef] [PubMed]
- Klümper, N.; von Danwitz, M.; Stein, J.; Schmidt, D.; Schmidt, A.; Kristiansen, G.; Muders, M.; Hölzel, M.; Ritter, M.; Alajati, A.; et al. Downstream Neighbor of SON (DONSON) Expression Is Enhanced in Phenotypically Aggressive Prostate Cancers. Cancers 2020, 12, 3439. [Google Scholar] [CrossRef] [PubMed]
- Stael, S.; Miller, L.P.; Fernández-Fernández, Á.D.; Van Breusegem, F. Detection of Damage-Activated Metacaspase ActivityActivitiesby Western Blot in Plants. In Plant Proteases and Plant Cell Death: Methods and Protocols; Klemenčič, M., Stael, S., Huesgen, P.F., Eds.; Methods in Molecular Biology; Springer: New York, NY, USA, 2022; pp. 127–137. ISBN 978-1-07-162079-3. [Google Scholar]
- Abeshouse, A.; Ahn, J.; Akbani, R.; Ally, A.; Amin, S.; Andry, C.D.; Annala, M.; Aprikian, A.; Armenia, J.; Arora, A.; et al. The Molecular Taxonomy of Primary Prostate Cancer. Cell 2015, 163, 1011–1025. [Google Scholar] [CrossRef] [PubMed]
- Penney, K.L.; Pettersson, A.; Shui, I.M.; Graff, R.E.; Kraft, P.; Lis, R.T.; Sesso, H.D.; Loda, M.; Mucci, L.A. Association of Prostate Cancer Risk Variants with TMPRSS2:ERG Status: Evidence for Distinct Molecular Subtypes. Cancer Epidemiol. Biomark. Prev. 2016, 25, 745–749. [Google Scholar] [CrossRef]
- Adamo, P.; Ladomery, M.R. The Oncogene ERG: A Key Factor in Prostate Cancer. Oncogene 2016, 35, 403–414. [Google Scholar] [CrossRef]
- Tomlins, S.A.; Rhodes, D.R.; Perner, S.; Dhanasekaran, S.M.; Mehra, R.; Sun, X.-W.; Varambally, S.; Cao, X.; Tchinda, J.; Kuefer, R.; et al. Recurrent Fusion of TMPRSS2 and ETS Transcription Factor Genes in Prostate Cancer. Science 2005, 310, 644–648. [Google Scholar] [CrossRef]
- Kim, T.S.; Heinlein, C.; Hackman, R.C.; Nelson, P.S. Phenotypic Analysis of Mice Lacking the Tmprss2-Encoded Protease. Mol. Cell. Biol. 2006, 26, 965–975. [Google Scholar] [CrossRef]
- Carver, B.S.; Tran, J.; Chen, Z.; Carracedo-Perez, A.; Alimonti, A.; Nardella, C.; Gopalan, A.; Scardino, P.T.; Cordon-Cardo, C.; Gerald, W.; et al. ETS Rearrangements and Prostate Cancer Initiation. Nature 2009, 457, E1. [Google Scholar] [CrossRef]
- Deramaudt, T.B.; Remy, P.; Stiegler, P. Identification of Interaction Partners for Two Closely-Related Members of the ETS Protein Family, FLI and ERG. Gene 2001, 274, 169–177. [Google Scholar] [CrossRef]
- Shimizu, K.; Ichikawa, H.; Tojo, A.; Kaneko, Y.; Maseki, N.; Hayashi, Y.; Ohira, M.; Asano, S.; Ohki, M. An Ets-Related Gene, ERG, Is Rearranged in Human Myeloid Leukemia with t(16;21) Chromosomal Translocation. Proc. Natl. Acad. Sci. USA 1993, 90, 10280–10284. [Google Scholar] [CrossRef]
- Tomlins, S.A.; Bjartell, A.; Chinnaiyan, A.M.; Jenster, G.; Nam, R.K.; Rubin, M.A.; Schalken, J.A. ETS Gene Fusions in Prostate Cancer: From Discovery to Daily Clinical Practice. Eur. Urol. 2009, 56, 275–286. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Yu, K.; Chen, K.; Zhu, X.; Yang, Z.; Wang, Q.; Gao, J.; Wang, Y.; Cao, T.; Xu, H.; et al. Fbxo45 Facilitates Pancreatic Carcinoma Progression by Targeting USP49 for Ubiquitination and Degradation. Cell Death Dis. 2022, 13, 231. [Google Scholar] [CrossRef]
- Burikhanov, R.; Rangnekar, V.M. Par-4 as a Therapeutic Target in Cancer and Other Diseases. In Tumor Suppressor Par-4: Role in Cancer and Other Diseases; Rangnekar, V.M., Ed.; Springer International Publishing: Cham, Switzerland, 2021; pp. 313–320. ISBN 978-3-030-80558-6. [Google Scholar]
- Qiu, X.; Boufaied, N.; Hallal, T.; Feit, A.; de Polo, A.; Luoma, A.M.; Alahmadi, W.; Larocque, J.; Zadra, G.; Xie, Y.; et al. MYC Drives Aggressive Prostate Cancer by Disrupting Transcriptional Pause Release at Androgen Receptor Targets. Nat. Commun. 2022, 13, 2559. [Google Scholar] [CrossRef] [PubMed]
- Reiner, T.; de las Pozas, A.; Parrondo, R.; Palenzuela, D.; Cayuso, W.; Rai, P.; Perez-Stable, C. Mcl-1 Protects Prostate Cancer Cells from Cell Death Mediated by Chemotherapy-Induced DNA Damage. Oncoscience 2015, 2, 703–715. [Google Scholar] [CrossRef] [PubMed]
- Richter, K.T.; Kschonsak, Y.T.; Vodicska, B.; Hoffmann, I. FBXO45-MYCBP2 Regulates Mitotic Cell Fate by Targeting FBXW7 for Degradation. Cell Death Differ. 2020, 27, 758–772. [Google Scholar] [CrossRef]
- Ganapathy, K.; Ngo, C.; Andl, T.; Coppola, D.; Park, J.; Chakrabarti, R. Anticancer Function of microRNA -30e Is Mediated by Negative Regulation of HELLPAR, a Noncoding macroRNA, and Genes Involved in Ubiquitination and Cell Cycle Progression in Prostate Cancer. Mol. Oncol. 2022, 16, 2936–2958. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Parameters | p-Value | Hazard Ratio | 95% Confidence Interval | |
|---|---|---|---|---|
| Low | High | |||
| FBX045 | 0.022 | 4.483 | 1.246 | 16.136 |
| pT | 0.372 | 1.943 | 0.452 | 8.357 |
| pN | 0.471 | 0.404 | 0.034 | 4.751 |
| ISUP | 0.636 | 0.972 | 0.866 | 1.092 |
| Age | 0.636 | 0.972 | 0.866 | 1.092 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
von Danwitz, M.; Klümper, N.; Bernhardt, M.; Cox, A.; Krausewitz, P.; Alajati, A.; Kristiansen, G.; Ritter, M.; Ellinger, J.; Stein, J. Identification of F-Box/SPRY Domain-Containing Protein 1 (FBXO45) as a Prognostic Biomarker for TMPRSS2–ERG-Positive Primary Prostate Cancers. Cancers 2023, 15, 1890. https://doi.org/10.3390/cancers15061890
von Danwitz M, Klümper N, Bernhardt M, Cox A, Krausewitz P, Alajati A, Kristiansen G, Ritter M, Ellinger J, Stein J. Identification of F-Box/SPRY Domain-Containing Protein 1 (FBXO45) as a Prognostic Biomarker for TMPRSS2–ERG-Positive Primary Prostate Cancers. Cancers. 2023; 15(6):1890. https://doi.org/10.3390/cancers15061890
Chicago/Turabian Stylevon Danwitz, Marthe, Niklas Klümper, Marit Bernhardt, Alexander Cox, Philipp Krausewitz, Abdullah Alajati, Glen Kristiansen, Manuel Ritter, Jörg Ellinger, and Johannes Stein. 2023. "Identification of F-Box/SPRY Domain-Containing Protein 1 (FBXO45) as a Prognostic Biomarker for TMPRSS2–ERG-Positive Primary Prostate Cancers" Cancers 15, no. 6: 1890. https://doi.org/10.3390/cancers15061890
APA Stylevon Danwitz, M., Klümper, N., Bernhardt, M., Cox, A., Krausewitz, P., Alajati, A., Kristiansen, G., Ritter, M., Ellinger, J., & Stein, J. (2023). Identification of F-Box/SPRY Domain-Containing Protein 1 (FBXO45) as a Prognostic Biomarker for TMPRSS2–ERG-Positive Primary Prostate Cancers. Cancers, 15(6), 1890. https://doi.org/10.3390/cancers15061890