
Hyaluronan-Induced CD44-iASPP Interaction Affects Fibroblast Migration and Survival

 , ,
, ,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. DNA Constructs
2.2. Cell Culture and Antibodies
2.3. Transfections
2.4. Cell Lysis, Subcellular Fractionation, Immunoblotting, and Co-Immunoprecipitation
2.5. Proximity Ligation Assay
2.6. In Silico Analysis
2.7. Cell Proliferation Assay
2.8. Immunofluorescence Staining
2.9. Scratch Wound Migration Assay
2.10. Adhesion Assay
2.11. Measurement of Intracellular ROS Levels
3. Results
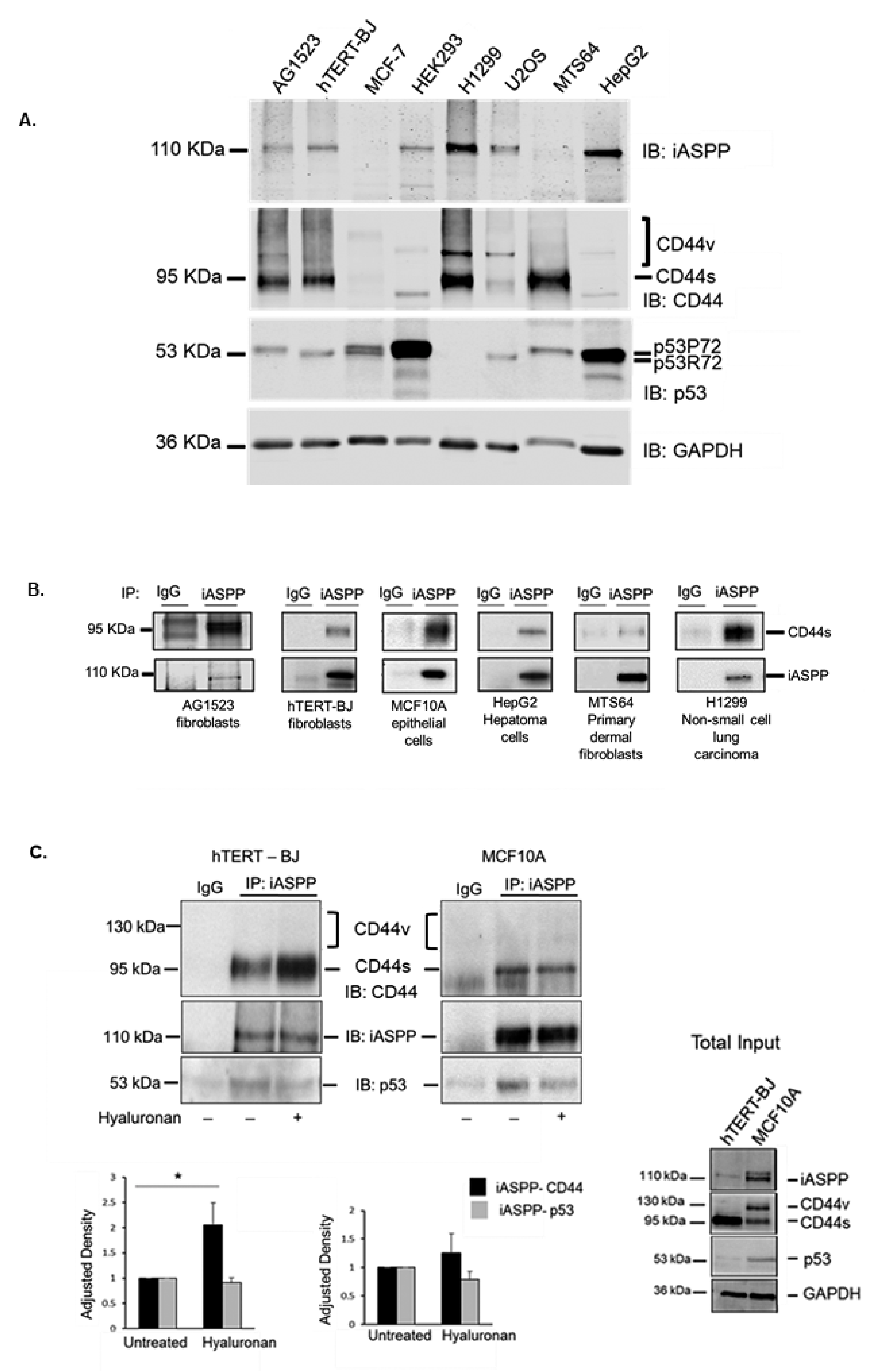
3.1. The Standard Isoform of CD44 Binds to iASPP in Mesenchymal and Epithelial Cells
3.2. Hyaluronan Enhanced the Amount of iASPP-CD44s Complexes in Mesenchymal Cells but Not in Epithelial Cells
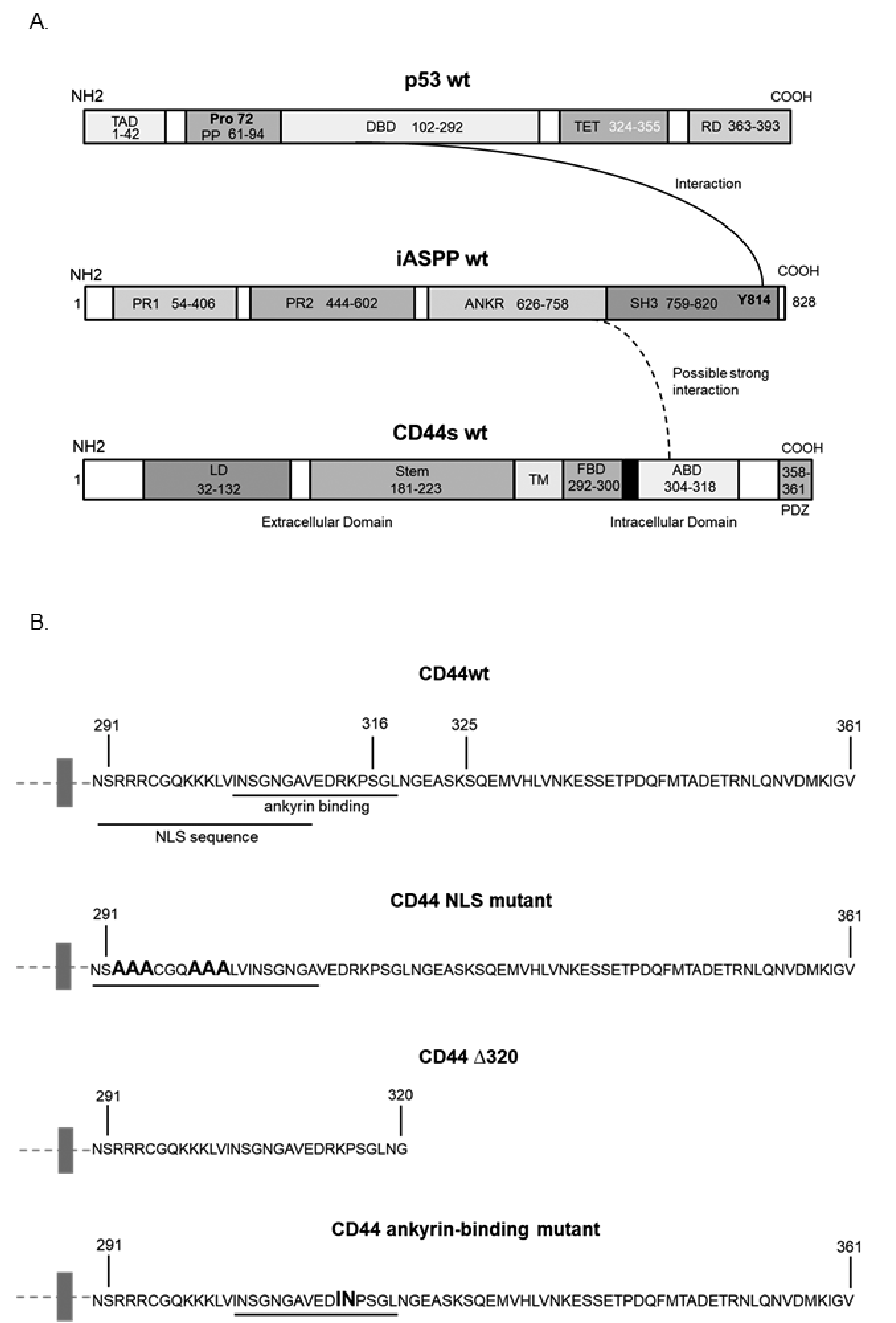
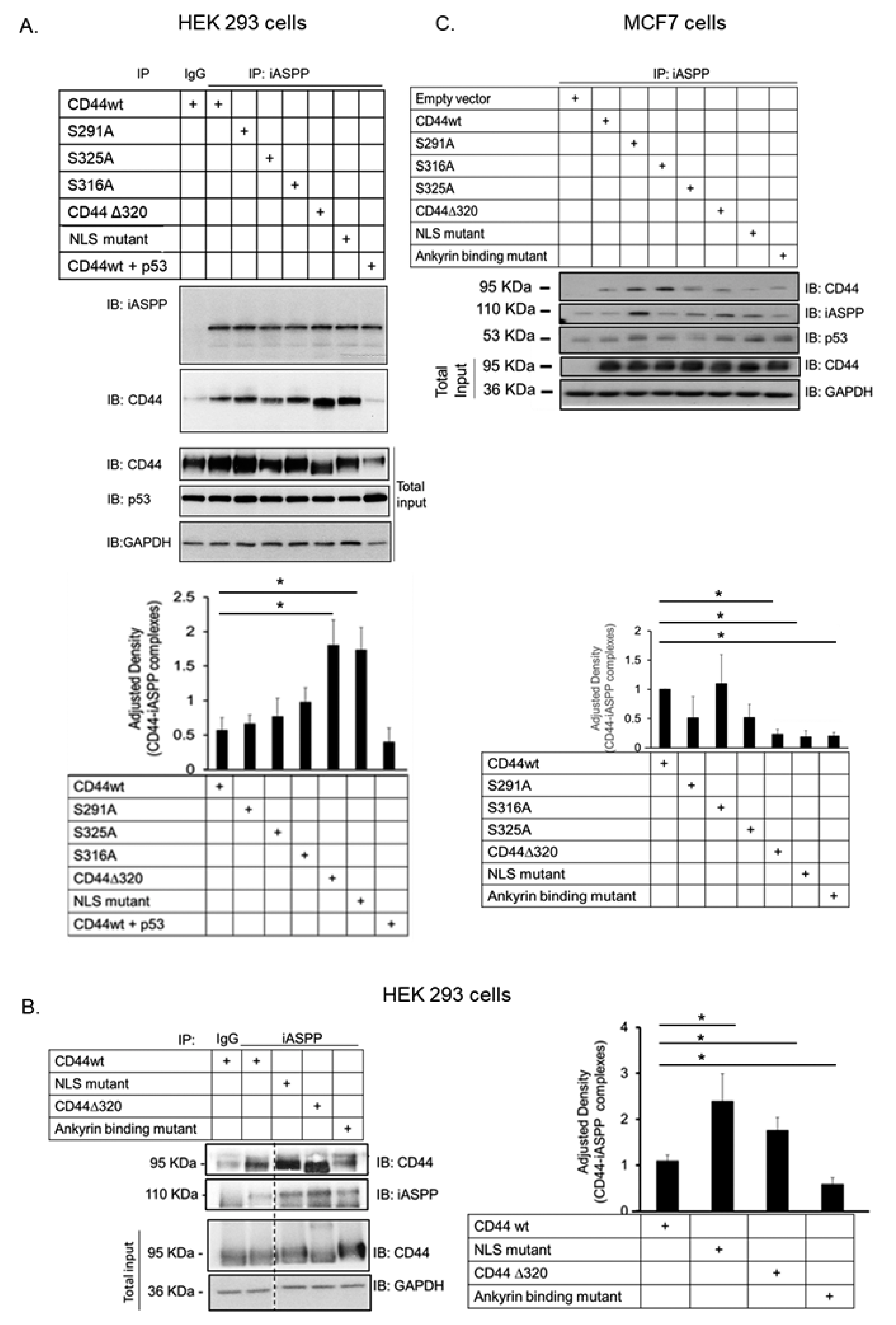
3.3. Characterization of the Epitopes Involved in the Interaction between CD44 and iASPP
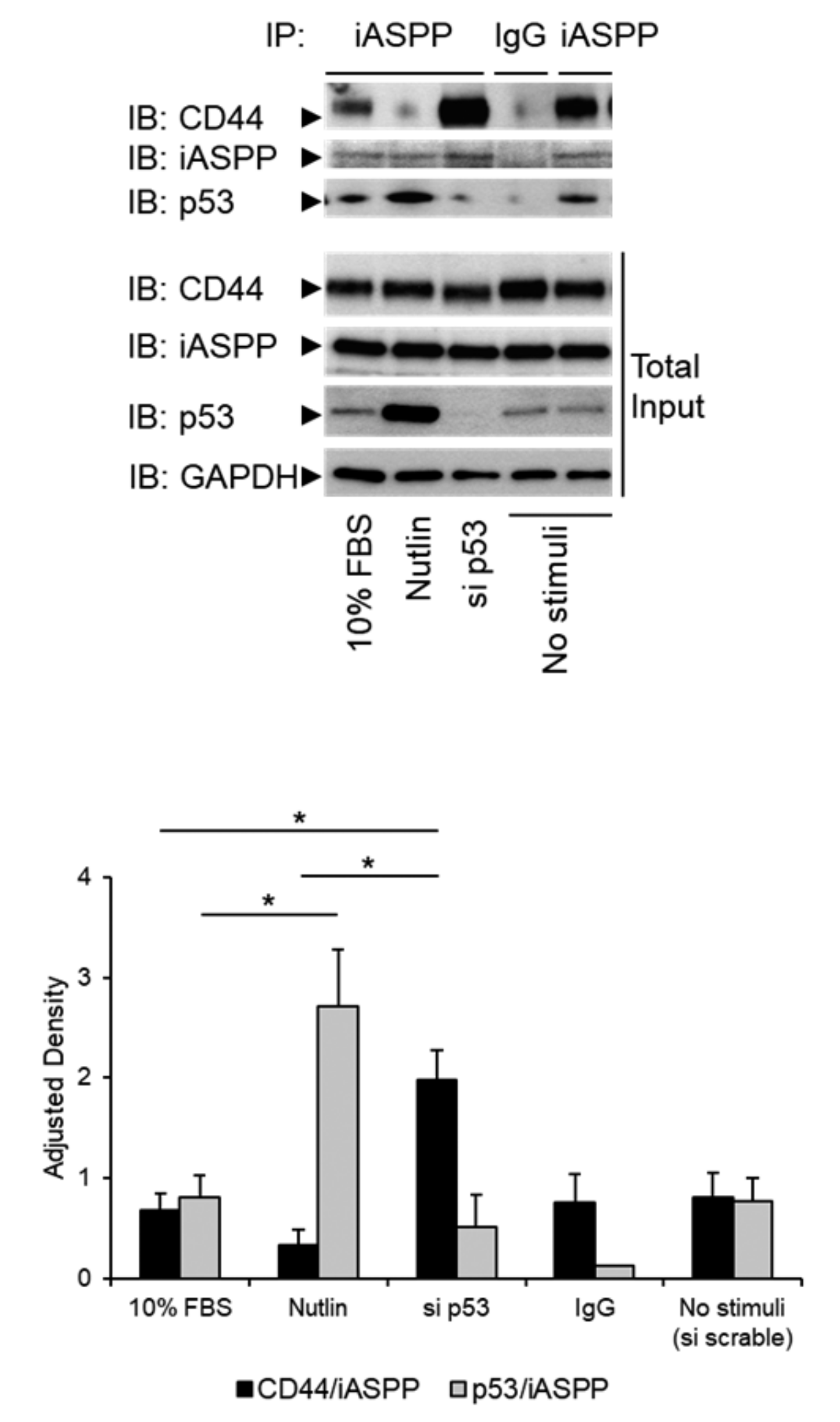
3.4. The Level of p53 in hTERT-BJ Cells Affects the Formation of the iASPP-CD44 Complex
3.5. CD44 Expression Modulates the Sub-Cellular Localization of iASPP-p53 Complexes
3.6. Differential Expression of CD44, iASPP, and p53 in Different Tumor Types
3.7. CD44 Suppresses Cell Growth
3.8. iASPP Is Required for Hyaluronan-Activated CD44-Induced Fibroblast Migration
3.9. p53 Is Required for CD44-Dependent Cell Adhesion
3.10. CD44 Increases p53-Mediated ROS Production
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Laurent, T.C.; Fraser, J.R.E. Hyaluronan. FASEB J. 1992, 6, 2397–2404. [Google Scholar] [CrossRef] [PubMed]
- Toole, B.P. Hyaluronan: From extracellular glue to pericellular cue. Nat. Rev. Cancer 2004, 4, 528–539. [Google Scholar] [CrossRef] [PubMed]
- Toole, B.P. Hyaluronan-CD44 Interactions in Cancer: Paradoxes and Possibilities. Clin. Cancer Res. 2009, 15, 7462–7468. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, N.; Miyoshi, S.; Mikami, T.; Koyama, H.; Kitazawa, M.; Takeoka, M.; Sano, K.; Amano, J.; Isogai, Z.; Niida, S.; et al. Hyaluronan deficiency in tumor stroma impairs macrophage trafficking and tumor neovascularization. Cancer Res. 2010, 70, 7073–7083. [Google Scholar] [CrossRef]
- Heldin, P. Growth Factor Regulation of Hyaluronan Metabolism in Tumor Progression. 2015. Available online: http://www.glycoforum.gr.jp/ (accessed on 12 June 2009).
- Tammi, R.H.; Passi, A.G.; Rilla, K.; Karousou, E.; Vigetti, D.; Makkonen, K.; Tammi, M. Transcriptional and post-translational regulation of hyaluronan synthesis. FEBS J. 2011, 278, 1419–1428. [Google Scholar] [CrossRef]
- Heldin, P.; Basu, K.; Kozlova, I.; Porsch, H. HAS2 and CD44 in Breast Tumorigenesis. Adv. Cancer Res. 2014, 123, 211–229. [Google Scholar] [CrossRef]
- Heldin, P.; Basu, K.; Olofsson, B.; Porsch, H.; Kozlova, I.; Kahata, K. Deregulation of hyaluronan synthesis, degradation and binding promotes breast cancer. J. Biochem. 2013, 154, 395–408. [Google Scholar] [CrossRef]
- Ponta, H.; Sherman, L.; Herrlich, P.A. CD44: From adhesion molecules to signalling regulators. Nat. Rev. Mol. Cell Biol. 2003, 4, 33–45. [Google Scholar] [CrossRef]
- Thorne, R.F.; Legg, J.W.; Isacke, C.M. The role of the CD44 transmembrane and cytoplasmic domains in co-ordinating adhesive and signalling events. J. Cell Sci. 2004, 117, 373–380. [Google Scholar] [CrossRef]
- Hamilton, S.R.; Fard, S.F.; Paiwand, F.F.; Tolg, C.; Veiseh, M.; Wang, C.; McCarthy, J.B.; Bissell, M.J.; Koropatnick, J.; Turley, E.A. The hyaluronan receptors CD44 and Rhamm (CD168) form complexes with ERK1,2 that sustain high basal motility in breast cancer cells. J. Biol. Chem. 2007, 282, 16667–16680. [Google Scholar] [CrossRef]
- Orian-Rousseau, V.; Ponta, H. Adhesion proteins meet receptors: A common theme? Adv. Cancer Res. 2008, 101, 63–92. [Google Scholar] [CrossRef]
- Stoop, R.; Kotani, H.; McNeish, J.D.; Otterness, I.G.; Mikecz, K. Increased resistance to collagen-induced arthritis in CD44-deficient DBA/1 mice. Arthritis Rheum. 2001, 44, 2922–2931. [Google Scholar] [CrossRef]
- Henke, C.; Bitterman, P.; Roongta, U.; Ingbar, D.; Polunovsky, V. Induction of fibroblast apoptosis by anti-CD44 antibody: Implications for the treatment of fibroproliferative lung disease. Am. J. Pathol. 1996, 149, 1639–1650. [Google Scholar]
- Draffin, J.E.; McFarlane, S.; Hill, A.; Johnston, P.G.; Waugh, D.J. CD44 potentiates the adherence of metastatic prostate and breast cancer cells to bone marrow endothelial cells. Cancer Res. 2004, 64, 5702–5711. [Google Scholar] [CrossRef]
- Olofsson, B.; Porsch, H.; Heldin, P. Knock-Down of CD44 Regulates Endothelial Cell Differentiation via NFkappaB-Mediated Chemokine Production. PLoS ONE 2014, 9, e90921. [Google Scholar] [CrossRef]
- Teder, P.; Bergh, J.; Heldin, P. Functional hyaluronan receptors are expressed on squamous cell lung carcinoma cell line but not on other lung carcinoma cell lines. Cancer Res. 1995, 55, 3908–3914. [Google Scholar]
- Kim, Y.; Lee, Y.S.; Choe, J.; Lee, H.; Kim, Y.M.; Jeoung, D. CD44-epidermal growth factor receptor interaction mediates hyaluronic acid-promoted cell motility by activating protein kinase C signaling involving Akt, Rac1, Phox, reactive oxygen species, focal adhesion kinase, and MMP-2. J. Biol. Chem. 2008, 283, 22513–22528. [Google Scholar] [CrossRef]
- Bourguignon, L.Y.; Gilad, E.; Peyrollier, K. Heregulin-mediated ErbB2-ERK signaling activates hyaluronan synthases leading to CD44-dependent ovarian tumor cell growth and migration. J. Biol. Chem. 2007, 282, 19426–19441. [Google Scholar] [CrossRef]
- Sun, C.X.; Robb, V.A.; Gutmann, D.H. Protein 4.1 tumor suppressors: Getting a FERM grip on growth regulation. J. Cell Sci. 2002, 115, 3991–4000. [Google Scholar] [CrossRef]
- Gourlay, C.W.; Ayscough, K.R. The actin cytoskeleton: A key regulator of apoptosis and ageing? Nat. Rev. Mol. Cell Biol. 2005, 6, 583–589. [Google Scholar] [CrossRef]
- Mielgo, A.; van Driel, M.; Bloem, A.; Landmann, L.; Gunthert, U. A novel antiapoptotic mechanism based on interference of Fas signaling by CD44 variant isoforms. Cell Death Differ. 2006, 13, 465–477. [Google Scholar] [CrossRef] [PubMed]
- Fujii, K.; Fujii, Y.; Hubscher, S.; Tanaka, Y. CD44 is the physiological trigger of Fas up-regulation on rheumatoid synovial cells. J. Immunol. 2001, 167, 1198–1203. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Heldin, C.H.; Heldin, P. Inhibition of platelet-derived growth factor-BB-induced receptor activation and fibroblast migration by hyaluronan activation of CD44. J. Biol. Chem. 2006, 281, 26512–26519. [Google Scholar] [CrossRef] [PubMed]
- Skandalis, S.S.; Kozlova, I.; Engstrom, U.; Hellman, U.; Heldin, P. Proteomic identification of CD44 interacting proteins. IUBMB Life 2010, 62, 833–840. [Google Scholar] [CrossRef] [PubMed]
- Kozlova, I.; Ruusala, A.; Voytyuk, O.; Skandalis, S.S.; Heldin, P. IQGAP1 regulates hyaluronan-mediated fibroblast motility and proliferation. Cell Signal. 2012, 24, 1856–1862. [Google Scholar] [CrossRef]
- Bergamaschi, D.; Samuels, Y.; O’Neil, N.J.; Trigiante, G.; Crook, T.; Hsieh, J.K.; O’Connor, D.J.; Zhong, S.; Campargue, I.; Tomlinson, M.L.; et al. iASPP oncoprotein is a key inhibitor of p53 conserved from worm to human. Nat. Genet. 2003, 33, 162–167. [Google Scholar] [CrossRef]
- Sullivan, A.; Lu, X. ASPP: A new family of oncogenes and tumour suppressor genes. Br. J. Cancer 2007, 96, 196–200. [Google Scholar] [CrossRef]
- Yang, J.P.; Hori, M.; Sanda, T.; Okamoto, T. Identification of a novel inhibitor of nuclear factor-kappa B RelA-associated inhibitor. J. Biol. Chem. 1999, 274, 15662–15670. [Google Scholar] [CrossRef]
- Trigiante, G.; Lu, X. ASPPs and cancer. Nat. Rev. Cancer 2006, 6, 217–226. [Google Scholar] [CrossRef]
- Morris, E.V.; Cerundolo, L.; Lu, M.; Verrill, C.; Fritzsche, F.; White, M.J.; Thalmann, G.N.; ten Donkelaar, C.S.; Ratnayaka, I.; Salter, V.; et al. Nuclear iASPP may facilitate prostate cancer progression. Cell Death Dis. 2014, 5, e1492. [Google Scholar] [CrossRef]
- Zhang, X.W.; Wang, M.; Zhou, C.L.; Chen, S.; Wang, J.X. The expression of iASPP in acute leukemias. Leuk. Res. 2005, 29, 179–183. [Google Scholar] [CrossRef]
- Green, D.R.; Kroemer, G. Cytoplasmic functions of the tumour suppressor p53. Nature 2009, 458, 1127–1130. [Google Scholar] [CrossRef]
- Godar, S.; Ince, T.A.; Bell, G.W.; Feldser, D.; Donaher, J.L.; Bergh, J.; Liu, A.; Miu, K.; Watnick, R.S.; Reinhardt, F.; et al. Growth-inhibitory and tumor- suppressive functions of p53 depend on its repression of CD44 expression. Cell 2008, 134, 62–73. [Google Scholar] [CrossRef]
- Mellor, L.; Knudson, C.B.; Hida, D.; Askew, E.B.; Knudson, W. Intracellular Domain Fragment of CD44 Alters CD44 Function in Chondrocytes. J. Biol. Chem. 2013, 288, 25838–25850. [Google Scholar] [CrossRef]
- Papadopoulos, N.; Lennartsson, J.; Heldin, C.H. PDGFR beta translocates to the nucleus and regulates chromatin remodeling via TATA element-modifying factor 1. J. Cell Biol. 2018, 217, 1701–1717. [Google Scholar] [CrossRef]
- Murakami, Y.; Mizuguchi, K. Homology-based prediction of interactions between proteins using Averaged One-Dependence Estimators. BMC Bioinform. 2014, 15, 213. [Google Scholar] [CrossRef]
- Cerami, E.; Gao, J.; Dogrusoz, U.; Gross, B.E.; Sumer, S.O.; Aksoy, B.A. The cBio Cancer Genomics Portal: An Open Platform for Exploring Multidimensional Cancer Genomics Data (vol 2, pg 401, 2012). Cancer Discov. 2012, 2, 960. [Google Scholar] [CrossRef]
- Gao, J.J.; Aksoy, B.A.; Dogrusoz, U.; Dresdner, G.; Gross, B.; Sumer, S.O.; Sun, Y.C.; Jacobsen, A.; Sinha, R.; Larsson, E.; et al. Integrative Analysis of Complex Cancer Genomics and Clinical Profiles Using the cBioPortal. Sci. Signal. 2013, 6, pl1. [Google Scholar] [CrossRef]
- Canisius, S.; Martens, J.W.M.; Wessels, L.F.A. A novel independence test for somatic alterations in cancer shows that biology drives mutual exclusivity but chance explains most co-occurrence. Genome Biol. 2016, 17, 261. [Google Scholar] [CrossRef]
- Gunthert, A.R.; Strater, J.; von Reyher, U.; Henne, C.; Joos, S.; Koretz, K.; Moldenhauer, G.; Krammer, P.H.; Moller, P. Early detachment of colon carcinoma cells during CD95(APO-1/Fas)-mediated apoptosis.1. De-adhesion from hyaluronate by shedding of CD44. J. Cell Biol. 1996, 134, 1089–1096. [Google Scholar] [CrossRef]
- Razavipour, S.F.; Harikumar, K.B.; Slingerland, J.M. p27 as a Transcriptional Regulator: New Roles in Development and Cancer. Cancer Res. 2020, 80, 3451–3458. [Google Scholar] [CrossRef] [PubMed]
- Dimri, G.P.; Lee, X.H.; Basile, G.; Acosta, M.; Scott, C.; Roskelley, C.; Medrano, E.E.; Linskens, M.; Rubelj, I.; Pereirasmith, O.; et al. A Biomarker That Identifies Senescent Human-Cells in Culture and in Aging Skin in-Vivo. Proc. Natl. Acad. Sci. USA 1995, 92, 9363–9367. [Google Scholar] [CrossRef] [PubMed]
- Vendrov, A.E.; Madamanchi, N.R.; Niu, X.L.; Molnar, K.C.; Runge, M.; Szyndralewiez, C.; Page, P.; Runge, M.S. NADPH oxidases regulate CD44 and hyaluronic acid expression in thrombin-treated vascular smooth muscle cells and in atherosclerosis. J. Biol. Chem. 2010, 285, 26545–26557. [Google Scholar] [CrossRef] [PubMed]
- Bergamaschi, D.; Samuels, Y.; Sullivan, A.; Zvelebil, M.; Breyssens, H.; Bisso, A.; Del Sal, G.; Syed, N.; Smith, P.; Gasco, M.; et al. iASPP preferentially binds p53 proline-rich region and modulates apoptotic function of codon 72 polymorphic p53. Nat. Genet. 2006, 38, 1133–1141. [Google Scholar] [CrossRef]
- Ishimoto, T.; Nagano, O.; Yae, T.; Tamada, M.; Motohara, T.; Oshima, H.; Oshima, M.; Ikeda, T.; Asaba, R.; Yagi, H.; et al. CD44 Variant Regulates Redox Status in Cancer Cells by Stabilizing the xCT Subunit of System xc(-) and Thereby Promotes Tumor Growth. Cancer Cell 2011, 19, 387–400. [Google Scholar] [CrossRef]
- Tamada, M.; Nagano, O.; Tateyama, S.; Ohmura, M.; Yae, T.; Ishimoto, T.; Sugihara, E.; Onishi, N.; Yamamoto, T.; Yanagawa, H.; et al. Modulation of Glucose Metabolism by CD44 Contributes to Antioxidant Status and Drug Resistance in Cancer Cells. Cancer Res. 2012, 72, 1438–1448. [Google Scholar] [CrossRef]
- Brown, R.L.; Reinke, L.M.; Damerow, M.S.; Perez, D.; Chodosh, L.A.; Yang, J.; Cheng, C.H. CD44 splice isoform switching in human and mouse epithelium is essential for epithelial-mesenchymal transition and breast cancer progression. J. Clin. Investig. 2011, 121, 1064–1074. [Google Scholar] [CrossRef]
- Freeman, S.A.; Vega, A.; Riedl, M.; Collins, R.F.; Ostrowski, P.P.; Woods, E.C.; Bertozzi, C.R.; Tammi, M.I.; Lidke, D.S.; Johnson, P.; et al. Transmembrane Pickets Connect Cyto- and Pericellular Skeletons Forming Barriers to Receptor Engagement. Cell 2018, 172, 305–317. [Google Scholar] [CrossRef]
- Laska, M.J.; Lowe, S.W.; Zender, L.; Hearn, S.; Vogel, U.; Jensen, U.B.; Bric, A.; Nexo, B.A. Enforced Expression of PPP1R13L Increases Tumorigenesis and Invasion Through p53-Dependent and p53-Independent Mechanisms. Mol. Carcinog. 2009, 48, 832–842. [Google Scholar] [CrossRef]
- Zhang, B.; Xiao, H.J.; Chen, J.; Tao, X.; Cai, L.H. Inhibitory member of the apoptosis-stimulating protein of p53 (ASPP) family promotes growth and tumorigenesis in human p53-deficient prostate cancer cells. Prostate Cancer Prostatic Dis. 2011, 14, 219–224. [Google Scholar] [CrossRef]
- Ge, W.J.; Zhao, K.M.; Wang, X.W.; Li, H.Y.; Yu, M.; He, M.M.; Xue, X.T.; Zhu, Y.F.; Zhang, C.; Cheng, Y.W.; et al. iASPP Is an Antioxidative Factor and Drives Cancer Growth and Drug Resistance by Competing with Nrf2 for Keap1 Binding. Cancer Cell 2017, 32, 561–573.e6. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, C.-Y.; Basu, K.; Ruusala, A.; Kozlova, I.; Li, Y.-S.; Skandalis, S.S.; Heldin, C.-H.; Heldin, P. Hyaluronan-Induced CD44-iASPP Interaction Affects Fibroblast Migration and Survival. Cancers 2023, 15, 1082. https://doi.org/10.3390/cancers15041082
Lin C-Y, Basu K, Ruusala A, Kozlova I, Li Y-S, Skandalis SS, Heldin C-H, Heldin P. Hyaluronan-Induced CD44-iASPP Interaction Affects Fibroblast Migration and Survival. Cancers. 2023; 15(4):1082. https://doi.org/10.3390/cancers15041082
Chicago/Turabian StyleLin, Chun-Yu, Kaustuv Basu, Aino Ruusala, Inna Kozlova, Yan-Shuang Li, Spyridon S. Skandalis, Carl-Henrik Heldin, and Paraskevi Heldin. 2023. "Hyaluronan-Induced CD44-iASPP Interaction Affects Fibroblast Migration and Survival" Cancers 15, no. 4: 1082. https://doi.org/10.3390/cancers15041082
APA StyleLin, C.-Y., Basu, K., Ruusala, A., Kozlova, I., Li, Y.-S., Skandalis, S. S., Heldin, C.-H., & Heldin, P. (2023). Hyaluronan-Induced CD44-iASPP Interaction Affects Fibroblast Migration and Survival. Cancers, 15(4), 1082. https://doi.org/10.3390/cancers15041082