The Role of Pentraxin 3 in Gastrointestinal Cancers



Abstract
:Simple Summary
Abstract
1. Gastrointestinal Cancers
2. Pentraxin Family
3. Role of Pentraxin-3 in GI Cancers
4. Literature Search and Data Extraction
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Arnold, M.; Abnet, C.C.; Neale, R.E.; Vignat, J.; Giovannucci, E.L.; McGlynn, K.A.; Bray, F. Global Burden of 5 Major Types of Gastrointestinal Cancer. Gastroenterology 2020, 159, 335–349.e15. [Google Scholar] [CrossRef] [PubMed]
- Meng, C.; Bai, C.; Brown, T.D.; Hood, L.E.; Tian, Q. Human Gut Microbiota and Gastrointestinal Cancer. Genom. Proteom. Bioinform. 2018, 16, 33–49. [Google Scholar] [CrossRef]
- Ferlay, J.; Ervik, M.; Lam, F.; Colombet, M.; Mery, L.; Piñeros, M.; Znaor, A.; Soerjomataram, I.; Bray, F. Global Cancer Observatory: Cancer Today. Lyon, France: International Agency for Research on Cancer. 2020. Available online: https://gco.iarc.fr/today (accessed on 21 August 2023).
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Ferlay, J.; Colombet, M.; Soerjomataram, I.; Parkin, D.M.; Piñeros, M.; Znaor, A.; Bray, F. Cancer statistics for the year 2020: An overview. Int. J. Cancer 2021, 149, 778–789. [Google Scholar] [CrossRef]
- Patel, S.G.; Karlitz, J.J.; Yen, T.; Lieu, C.H.; Boland, C.R. The rising tide of early-onset colorectal cancer: A comprehensive review of epidemiology, clinical features, biology, risk factors, prevention, and early detection. Lancet Gastroenterol. Hepatol. 2022, 7, 262–274. [Google Scholar] [CrossRef] [PubMed]
- Gewartowska, M.; Stańczyk, M. Nowotwory dziedziczne przewodu pokarmowego. Chir. Po Dyplomie 2019, 14, 14–25. [Google Scholar]
- Tong, Y.; Gao, H.; Qi, Q.; Liu, X.; Li, J.; Gao, J.; Li, P.; Wang, Y.; Du, L.; Wang, C. High fat diet, gut microbiome and gastrointestinal cancer. Theranostics 2021, 11, 5889–5910. [Google Scholar] [CrossRef] [PubMed]
- DiSiena, M.; Perelman, A.; Birk, J.; Rezaizadeh, H. Esophageal Cancer: An Updated Review. South. Med. J. 2021, 114, 161–168. [Google Scholar] [CrossRef]
- Chevallay, M.; Bollschweiler, E.; Chandramohan, S.M.; Schmidt, T.; Koch, O.; Demanzoni, G.; Mönig, S.; Allum, W. Cancer of the gastroesophageal junction: A diagnosis, classification, and management review. Ann. N. Y. Acad Sci. 2018, 1434, 132–138. [Google Scholar] [CrossRef]
- Liu, K.; Zhao, T.; Wang, J.; Chen, Y.; Zhang, R.; Lan, X.; Que, J. Etiology, cancer stem cells and potential diagnostic biomarkers for esophageal cancer. Cancer Lett. 2019, 458, 21–28. [Google Scholar] [CrossRef]
- Smyth, E.C.; Nilsson, M.; Grabsch, H.I.; van Grieken, N.C.; Lordick, F. Gastric cancer. Lancet 2020, 396, 635–648. [Google Scholar] [CrossRef] [PubMed]
- Machlowska, J.; Baj, J.; Sitarz, M.; Maciejewski, R.; Sitarz, R. Gastric Cancer: Epidemiology, Risk Factors, Classification, Genomic Characteristics and Treatment Strategies. Int. J. Mol. Sci. 2020, 21, 4012. [Google Scholar] [CrossRef] [PubMed]
- López, M.J.; Carbajal, J.; Alfaro, A.L.; Saravia, L.G.; Zanabria, D.; Araujo, J.M.; Quispe, L.; Zevallos, A.; Buleje, J.L.; Cho, C.E.; et al. Characteristics of gastric cancer around the world. Crit. Rev. Oncol. 2023, 181, 103841. [Google Scholar] [CrossRef] [PubMed]
- Röcken, C. Predictive biomarkers in gastric cancer. J. Cancer Res. Clin. Oncol. 2023, 149, 467–481. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Wang, C.; Xu, H.; Gao, Y. Aldehyde Dehydrogenase, Liver Disease and Cancer. Int. J. Biol. Sci. 2020, 16, 921–934. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.S.; Zeng, X.F.; Liu, Z.N.; Zhao, Q.H.; Tan, Y.T.; Gao, J.; Li, H.L.; Xiang, Y.B. Diet and liver cancer risk: A narrative review of epidemiological evidence. Br. J. Nutr. 2020, 124, 330–340. [Google Scholar] [CrossRef]
- Llovet, J.M.; Kelley, R.K.; Villanueva, A.; Singal, A.G.; Pikarsky, E.; Roayaie, S.; Lencioni, R.; Koike, K.; Zucman-Rossi, J.; Finn, R.S. Hepatocellular carcinoma. Nat. Rev. Dis. Primers 2021, 7, 6. [Google Scholar] [CrossRef]
- Roa, J.C.; García, P.; Kapoor, V.K.; Maithel, S.K.; Javle, M.; Koshiol, J. Gallbladder cancer. Nat. Rev. Dis. Prim. 2022, 8, 69. [Google Scholar] [CrossRef]
- Schmidt, M.A.; Marcano-Bonilla, L.; Roberts, L.R. Gallbladder cancer: Epidemiology and genetic risk associations. Chin. Clin. Oncol. 2019, 8, 31. [Google Scholar] [CrossRef]
- Baiu, I.; Visser, B. Gallbladder Cancer. JAMA 2018, 320, 1294. [Google Scholar] [CrossRef]
- Lam, R.; Zakko, A.; Petrov, J.C.; Kumar, P.; Duffy, A.J.; Muniraj, T. Gallbladder Disorders: A Comprehensive Review. Disease-a-Month 2021, 67, 101130. [Google Scholar] [CrossRef]
- Wood, L.D.; Canto, M.I.; Jaffee, E.M.; Simeone, D.M. Pancreatic Cancer: Pathogenesis, Screening, Diagnosis, and Treatment. Gastroenterology 2022, 163, 386–402.e1. [Google Scholar] [CrossRef]
- Mizrahi, J.D.; Surana, R.; Valle, J.W.; Shroff, R.T. Pancreatic cancer. Lancet 2020, 395, 2008–2020. [Google Scholar] [CrossRef]
- Khalaf, N.; El-Serag, H.B.; Abrams, H.R.; Thrift, A.P. Burden of Pancreatic Cancer: From Epidemiology to Practice. Clin. Gastroenterol. Hepatol. 2021, 19, 876–884. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Liu, W. Pancreatic Cancer: A Review of Risk Factors, Diagnosis, and Treatment. Technol. Cancer Res. Treat. 2020, 19, 1533033820962117. [Google Scholar] [CrossRef]
- Klein, A.P. Pancreatic cancer epidemiology: Understanding the role of lifestyle and inherited risk factors. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 493–502. [Google Scholar] [CrossRef] [PubMed]
- Kolbeinsson, H.M.; Chandana, S.; Wright, G.P.; Chung, M. Pancreatic Cancer: A Review of Current Treatment and Novel Therapies. J. Investig. Surg. 2023, 36, 2129884. [Google Scholar] [CrossRef] [PubMed]
- Symons, R.; Daly, D.; Gandy, R.; Goldstein, D.; Aghmesheh, M. Progress in the Treatment of Small Intestine Cancer. Curr. Treat. Options Oncol. 2023, 24, 241–261. [Google Scholar] [CrossRef] [PubMed]
- Nagel, G.; Bjørge, T.; Jaensch, A.; Peter, R.S.; Häggström, C.; Lang, A.; Engeland, A.; Teleka, S.; Jirström, K.; Lindquist, D.; et al. Metabolic factors and the risk of small intestine cancers: Pooled study of 800 000 individuals in the metabolic syndrome and cancer project. Int. J. Cancer 2021, 149, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Assumpção, P.; Khayat, A.; Araújo, T.; Barra, W.; Ishak, G.; Cruz, A.; Santos, S.; Demachki, S.; Assumpção, P.; Calcagno, D.; et al. The Small Bowel Cancer Incidence Enigma. Pathol. Oncol. Res. 2020, 26, 635–639. [Google Scholar] [CrossRef] [PubMed]
- Dekker, E.; Tanis, P.J.; Vleugels, J.L.A.; Kasi, P.M.; Wallace, M.B. Colorectal cancer. Lancet 2019, 394, 1467–1480. [Google Scholar] [CrossRef] [PubMed]
- Baidoun, F.; Elshiwy, K.; Elkeraie, Y.; Merjaneh, Z.; Khoudari, G.; Sarmini, M.T.; Gad, M.; Al-Husseini, M.; Saad, A. Colorectal Cancer Epidemiology: Recent Trends and Impact on Outcomes. Curr. Drug Targets 2021, 22, 998–1009. [Google Scholar] [CrossRef] [PubMed]
- Sninsky, J.A.; Shore, B.M.; Lupu, G.V.; Crockett, S.D. Risk Factors for Colorectal Polyps and Cancer. Gastrointest. Endosc. Clin. N. Am. 2022, 32, 195–213. [Google Scholar] [CrossRef] [PubMed]
- Young, A.N.; Jacob, E.; Willauer, P.; Smucker, L.; Monzon, R.; Oceguera, L. Anal Cancer. Surg. Clin. N. Am. 2020, 100, 629–634. [Google Scholar] [CrossRef] [PubMed]
- Gondal, T.A.; Chaudhary, N.; Bajwa, H.; Rauf, A.; Le, D.; Ahmed, S. Anal Cancer: The Past, Present and Future. Curr. Oncol. 2023, 30, 3232–3250. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Wang, X.; Zou, H.; Dai, Z.; Feng, S.; Zhang, M.; Xiao, G.; Liu, Z.; Cheng, Q. The Basic Characteristics of the Pentraxin Family and Their Functions in Tumor Progression. Front. Immunol. 2020, 11, 1757. [Google Scholar] [CrossRef]
- Armstrong, P.B. Comparative biology of the pentraxin protein family: Evolutionarily conserved component of innate immune system. Int. Rev. Cell Mol. Biol. 2015, 316, 1–47. [Google Scholar] [CrossRef]
- Pathak, A.; Agrawal, A. Evolution of C-Reactive Protein. Front. Immunol. 2019, 10, 943. [Google Scholar] [CrossRef]
- Dyer, E.M.; Waterfield, T.; Baynes, H. How to use C-reactive protein. Arch. Dis. Child. Educ. Pract. Ed. 2018, 104, 150–153. [Google Scholar] [CrossRef]
- Noushad, S.; Ahmed, S.; Ansari, B.; Mustafa, U.H.; Saleem, Y.; Hazrat, H. Physiological biomarkers of chronic stress: A systematic review. Int. J. Health Sci. 2021, 15, 46–59. [Google Scholar]
- Ponti, G.; Maccaferri, M.; Ruini, C.; Tomasi, A.; Ozben, T. Biomarkers associated with COVID-19 disease progression. Crit. Rev. Clin. Lab. Sci. 2020, 57, 389–399. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.; Huang, Y.; Shi, F.; Tan, K.; Ma, Q.; Chen, Y.; Jiang, X.; Li, X. C-reactive protein correlates with computed tomographic findings and predicts severe COVID-19 early. J. Med. Virol. 2020, 92, 856–862. [Google Scholar] [CrossRef] [PubMed]
- Xi, D.; Luo, T.; Xiong, H.; Liu, J.; Lu, H.; Li, M.; Hou, Y.; Guo, Z. SAP: Structure, function, and its roles in immune-related diseases. Int. J. Cardiol. 2015, 187, 20–26. [Google Scholar] [CrossRef]
- Pilling, D.; Gomer, R.H. The Development of Serum Amyloid P as a Possible Therapeutic. Front. Immunol. 2018, 9, 2328. [Google Scholar] [CrossRef] [PubMed]
- Bogdan, M.; Meca, A.-D.; Turcu-Stiolica, A.; Oancea, C.N.; Kostici, R.; Surlin, M.V.; Florescu, C. Insights into the Relationship between Pentraxin-3 and Cancer. Int. J. Mol. Sci. 2022, 23, 15302. [Google Scholar] [CrossRef]
- Bonavita, E.; Gentile, S.; Rubino, M.; Maina, V.; Papait, R.; Kunderfranco, P.; Greco, C.; Feruglio, F.; Molgora, M.; Laface, I.; et al. PTX3 Is an Extrinsic Oncosuppressor Regulating Complement-Dependent Inflammation in Cancer. Cell 2015, 160, 700–714. [Google Scholar] [CrossRef] [PubMed]
- Chiari, D.; Pirali, B.; Perano, V.; Leone, R.; Mantovani, A.; Bottazzi, B. The crossroad between autoimmune disorder, tissue remodeling and cancer of the thyroid: The long pentraxin 3 (PTX3). Front. Endocrinol. 2023, 14, 1146017. [Google Scholar] [CrossRef]
- Brunetta, E.; Folci, M.; Bottazzi, B.; De Santis, M.; Gritti, G.; Protti, A.; Mapelli, S.N.; Bonovas, S.; Piovani, D.; Leone, R.; et al. Macrophage expression and prognostic significance of the long pentraxin PTX3 in COVID-19. Nat. Immunol. 2021, 22, 19–24. [Google Scholar] [CrossRef]
- Mantovani, A.; Morrone, M.C.; Patrono, C.; Santoro, M.G.; Schiaffino, S.; Remuzzi, G.; Bussolati, G.; Cappuccinelli, P. Long Covid: Where we stand and challenges ahead. Cell Death Differ. 2022, 29, 1891–1900. [Google Scholar] [CrossRef]
- Chi, J.-Y.; Hsiao, Y.-W.; Li, C.-F.; Lo, Y.-C.; Lin, Z.-Y.; Hong, J.-Y.; Liu, Y.-M.; Han, X.; Wang, S.-M.; Chen, B.-K.; et al. Targeting chemotherapy-induced PTX3 in tumor stroma to prevent the progression of drug-resistant cancers. Oncotarget 2015, 6, 23987–24001. [Google Scholar] [CrossRef]
- Alessi, P.; Leali, D.; Camozzi, M.; Cantelmo, A.; Albini, A.; Presta, M. Anti-FGF2 approaches as a strategy to compensate resistance to anti-VEGF therapy: Long-pentraxin 3 as a novel antiangiogenic FGF2-antagonist. Eur. Cytokine Netw. 2009, 20, 225–234. [Google Scholar] [CrossRef]
- de la Torre, Y.M.; Fabbri, M.; Jaillon, S.; Bastone, A.; Nebuloni, M.; Vecchi, A.; Mantovani, A.; Garlanda, C. Evolution of the Pentraxin Family: The New Entry PTX4. J. Immunol. 2010, 184, 5055–5064. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, S.; Liu, J.; Zhang, Y.; Zhang, N.; Cheng, Q.; Zhang, H.; Wu, X. The pentraxin family in autoimmune disease. Clin. Chim. Acta 2023, 551, 117592. [Google Scholar] [CrossRef]
- Oggioni, M.; Mercurio, D.; Minuta, D.; Fumagalli, S.; Popiolek-Barczyk, K.; Sironi, M.; Ciechanowska, A.; Ippati, S.; De Blasio, D.; Perego, C.; et al. Long pentraxin PTX3 is upregulated systemically and centrally after experimental neurotrauma, but its depletion leaves unaltered sensorimotor deficits or histopathology. Sci. Rep. 2021, 11, 9616. [Google Scholar] [CrossRef]
- Liu, H.; Kong, X.; Chen, F. Mkrn3 functions as a novel ubiquitin E3 ligase to inhibit Nptx1 during puberty initiation. Oncotarget 2017, 8, 85102–85109. [Google Scholar] [CrossRef]
- Manchia, M.; Piras, I.S.; Huentelman, M.J.; Pinna, F.; Zai, C.C.; Kennedy, J.L.; Carpiniello, B. Pattern of gene expression in different stages of schizophrenia: Down-regulation of NPTX2 gene revealed by a meta-analysis of microarray datasets. Eur. Neuropsychopharmacol. 2017, 27, 1054–1063. [Google Scholar] [CrossRef]
- Begcevic, I.; Tsolaki, M.; Brinc, D.; Brown, M.; Martinez-Morillo, E.; Lazarou, I.; Kozori, M.; Tagaraki, F.; Nenopoulou, S.; Gkioka, M.; et al. Neuronal pentraxin receptor-1 is a new cerebrospinal fluid biomarker of Alzheimer’s disease progression. F1000Research 2018, 7, 1012. [Google Scholar] [CrossRef]
- Goetzl, E.J.; Abner, E.L.; Jicha, G.A.; Kapogiannis, D.; Schwartz, J.B. Declining levels of functionally specialized synaptic proteins in plasma neuronal exosomes with progression of Alzheimer’s disease. FASEB J. 2018, 32, 888–893. [Google Scholar] [CrossRef]
- Dulewicz, M.; Kulczyńska-Przybik, A.; Słowik, A.; Borawska, R.; Mroczko, B. Neurogranin and Neuronal Pentraxin Receptor as Synaptic Dysfunction Biomarkers in Alzheimer’s Disease. J. Clin. Med. 2021, 10, 4575. [Google Scholar] [CrossRef]
- Kulczyńska-Przybik, A.; Dulewicz, M.; Doroszkiewicz, J.; Borawska, R.; Słowik, A.; Zetterberg, H.; Hanrieder, J.; Blennow, K.; Mroczko, B. The Relationships between Cerebrospinal Fluid Glial (CXCL12, CX3CL, YKL-40) and Synaptic Biomarkers (Ng, NPTXR) in Early Alzheimer’s Disease. Int. J. Mol. Sci. 2023, 24, 13166. [Google Scholar] [CrossRef]
- Li, D.; Hao, Z.; Nan, Y.; Chen, Y. Role of long pentraxin PTX3 in cancer. Clin. Exp. Med. 2023, 1–11. [Google Scholar] [CrossRef]
- Presta, M.; Foglio, E.; Schuind, A.C.; Ronca, R. Long Pentraxin-3 Modulates the Angiogenic Activity of Fibroblast Growth Factor-2. Front. Immunol. 2018, 9, 2327. [Google Scholar] [CrossRef]
- Liu, B.; Zhao, Y.; Guo, L. Increased serum pentraxin-3 level predicts poor prognosis in patients with colorectal cancer after curative surgery, a cohort study. Medicine 2018, 97, e11780. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, T.-Y.; Niu, X.-C. Increased Plasma Levels of Pentraxin 3 Are Associated with Poor Prognosis of Colorectal Carcinoma Patients. Tohoku J. Exp. Med. 2016, 240, 39–46. [Google Scholar] [CrossRef]
- Di Caro, G.; Carvello, M.; Pesce, S.; Erreni, M.; Marchesi, F.; Todoric, J.; Sacchi, M.; Montorsi, M.; Allavena, P.; Spinelli, A. Circulating Inflammatory Mediators as Potential Prognostic Markers of Human Colorectal Cancer. PLoS ONE 2016, 11, e0148186. [Google Scholar] [CrossRef]
- Papila, K.; Sozer, V.; Cigdem, K.; Durmus, S.; Kurtulus, D.; Papila, C.; Gelisgen, R.; Uzun, H. Circulating nuclear factor-kappa B mediates cancer-associated inflammation in human breast and colon cancer. J. Med. Biochem. 2021, 40, 150–159. [Google Scholar] [CrossRef]
- Rubino, M.; Kunderfranco, P.; Basso, G.; Greco, C.M.; Pasqualini, F.; Serio, S.; Roncalli, M.; Laghi, L.; Mantovani, A.; Papait, R.; et al. Epigenetic regulation of the extrinsic oncosuppressor PTX3 gene in inflammation and cancer. OncoImmunology 2017, 6, e1333215. [Google Scholar] [CrossRef]
- An, S.; Kim, S.-K.; Kwon, H.Y.; Kim, C.S.; Bang, H.-J.; Do, H.; Kim, B.; Kim, K.; Kim, Y. Expression of Immune-Related and Inflammatory Markers and Their Prognostic Impact in Colorectal Cancer Patients. Int. J. Mol. Sci. 2023, 24, 11579. [Google Scholar] [CrossRef]
- Chen, D.; Wang, R.; Long, M.; Li, W.; Xiao, B.; Deng, H.; Weng, K.; Gong, D.; Liu, F.; Luo, S.; et al. Identification of in vitro and in vivo oncolytic effect in colorectal cancer cells by Orf virus strain NA1/11. Oncol. Rep. 2021, 45, 535–546. [Google Scholar] [CrossRef]
- Mathieu, V.; Laguera, B.; Masi, M.; Dulanto, S.A.; Bingham, T.W.; Hernandez, L.W.; Sarlah, D.; Evidente, A.; Lafontaine, D.L.J.; Kornienko, A.; et al. Amaryllidaceae Alkaloids Decrease the Proliferation, Invasion, and Secretion of Clinically Relevant Cytokines by Cultured Human Colon Cancer Cells. Biomolecules 2022, 12, 1267. [Google Scholar] [CrossRef]
- Deng, H.; Fan, X.; Wang, X.; Zeng, L.; Zhang, K.; Zhang, X.; Li, N.; Han, Q.; Lv, Y.; Liu, Z. Serum pentraxin 3 as a biomarker of hepatocellular carcinoma in chronic hepatitis B virus infection. Sci. Rep. 2020, 10, 20276. [Google Scholar] [CrossRef]
- Mehanna, R.G.; Mohiedeen, K.; Kassem, M.; Deghady, A.; Abouelkheir, H. Assessment of serum CXCL9 and pentraxin 3 as novel markers for hepatocellular carcinoma in cirrhotic hepatitis C patients. Clin. Exp. Hepatol. 2023, 9, 14–20. [Google Scholar] [CrossRef]
- Carmo, R.F.; Aroucha, D.; Vasconcelos, L.R.S.; Pereira, L.; Moura, P.; Cavalcanti, M. Genetic variation in PTX3 and plasma levels associated with hepatocellular carcinoma in patients with HCV. J. Viral Hepat. 2015, 23, 116–122. [Google Scholar] [CrossRef]
- Song, T.; Wang, C.; Guo, C.; Liu, Q.; Zheng, X. Pentraxin 3 overexpression accelerated tumor metastasis and indicated poor prognosis in hepatocellular carcinoma via driving epithelial-mesenchymal transition. J. Cancer 2018, 9, 2650–2658. [Google Scholar] [CrossRef]
- Cabiati, M.; Gaggini, M.; De Simone, P.; Del Ry, S. Do pentraxin 3 and neural pentraxin 2 have different facet function in hepatocellular carcinoma? Clin. Exp. Med. 2021, 21, 555–562. [Google Scholar] [CrossRef]
- Feder, S.; Haberl, E.M.; Spirk, M.; Weiss, T.S.; Wiest, R.; Buechler, C. Pentraxin-3 is not related to disease severity in cirrhosis and hepatocellular carcinoma patients. Clin. Exp. Med. 2020, 20, 289–297. [Google Scholar] [CrossRef]
- Fan, Z.; Zheng, Y.; Li, X.; Deng, X.; Ba, Y.; Feng, K.; Su, J.; Wang, H.; Suo, Z.; Li, L. Promoting role of pentraxin-3 in esophageal squamous cell carcinoma. Mol. Ther.-Oncolytics 2022, 24, 772–787. [Google Scholar] [CrossRef]
- Ma, D.; Zong, Y.; Zhu, S.-T.; Wang, Y.-J.; Li, P.; Zhang, S.-T. Inhibitory Role of Pentraxin-3 in Esophageal Squamous Cell Carcinoma. Chin. Med. J. 2016, 129, 2233–2240. [Google Scholar] [CrossRef]
- Zhao, S.; Jiang, Y.; Tian, T.; Zhao, J.; Xie, Y.; Chen, X.; Lu, J.; Yang, F.; Li, H.; Liu, K.; et al. Jie Du Tong Ye San Prevents N-Nitrosomethylbenzylamine-Induced Esophageal Carcinogenesis via Inhibition of Inflammation and Proliferation. Evidence-Based Complement. Altern. Med. 2019, 2019, 5752670. [Google Scholar] [CrossRef]
- Peiffer, D.S.; Zimmerman, N.P.; Wang, L.-S.; Ransom, B.W.; Carmella, S.G.; Kuo, C.-T.; Siddiqui, J.; Chen, J.-H.; Oshima, K.; Huang, Y.-W.; et al. Chemoprevention of Esophageal Cancer with Black Raspberries, Their Component Anthocyanins, and a Major Anthocyanin Metabolite, Protocatechuic Acid. Cancer Prev. Res. 2014, 7, 574–584. [Google Scholar] [CrossRef]
- Cui, X.; Qin, T.; Zhao, Z.; Yang, G.; Sanches, J.G.P.; Zhang, Q.; Fan, S.; Cao, L.; Hu, X. Pentraxin-3 inhibits milky spots metastasis of gastric cancer by inhibiting M2 macrophage polarization. J. Cancer 2021, 12, 4686–4697. [Google Scholar] [CrossRef]
- Yeni, M.; Korkut, E.; Aksungur, N.; Kara, S.; Askin, S.; Kartal, M. Determination of Pentraxin-3, Interleukin-8 and Vascular Endothelial Growth Factor Levels in Patients with Gastric Adenocarcinoma. Asian Pac. J. Cancer Prev. 2021, 22, 1507–1512. [Google Scholar] [CrossRef]
- Choi, B.; Lee, E.-J.; Park, Y.S.; Kim, S.-M.; Kim, E.-Y.; Song, Y.; Kang, S.-W.; Rhu, M.-H.; Chang, E.-J. Pentraxin-3 Silencing Suppresses Gastric Cancer-related Inflammation by Inhibiting Chemotactic Migration of Macrophages. Anticancer Res. 2015, 35, 2663–2668. [Google Scholar] [PubMed]
- Choi, B.; Lee, E.-J.; Shin, M.-K.; Park, Y.S.; Ryu, M.-H.; Kim, S.-M.; Kim, E.-Y.; Lee, H.K.; Chang, E.-J. Upregulation of brain-derived neurotrophic factor in advanced gastric cancer contributes to bone metastatic osteolysis by inducing long pentraxin. Oncotarget 2016, 7, 55506–55517. [Google Scholar] [CrossRef] [PubMed]
- Goulart, M.R.; Watt, J.; Siddiqui, I.; Lawlor, R.T.; Imrali, A.; Hughes, C.; Saad, A.; ChinAleong, J.; Hurt, C.; Cox, C.; et al. Pentraxin 3 is a stromally-derived biomarker for detection of pancreatic ductal adenocarcinoma. npj Precis. Oncol. 2021, 5, 61. [Google Scholar] [CrossRef] [PubMed]
- Kamal, M.A.; Siddiqui, I.; Belgiovine, C.; Barbagallo, M.; Paleari, V.; Pistillo, D.; Chiabrando, C.; Schiarea, S.; Bottazzi, B.; Leone, R.; et al. Oncogenic KRAS-Induced Protein Signature in the Tumor Secretome Identifies Laminin-C2 and Pentraxin-3 as Useful Biomarkers for the Early Diagnosis of Pancreatic Cancer. Cancers 2022, 14, 2653. [Google Scholar] [CrossRef]
- Kocher, H.M.; Basu, B.; Froeling, F.E.M.; Sarker, D.; Slater, S.; Carlin, D.; Desouza, N.M.; De Paepe, K.N.; Goulart, M.R.; Hughes, C.; et al. Phase I clinical trial repurposing all-trans retinoic acid as a stromal targeting agent for pancreatic cancer. Nat. Commun. 2020, 11, 4841. [Google Scholar] [CrossRef]
- Kondo, S.; Ueno, H.; Hosoi, H.; Hashimoto, J.; Morizane, C.; Koizumi, F.; Tamura, K.; Okusaka, T. Clinical impact of pentraxin family expression on prognosis of pancreatic carcinoma. Br. J. Cancer 2013, 109, 739–746. [Google Scholar] [CrossRef]
- Rosendahl, A.H.; Gundewar, C.; Hilmersson, K.S.; Ni, L.; Saleem, M.A.; Andersson, R. Conditionally immortalized human pancreatic stellate cell lines demonstrate enhanced proliferation and migration in response to IGF-I. Exp. Cell Res. 2015, 330, 300–310. [Google Scholar] [CrossRef]
- A McCarthy, G.; Di Niro, R.; Finan, J.M.; Jain, A.; Guo, Y.; Wyatt, C.R.; Guimaraes, A.R.; A Waugh, T.; Keith, D.; Morgan, T.K.; et al. Deletion of the mRNA stability factor ELAVL1 (HuR) in pancreatic cancer cells disrupts the tumor microenvironment integrity. NAR Cancer 2023, 5, zcad016. [Google Scholar] [CrossRef]
- Sato, K.; Hikita, H.; Shigekawa, M.; Kato, S.; Sasaki, Y.; Shinkai, K.; Fukuoka, M.; Kudo, S.; Sato, Y.; Fukumoto, K.; et al. Pentraxin 3 is an adipose tissue-related serum marker for pancreatic cancer cachexia predicting subsequent muscle mass and visceral fat loss. Cancer Sci. 2022, 113, 4311–4326. [Google Scholar] [CrossRef] [PubMed]
- Page, M.J.; E McKenzie, J.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; A Akl, E.; E Brennan, S.; et al. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. BMJ 2021, 372, n71. [Google Scholar] [CrossRef] [PubMed]
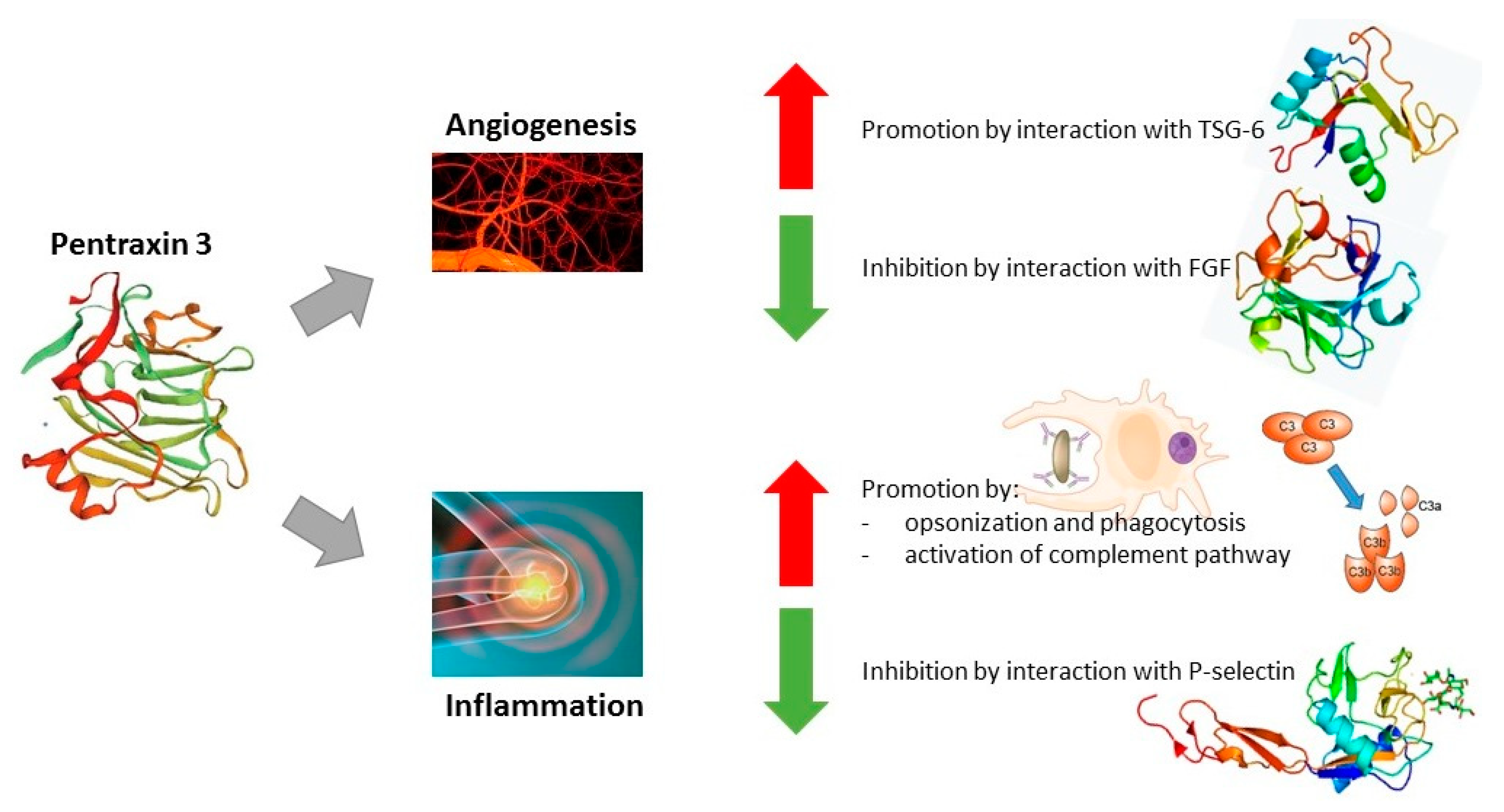
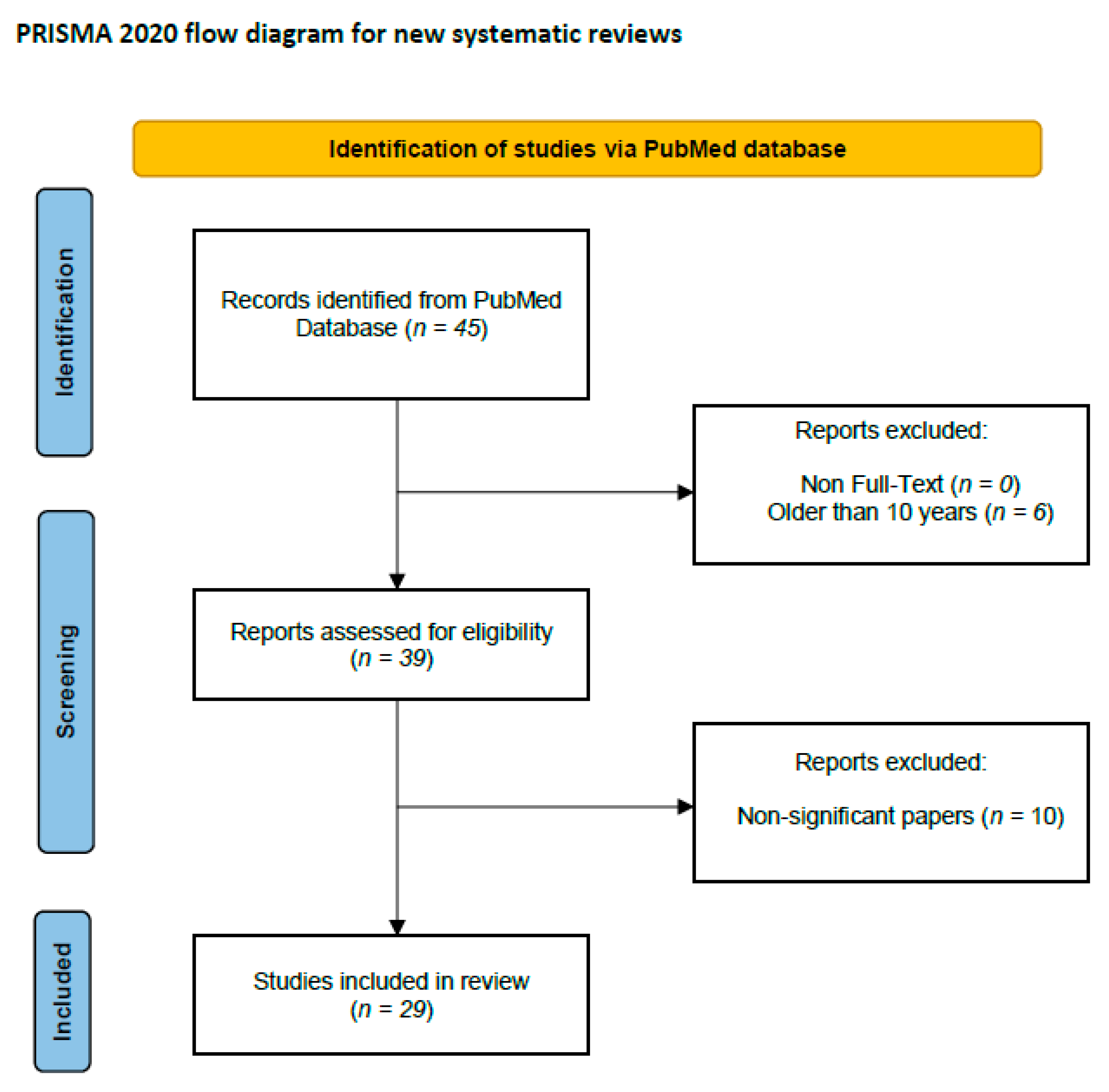

{kind=link}
{kind=link}
| Material/Cancer Type | Serum/Plasma | Tissue | Cell Line |
|---|---|---|---|
| Colorectal cancer | ↑ | n/s | ↑ |
| Liver cancer | ↑ | ↑ | - |
| Esophageal cancer | ↓ | ↓ | ↑ |
| Pancreatic cancer | ↑ | ↑ | ↑ |
| Gastric cancer | ↓/↑ | - | ↑ |
| Entry | Number of Available Publications | Number of Publications from Last 10 Years with Full Text | Number of Irrelevant Publications | Number of Publications Taken into Analysis |
|---|---|---|---|---|
| (colorectal/colon cancer) AND (pentraxin 3) | 10 | 10 | 1 (irrelevant) | 9 |
| (liver cancer) AND (pentraxin 3) | 17 | 13 | 7 (irrelevant) | 6 |
| (esophageal cancer) AND (pentraxin 3) | 5 | 4 | 0 | 4 |
| (pancreatic cancer) AND (pentraxin 3) | 8 | 7 | 1 (retracted) | 6 |
| (gastric cancer) AND (pentraxin 3) | 5 | 5 | 1 (retracted) | 4 |
| (gallbladder cancer) AND (pentraxin 3) | 0 | 0 | 0 | 0 |
| (anus cancer) AND (pentraxin 3) | 0 | 0 | 0 | 0 |
| (small intestine cancer) AND (pentraxin 3) | 0 | 0 | 0 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zajkowska, M.; Mroczko, B. The Role of Pentraxin 3 in Gastrointestinal Cancers. Cancers 2023, 15, 5832. https://doi.org/10.3390/cancers15245832
Zajkowska M, Mroczko B. The Role of Pentraxin 3 in Gastrointestinal Cancers. Cancers. 2023; 15(24):5832. https://doi.org/10.3390/cancers15245832
Chicago/Turabian StyleZajkowska, Monika, and Barbara Mroczko. 2023. "The Role of Pentraxin 3 in Gastrointestinal Cancers" Cancers 15, no. 24: 5832. https://doi.org/10.3390/cancers15245832
APA StyleZajkowska, M., & Mroczko, B. (2023). The Role of Pentraxin 3 in Gastrointestinal Cancers. Cancers, 15(24), 5832. https://doi.org/10.3390/cancers15245832