
The BAFF-APRIL System in Cancer



Abstract
Simple Summary
Abstract
1. Introduction
2. Expression Profile of BAFF, APRIL and Receptors
3. BAFF and APRIL Signaling in Immune Homeostasis
3.1. Effect on B Cells
3.1.1. B Cell Development and Differentiation
3.1.2. B-Cell Function
3.2. Effect on T Cells
3.3. Effect on Myeloid Cells
3.4. Effect on Other Immune and Non-Immune Cells
4. BAFF-APRIL System in the Pathogenesis of Inflammatory Diseases
5. BAFF and APRIL in Cancer Pathogenesis
5.1. Hematological Cancers
5.1.1. Acute Lymphoblastic Leukemia (B-ALL)
5.1.2. Hodgkin’s Lymphoma (HL)
5.1.3. Multiple Myeloma (MM)
5.1.4. Burkitt’s Lymphoma (BL)
5.1.5. Chronic Lymphocytic Leukemia (CLL)
5.1.6. Central Nervous System Lymphoma (CNSL)
5.1.7. Diffuse Large B-Cell Lymphoma (DLBCL)
5.1.8. Follicular Lymphoma (FL)
5.1.9. Hairy Cell Leukemia (HCL)
5.1.10. Mantle Cell Lymphoma (MCL)
5.1.11. Marginal Zone Lymphoma (MZL)
5.2. Solid Cancers
5.2.1. Breast Cancer
5.2.2. Colorectal Cancer (CRC)
5.2.3. Glioma
5.2.4. Hepatocellular Carcinoma
5.2.5. Lung Cancer
5.2.6. Skin Cancers
5.2.7. Other Solid Cancers
6. Therapeutic Targeting of the BAFF/APRIL Pathway
6.1. BAFF Targeted Therapies
6.1.1. Belimumab
6.1.2. Tabalumab
6.1.3. BAFF Expressing CAR-T Cell
6.2. APRIL Targeted Therapies
6.3. BAFF-R Targeted Therapies
6.4. BCMA Target Therapies
6.4.1. Bi-Specific Antibodies
6.4.2. Antibody-Drug Conjugates (ADC)
6.4.3. BCMA Expressing CAR-T Cells
6.4.4. BCMA Decoy Receptor
6.5. TACI Target Therapies
7. Conclusions and Perspective
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Schneider, P.; MacKay, F.; Steiner, V.; Hofmann, K.; Bodmer, J.L.; Holler, N.; Ambrose, C.; Lawton, P.; Bixler, S.; Acha-Orbea, H.; et al. BAFF, a novel ligand of the tumor necrosis factor family, stimulates B cell growth. J. Exp. Med. 1999, 189, 1747–1756. [Google Scholar] [CrossRef] [PubMed]
- Hahne, M.; Kataoka, T.; Schröter, M.; Hofmann, K.; Irmler, M.; Bodmer, J.L.; Schneider, P.; Bornand, T.; Holler, N.; French, L.E.; et al. APRIL, a new ligand of the tumor necrosis factor family, stimulates tumor cell growth. J. Exp. Med. 1998, 188, 1185–1190. [Google Scholar] [CrossRef] [PubMed]
- Roschke, V.; Sosnovtseva, S.; Ward, C.D.; Hong, J.S.; Smith, R.; Albert, V.; Stohl, W.; Baker, K.P.; Ullrich, S.; Nardelli, B.; et al. BLyS and APRIL form biologically active heterotrimers that are expressed in patients with systemic immune-based rheumatic diseases. J. Immunol. 2002, 169, 4314–4321. [Google Scholar] [CrossRef]
- Wallweber, H.J.; Compaan, D.M.; Starovasnik, M.A.; Hymowitz, S.G. The crystal structure of a proliferation-inducing ligand, APRIL. J. Mol. Biol. 2004, 343, 283–290. [Google Scholar] [CrossRef]
- Liu, Y.; Hong, X.; Kappler, J.; Jiang, L.; Zhang, R.; Xu, L.; Pan, C.H.; Martin, W.E.; Murphy, R.C.; Shu, H.B.; et al. Ligand-receptor binding revealed by the TNF family member TALL-1. Nature 2003, 423, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Day, E.S.; Cachero, T.G.; Qian, F.; Sun, Y.; Wen, D.; Pelletier, M.; Hsu, Y.M.; Whitty, A. Selectivity of BAFF/BLyS and APRIL for binding to the TNF family receptors BAFFR/BR3 and BCMA. Biochemistry 2005, 44, 1919–1931. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Bressette, D.; Carrell, J.A.; Kaufman, T.; Feng, P.; Taylor, K.; Gan, Y.; Cho, Y.H.; Garcia, A.D.; Gollatz, E.; et al. Tumor necrosis factor (TNF) receptor superfamily member TACI is a high affinity receptor for TNF family members APRIL and BLyS. J. Biol. Chem. 2000, 275, 35478–35485. [Google Scholar] [CrossRef]
- Schneider, P.; Takatsuka, H.; Wilson, A.; Mackay, F.; Tardivel, A.; Lens, S.; Cachero, T.G.; Finke, D.; Beermann, F.; Tschopp, J. Maturation of marginal zone and follicular B cells requires B cell activating factor of the tumor necrosis factor family and is independent of B cell maturation antigen. J. Exp. Med. 2001, 194, 1691–1697. [Google Scholar] [CrossRef]
- Hoffmann, F.S.; Kuhn, P.H.; Laurent, S.A.; Hauck, S.M.; Berer, K.; Wendlinger, S.A.; Krumbholz, M.; Khademi, M.; Olsson, T.; Dreyling, M.; et al. The immunoregulator soluble TACI is released by ADAM10 and reflects B cell activation in autoimmunity. J. Immunol. 2015, 194, 542–552. [Google Scholar] [CrossRef]
- Laurent, S.A.; Hoffmann, F.S.; Kuhn, P.H.; Cheng, Q.; Chu, Y.; Schmidt-Supprian, M.; Hauck, S.M.; Schuh, E.; Krumbholz, M.; Rübsamen, H.; et al. γ-Secretase directly sheds the survival receptor BCMA from plasma cells. Nat. Commun. 2015, 6, 7333. [Google Scholar] [CrossRef]
- Thompson, J.S.; Bixler, S.A.; Qian, F.; Vora, K.; Scott, M.L.; Cachero, T.G.; Hession, C.; Schneider, P.; Sizing, I.D.; Mullen, C.; et al. BAFF-R, a newly identified TNF receptor that specifically interacts with BAFF. Science 2001, 293, 2108–2111. [Google Scholar] [CrossRef] [PubMed]
- Hendriks, J.; Planelles, L.; de Jong-Odding, J.; Hardenberg, G.; Pals, S.T.; Hahne, M.; Spaargaren, M.; Medema, J.P. Heparan sulfate proteoglycan binding promotes APRIL-induced tumor cell proliferation. Cell Death Differ. 2005, 12, 637–648. [Google Scholar] [CrossRef] [PubMed]
- Moore, P.A.; Belvedere, O.; Orr, A.; Pieri, K.; LaFleur, D.W.; Feng, P.; Soppet, D.; Charters, M.; Gentz, R.; Parmelee, D.; et al. BLyS: Member of the tumor necrosis factor family and B lymphocyte stimulator. Science 1999, 285, 260–263. [Google Scholar] [CrossRef] [PubMed]
- Mackay, F.; Schneider, P. Cracking the BAFF code. Nat. Rev. Immunol. 2009, 9, 491–502. [Google Scholar] [CrossRef] [PubMed]
- Vincent, F.B.; Saulep-Easton, D.; Figgett, W.A.; Fairfax, K.A.; Mackay, F. The BAFF/APRIL system: Emerging functions beyond B cell biology and autoimmunity. Cytokine Growth Factor Rev. 2013, 24, 203–215. [Google Scholar] [CrossRef]
- Litinskiy, M.B.; Nardelli, B.; Hilbert, D.M.; He, B.; Schaffer, A.; Casali, P.; Cerutti, A. DCs induce CD40-independent immunoglobulin class switching through BLyS and APRIL. Nat. Immunol. 2002, 3, 822–829. [Google Scholar] [CrossRef]
- Stein, J.V.; López-Fraga, M.; Elustondo, F.A.; Carvalho-Pinto, C.E.; Rodríguez, D.; Gómez-Caro, R.; De Jong, J.; Martínez, A.C.; Medema, J.P.; Hahne, M. APRIL modulates B and T cell immunity. J. Clin. Investig. 2002, 109, 1587–1598. [Google Scholar] [CrossRef]
- de Graaf, C.A.; Choi, J.; Baldwin, T.M.; Bolden, J.E.; Fairfax, K.A.; Robinson, A.J.; Biben, C.; Morgan, C.; Ramsay, K.; Ng, A.P.; et al. Haemopedia: An Expression Atlas of Murine Hematopoietic Cells. Stem Cell Rep. 2016, 7, 571–582. [Google Scholar] [CrossRef]
- Suzuki, K.; Setoyama, Y.; Yoshimoto, K.; Tsuzaka, K.; Abe, T.; Takeuchi, T. Effect of interleukin-2 on synthesis of B cell activating factor belonging to the tumor necrosis factor family (BAFF) in human peripheral blood mononuclear cells. Cytokine 2008, 44, 44–48. [Google Scholar] [CrossRef]
- Krumbholz, M.; Theil, D.; Derfuss, T.; Rosenwald, A.; Schrader, F.; Monoranu, C.M.; Kalled, S.L.; Hess, D.M.; Serafini, B.; Aloisi, F.; et al. BAFF is produced by astrocytes and up-regulated in multiple sclerosis lesions and primary central nervous system lymphoma. J. Exp. Med. 2005, 201, 195–200. [Google Scholar] [CrossRef]
- Thangarajh, M.; Masterman, T.; Hillert, J.; Moerk, S.; Jonsson, R. A proliferation-inducing ligand (APRIL) is expressed by astrocytes and is increased in multiple sclerosis. Scand. J. Immunol. 2007, 65, 92–98. [Google Scholar] [CrossRef]
- Kato, A.; Truong-Tran, A.Q.; Scott, A.L.; Matsumoto, K.; Schleimer, R.P. Airway epithelial cells produce B cell-activating factor of TNF family by an IFN-beta-dependent mechanism. J. Immunol. 2006, 177, 7164–7172. [Google Scholar] [CrossRef] [PubMed]
- Huard, B.; McKee, T.; Bosshard, C.; Durual, S.; Matthes, T.; Myit, S.; Donze, O.; Frossard, C.; Chizzolini, C.; Favre, C.; et al. APRIL secreted by neutrophils binds to heparan sulfate proteoglycans to create plasma cell niches in human mucosa. J. Clin. Investig. 2008, 118, 2887–2895. [Google Scholar] [CrossRef]
- Ittah, M.; Miceli-Richard, C.; Gottenberg, J.E.; Sellam, J.; Eid, P.; Lebon, P.; Pallier, C.; Lepajolec, C.; Mariette, X. Viruses induce high expression of BAFF by salivary gland epithelial cells through TLR- and type-I IFN-dependent and -independent pathways. Eur. J. Immunol. 2008, 38, 1058–1064. [Google Scholar] [CrossRef] [PubMed]
- Abe, M.; Kido, S.; Hiasa, M.; Nakano, A.; Oda, A.; Amou, H.; Matsumoto, T. BAFF and APRIL as osteoclast-derived survival factors for myeloma cells: A rationale for TACI-Fc treatment in patients with multiple myeloma. Leukemia 2006, 20, 1313–1315. [Google Scholar] [CrossRef] [PubMed]
- Geffroy-Luseau, A.; Jégo, G.; Bataille, R.; Campion, L.; Pellat-Deceunynck, C. Osteoclasts support the survival of human plasma cells in vitro. Int. Immunol. 2008, 20, 775–782. [Google Scholar] [CrossRef] [PubMed]
- Langat, D.L.; Wheaton, D.A.; Platt, J.S.; Sifers, T.; Hunt, J.S. Signaling pathways for B cell-activating factor (BAFF) and a proliferation-inducing ligand (APRIL) in human placenta. Am. J. Pathol. 2008, 172, 1303–1311. [Google Scholar] [CrossRef] [PubMed]
- Alsaleh, G.; Messer, L.; Semaan, N.; Boulanger, N.; Gottenberg, J.E.; Sibilia, J.; Wachsmann, D. BAFF synthesis by rheumatoid synoviocytes is positively controlled by alpha5beta1 integrin stimulation and is negatively regulated by tumor necrosis factor alpha and Toll-like receptor ligands. Arthritis Rheum. 2007, 56, 3202–3214. [Google Scholar] [CrossRef]
- Pelekanou, V.; Kampa, M.; Kafousi, M.; Darivianaki, K.; Sanidas, E.; Tsiftsis, D.D.; Stathopoulos, E.N.; Tsapis, A.; Castanas, E. Expression of TNF-superfamily members BAFF and APRIL in breast cancer: Immunohistochemical study in 52 invasive ductal breast carcinomas. BMC Cancer 2008, 8, 76. [Google Scholar] [CrossRef]
- Darce, J.R.; Arendt, B.K.; Wu, X.; Jelinek, D.F. Regulated expression of BAFF-binding receptors during human B cell differentiation. J. Immunol. 2007, 179, 7276–7286. [Google Scholar] [CrossRef]
- Ng, L.G.; Sutherland, A.P.; Newton, R.; Qian, F.; Cachero, T.G.; Scott, M.L.; Thompson, J.S.; Wheway, J.; Chtanova, T.; Groom, J.; et al. B cell-activating factor belonging to the TNF family (BAFF)-R is the principal BAFF receptor facilitating BAFF costimulation of circulating T and B cells. J. Immunol. 2004, 173, 807–817. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, B.P.; Raman, V.S.; Erickson, L.D.; Cook, W.J.; Weaver, L.K.; Ahonen, C.; Lin, L.L.; Mantchev, G.T.; Bram, R.J.; Noelle, R.J. BCMA is essential for the survival of long-lived bone marrow plasma cells. J. Exp. Med. 2004, 199, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Chiu, A.; Xu, W.; He, B.; Dillon, S.R.; Gross, J.A.; Sievers, E.; Qiao, X.; Santini, P.; Hyjek, E.; Lee, J.W.; et al. Hodgkin lymphoma cells express TACI and BCMA receptors and generate survival and proliferation signals in response to BAFF and APRIL. Blood 2007, 109, 729–739. [Google Scholar] [CrossRef]
- Carpenter, R.O.; Evbuomwan, M.O.; Pittaluga, S.; Rose, J.J.; Raffeld, M.; Yang, S.; Gress, R.E.; Hakim, F.T.; Kochenderfer, J.N. B-cell Maturation Antigen Is a Promising Target for Adoptive T-cell Therapy of Multiple Myeloma. Clin. Cancer Res. 2013, 19, 2048–2060. [Google Scholar] [CrossRef] [PubMed]
- Groom, J.R.; Fletcher, C.A.; Walters, S.N.; Grey, S.T.; Watt, S.V.; Sweet, M.J.; Smyth, M.J.; Mackay, C.R.; Mackay, F. BAFF and MyD88 signals promote a lupuslike disease independent of T cells. J. Exp. Med. 2007, 204, 1959–1971. [Google Scholar] [CrossRef] [PubMed]
- Sindhava, V.J.; Scholz, J.L.; Stohl, W.; Cancro, M.P. APRIL mediates peritoneal B-1 cell homeostasis. Immunol. Lett. 2014, 160, 120–127. [Google Scholar] [CrossRef]
- Bloom, D.D.; Reshetylo, S.; Nytes, C.; Goodsett, C.T.; Hematti, P. Blockade of BAFF Receptor BR3 on T Cells Enhances Their Activation and Cytotoxicity. J. Immunother. 2018, 41, 213–223. [Google Scholar] [CrossRef]
- Marino, E.; Villanueva, J.; Walters, S.; Liuwantara, D.; Mackay, F.; Grey, S.T. CD4(+)CD25(+) T-cells control autoimmunity in the absence of B-cells. Diabetes 2009, 58, 1568–1577. [Google Scholar] [CrossRef]
- Walters, S.; Webster, K.E.; Sutherland, A.; Gardam, S.; Groom, J.; Liuwantara, D.; Marino, E.; Thaxton, J.; Weinberg, A.; Mackay, F.; et al. Increased CD4+Foxp3+ T cells in BAFF-transgenic mice suppress T cell effector responses. J. Immunol. 2009, 182, 793–801. [Google Scholar] [CrossRef]
- von Bülow, G.U.; Bram, R.J. NF-AT activation induced by a CAML-interacting member of the tumor necrosis factor receptor superfamily. Science 1997, 278, 138–141. [Google Scholar] [CrossRef]
- Chang, S.K.; Arendt, B.K.; Darce, J.R.; Wu, X.; Jelinek, D.F. A role for BLyS in the activation of innate immune cells. Blood 2006, 108, 2687–2694. [Google Scholar] [CrossRef]
- Chang, S.K.; Mihalcik, S.A.; Jelinek, D.F. B lymphocyte stimulator regulates adaptive immune responses by directly promoting dendritic cell maturation. J. Immunol. 2008, 180, 7394–7403. [Google Scholar] [CrossRef] [PubMed]
- Kampa, M.; Notas, G.; Stathopoulos, E.N.; Tsapis, A.; Castanas, E. The TNFSF Members APRIL and BAFF and Their Receptors TACI, BCMA, and BAFFR in Oncology, With a Special Focus in Breast Cancer. Front. Oncol. 2020, 10, 827. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Yang, M.; Long, D.; Li, Q.; Zhao, M.; Wu, H.; Lu, Q. Abnormal expression of BAFF and its receptors in peripheral blood and skin lesions from systemic lupus erythematosus patients. Autoimmunity 2020, 53, 192–200. [Google Scholar] [CrossRef] [PubMed]
- Schuh, E.; Musumeci, A.; Thaler, F.S.; Laurent, S.; Ellwart, J.W.; Hohlfeld, R.; Krug, A.; Meinl, E. Human Plasmacytoid Dendritic Cells Display and Shed B Cell Maturation Antigen upon TLR Engagement. J. Immunol. 2017, 198, 3081–3088. [Google Scholar] [CrossRef]
- Tai, Y.T.; Mayes, P.A.; Acharya, C.; Zhong, M.Y.; Cea, M.; Cagnetta, A.; Craigen, J.; Yates, J.; Gliddon, L.; Fieles, W.; et al. Novel anti-B-cell maturation antigen antibody-drug conjugate (GSK2857916) selectively induces killing of multiple myeloma. Blood 2014, 123, 3128–3138. [Google Scholar] [CrossRef]
- Garcillan, B.; Figgett, W.A.; Infantino, S.; Lim, E.X.; Mackay, F. Molecular control of B-cell homeostasis in health and malignancy. Immunol. Cell Biol. 2018, 96, 453–462. [Google Scholar] [CrossRef]
- Rowland, S.L.; Leahy, K.F.; Halverson, R.; Torres, R.M.; Pelanda, R. BAFF receptor signaling aids the differentiation of immature B cells into transitional B cells following tonic BCR signaling. J. Immunol. 2010, 185, 4570–4581. [Google Scholar] [CrossRef]
- Patke, A.; Mecklenbräuker, I.; Tarakhovsky, A. Survival signaling in resting B cells. Curr. Opin. Immunol. 2004, 16, 251–255. [Google Scholar] [CrossRef]
- Samy, E.; Wax, S.; Huard, B.; Hess, H.; Schneider, P. Targeting BAFF and APRIL in systemic lupus erythematosus and other antibody-associated diseases. Int. Rev. Immunol. 2017, 36, 3–19. [Google Scholar] [CrossRef]
- Muller-Winkler, J.; Mitter, R.; Rappe, J.C.F.; Vanes, L.; Schweighoffer, E.; Mohammadi, H.; Wack, A.; Tybulewicz, V.L.J. Critical requirement for BCR, BAFF, and BAFFR in memory B cell survival. J. Exp. Med. 2021, 218, e20191393. [Google Scholar] [CrossRef] [PubMed]
- Yan, M.; Brady, J.R.; Chan, B.; Lee, W.P.; Hsu, B.; Harless, S.; Cancro, M.; Grewal, I.S.; Dixit, V.M. Identification of a novel receptor for B lymphocyte stimulator that is mutated in a mouse strain with severe B cell deficiency. Curr. Biol. 2001, 11, 1547–1552. [Google Scholar] [CrossRef] [PubMed]
- Schiemann, B.; Gommerman, J.L.; Vora, K.; Cachero, T.G.; Shulga-Morskaya, S.; Dobles, M.; Frew, E.; Scott, M.L. An essential role for BAFF in the normal development of B cells through a BCMA-independent pathway. Science 2001, 293, 2111–2114. [Google Scholar] [CrossRef]
- Gross, J.A.; Dillon, S.R.; Mudri, S.; Johnston, J.; Littau, A.; Roque, R.; Rixon, M.; Schou, O.; Foley, K.P.; Haugen, H.; et al. TACI-Ig neutralizes molecules critical for B cell development and autoimmune disease. impaired B cell maturation in mice lacking BLyS. Immunity 2001, 15, 289–302. [Google Scholar] [CrossRef] [PubMed]
- Vora, K.A.; Wang, L.C.; Rao, S.P.; Liu, Z.Y.; Majeau, G.R.; Cutler, A.H.; Hochman, P.S.; Scott, M.L.; Kalled, S.L. Cutting edge: Germinal centers formed in the absence of B cell-activating factor belonging to the TNF family exhibit impaired maturation and function. J. Immunol. 2003, 171, 547–551. [Google Scholar] [CrossRef]
- Shulga-Morskaya, S.; Dobles, M.; Walsh, M.E.; Ng, L.G.; MacKay, F.; Rao, S.P.; Kalled, S.L.; Scott, M.L. B cell-activating factor belonging to the TNF family acts through separate receptors to support B cell survival and T cell-independent antibody formation. J. Immunol. 2004, 173, 2331–2341. [Google Scholar] [CrossRef]
- Gross, J.A.; Johnston, J.; Mudri, S.; Enselman, R.; Dillon, S.R.; Madden, K.; Xu, W.; Parrish-Novak, J.; Foster, D.; Lofton-Day, C.; et al. TACI and BCMA are receptors for a TNF homologue implicated in B-cell autoimmune disease. Nature 2000, 404, 995–999. [Google Scholar] [CrossRef]
- Mackay, F.; Woodcock, S.A.; Lawton, P.; Ambrose, C.; Baetscher, M.; Schneider, P.; Tschopp, J.; Browning, J.L. Mice transgenic for BAFF develop lymphocytic disorders along with autoimmune manifestations. J. Exp. Med. 1999, 190, 1697–1710. [Google Scholar] [CrossRef]
- Vincent, F.B.; Morand, E.F.; Schneider, P.; Mackay, F. The BAFF/APRIL system in SLE pathogenesis. Nat. Rev. Rheumatol. 2014, 10, 365–373. [Google Scholar] [CrossRef]
- Hardenberg, G.; van Bostelen, L.; Hahne, M.; Medema, J.P. Thymus-independent class switch recombination is affected by APRIL. Immunol. Cell Biol. 2008, 86, 530–534. [Google Scholar] [CrossRef]
- Belnoue, E.; Pihlgren, M.; McGaha, T.L.; Tougne, C.; Rochat, A.F.; Bossen, C.; Schneider, P.; Huard, B.; Lambert, P.H.; Siegrist, C.A. APRIL is critical for plasmablast survival in the bone marrow and poorly expressed by early-life bone marrow stromal cells. Blood 2008, 111, 2755–2764. [Google Scholar] [CrossRef]
- Benson, M.J.; Dillon, S.R.; Castigli, E.; Geha, R.S.; Xu, S.; Lam, K.P.; Noelle, R.J. Cutting edge: The dependence of plasma cells and independence of memory B cells on BAFF and APRIL. J. Immunol. 2008, 180, 3655–3659. [Google Scholar] [CrossRef] [PubMed]
- Reijmers, R.M.; Groen, R.W.; Kuil, A.; Weijer, K.; Kimberley, F.C.; Medema, J.P.; van Kuppevelt, T.H.; Li, J.P.; Spaargaren, M.; Pals, S.T. Disruption of heparan sulfate proteoglycan conformation perturbs B-cell maturation and APRIL-mediated plasma cell survival. Blood 2011, 117, 6162–6171. [Google Scholar] [CrossRef] [PubMed]
- Castigli, E.; Wilson, S.A.; Elkhal, A.; Ozcan, E.; Garibyan, L.; Geha, R.S. Transmembrane activator and calcium modulator and cyclophilin ligand interactor enhances CD40-driven plasma cell differentiation. J. Allergy Clin. Immunol. 2007, 120, 885–891. [Google Scholar] [CrossRef] [PubMed]
- Tsuji, S.; Cortesao, C.; Bram, R.J.; Platt, J.L.; Cascalho, M. TACI deficiency impairs sustained Blimp-1 expression in B cells decreasing long-lived plasma cells in the bone marrow. Blood 2011, 118, 5832–5839. [Google Scholar] [CrossRef]
- Seshasayee, D.; Valdez, P.; Yan, M.; Dixit, V.M.; Tumas, D.; Grewal, I.S. Loss of TACI causes fatal lymphoproliferation and autoimmunity, establishing TACI as an inhibitory BLyS receptor. Immunity 2003, 18, 279–288. [Google Scholar] [CrossRef]
- Yan, M.; Wang, H.; Chan, B.; Roose-Girma, M.; Erickson, S.; Baker, T.; Tumas, D.; Grewal, I.S.; Dixit, V.M. Activation and accumulation of B cells in TACI-deficient mice. Nat. Immunol. 2001, 2, 638–643. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.; Boone, T.; Delaney, J.; Hawkins, N.; Kelley, M.; Ramakrishnan, M.; McCabe, S.; Qiu, W.R.; Kornuc, M.; Xia, X.Z.; et al. APRIL and TALL-I and receptors BCMA and TACI: System for regulating humoral immunity. Nat. Immunol. 2000, 1, 252–256. [Google Scholar] [CrossRef]
- Patke, A.; Mecklenbräuker, I.; Erdjument-Bromage, H.; Tempst, P.; Tarakhovsky, A. BAFF controls B cell metabolic fitness through a PKCβ- and Akt-dependent mechanism. J. Exp. Med. 2006, 203, 2551–2562. [Google Scholar] [CrossRef]
- Caro-Maldonado, A.; Wang, R.; Nichols, A.G.; Kuraoka, M.; Milasta, S.; Sun, L.D.; Gavin, A.L.; Abel, E.D.; Kelsoe, G.; Green, D.R.; et al. Metabolic Reprogramming Is Required for Antibody Production That Is Suppressed in Anergic but Exaggerated in Chronically BAFF-Exposed B Cells. J. Immunol. 2014, 192, 3626–3636. [Google Scholar] [CrossRef]
- Park, S.; Jang, J.-W.; Moon, E.-Y. BAFF attenuates oxidative stress-induced cell death by the regulation of mitochondria membrane potential via Syk activation in WiL2-NS B lymphoblasts. Sci. Rep. 2020, 10, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Qin, J.; Ma, J.; Zeng, Q.; Zhang, H.; Zhang, R.; Liu, C.; Xu, C.; Zhang, S.; Huang, S. BAFF inhibits autophagy promoting cell proliferation and survival by activating Ca2+-Ca.aMKII-dependent Akt/mTOR signaling pathway in normal and neoplastic B-lymphoid cells. Cell. Signal. 2019, 53, 68–79. [Google Scholar] [CrossRef] [PubMed]
- Thien, M.; Phan, T.G.; Gardam, S.; Amesbury, M.; Basten, A.; Mackay, F.; Brink, R. Excess BAFF rescues self-reactive B cells from peripheral deletion and allows them to enter forbidden follicular and marginal zone niches. Immunity 2004, 20, 785–798. [Google Scholar] [CrossRef]
- Yang, M.; Sun, L.; Wang, S.; Ko, K.-H.; Xu, H.; Zheng, B.-J.; Cao, X.; Lu, L. Cutting Edge: Novel Function of B Cell-Activating Factor in the Induction of IL-10–Producing Regulatory B Cells. J. Immunol. 2010, 184, 3321–3325. [Google Scholar] [CrossRef]
- Hua, C.; Audo, R.; Yeremenko, N.; Baeten, D.; Hahne, M.; Combe, B.; Morel, J.; Daïen, C. A proliferation inducing ligand (APRIL) promotes IL-10 production and regulatory functions of human B cells. J. Autoimmun. 2016, 73, 64–72. [Google Scholar] [CrossRef]
- Fehres, C.M.; van Uden, N.O.; Yeremenko, N.G.; Fernandez, L.; Franco Salinas, G.; van Duivenvoorde, L.M.; Huard, B.; Morel, J.; Spits, H.; Hahne, M.; et al. APRIL Induces a Novel Subset of IgA+ Regulatory B Cells That Suppress Inflammation via Expression of IL-10 and PD-L1. Front. Immunol. 2019, 10, 1368. [Google Scholar] [CrossRef]
- Yanaba, K.; Bouaziz, J.D.; Haas, K.M.; Poe, J.C.; Fujimoto, M.; Tedder, T.F. A regulatory B cell subset with a unique CD1dhiCD5+ phenotype controls T cell-dependent inflammatory responses. Immunity 2008, 28, 639–650. [Google Scholar] [CrossRef]
- Saulep-Easton, D.; Vincent, F.B.; Quah, P.S.; Wei, A.; Ting, S.B.; Croce, C.M.; Tam, C.; Mackay, F. The BAFF receptor TACI controls IL-10 production by regulatory B cells and CLL B cells. Leukemia 2016, 30, 163–172. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, J.; Zhou, N.; Zhang, Y.; Wu, M.; Xu, J.; Shen, C.; An, X.; Shen, G.; Yang, M.; et al. The Unknown Aspect of BAFF: Inducing IL-35 Production by a CD5(+)CD1d(hi)FcγRIIb(hi) Regulatory B-Cell Subset in Lupus. J. Investig. Derm. 2017, 137, 2532–2543. [Google Scholar] [CrossRef] [PubMed]
- Do, R.K.; Hatada, E.; Lee, H.; Tourigny, M.R.; Hilbert, D.; Chen-Kiang, S. Attenuation of apoptosis underlies B lymphocyte stimulator enhancement of humoral immune response. J. Exp. Med. 2000, 192, 953–964. [Google Scholar] [CrossRef]
- Avery, D.T.; Kalled, S.L.; Ellyard, J.I.; Ambrose, C.; Bixler, S.A.; Thien, M.; Brink, R.; Mackay, F.; Hodgkin, P.D.; Tangye, S.G. BAFF selectively enhances the survival of plasmablasts generated from human memory B cells. J. Clin. Investig. 2003, 112, 286–297. [Google Scholar] [CrossRef]
- Castigli, E.; Wilson, S.A.; Scott, S.; Dedeoglu, F.; Xu, S.; Lam, K.P.; Bram, R.J.; Jabara, H.; Geha, R.S. TACI and BAFF-R mediate isotype switching in B cells. J. Exp. Med. 2005, 201, 35–39. [Google Scholar] [CrossRef] [PubMed]
- Sakurai, D.; Hase, H.; Kanno, Y.; Kojima, H.; Okumura, K.; Kobata, T. TACI regulates IgA production by APRIL in collaboration with HSPG. Blood 2007, 109, 2961–2967. [Google Scholar] [CrossRef] [PubMed]
- Figgett, W.A.; Fairfax, K.; Vincent, F.B.; Le Page, M.A.; Katik, I.; Deliyanti, D.; Quah, P.S.; Verma, P.; Grumont, R.; Gerondakis, S.; et al. The TACI receptor regulates T-cell-independent marginal zone B cell responses through innate activation-induced cell death. Immunity 2013, 39, 573–583. [Google Scholar] [CrossRef]
- Xu, S.; Lam, K.P. B-cell maturation protein, which binds the tumor necrosis factor family members BAFF and APRIL, is dispensable for humoral immune responses. Mol. Cell Biol. 2001, 21, 4067–4074. [Google Scholar] [CrossRef]
- von Bulow, G.U.; van Deursen, J.M.; Bram, R.J. Regulation of the T-independent humoral response by TACI. Immunity 2001, 14, 573–582. [Google Scholar] [CrossRef] [PubMed]
- Mantchev, G.T.; Cortesao, C.S.; Rebrovich, M.; Cascalho, M.; Bram, R.J. TACI is required for efficient plasma cell differentiation in response to T-independent type 2 antigens. J. Immunol. 2007, 179, 2282–2288. [Google Scholar] [CrossRef]
- Salzer, U.; Chapel, H.M.; Webster, A.D.; Pan-Hammarstrom, Q.; Schmitt-Graeff, A.; Schlesier, M.; Peter, H.H.; Rockstroh, J.K.; Schneider, P.; Schaffer, A.A.; et al. Mutations in TNFRSF13B encoding TACI are associated with common variable immunodeficiency in humans. Nat. Genet. 2005, 37, 820–828. [Google Scholar] [CrossRef]
- Castigli, E.; Wilson, S.A.; Garibyan, L.; Rachid, R.; Bonilla, F.; Schneider, L.; Geha, R.S. TACI is mutant in common variable immunodeficiency and IgA deficiency. Nat. Genet. 2005, 37, 829–834. [Google Scholar] [CrossRef]
- Salzer, U.; Grimbacher, B. TACI deficiency—A complex system out of balance. Curr. Opin. Immunol. 2021, 71, 81–88. [Google Scholar] [CrossRef]
- Huard, B.; Schneider, P.; Mauri, D.; Tschopp, J.; French, L.E. T cell costimulation by the TNF ligand BAFF. J. Immunol. 2001, 167, 6225–6231. [Google Scholar] [CrossRef]
- Hu, S.; Wang, R.; Zhang, M.; Liu, K.; Tao, J.; Tai, Y.; Zhou, W.; Wang, Q.; Wei, W. BAFF promotes T cell activation through the BAFF-BAFF-R-PI3K-Akt signaling pathway. Biomed. Pharmacother. 2019, 114, 108796. [Google Scholar] [CrossRef] [PubMed]
- Allman, W.R.; Dey, R.; Liu, L.; Siddiqui, S.; Coleman, A.S.; Bhattacharya, P.; Yano, M.; Uslu, K.; Takeda, K.; Nakhasi, H.L.; et al. TACI deficiency leads to alternatively activated macrophage phenotype and susceptibility to Leishmania infection. Proc. Natl. Acad. Sci. USA 2015, 112, E4094–E4103. [Google Scholar] [CrossRef] [PubMed]
- Quah, P.S.; Sutton, V.; Whitlock, E.; Figgett, W.A.; Andrews, D.M.; Fairfax, K.A.; Mackay, F. The effects of B cell-activating factor (BAFF) on the population size, maturation, and function of murine natural killer cells. Immunol. Cell Biol. 2022, 100, 761–776. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Wen, L.; Huang, X.; Liang, J.; Gao, W.; Zhang, S.; Chen, L. hsBAFF enhances activity of NK cells by regulation of CD4(+) T lymphocyte function. Immunol. Lett. 2008, 120, 96–102. [Google Scholar] [CrossRef]
- Bonci, D.; Hahne, M.; Felli, N.; Peschle, C.; De Maria, R. Potential role of APRIL as autocrine growth factor for megakaryocytopoiesis. Blood 2004, 104, 3169–3172. [Google Scholar] [CrossRef] [PubMed]
- Levy, R.A.; Gonzalez-Rivera, T.; Khamashta, M.; Fox, N.L.; Jones-Leone, A.; Rubin, B.; Burriss, S.W.; Gairy, K.; Maurik, A.V.; Roth, D.A. 10 Years of belimumab experience: What have we learnt? Lupus 2021, 30, 1705–1721. [Google Scholar] [CrossRef]
- Jackson, S.W.; Scharping, N.E.; Jacobs, H.M.; Wang, S.; Chait, A.; Rawlings, D.J. Cutting Edge: BAFF Overexpression Reduces Atherosclerosis via TACI-Dependent B Cell Activation. J. Immunol. 2016, 197, 4529–4534. [Google Scholar] [CrossRef]
- Tsiantoulas, D.; Sage, A.P.; Göderle, L.; Ozsvar-Kozma, M.; Murphy, D.; Porsch, F.; Pasterkamp, G.; Menche, J.; Schneider, P.; Mallat, Z.; et al. B Cell-Activating Factor Neutralization Aggravates Atherosclerosis. Circulation 2018, 138, 2263–2273. [Google Scholar] [CrossRef]
- Tsiantoulas, D.; Eslami, M.; Obermayer, G.; Clement, M.; Smeets, D.; Mayer, F.J.; Kiss, M.G.; Enders, L.; Weißer, J.; Göderle, L.; et al. APRIL limits atherosclerosis by binding to heparan sulfate proteoglycans. Nature 2021, 597, 92–96. [Google Scholar] [CrossRef]
- Varin, M.M.; Le Pottier, L.; Youinou, P.; Saulep, D.; Mackay, F.; Pers, J.O. B-cell tolerance breakdown in Sjogren’s syndrome: Focus on BAFF. Autoimmun. Rev. 2010, 9, 604–608. [Google Scholar] [CrossRef] [PubMed]
- Dillon, S.R.; Harder, B.; Lewis, K.B.; Moore, M.D.; Liu, H.; Bukowski, T.R.; Hamacher, N.B.; Lantry, M.M.; Maurer, M.; Krejsa, C.M.; et al. B-lymphocyte stimulator/a proliferation-inducing ligand heterotrimers are elevated in the sera of patients with autoimmune disease and are neutralized by atacicept and B-cell maturation antigen-immunoglobulin. Arthritis Res. 2010, 12, R48. [Google Scholar] [CrossRef] [PubMed]
- Lai Kwan Lam, Q.; King Hung Ko, O.; Zheng, B.J.; Lu, L. Local BAFF gene silencing suppresses Th17-cell generation and ameliorates autoimmune arthritis. Proc. Natl. Acad. Sci. USA 2008, 105, 14993–14998. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Guo, J.; Wang, L.; Zhou, W.; Zhang, Z. The role of a proliferation-inducing ligand (APRIL) in the pathogenesis of rheumatoid arthritis. Scand. J. Rheumatol 2014, 43, 462–469. [Google Scholar] [CrossRef] [PubMed]
- Kappos, L.; Hartung, H.-P.; Freedman, M.S.; Boyko, A.; Radü, E.W.; Mikol, D.D.; Lamarine, M.; Hyvert, Y.; Freudensprung, U.; Plitz, T. Atacicept in multiple sclerosis (ATAMS): A randomised, placebo-controlled, double-blind, phase 2 trial. Lancet Neurol. 2014, 13, 353–363. [Google Scholar] [CrossRef]
- Schubert, R.D.; Hu, Y.; Kumar, G.; Szeto, S.; Abraham, P.; Winderl, J.; Guthridge, J.M.; Pardo, G.; Dunn, J.; Steinman, L.; et al. IFN-β treatment requires B cells for efficacy in neuroautoimmunity. J. Immunol. 2015, 194, 2110–2116. [Google Scholar] [CrossRef]
- Rizzo, F.; Giacomini, E.; Mechelli, R.; Buscarinu, M.C.; Salvetti, M.; Severa, M.; Coccia, E.M. Interferon-β therapy specifically reduces pathogenic memory B cells in multiple sclerosis patients by inducing a FAS-mediated apoptosis. Immunol. Cell Biol. 2016, 94, 886–894. [Google Scholar] [CrossRef]
- Rojas, O.L.; Probstel, A.K.; Porfilio, E.A.; Wang, A.A.; Charabati, M.; Sun, T.; Lee, D.S.W.; Galicia, G.; Ramaglia, V.; Ward, L.A.; et al. Recirculating Intestinal IgA-Producing Cells Regulate Neuroinflammation via IL-10. Cell 2019, 176, 610–624.e618. [Google Scholar] [CrossRef]
- McNamara, P.S.; Fonceca, A.M.; Howarth, D.; Correia, J.B.; Slupsky, J.R.; Trinick, R.E.; Al Turaiki, W.; Smyth, R.L.; Flanagan, B.F. Respiratory syncytial virus infection of airway epithelial cells, in vivo and in vitro, supports pulmonary antibody responses by inducing expression of the B cell differentiation factor BAFF. Thorax 2013, 68, 76. [Google Scholar] [CrossRef]
- Toubi, E.; Gordon, S.; Kessel, A.; Rosner, I.; Rozenbaum, M.; Shoenfeld, Y.; Zuckerman, E. Elevated serum B-Lymphocyte activating factor (BAFF) in chronic hepatitis C virus infection: Association with autoimmunity. J. Autoimmun. 2006, 27, 134–139. [Google Scholar] [CrossRef]
- Rodriguez, B.; Valdez, H.; Freimuth, W.; Butler, T.; Asaad, R.; Lederman, M.M. Plasma levels of B-lymphocyte stimulator increase with HIV disease progression. Aids 2003, 17, 1983–1985. [Google Scholar] [CrossRef] [PubMed]
- Adalid-Peralta, L.; Godot, V.; Colin, C.; Krzysiek, R.; Tran, T.; Poignard, P.; Venet, A.; Hosmalin, A.; Lebon, P.; Rouzioux, C.; et al. Stimulation of the primary anti-HIV antibody response by IFN-α in patients with acute HIV-1 infection. J. Leukoc. Biol. 2008, 83, 1060–1067. [Google Scholar] [CrossRef]
- Wolf, A.I.; Mozdzanowska, K.; Quinn, W.J., 3rd; Metzgar, M.; Williams, K.L.; Caton, A.J.; Meffre, E.; Bram, R.J.; Erickson, L.D.; Allman, D.; et al. Protective antiviral antibody responses in a mouse model of influenza virus infection require TACI. J. Clin. Investig. 2011, 121, 3954–3964. [Google Scholar] [CrossRef]
- Jabłońska, E.; Dakowicz, L.; Ratajczak-Wrona, W.; Garley, M.; Sawicka-Powierza, J.; Krawczuk-Rybak, M. TNF superfamily proteins in the serum of patients with B-ALL--preliminary study. Clin. Lab. 2014, 60, 1757–1764. [Google Scholar] [CrossRef]
- Onda, K.; Iijima, K.; Katagiri, Y.U.; Okita, H.; Saito, M.; Shimizu, T.; Kiyokawa, N. Differential effects of BAFF on B cell precursor acute lymphoblastic leukemia and Burkitt lymphoma. Int. J. Hematol. 2010, 91, 808–819. [Google Scholar] [CrossRef]
- Parameswaran, R.; Müschen, M.; Kim, Y.-m.; Groffen, J.; Heisterkamp, N. A Functional Receptor for B-Cell–Activating Factor Is Expressed on Human Acute Lymphoblastic Leukemias. Cancer Res. 2010, 70, 4346–4356. [Google Scholar] [CrossRef]
- Oki, Y.; Georgakis, G.V.; Migone, T.-S.; Kwak, L.W.; Younes, A. Serum BLyS Level as a Prognostic Marker in Patients with Lymphoma. Blood 2005, 106, 1926. [Google Scholar] [CrossRef]
- Moreaux, J.; Legouffe, E.; Jourdan, E.; Quittet, P.; Rème, T.; Lugagne, C.; Moine, P.; Rossi, J.F.; Klein, B.; Tarte, K. BAFF and APRIL protect myeloma cells from apoptosis induced by interleukin 6 deprivation and dexamethasone. Blood 2004, 103, 3148–3157. [Google Scholar] [CrossRef]
- Novak, A.J.; Darce, J.R.; Arendt, B.K.; Harder, B.; Henderson, K.; Kindsvogel, W.; Gross, J.A.; Greipp, P.R.; Jelinek, D.F. Expression of BCMA, TACI, and BAFF-R in multiple myeloma: A mechanism for growth and survival. Blood 2004, 103, 689–694. [Google Scholar] [CrossRef] [PubMed]
- Rihacek, M.; Bienertova-Vasku, J.; Valik, D.; Sterba, J.; Pilatova, K.; Zdrazilova-Dubska, L. B-Cell Activating Factor as a Cancer Biomarker and Its Implications in Cancer-Related Cachexia. BioMed Res. Int. 2015, 2015, 792187. [Google Scholar] [CrossRef] [PubMed]
- Fragioudaki, M.; Tsirakis, G.; Pappa, C.A.; Aristeidou, I.; Tsioutis, C.; Alegakis, A.; Kyriakou, D.S.; Stathopoulos, E.N.; Alexandrakis, M.G. Serum BAFF levels are related to angiogenesis and prognosis in patients with multiple myeloma. Leuk. Res. 2012, 36, 1004–1008. [Google Scholar] [CrossRef]
- Lemancewicz, D.; Bolkun, L.; Jablonska, E.; Kulczynska, A.; Bolkun-Skornicka, U.; Kloczko, J.; Dzieciol, J. Evaluation of TNF superfamily molecules in multiple myeloma patients: Correlation with biological and clinical features. Leuk. Res. 2013, 37, 1089–1093. [Google Scholar] [CrossRef]
- Sanchez, E.; Li, M.; Kitto, A.; Li, J.; Wang, C.S.; Kirk, D.T.; Yellin, O.; Nichols, C.M.; Dreyer, M.P.; Ahles, C.P.; et al. Serum B-cell maturation antigen is elevated in multiple myeloma and correlates with disease status and survival. Br. J. Haematol. 2012, 158, 727–738. [Google Scholar] [CrossRef] [PubMed]
- De Vos, J.; Thykjaer, T.; Tarte, K.; Ensslen, M.; Raynaud, P.; Requirand, G.; Pellet, F.; Pantesco, V.; Rème, T.; Jourdan, M.; et al. Comparison of gene expression profiling between malignant and normal plasma cells with oligonucleotide arrays. Oncogene 2002, 21, 6848–6857. [Google Scholar] [CrossRef]
- Ghermezi, M.; Li, M.; Vardanyan, S.; Harutyunyan, N.M.; Gottlieb, J.; Berenson, A.; Spektor, T.M.; Andreu-Vieyra, C.; Petraki, S.; Sanchez, E.; et al. Serum B-cell maturation antigen: A novel biomarker to predict outcomes for multiple myeloma patients. Haematologica 2017, 102, 785–795. [Google Scholar] [CrossRef]
- Tarte, K.; De Vos, J.; Thykjaer, T.; Zhan, F.; Fiol, G.; Costes, V.; Rème, T.; Legouffe, E.; Rossi, J.F.; Shaughnessy, J., Jr.; et al. Generation of polyclonal plasmablasts from peripheral blood B cells: A normal counterpart of malignant plasmablasts. Blood 2002, 100, 1113–1122. [Google Scholar] [CrossRef] [PubMed]
- Moreaux, J.; Cremer, F.W.; Reme, T.; Raab, M.; Mahtouk, K.; Kaukel, P.; Pantesco, V.; De Vos, J.; Jourdan, E.; Jauch, A.; et al. The level of TACI gene expression in myeloma cells is associated with a signature of microenvironment dependence versus a plasmablastic signature. Blood 2005, 106, 1021–1030. [Google Scholar] [CrossRef]
- Bojarska-Junak, A.; Hus, I.; Chocholska, S.; Wasik-Szczepanek, E.; Sieklucka, M.; Dmoszyńska, A.; Roliński, J. BAFF and APRIL expression in B-cell chronic lymphocytic leukemia: Correlation with biological and clinical features. Leuk. Res. 2009, 33, 1319–1327. [Google Scholar] [CrossRef]
- Ferrer, G.; Hodgson, K.; Pereira, A.; Juan, M.; Elena, M.; Colomer, D.; Roue, G.; Aymerich, M.; Baumann, T.; Montserrat, E.; et al. Combined analysis of levels of serum B-cell activating factor and a proliferation-inducing ligand as predictor of disease progression in patients with chronic lymphocytic leukemia. Leuk. Lymphoma 2011, 52, 2064–2068. [Google Scholar] [CrossRef]
- Endo, T.; Nishio, M.; Enzler, T.; Cottam, H.B.; Fukuda, T.; James, D.F.; Karin, M.; Kipps, T.J. BAFF and APRIL support chronic lymphocytic leukemia B-cell survival through activation of the canonical NF-kappaB pathway. Blood 2007, 109, 703–710. [Google Scholar] [CrossRef] [PubMed]
- Kern, C.; Cornuel, J.F.; Billard, C.; Tang, R.; Rouillard, D.; Stenou, V.; Defrance, T.; Ajchenbaum-Cymbalista, F.; Simonin, P.Y.; Feldblum, S.; et al. Involvement of BAFF and APRIL in the resistance to apoptosis of B-CLL through an autocrine pathway. Blood 2004, 103, 679–688. [Google Scholar] [CrossRef] [PubMed]
- Cols, M.; Barra, C.M.; He, B.; Puga, I.; Xu, W.; Chiu, A.; Tam, W.; Knowles, D.M.; Dillon, S.R.; Leonard, J.P.; et al. Stromal endothelial cells establish a bidirectional crosstalk with chronic lymphocytic leukemia cells through the TNF-related factors BAFF, APRIL, and CD40L. J. Immunol. 2012, 188, 6071–6083. [Google Scholar] [CrossRef] [PubMed]
- Briones, J.; Timmerman, J.M.; Hilbert, D.M.; Levy, R. BLyS and BLyS receptor expression in non-Hodgkin’s lymphoma. Exp. Hematol. 2002, 30, 135–141. [Google Scholar] [CrossRef]
- Udd, K.A.; Bujarski, S.; Wirtschafter, E.; Spektor, T.M.; Ghermezi, M.; Rassenti, L.Z.; David, M.E.; Nosrati, J.D.; Rahbari, A.A.; Wang, J.; et al. Plasma B-Cell Maturation Antigen Levels are Elevated and Correlate with Disease Activity in Patients with Chronic Lymphocytic Leukemia. Target. Oncol. 2019, 14, 551–561. [Google Scholar] [CrossRef] [PubMed]
- Soof, C.M.; Spektor, T.M.; Parikh, S.A.; Slager, S.L.; Rabe, K.G.; Call, T.G.; Kenderian, S.S.; Ding, W.; Muchtar, E.; Ghermezi, M.; et al. Serum B-Cell maturation antigen is an independent prognostic marker in previously untreated chronic lymphocytic leukemia. Exp. Hematol. 2022, 111, 32–40. [Google Scholar] [CrossRef]
- Mulazzani, M.; Huber, M.; Borchard, S.; Langer, S.; Angele, B.; Schuh, E.; Meinl, E.; Dreyling, M.; Birnbaum, T.; Straube, A.; et al. APRIL and BAFF: Novel biomarkers for central nervous system lymphoma. J. Hematol. Oncol. 2019, 12, 102. [Google Scholar] [CrossRef] [PubMed]
- Mizutani, H.; Nakane, S.; Ikeda, T.; Nakamura, H.; Takamatsu, K.; Makino, K.; Tawara, N.; Mukaino, A.; Watari, M.; Matsui, H.; et al. CSF TACI and BAFF levels in patients with primary CNS lymphoma as novel diagnostic biomarkers. Ann. Clin. Transl. Neurol. 2018, 5, 1611–1616. [Google Scholar] [CrossRef]
- Thaler, F.S.; Laurent, S.A.; Huber, M.; Mulazzani, M.; Dreyling, M.; Kodel, U.; Kumpfel, T.; Straube, A.; Meinl, E.; von Baumgarten, L. Soluble TACI and soluble BCMA as biomarkers in primary central nervous system lymphoma. Neuro. Oncol. 2017, 19, 1618–1627. [Google Scholar] [CrossRef]
- Schwaller, J.; Schneider, P.; Mhawech-Fauceglia, P.; McKee, T.; Myit, S.; Matthes, T.; Tschopp, J.; Donze, O.; Le Gal, F.A.; Huard, B. Neutrophil-derived APRIL concentrated in tumor lesions by proteoglycans correlates with human B-cell lymphoma aggressiveness. Blood 2007, 109, 331–338. [Google Scholar] [CrossRef]
- Novak, A.J.; Slager, S.L.; Fredericksen, Z.S.; Wang, A.H.; Manske, M.M.; Ziesmer, S.; Liebow, M.; Macon, W.R.; Dillon, S.R.; Witzig, T.E.; et al. Genetic variation in B-cell-activating factor is associated with an increased risk of developing B-cell non-Hodgkin lymphoma. Cancer Res. 2009, 69, 4217–4224. [Google Scholar] [CrossRef]
- Rodig, S.J.; Shahsafaei, A.; Li, B.; Mackay, C.R.; Dorfman, D.M. BAFF-R, the major B cell-activating factor receptor, is expressed on most mature B cells and B-cell lymphoproliferative disorders. Hum. Pathol. 2005, 36, 1113–1119. [Google Scholar] [CrossRef]
- Novak, A.J.; Grote, D.M.; Stenson, M.; Ziesmer, S.C.; Witzig, T.E.; Habermann, T.M.; Harder, B.; Ristow, K.M.; Bram, R.J.; Jelinek, D.F.; et al. Expression of BLyS and its receptors in B-cell non-Hodgkin lymphoma: Correlation with disease activity and patient outcome. Blood 2004, 104, 2247–2253. [Google Scholar] [CrossRef] [PubMed]
- Linabery, A.M.; Ross, J.A. Trends in childhood cancer incidence in the U.S. (1992–2004). Cancer 2008, 112, 416–432. [Google Scholar] [CrossRef]
- Rafei, H.; Kantarjian, H.M.; Jabbour, E.J. Recent advances in the treatment of acute lymphoblastic leukemia. Leuk. Lymphoma 2019, 60, 2606–2621. [Google Scholar] [CrossRef] [PubMed]
- Sevdali, E.; Katsantoni, E.; Smulski, C.R.; Moschovi, M.; Palassopoulou, M.; Kolokotsa, E.N.; Argentou, N.; Giannakoulas, N.; Adamaki, M.; Vassilopoulos, G.; et al. BAFF/APRIL System Is Functional in B-Cell Acute Lymphoblastic Leukemia in a Disease Subtype Manner. Front. Oncol. 2019, 9, 594. [Google Scholar] [CrossRef]
- Li, Y.; Xie, X.; Jie, Z.; Zhu, L.; Yang, J.Y.; Ko, C.J.; Gao, T.; Jain, A.; Jung, S.Y.; Baran, N.; et al. DYRK1a mediates BAFF-induced noncanonical NF-kB activation to promote autoimmunity and B cell leukemogenesis. Blood 2021, 138, 2360–2371. [Google Scholar] [CrossRef]
- Fazio, G.; Turazzi, N.; Cazzaniga, V.; Kreuzaler, M.; Maglia, O.; Magnani, C.F.; Biagi, E.; Rolink, A.; Biondi, A.; Cazzaniga, G. TNFRSF13C (BAFFR) positive blasts persist after early treatment and at relapse in childhood B-cell precursor acute lymphoblastic leukaemia. Br. J. Haematol. 2018, 182, 434–436. [Google Scholar] [CrossRef]
- Küppers, R.; Rajewsky, K. The origin of Hodgkin and Reed/Sternberg cells in Hodgkin’s disease. Annu. Rev. Immunol. 1998, 16, 471–493. [Google Scholar] [CrossRef] [PubMed]
- Marafioti, T.; Hummel, M.; Foss, H.D.; Laumen, H.; Korbjuhn, P.; Anagnostopoulos, I.; Lammert, H.; Demel, G.; Theil, J.; Wirth, T.; et al. Hodgkin and reed-sternberg cells represent an expansion of a single clone originating from a germinal center B-cell with functional immunoglobulin gene rearrangements but defective immunoglobulin transcription. Blood 2000, 95, 1443–1450. [Google Scholar] [CrossRef]
- Mayani, H. Hematopoietic and microenvironment alterations in bone marrow from patients with multiple myeloma. Leuk. Res. 2013, 37, 228–229. [Google Scholar] [CrossRef]
- van Nieuwenhuijzen, N.; Spaan, I.; Raymakers, R.; Peperzak, V. From MGUS to Multiple Myeloma, a Paradigm for Clonal Evolution of Premalignant Cells. Cancer Res. 2018, 78, 2449–2456. [Google Scholar] [CrossRef]
- Matthes, T.; Dunand-Sauthier, I.; Santiago-Raber, M.L.; Krause, K.H.; Donze, O.; Passweg, J.; McKee, T.; Huard, B. Production of the plasma-cell survival factor a proliferation-inducing ligand (APRIL) peaks in myeloid precursor cells from human bone marrow. Blood 2011, 118, 1838–1844. [Google Scholar] [CrossRef]
- Tai, Y.T.; Acharya, C.; An, G.; Moschetta, M.; Zhong, M.Y.; Feng, X.; Cea, M.; Cagnetta, A.; Wen, K.; van Eenennaam, H.; et al. APRIL and BCMA promote human multiple myeloma growth and immunosuppression in the bone marrow microenvironment. Blood 2016, 127, 3225–3236. [Google Scholar] [CrossRef]
- Shen, X.; Guo, Y.; Qi, J.; Shi, W.; Wu, X.; Ju, S. Binding of B-cell maturation antigen to B-cell activating factor induces survival of multiple myeloma cells by activating Akt and JNK signaling pathways. Cell Biochem. Funct. 2016, 34, 104–110. [Google Scholar] [CrossRef]
- Tai, Y.T.; Li, X.F.; Breitkreutz, I.; Song, W.; Neri, P.; Catley, L.; Podar, K.; Hideshima, T.; Chauhan, D.; Raje, N.; et al. Role of B-cell-activating factor in adhesion and growth of human multiple myeloma cells in the bone marrow microenvironment. Cancer Res. 2006, 66, 6675–6682. [Google Scholar] [CrossRef]
- Tai, Y.T.; Lin, L.; Xing, L.; Cho, S.F.; Yu, T.; Acharya, C.; Wen, K.; Hsieh, P.A.; Dulos, J.; van Elsas, A.; et al. APRIL signaling via TACI mediates immunosuppression by T regulatory cells in multiple myeloma: Therapeutic implications. Leukemia 2019, 33, 426–438. [Google Scholar] [CrossRef]
- Küppers, R.; Klein, U.; Hansmann, M.L.; Rajewsky, K. Cellular origin of human B-cell lymphomas. N. Engl. J. Med. 1999, 341, 1520–1529. [Google Scholar] [CrossRef] [PubMed]
- Ogden, C.A.; Pound, J.D.; Batth, B.K.; Owens, S.; Johannessen, I.; Wood, K.; Gregory, C.D. Enhanced apoptotic cell clearance capacity and B cell survival factor production by IL-10-activated macrophages: Implications for Burkitt’s lymphoma. J. Immunol. 2005, 174, 3015–3023. [Google Scholar] [CrossRef] [PubMed]
- Hallek, M.; Shanafelt, T.D.; Eichhorst, B. Chronic lymphocytic leukaemia. Lancet 2018, 391, 1524–1537. [Google Scholar] [CrossRef] [PubMed]
- Landau, D.A.; Tausch, E.; Taylor-Weiner, A.N.; Stewart, C.; Reiter, J.G.; Bahlo, J.; Kluth, S.; Bozic, I.; Lawrence, M.; Böttcher, S.; et al. Mutations driving CLL and their evolution in progression and relapse. Nature 2015, 526, 525–530. [Google Scholar] [CrossRef]
- Lin, K.I.; Tam, C.S.; Keating, M.J.; Wierda, W.G.; O’Brien, S.; Lerner, S.; Coombes, K.R.; Schlette, E.; Ferrajoli, A.; Barron, L.L.; et al. Relevance of the immunoglobulin VH somatic mutation status in patients with chronic lymphocytic leukemia treated with fludarabine, cyclophosphamide, and rituximab (FCR) or related chemoimmunotherapy regimens. Blood 2009, 113, 3168–3171. [Google Scholar] [CrossRef]
- Eichhorst, B.; Fink, A.M.; Bahlo, J.; Busch, R.; Kovacs, G.; Maurer, C.; Lange, E.; Köppler, H.; Kiehl, M.; Sökler, M.; et al. First-line chemoimmunotherapy with bendamustine and rituximab versus fludarabine, cyclophosphamide, and rituximab in patients with advanced chronic lymphocytic leukaemia (CLL10): An international, open-label, randomised, phase 3, non-inferiority trial. Lancet Oncol. 2016, 17, 928–942. [Google Scholar] [CrossRef] [PubMed]
- Decker, T.; Hipp, S.; Ringshausen, I.; Bogner, C.; Oelsner, M.; Schneller, F.; Peschel, C. Rapamycin-induced G1 arrest in cycling B-CLL cells is associated with reduced expression of cyclin D3, cyclin E, cyclin A, and survivin. Blood 2003, 101, 278–285. [Google Scholar] [CrossRef]
- Raval, A.; Tanner, S.M.; Byrd, J.C.; Angerman, E.B.; Perko, J.D.; Chen, S.S.; Hackanson, B.; Grever, M.R.; Lucas, D.M.; Matkovic, J.J.; et al. Downregulation of death-associated protein kinase 1 (DAPK1) in chronic lymphocytic leukemia. Cell 2007, 129, 879–890. [Google Scholar] [CrossRef]
- Vaca, A.M.; Ioannou, N.; Sivina, M.; Vlachonikola, E.; Clise-Dwyer, K.; Kim, E.; Li, D.; Ma, Q.; Ferrajoli, A.; Estrov, Z.; et al. Activation and expansion of T-follicular helper cells in chronic lymphocytic leukemia nurselike cell co-cultures. Leukemia 2022, 36, 1324–1335. [Google Scholar] [CrossRef]
- Nishio, M.; Endo, T.; Tsukada, N.; Ohata, J.; Kitada, S.; Reed, J.C.; Zvaifler, N.J.; Kipps, T.J. Nurselike cells express BAFF and APRIL, which can promote survival of chronic lymphocytic leukemia cells via a paracrine pathway distinct from that of SDF-1alpha. Blood 2005, 106, 1012–1020. [Google Scholar] [CrossRef]
- Bichi, R.; Shinton, S.A.; Martin, E.S.; Koval, A.; Calin, G.A.; Cesari, R.; Russo, G.; Hardy, R.R.; Croce, C.M. Human chronic lymphocytic leukemia modeled in mouse by targeted TCL1 expression. Proc. Natl. Acad. Sci. USA 2002, 99, 6955–6960. [Google Scholar] [CrossRef]
- Saulep-Easton, D.; Vincent, F.B.; Le Page, M.; Wei, A.; Ting, S.B.; Croce, C.M.; Tam, C.; Mackay, F. Cytokine-driven loss of plasmacytoid dendritic cell function in chronic lymphocytic leukemia. Leukemia 2014, 28, 2005–2015. [Google Scholar] [CrossRef]
- Planelles, L.; Carvalho-Pinto, C.E.; Hardenberg, G.; Smaniotto, S.; Savino, W.; Gomez-Caro, R.; Alvarez-Mon, M.; de Jong, J.; Eldering, E.; Martinez, A.C.; et al. APRIL promotes B-1 cell-associated neoplasm. Cancer Cell 2004, 6, 399–408. [Google Scholar] [CrossRef] [PubMed]
- Lascano, V.; Guadagnoli, M.; Schot, J.G.; Luijks, D.M.; Guikema, J.E.; Cameron, K.; Hahne, M.; Pals, S.; Slinger, E.; Kipps, T.J.; et al. Chronic lymphocytic leukemia disease progression is accelerated by APRIL-TACI interaction in the TCL1 transgenic mouse model. Blood 2013, 122, 3960–3963. [Google Scholar] [CrossRef] [PubMed]
- Deckert, M.; Brunn, A.; Montesinos-Rongen, M.; Terreni, M.R.; Ponzoni, M. Primary lymphoma of the central nervous system—A diagnostic challenge. Hematol. Oncol. 2014, 32, 57–67. [Google Scholar] [CrossRef]
- Zhou, X.; Mulazzani, M.; von Mücke-Heim, I.A.; Langer, S.; Zhang, W.; Ishikawa-Ankerhold, H.; Dreyling, M.; Straube, A.; von Baumgarten, L. The Role of BAFF-R Signaling in the Growth of Primary Central Nervous System Lymphoma. Front. Oncol. 2020, 10, 682. [Google Scholar] [CrossRef] [PubMed]
- Pham, L.V.; Fu, L.; Tamayo, A.T.; Bueso-Ramos, C.; Drakos, E.; Vega, F.; Medeiros, L.J.; Ford, R.J. Constitutive BR3 receptor signaling in diffuse, large B-cell lymphomas stabilizes nuclear factor-κB-inducing kinase while activating both canonical and alternative nuclear factor-κB pathways. Blood 2011, 117, 200–210. [Google Scholar] [CrossRef]
- Bohn, J.P.; Salcher, S.; Pircher, A.; Untergasser, G.; Wolf, D. The Biology of Classic Hairy Cell Leukemia. Int. J. Mol. Sci. 2021, 22, 7780. [Google Scholar] [CrossRef] [PubMed]
- Chiu, A.; Chen, K.; He, B.; Dillon, S.; Chadburn, A.; Knowles, D.M.; Xu, W.; Cerutti, A. Malignant B Cells from Hairy Cell Leukemia Express an Innate Phenotype and Undergo IgD Class Switching in Response to Innate Environmental Factors, Including BAFF and APRIL. Blood 2007, 110, 4707. [Google Scholar] [CrossRef]
- Chiu, A.; Xu, W.; He, B.; Santini, P.; Dillon, S.R.; Chadburn, A.; Knowles, D.M.; Cerutti, A. Splenic Sinusoids Stimulate the Survival and Proliferation of Hairy Cell Leukemia B Cells through BAFF, APRIL and Heparan-Sulphate Proteoglycans. Blood 2006, 108, 4959. [Google Scholar] [CrossRef]
- Zhang, K.; Roy, N.K.; Vicioso, Y.; Woo, J.; Beck, R.; de Lima, M.; Caimi, P.; Feinberg, D.; Parameswaran, R. BAFF receptor antibody for mantle cell lymphoma therapy. Oncoimmunology 2021, 10, 1893501. [Google Scholar] [CrossRef]
- Swerdlow, S.H.; Campo, E.; Harris, N.L.; Jaffe, E.S.; Pileri, S.A.; Stein, H.; Thiele, J.; Vardiman, J.W. WHO Classification of Tumours of Haematopoietic and Lymphoid Tissues; International Agency for Research on Cancer Lyon: Lyon, France, 2008; Volume 2. [Google Scholar]
- Rossi, D.; Bertoni, F.; Zucca, E. Marginal-Zone Lymphomas. N. Engl. J. Med. 2022, 386, 568–581. [Google Scholar] [CrossRef] [PubMed]
- Mhawech-Fauceglia, P.; Kaya, G.; Sauter, G.; McKee, T.; Donze, O.; Schwaller, J.; Huard, B. The source of APRIL up-regulation in human solid tumor lesions. J. Leukoc. Biol. 2006, 80, 697–704. [Google Scholar] [CrossRef]
- Moreaux, J.; Veyrune, J.L.; De Vos, J.; Klein, B. APRIL is overexpressed in cancer: Link with tumor progression. BMC Cancer 2009, 9, 83. [Google Scholar] [CrossRef]
- Hinterleitner, C.; Zhou, Y.; Tandler, C.; Heitmann, J.S.; Kropp, K.N.; Hinterleitner, M.; Koch, A.; Hartkopf, A.D.; Zender, L.; Salih, H.R.; et al. Platelet-Expressed TNFRSF13B (TACI) Predicts Breast Cancer Progression. Front. Oncol. 2021, 11, 642170. [Google Scholar] [CrossRef] [PubMed]
- Ding, W.; Ju, S.; Jiang, S.; Zhu, L.; Wang, Y.; Wang, H. Reduced APRIL expression induces cellular senescence via a HSPG-dependent pathway. Pathol. Oncol. Res. 2009, 15, 693–701. [Google Scholar] [CrossRef] [PubMed]
- Pelekanou, V.; Notas, G.; Kampa, M.; Tsentelierou, E.; Stathopoulos, E.N.; Tsapis, A.; Castanas, E. BAFF, APRIL, TWEAK, BCMA, TACI and Fn14 proteins are related to human glioma tumor grade: Immunohistochemistry and public microarray data meta-analysis. PLoS ONE 2013, 8, e83250. [Google Scholar] [CrossRef] [PubMed]
- Deshayes, F.; Laprée, G.; Portier, A.; Richard, Y.; Pencalet, P.; Mahieu-Caputo, D.; Horellou, P.; Tsapis, A. Abnormal production of the TNF-homologue APRIL increases the proliferation of human malignant glioblastoma cell lines via a specific receptor. Oncogene 2004, 23, 3005–3012. [Google Scholar] [CrossRef]
- Notas, G.; Alexaki, V.I.; Kampa, M.; Pelekanou, V.; Charalampopoulos, I.; Sabour-Alaoui, S.; Pediaditakis, I.; Dessirier, V.; Gravanis, A.; Stathopoulos, E.N.; et al. APRIL binding to BCMA activates a JNK2-FOXO3-GADD45 pathway and induces a G2/M cell growth arrest in liver cells. J. Immunol. 2012, 189, 4748–4758. [Google Scholar] [CrossRef]
- Warakomska, M.; Tynecka, M.; Lemancewicz, D.; Grubczak, K.; Dzieciol, J.; Moniuszko, M.; Eljaszewicz, A.; Bolkun, L. The effects of BAFF and APRIL signaling on non-small cell lung cancer cell proliferation and invasiveness. Oncol. Lett. 2021, 22, 728. [Google Scholar] [CrossRef]
- Fabris, M.; Tonutti, E.; Panighel, C.; Blasone, N.; Visentini, D.; Pizzolitto, S.; De Maglio, G.; Isola, M.; Curcio, F.; Grimaldi, F. The role of B-lymphocyte stimulator in neuroendocrine tumors: Correlation with tumor differentiation, disease status and the presence of metastases. Immunol. Endocr. Metab. Agents Med. Chem. (Former. Curr. Med. Chem.-Immunol. Endocr. Metab. Agents) 2011, 11, 306–314. [Google Scholar] [CrossRef]
- Grimaldi, F.; Vescini, F.; Tonelli, V.; Pistis, C.; Kara, E.; Triggiani, V.; Tonutti, E.; Curcio, F.; Fabris, M. Exploring the possible prognostic role of B-Lymphocyte Stimulator (BLyS) in a large series of patients with neuroendocrine tumors. Endocr. Metab. Immune Disord.-Drug Targets (Former. Curr. Drug Targets-Immune Endocr. Metab. Disord.) 2018, 18, 618–625. [Google Scholar] [CrossRef]
- Jablonska, E.; Iwaniuk, A.; Ratajczak-Wrona, W.; Grubczak, K.; Dziemianczyk-Pakiela, D.; Moniuszko, M.; Nowak, K.; Borys, J.; Garley, M. The promoting effect of neutrophil-derived BAFF molecule on the proliferation and life span of CAL-27 oral squamous carcinoma cells. Immunobiology 2022, 227, 152247. [Google Scholar] [CrossRef]
- Jabłońska, E.; Wawrusiewicz-Kurylonek, N.; Garley, M.; Ratajczak-Wrona, W.; Antonowicz, B.; Dziemiańczyk-Pakieła, D.; Jabłoński, J.; Krętowski, A.; Grabowska, S.Z. A proliferation-inducing ligand (APRIL) in neutrophils of patients with oral cavity squamous cell carcinoma. Eur. Cytokine Netw. 2012, 23, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Koizumi, M.; Hiasa, Y.; Kumagi, T.; Yamanishi, H.; Azemoto, N.; Kobata, T.; Matsuura, B.; Abe, M.; Onji, M. Increased B cell-activating factor promotes tumor invasion and metastasis in human pancreatic cancer. PLoS ONE 2013, 8, e71367. [Google Scholar] [CrossRef] [PubMed]
- Pelekanou, V.; Notas, G.; Theodoropoulou, K.; Kampa, M.; Takos, D.; Alexaki, V.I.; Radojicic, J.; Sofras, F.; Tsapis, A.; Stathopoulos, E.N.; et al. Detection of the TNFSF members BAFF, APRIL, TWEAK and their receptors in normal kidney and renal cell carcinomas. Anal. Cell Pathol. 2011, 34, 49–60. [Google Scholar] [CrossRef]
- Alexaki, V.I.; Pelekanou, V.; Notas, G.; Venihaki, M.; Kampa, M.; Dessirier, V.; Sabour-Alaoui, S.; Stathopoulos, E.N.; Tsapis, A.; Castanas, E. B-cell maturation antigen (BCMA) activation exerts specific proinflammatory effects in normal human keratinocytes and is preferentially expressed in inflammatory skin pathologies. Endocrinology 2012, 153, 739–749. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Süsskind, D. Exploring the role of BAFF as biomarker in the detection of uveal melanoma metastases. J. Cancer Res. Clin. Oncol. 2021, 147, 1389–1405. [Google Scholar] [CrossRef]
- Bienertova-Vasku, J.; Lungova, A.; Bienert, P.; Zlamal, F.; Tomandl, J.; Tomandlova, M.; Splichal, Z.; Sterba, J. Circulating levels of B-cell activating factor in paediatric patients with malignancy with or without cancer-related cachexia. Klin. Onkol. 2012, 25, S58–S63. [Google Scholar]
- García-Castro, A.; Zonca, M.; Florindo-Pinheiro, D.; Carvalho-Pinto, C.E.; Cordero, A.; Gutiérrez del Fernando, B.; García-Grande, A.; Mañes, S.; Hahne, M.; González-Suárez, E.; et al. APRIL promotes breast tumor growth and metastasis and is associated with aggressive basal breast cancer. Carcinogenesis 2015, 36, 574–584. [Google Scholar] [CrossRef]
- Pelekanou, V.; Notas, G.; Athanasouli, P.; Alexakis, K.; Kiagiadaki, F.; Peroulis, N.; Kalyvianaki, K.; Kampouri, E.; Polioudaki, H.; Theodoropoulos, P.; et al. BCMA (TNFRSF17) Induces APRIL and BAFF Mediated Breast Cancer Cell Stemness. Front. Oncol. 2018, 8, 301. [Google Scholar] [CrossRef]
- Abo-Elfadl, M.T.; Gamal-Eldeen, A.M.; Ismail, M.F.; Shahin, N.N. Silencing of the cytokine receptor TNFRSF13B: A new therapeutic target for triple-negative breast cancer. Cytokine 2020, 125, 154790. [Google Scholar] [CrossRef]
- Lascano, V.; Zabalegui, L.F.; Cameron, K.; Guadagnoli, M.; Jansen, M.; Burggraaf, M.; Versloot, M.; Rodermond, H.; van der Loos, C.; Carvalho-Pinto, C.E.; et al. The TNF family member APRIL promotes colorectal tumorigenesis. Cell Death Differ. 2012, 19, 1826–1835. [Google Scholar] [CrossRef]
- Ding, W.; Wang, G.; Shao, K.; Wang, F.; Huang, H.; Ju, S.; Cong, H.; Wang, H. Amelioration of colorectal cancer using negative lipidoid nanoparticles to encapsulate siRNA against APRIL by enema delivery mode. Pathol. Oncol. Res. 2014, 20, 953–964. [Google Scholar] [CrossRef]
- Wang, J.; Ding, W.; Sun, B.; Jing, R.; Huang, H.; Shi, G.; Wang, H. Targeting of colorectal cancer growth, metastasis, and anti-apoptosis in BALB/c nude mice via APRIL siRNA. Mol. Cell. Biochem. 2012, 363, 1–10. [Google Scholar] [CrossRef]
- Roth, W.; Wagenknecht, B.; Klumpp, A.; Naumann, U.; Hahne, M.; Tschopp, J.; Weller, M. APRIL, a new member of the tumor necrosis factor family, modulates death ligand-induced apoptosis. Cell Death Differ. 2001, 8, 403–410. [Google Scholar] [CrossRef]
- Gao, L.; Morine, Y.; Yamada, S.; Saito, Y.; Ikemoto, T.; Tokuda, K.; Miyazaki, K.; Okikawa, S.; Takasu, C.; Shimada, M. The BAFF/NFκB axis is crucial to interactions between sorafenib-resistant HCC cells and cancer-associated fibroblasts. Cancer Sci. 2021, 112, 3545–3554. [Google Scholar] [CrossRef] [PubMed]
- Dou, H.; Yan, Z.; Zhang, M.; Xu, X. APRIL, BCMA and TACI proteins are abnormally expressed in non-small cell lung cancer. Oncol. Lett. 2016, 12, 3351–3355. [Google Scholar] [CrossRef] [PubMed]
- Yarchoan, M.; Ho, W.J.; Mohan, A.; Shah, Y.; Vithayathil, T.; Leatherman, J.; Dennison, L.; Zaidi, N.; Ganguly, S.; Woolman, S.; et al. Effects of B cell-activating factor on tumor immunity. JCI Insight 2020, 5. [Google Scholar] [CrossRef] [PubMed]
- Di Carlo, E.; D’Antuono, T.; Pompa, P.; Giuliani, R.; Rosini, S.; Stuppia, L.; Musiani, P.; Sorrentino, C. The lack of epithelial interleukin-7 and BAFF/BLyS gene expression in prostate cancer as a possible mechanism of tumor escape from immunosurveillance. Clin. Cancer Res. 2009, 15, 2979–2987. [Google Scholar] [CrossRef]
- Jianyi, D.; Haili, G.; Bo, Y.; Meiqin, Y.; Baoyou, H.; Haoran, H.; Fang, L.; Qingliang, Z.; Lingfei, H. Myeloid-derived suppressor cells cross-talk with B10 cells by BAFF/BAFF-R pathway to promote immunosuppression in cervical cancer. Cancer Immunol. Immunother. 2022, 72, 73–85. [Google Scholar] [CrossRef]
- Tandler, C.; Schmidt, M.; Heitmann, J.S.; Hierold, J.; Schmidt, J.; Schneider, P.; Dorfel, D.; Walz, J.; Salih, H.R. Neutralization of B-Cell Activating Factor (BAFF) by Belimumab Reinforces Small Molecule Inhibitor Treatment in Chronic Lymphocytic Leukemia. Cancers 2020, 12, 2725. [Google Scholar] [CrossRef]
- Wild, J.; Schmiedel, B.J.; Maurer, A.; Raab, S.; Prokop, L.; Stevanovic, S.; Dorfel, D.; Schneider, P.; Salih, H.R. Neutralization of (NK-cell-derived) B-cell activating factor by Belimumab restores sensitivity of chronic lymphoid leukemia cells to direct and Rituximab-induced NK lysis. Leukemia 2015, 29, 1676–1683. [Google Scholar] [CrossRef]
- Raje, N.S.; Faber, E.A., Jr.; Richardson, P.G.; Schiller, G.; Hohl, R.J.; Cohen, A.D.; Forero, A.; Carpenter, S.; Nguyen, T.S.; Conti, I.; et al. Phase 1 Study of Tabalumab, a Human Anti-B-Cell Activating Factor Antibody, and Bortezomib in Patients with Relapsed/Refractory Multiple Myeloma. Clin. Cancer Res. 2016, 22, 5688–5695. [Google Scholar] [CrossRef]
- Raje, N.S.; Moreau, P.; Terpos, E.; Benboubker, L.; Grząśko, N.; Holstein, S.A.; Oriol, A.; Huang, S.Y.; Beksac, M.; Kuliczkowski, K.; et al. Phase 2 study of tabalumab, a human anti-B-cell activating factor antibody, with bortezomib and dexamethasone in patients with previously treated multiple myeloma. Br. J. Haematol. 2017, 176, 783–795. [Google Scholar] [CrossRef]
- Iida, S.; Ogiya, D.; Abe, Y.; Taniwaki, M.; Asou, H.; Maeda, K.; Uenaka, K.; Nagaoka, S.; Ishiki, T.; Conti, I.; et al. Dose-escalation study of tabalumab with bortezomib and dexamethasone in Japanese patients with multiple myeloma. Cancer Sci. 2016, 107, 1281–1289. [Google Scholar] [CrossRef] [PubMed]
- Wong, D.P.; Roy, N.K.; Zhang, K.; Anukanth, A.; Asthana, A.; Shirkey-Son, N.J.; Dunmire, S.; Jones, B.J.; Lahr, W.S.; Webber, B.R.; et al. A BAFF ligand-based CAR-T cell targeting three receptors and multiple B cell cancers. Nat. Commun. 2022, 13, 217. [Google Scholar] [CrossRef] [PubMed]
- Phase 1 Study of BAFF CAR-T Cells (LMY-920) for Non-Hodgkin Lymphoma. Available online: https://www.clinicaltrials.gov/ct2/show/NCT05312801?term=BAFF&cond=Cancer&draw=2&rank=2 (accessed on 8 August 2022).
- Guadagnoli, M.; Kimberley, F.C.; Phan, U.; Cameron, K.; Vink, P.M.; Rodermond, H.; Eldering, E.; Kater, A.P.; van Eenennaam, H.; Medema, J.P. Development and characterization of APRIL antagonistic monoclonal antibodies for treatment of B-cell lymphomas. Blood 2011, 117, 6856–6865. [Google Scholar] [CrossRef] [PubMed]
- Lee, L.; Draper, B.; Chaplin, N.; Philip, B.; Chin, M.; Galas-Filipowicz, D.; Onuoha, S.; Thomas, S.; Baldan, V.; Bughda, R.; et al. An APRIL-based chimeric antigen receptor for dual targeting of BCMA and TACI in multiple myeloma. Blood 2018, 131, 746–758. [Google Scholar] [CrossRef]
- Schmidts, A.; Ormhøj, M.; Choi, B.D.; Taylor, A.O.; Bouffard, A.A.; Scarfò, I.; Larson, R.C.; Frigault, M.J.; Gallagher, K.; Castano, A.P.; et al. Rational design of a trimeric APRIL-based CAR-binding domain enables efficient targeting of multiple myeloma. Blood Adv. 2019, 3, 3248–3260. [Google Scholar] [CrossRef]
- McWilliams, E.M.; Lucas, C.R.; Chen, T.; Harrington, B.K.; Wasmuth, R.; Campbell, A.; Rogers, K.A.; Cheney, C.M.; Mo, X.; Andritsos, L.A.; et al. Anti-BAFF-R antibody VAY-736 demonstrates promising preclinical activity in CLL and enhances effectiveness of ibrutinib. Blood Adv. 2019, 3, 447–460. [Google Scholar] [CrossRef]
- Vicioso, Y.; Gram, H.; Beck, R.; Asthana, A.; Zhang, K.; Wong, D.P.; Letterio, J.; Parameswaran, R. Combination Therapy for Treating Advanced Drug-Resistant Acute Lymphoblastic Leukemia. Cancer Immunol. Res. 2019, 7, 1106–1119. [Google Scholar] [CrossRef]
- Wang, X.; Dong, Z.; Awuah, D.; Chang, W.C.; Cheng, W.A.; Vyas, V.; Cha, S.C.; Anderson, A.J.; Zhang, T.; Wang, Z.; et al. CD19/BAFF-R dual-targeted CAR T cells for the treatment of mixed antigen-negative variants of acute lymphoblastic leukemia. Leukemia 2022, 36, 1015–1024. [Google Scholar] [CrossRef]
- Dong, Z.; Cheng, W.A.; Smith, D.L.; Huang, B.; Zhang, T.; Chang, W.C.; Wang, X.; Forman, S.J.; Kwak, L.W.; Qin, H. Antitumor efficacy of BAFF-R targeting CAR T cells manufactured under clinic-ready conditions. Cancer Immunol. Immunother. 2020, 69, 2139–2145. [Google Scholar] [CrossRef]
- Paul, B.; Rodriguez, C.; Usmani, S.Z. BCMA-Targeted Biologic Therapies: The Next Standard of Care in Multiple Myeloma Therapy. Drugs 2022, 82, 613–631. [Google Scholar] [CrossRef] [PubMed]
- Topp, M.S.; Duell, J.; Zugmaier, G.; Attal, M.; Moreau, P.; Langer, C.; Krönke, J.; Facon, T.; Salnikov, A.V.; Lesley, R.; et al. Anti-B-Cell Maturation Antigen BiTE Molecule AMG 420 Induces Responses in Multiple Myeloma. J. Clin. Oncol. 2020, 38, 775–783. [Google Scholar] [CrossRef] [PubMed]
- Hipp, S.; Tai, Y.T.; Blanset, D.; Deegen, P.; Wahl, J.; Thomas, O.; Rattel, B.; Adam, P.J.; Anderson, K.C.; Friedrich, M. A novel BCMA/CD3 bispecific T-cell engager for the treatment of multiple myeloma induces selective lysis in vitro and in vivo. Leukemia 2017, 31, 1743–1751. [Google Scholar] [CrossRef] [PubMed]
- Seckinger, A.; Delgado, J.A.; Moser, S.; Moreno, L.; Neuber, B.; Grab, A.; Lipp, S.; Merino, J.; Prosper, F.; Emde, M.; et al. Target Expression, Generation, Preclinical Activity, and Pharmacokinetics of the BCMA-T Cell Bispecific Antibody EM801 for Multiple Myeloma Treatment. Cancer Cell 2017, 31, 396–410. [Google Scholar] [CrossRef]
- Costa, L.J.; Wong, S.W.; Bermudez, A.; de la Rubia, J.; Mateos, M.-V.; Ocio, E.M.; Rodriguez-Otero, P.; San-Miguel, J.; Li, S.; Sarmiento, R. First clinical study of the B-cell maturation antigen (BCMA) 2+ 1 T cell engager (TCE) CC-93269 in patients (Pts) with relapsed/refractory multiple myeloma (RRMM): Interim results of a phase 1 multicenter trial. Blood 2019, 134, 143. [Google Scholar] [CrossRef]
- Cho, S.-F.; Lin, L.; Xing, L.; Wen, K.; Yu, T.; Hsieh, P.A.; Li, Y.; Munshi, N.C.; Wahl, J.; Matthes, K.; et al. AMG 701 Potently Induces Anti-Multiple Myeloma (MM) Functions of T Cells and IMiDs Further Enhance Its Efficacy to Prevent MM Relapse In Vivo. Blood 2019, 134, 135. [Google Scholar] [CrossRef]
- Usmani, S.Z.; Mateos, M.-V.; Nahi, H.; Krishnan, A.Y.; van de Donk, N.W.; San Miguel, J.; Oriol, A.; Rosiñol, L.; Chari, A.; Adams, H. Phase I study of teclistamab, a humanized B-cell maturation antigen (BCMA) x CD3 bispecific antibody, in relapsed/refractory multiple myeloma (R/R MM). J. Clin. Oncol. 2020, 38, 100. [Google Scholar] [CrossRef]
- Usmani, S.Z.; Garfall, A.L.; van de Donk, N.; Nahi, H.; San-Miguel, J.F.; Oriol, A.; Rosinol, L.; Chari, A.; Bhutani, M.; Karlin, L.; et al. Teclistamab, a B-cell maturation antigen × CD3 bispecific antibody, in patients with relapsed or refractory multiple myeloma (MajesTEC-1): A multicentre, open-label, single-arm, phase 1 study. Lancet 2021, 398, 665–674. [Google Scholar] [CrossRef]
- Zhou, X.; Einsele, H.; Danhof, S. Bispecific Antibodies: A New Era of Treatment for Multiple Myeloma. J. Clin. Med. 2020, 9, 2166. [Google Scholar] [CrossRef]
- Lambert, J.M.; Morris, C.Q. Antibody-Drug Conjugates (ADCs) for Personalized Treatment of Solid Tumors: A Review. Adv. Ther. 2017, 34, 1015–1035. [Google Scholar] [CrossRef]
- Baines, A.C.; Ershler, R.; Kanapuru, B.; Xu, Q.; Shen, G.; Li, L.; Ma, L.; Okusanya, O.O.; Simpson, N.E.; Nguyen, W.; et al. FDA Approval Summary: Belantamab Mafodotin for Patients with Relapsed or Refractory Multiple Myeloma. Clin. Cancer Res. 2022, 28, 4629–4633. [Google Scholar] [CrossRef]
- Kinneer, K.; Flynn, M.; Thomas, S.B.; Meekin, J.; Varkey, R.; Xiao, X.; Zhong, H.; Breen, S.; Hynes, P.G.; Fleming, R.; et al. Preclinical assessment of an antibody-PBD conjugate that targets BCMA on multiple myeloma and myeloma progenitor cells. Leukemia 2019, 33, 766–771. [Google Scholar] [CrossRef]
- Singh, R.K.; Jones, R.J.; Hong, S.; Shirazi, F.; Wang, H.; Kuiatse, I.; Pahl, A.; Orlowski, R.Z. HDP101, a Novel B-Cell Maturation Antigen (BCMA)-Targeted Antibody Conjugated to α-Amanitin, Is Active Against Myeloma with Preferential Efficacy Against Pre-Clinical Models of Deletion 17p. Blood 2018, 132, 593. [Google Scholar] [CrossRef]
- Ali, S.A.; Shi, V.; Maric, I.; Wang, M.; Stroncek, D.F.; Rose, J.J.; Brudno, J.N.; Stetler-Stevenson, M.; Feldman, S.A.; Hansen, B.G.; et al. T cells expressing an anti–B-cell maturation antigen chimeric antigen receptor cause remissions of multiple myeloma. Blood 2016, 128, 1688–1700. [Google Scholar] [CrossRef]
- Brudno, J.N.; Maric, I.; Hartman, S.D.; Rose, J.J.; Wang, M.; Lam, N.; Stetler-Stevenson, M.; Salem, D.; Yuan, C.; Pavletic, S.; et al. T Cells Genetically Modified to Express an Anti-B-Cell Maturation Antigen Chimeric Antigen Receptor Cause Remissions of Poor-Prognosis Relapsed Multiple Myeloma. J. Clin. Oncol. 2018, 36, 2267–2280. [Google Scholar] [CrossRef]
- Raje, N.; Berdeja, J.; Lin, Y.; Siegel, D.; Jagannath, S.; Madduri, D.; Liedtke, M.; Rosenblatt, J.; Maus, M.V.; Turka, A.; et al. Anti-BCMA CAR T-Cell Therapy bb2121 in Relapsed or Refractory Multiple Myeloma. N. Engl. J. Med. 2019, 380, 1726–1737. [Google Scholar] [CrossRef]
- Munshi, N.C.; Anderson, L.D., Jr.; Shah, N.; Madduri, D.; Berdeja, J.; Lonial, S.; Raje, N.; Lin, Y.; Siegel, D.; Oriol, A.; et al. Idecabtagene Vicleucel in Relapsed and Refractory Multiple Myeloma. N. Engl. J. Med. 2021, 384, 705–716. [Google Scholar] [CrossRef]
- Zhao, W.H.; Liu, J.; Wang, B.Y.; Chen, Y.X.; Cao, X.M.; Yang, Y.; Zhang, Y.L.; Wang, F.X.; Zhang, P.Y.; Lei, B.; et al. A phase 1, open-label study of LCAR-B38M, a chimeric antigen receptor T cell therapy directed against B cell maturation antigen, in patients with relapsed or refractory multiple myeloma. J. Hematol. Oncol. 2018, 11, 141. [Google Scholar] [CrossRef] [PubMed]
- Samur, M.K.; Fulciniti, M.; Aktas Samur, A.; Bazarbachi, A.H.; Tai, Y.T.; Prabhala, R.; Alonso, A.; Sperling, A.S.; Campbell, T.; Petrocca, F.; et al. Biallelic loss of BCMA as a resistance mechanism to CAR T cell therapy in a patient with multiple myeloma. Nat. Commun. 2021, 12, 868. [Google Scholar] [CrossRef] [PubMed]
- Ahn, S.; Leblay, N.; Neri, P. Understanding the Mechanisms of Resistance to T Cell-based Immunotherapies to Develop More Favorable Strategies in Multiple Myeloma. Hemasphere 2021, 5, e575. [Google Scholar] [CrossRef] [PubMed]
- Yan, Z.; Cao, J.; Cheng, H.; Qiao, J.; Zhang, H.; Wang, Y.; Shi, M.; Lan, J.; Fei, X.; Jin, L.; et al. A combination of humanised anti-CD19 and anti-BCMA CAR T cells in patients with relapsed or refractory multiple myeloma: A single-arm, phase 2 trial. Lancet Haematol. 2019, 6, e521–e529. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Yin, H.; Zhao, X.; Jin, D.; Liang, Y.; Xiong, T.; Li, L.; Tang, W.; Zhang, J.; Liu, M.; et al. High efficacy and safety of CD38 and BCMA bispecific CAR-T in relapsed or refractory multiple myeloma. J. Exp. Clin. Cancer Res. 2022, 41, 2. [Google Scholar] [CrossRef] [PubMed]
- Larson, R.; Castano, A.; Bouffard, A.; Scarfo, I.; Kann, M.; Schmidts, A.; Gallagher, K.; Maus, M.V. Bispecific CAR T cells for multiple myeloma: Natural ligand compared to tandem scFv design. J. Immunol. 2020, 204, 246.3. [Google Scholar] [CrossRef]
- Miao, Y.R.; Thakkar, K.; Cenik, C.; Jiang, D.; Mizuno, K.; Jia, C.; Li, C.G.; Zhao, H.; Diep, A.; Xu, Y.; et al. Developing high-affinity decoy receptors to treat multiple myeloma and diffuse large B cell lymphoma. J. Exp. Med. 2022, 219. [Google Scholar] [CrossRef] [PubMed]
- Yaccoby, S.; Pennisi, A.; Li, X.; Dillon, S.R.; Zhan, F.; Barlogie, B.; Shaughnessy, J.D., Jr. Atacicept (TACI-Ig) inhibits growth of TACI(high) primary myeloma cells in SCID-hu mice and in coculture with osteoclasts. Leukemia 2008, 22, 406–413. [Google Scholar] [CrossRef]
- Rossi, J.F. Phase I study of atacicept in relapsed/refractory multiple myeloma (MM) and Waldenström’s macroglobulinemia. Clin. Lymphoma Myeloma Leuk. 2011, 11, 136–138. [Google Scholar] [CrossRef]
- Kofler, D.M.; Gawlik, B.B.; Elter, T.; Gianella-Borradori, A.; Wendtner, C.M.; Hallek, M. Phase 1b trial of atacicept, a recombinant protein binding BLyS and APRIL, in patients with chronic lymphocytic leukemia. Leukemia 2012, 26, 841–844. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Hematological Cancers | Expression Profile and Clinical Relevancy |
|---|---|
| B-ALL | Serum BAFF and APRIL concentrations are higher in B-ALL patients [114]. BAFF-R expression in primary B-ALL cases is heterogeneous (0.9% to 93% of B-ALL cells express BAFF-R) [115]. TACI and BCMA expression is relatively low to negative in B-ALL cells [115,116]. |
| HL | Increased serum levels of BAFF and APRIL in HL [117]. |
| MM | Serum levels BAFF and APRIL are three to five times higher in MM [118,119]. Higher BAFF expression is associated with disease progression and shorter progression-free survival [118,120,121,122]. BAFF-R expression is very low or absent in primary MM cells and in MM cell lines [119]. BCMA is highly expressed in the plasma cells of MM patients, and serum soluble BCMA (sBCMA) levels are used as a biomarker for MM disease status [123,124]. Patients with higher levels (above 326.4 ng/mL) of sBCMA have significantly shorter progression-free (3.6 months) and overall survival (98 months) than patients with less than 326.4 ng/mL of serum sBCMA (9.0 and 155 months, respectively) [125]. Higher TACI expression is observed in on MM cells [126,127]; however, lower TACI expression is associated with worse prognosis, including increased stage III MM probability, attenuated hemoglobin levels, and increased bone lesions [127]. |
| NHL | |
| BL | BAFF-R expression in primary BL cases is heterogeneous (0.04 to 81% of B-ALL cells express BAFF-R) [115]. |
| CLL | Increased BAFF level in CLL patients, specifically with unmutated IgHV, and increased BAFF expression is associated with worse outcomes [128,129]. The plasma level of APRIL is higher in CLL patients [128,129]. Higher intracellular APRIL and BAFF in CLL cells is associated with higher expression of adverse prognostic factors CD38 and ZAP70 and poorer clinical outcomes [128]. The expression of BAFF-R, TACI and BCMA on CLL B cells is comparable to healthy B cells [130,131,132]; however, lower BAFF-R expression on CLL B cells has also been reported [133]. CLL B cells with mutated IgHV express more TACI and BCMA than unmutated cells [132]. Plasma sBCMA levels are significantly higher in CLL patients, with the sBCMA concentration increasing with disease severity and associated with poorer outcomes [134,135]. |
| CNSL | Elevated levels of BAFF-APRIL and their receptors BCMA and TACI have been detected in the cerebrospinal fluid (CSF) and biopsies of CNSL patients, compared to patients with other neurological diseases [136,137,138]. The levels of BAFF and APRIL in CSF act as sensitive and specific biomarkers for CNSL diagnosis and therapeutic response. The serum levels of BAFF or APRIL remain unchanged, suggesting a localized response. |
| DLBCL | BAFF-R expression is comparable to healthy B cells [133]. Tumor cells also express TACI, BCMA and HSPGs [139]. Serum BAFF concentrations and APRIL expression in tumor lesions are higher and associated with poor prognosis [139,140]. |
| FL | Lower BAFF-R expression [133]. Three-fold higher serum BAFF expression compared to healthy donors [133]. |
| HCL | HCL cells express high levels of BAFF-R, TACI, BCMA, and HSPGs [141]. |
| MCL | Serum BAFF concentrations are higher and correlate to poor treatment response and relapse [142]. BAFF-R expression is comparable to healthy B cells [133]. |
| MZL | BAFF-R expression is comparable to healthy B cells [133]. |
| Solid Cancers | Expression Profile and Clinical Relevancy |
|---|---|
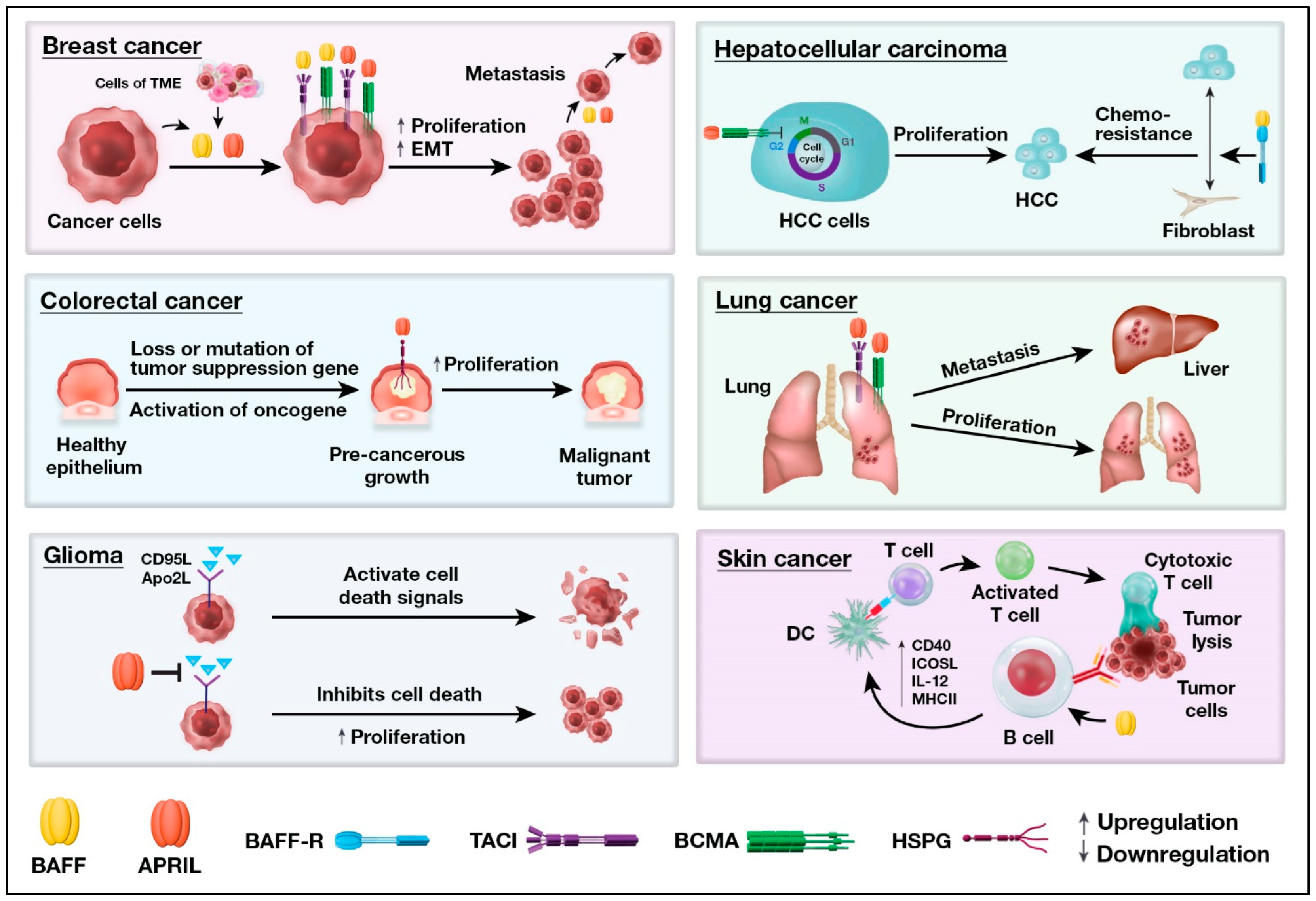
| Breast cancer | BAFF is ubiquitously expressed in malignant and non-malignant tissue, whereas APRIL expression is lower in malignant tissues than in non-malignant tissues [29]. Other studies reported overexpression of APRIL protein and mRNA in tissue lesions [180,181]. None of the receptors can be identified immunohistochemically, yet the mRNAs of the receptors are detectable, most likely related to tumor-infiltrating immune cells [29,182]. |
| Colorectal cancers | Higher levels of APRIL mRNA have been reported [2]. Colorectal cancer cell lines express higher levels of HSPG, with only marginal expression of BCMA and TACI observed [183]. |
| Glioma | Higher levels of APRIL, BCMA, and TACI expression and lower BAFF protein levels in glioma [184]. BAFF, APRIL, and their receptors are expressed by the majority of glioblastoma (a type of glioma) cell lines [185]. |
| HCC | APRIL and BCMA levels are higher, whereas BAFF and BAFF-R expressions are unchanged in HCC [186]. HCC does not express TACI. |
| Lung cancer | The non-small cell lung cancer (NSCLC) cell lines express BAFF and APRIL and the receptors BAFF-R and TACI, but do not express BCMA [187]. |
| Neuroendocrine tumors | Higher BAFF expression is associated with disease severity and refractory disease [188,189]. |
| Oral squamous cell carcinoma | Overexpression of BAFF and APRIL [190,191]. |
| Pancreatic cancer | Serum BAFF concentration is significantly higher and associated with disease severity and metastasis [192]. |
| Renal carcinoma | BAFF, APRIL, and TACI are highly expressed in cancer biopsies, while the expression of BAFF-R and BCMA is very low to undetectable [193]. Higher levels of APRIL expression in biopsies are associated with disease severity and negatively correlated with disease-free survival. |
| Skin cancer | Higher expression of BAFF, APRIL, and BCMA in skin cancers, specifically in uveal melanoma, has been reported [194,195]. High BAFF expression is a predictor of metastasis. |
| Thyroid carcinoma | Higher APRIL mRNA expression is found in thyroid carcinoma [2]. |
| Others | Serum concentrations of BAFF in nephroblastoma (Wilms’ tumor), Ewing sarcoma, and rhabdomyosarcoma were higher than in healthy controls and the childhood non-Hodgkin’s lymphoma subgroup [196]. |
| Target | Drug Name | Developmental Phase | Indication |
|---|---|---|---|
| Anti-BAFF antibody | Belimumab | Phase II | Relapsed and/or refractory CLL |
| Tabalumab | Phase II | Relapsed and/or refractory MM | |
| BAFF CAR-T cells | LMY-920 | Phase I | Relapsed and/or refractory NHL |
| Anti-APRIL antibody | BION-1301 | Phase I | Relapsed and/or refractory MM |
| APRIL CAR-T cells | APRIL CAR-T | Phase I | Relapsed and/or refractory MM |
| AUTO2 | Phase I/II | Relapsed and/or refractory MM | |
| Anti-BAFF-R antibody | VAY-736 | Phase I/Ib | NHLs (DLBCL, FL, MCL, MZL) |
| BAFF-R CAR-T cells | BAFF-R CAR-T | Phase I | Relapsed or refractory B-cell ALL and MCL |
| Anti-BCMA and anti-CD3ε bispecific antibody | AMG420 | Phase I | Relapsed and/or refractory MM |
| AMG701 | Phase I/II | Relapsed and/or refractory MM | |
| Teclistamab | Phase II/III | Relapsed and/or refractory MM | |
| Elranatamab | Phase II/III | MM | |
| REGN-5458 | Phase I/II | Relapsed and/or refractory MM | |
| TNB-383B | Phase I | Relapsed and/or refractory MM | |
| Anti-BCMA ADC | Belantamab Mafodotin | Phase III/FDA approved | Relapsed and/or refractory MM who has received at least four prior therapies |
| MEDI2228 | Phase I | Relapsed and/or refractory MM | |
| HDP-101 | Phase I/II | Relapsed and/or refractory MM | |
| BCMA CAR-T cells | Idecabtagene vicleucel | Phase II/FDA approved | Relapsed and/or refractory MM and MM patients received hematopoietic stem cell transplantation |
| LCAR-B38M CAR-T | Phase II | Relapsed and/or refractory MM | |
| BCMA/CD19 | Phase I | Relapsed and/or refractory MM, B-ALL | |
| BCMA/CD38 | Phase I/II | Relapsed and/or refractory MM | |
| TACI-Fc-chimera protein | Atacicept | Phase I | Relapsed and/or refractory MM and CLL |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ullah, M.A.; Mackay, F. The BAFF-APRIL System in Cancer. Cancers 2023, 15, 1791. https://doi.org/10.3390/cancers15061791
Ullah MA, Mackay F. The BAFF-APRIL System in Cancer. Cancers. 2023; 15(6):1791. https://doi.org/10.3390/cancers15061791
Chicago/Turabian StyleUllah, Md Ashik, and Fabienne Mackay. 2023. "The BAFF-APRIL System in Cancer" Cancers 15, no. 6: 1791. https://doi.org/10.3390/cancers15061791
APA StyleUllah, M. A., & Mackay, F. (2023). The BAFF-APRIL System in Cancer. Cancers, 15(6), 1791. https://doi.org/10.3390/cancers15061791