

Circular RNA from Tyrosylprotein Sulfotransferase 2 Gene Inhibits Cisplatin Sensitivity in Head and Neck Squamous Cell Carcinoma by Sponging miR-770-5p and Interacting with Nucleolin
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Patients and Follow-Up
2.2. CircRNA Microarray Analysis
2.3. Cell Cultures
2.4. Transfection Assay
2.5. Stable Cell Line Generation
2.6. Flow Cytometry
2.7. RNA Isolation and qRT–PCR
2.8. RNase R Digestion Treatment and Sanger Sequencing
2.9. CCK8 Experiment
2.10. Fluorescence In Situ Hybridization (FISH) Assay
2.11. RNA Pulldown Assay
2.12. Rapid Silver Staining Experiment for Mass Spectrometry
2.13. Western Blot Analysis
2.14. Immunohistochemistry
2.15. Statistical Analysis
3. Results
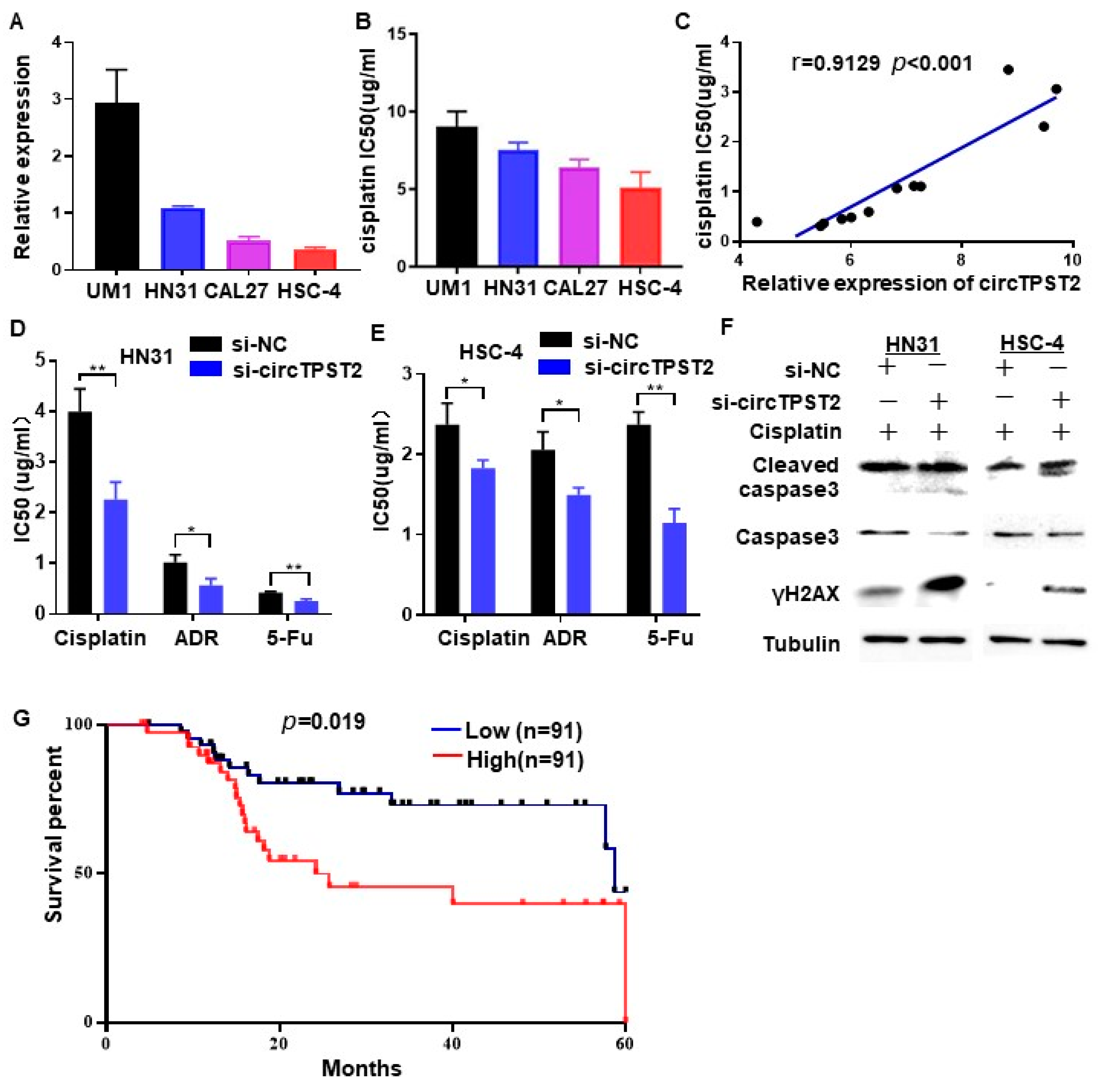
3.1. CircTPST2 Was Identified as a Novel Highly Expressed circRNA in HNSCC
3.2. The Expression Level of circTPST2 Is Negatively Correlated with the Cisplatin Sensitivity of HNSCC Cells
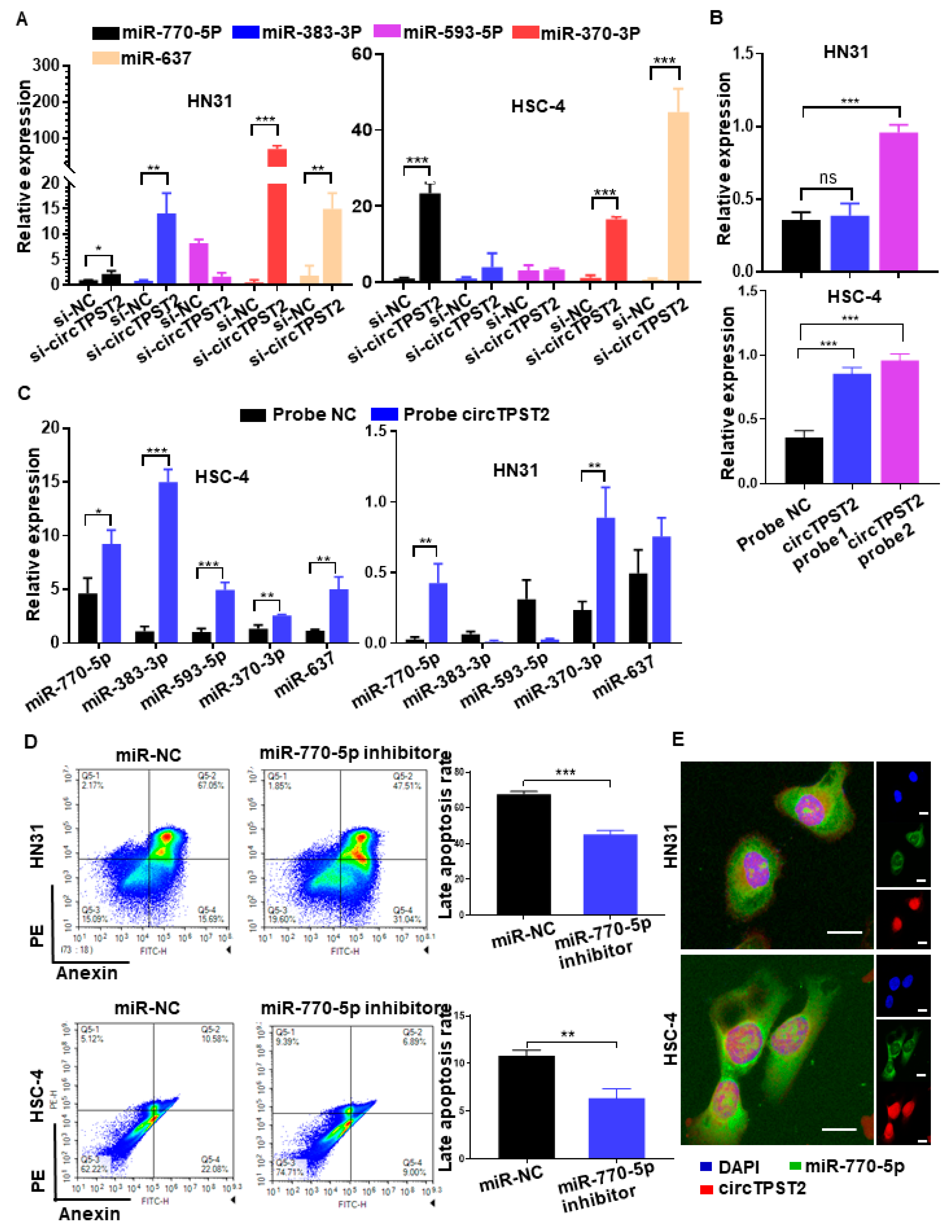
3.3. CircTPST2 Functions as a Sponge for miR-770-5p in HNSCC
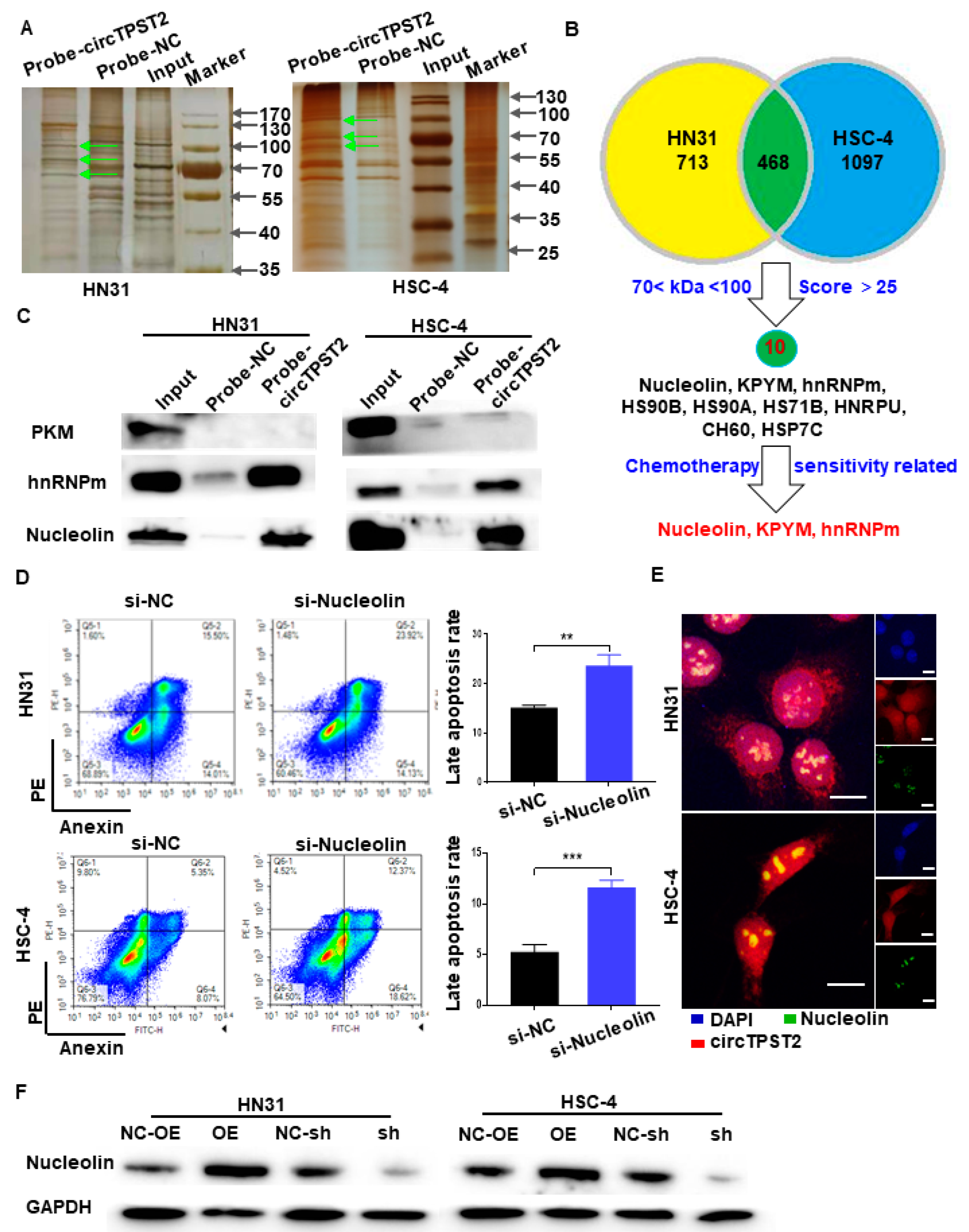
3.4. CircTPST2 Physically Interacts with the Nucleolin Protein in HNSCC
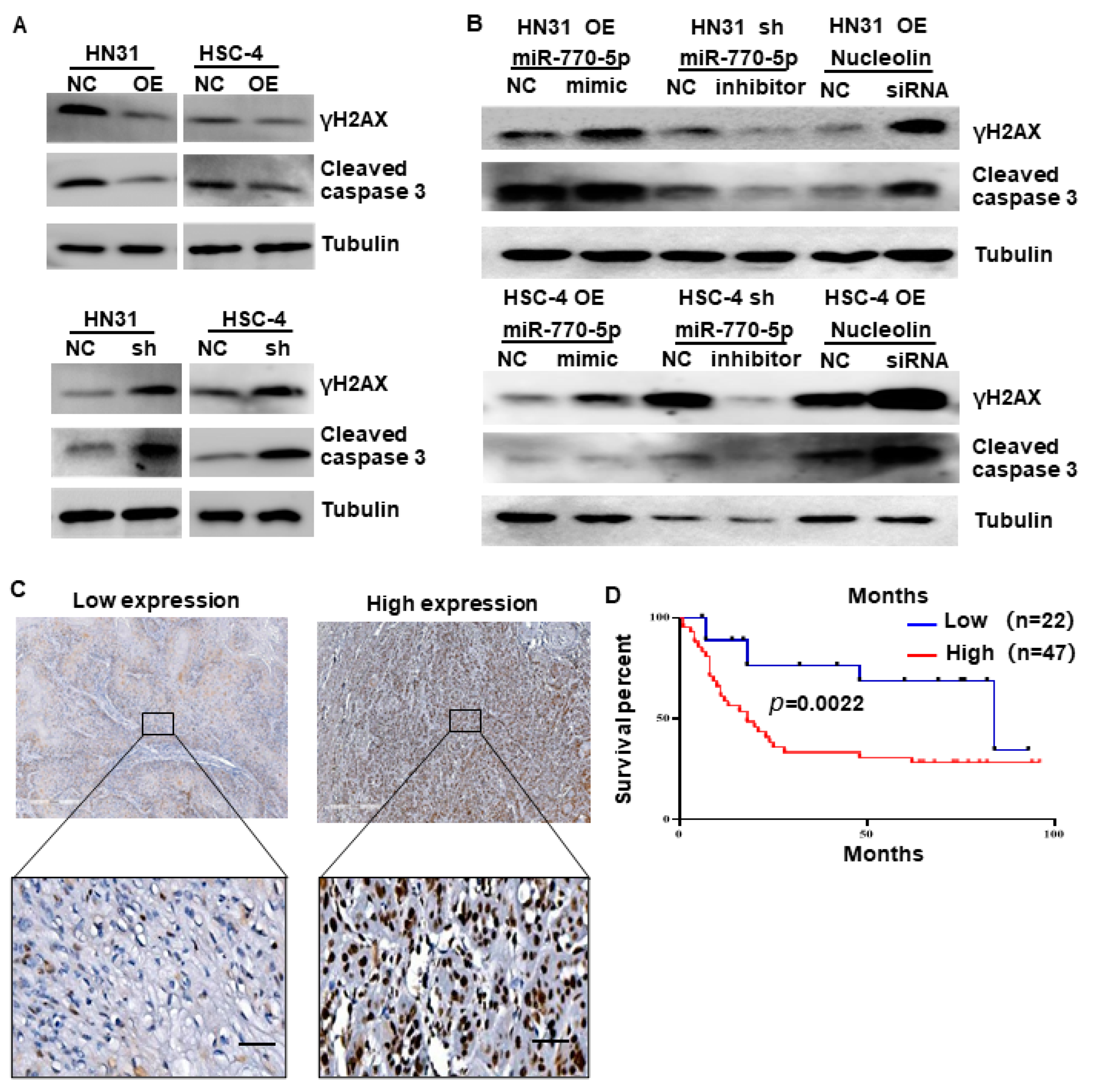
3.5. CircTPST2 Inhibits Cisplatin Sensitivity by Sponging miR-770-5p and Interacting with Nucleolin
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Bach, D.-H.; Lee, S.K.; Sood, A.K. Circular RNAs in Cancer. Mol. Ther. Nucleic Acids 2019, 16, 118–129. [Google Scholar] [CrossRef]
- Cramer, J.D.; Burtness, B.; Le, Q.T.; Ferris, R.L. The Changing Therapeutic Landscape of Head and Neck Cancer. Nat. Rev. Clin. Oncol. 2019, 16, 669–683. [Google Scholar] [CrossRef] [PubMed]
- Johnson, D.E.; Burtness, B.; Leemans, C.R.; Lui, V.W.Y.; Bauman, J.E.; Grandis, J.R. Head and Neck Squamous Cell Carcinoma. Nat. Rev. Dis. Primer 2020, 6, 92. [Google Scholar] [CrossRef]
- Holohan, C.; Van Schaeybroeck, S.; Longley, D.B.; Johnston, P.G. Cancer Drug Resistance: An Evolving Paradigm. Nat. Rev. Cancer 2013, 13, 714–726. [Google Scholar] [CrossRef] [PubMed]
- Senthebane, D.A.; Rowe, A.; Thomford, N.E.; Shipanga, H.; Munro, D.; Mazeedi, M.A.M.A.; Almazyadi, H.A.M.; Kallmeyer, K.; Dandara, C.; Pepper, M.S.; et al. The Role of Tumor Microenvironment in Chemoresistance: To Survive, Keep Your Enemies Closer. Int. J. Mol. Sci. 2017, 18, 1586. [Google Scholar] [CrossRef]
- Barrett, S.P.; Salzman, J. Circular RNAs: Analysis, Expression and Potential Functions. Development 2016, 143, 1838–1847. [Google Scholar] [CrossRef]
- Geng, Y.; Jiang, J.; Wu, C. Function and Clinical Significance of circRNAs in Solid Tumors. J. Hematol. Oncol. 2018, 11, 98. [Google Scholar] [CrossRef]
- Cui, C.; Yang, J.; Li, X.; Liu, D.; Fu, L.; Wang, X. Functions and Mechanisms of Circular RNAs in Cancer Radiotherapy and Chemotherapy Resistance. Mol. Cancer 2020, 19, 58. [Google Scholar] [CrossRef]
- Salzman, J. Circular RNA Expression: Its Potential Regulation and Function. Trends Genet. 2016, 32, 309–316. [Google Scholar] [CrossRef]
- Rawluk, J.; Waller, C.F. Gefitinib. Small Mol. Oncol. 2018, 211, 235–246. [Google Scholar] [CrossRef]
- Liu, F.; Zhang, J.; Qin, L.; Yang, Z.; Xiong, J.; Zhang, Y.; Li, R.; Li, S.; Wang, H.; Yu, B. Circular RNA EIF6 (Hsa_circ_0060060) Sponges miR-144-3p to Promote the Cisplatin-Resistance of Human Thyroid Carcinoma Cells by Autophagy Regulation. Aging 2018, 10, 3806. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Ji, M.; Wang, Q.; He, N.; Li, Y. Circular RNA Cdr1as Upregulates SCAI to Suppress Cisplatin Resistance in Ovarian Cancer via miR-1270 Suppression. Mol. Ther. Nucleic Acids 2019, 18, 24–33. [Google Scholar] [CrossRef] [PubMed]
- Xiong, D.; Dang, Y.; Lin, P.; Wen, D.; He, R.; Luo, D.; Feng, Z.; Chen, G. A circRNA–miRNA–mRNA Network Identification for Exploring Underlying Pathogenesis and Therapy Strategy of Hepatocellular Carcinoma. J. Transl. Med. 2018, 16, 220. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tang, S.; Li, L.; Sun, C.; Liu, Y.; Zhao, Y. Depletion of circPDSS1 Inhibits ITGA11 Production to Confer Cisplatin Sensitivity through miR-515-5p in Gastric Cancer. J. Chemother. 2022, 35, 514–526. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.; Wang, Y.; Peng, J.; Li, H.; Gu, J.; Ji, N.; Li, T.; Zhou, X.; Zeng, X.; Li, J. CircRFWD3 Promotes HNSCC Metastasis by Modulating miR-27a/b/PPARγ Signaling. Cell Death Discov. 2022, 8, 285. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Yu, X.; Ding, Y.; Zhao, J.; Wang, G.; Wu, X.; Jiang, J.; Peng, C.; Guo, G.Z.; Cui, S. MiR-770-5p Inhibits Cisplatin Chemoresistance in Human Ovarian Cancer by Targeting ERCC2. Oncotarget 2016, 7, 53254–53268. [Google Scholar] [CrossRef]
- Qu, W.; Huang, W.; Yang, F.; Ju, H.; Zhu, G. Long Noncoding RNA LINC00461 Mediates Cisplatin Resistance of Rectal Cancer via miR-593-5p/CCND1 Axis. Biomed. Pharmacother. 2020, 124, 109740. [Google Scholar] [CrossRef]
- Geng, X.; Zhang, Y.; Lin, X.; Zeng, Z.; Hu, J.; Hao, L.; Xu, J.; Wang, X.; Wang, H.; Li, Q. Exosomal circWDR62 Promotes Temozolomide Resistance and Malignant Progression through Regulation of the miR-370-3p/MGMT Axis in Glioma. Cell Death Dis. 2022, 13, 596. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, C.; Liu, X.; Wang, Y.; Zhao, R.; Yang, Y.; Zheng, X.; Zhang, Y.; Zhang, X. circHIPK3 Promotes Oxaliplatin-Resistance in Colorectal Cancer through Autophagy by Sponging miR-637. EBioMedicine 2019, 48, 277–288. [Google Scholar] [CrossRef]
- Zhang, X.; Xie, K.; Zhou, H.; Wu, Y.; Li, C.; Liu, Y.; Liu, Z.; Xu, Q.; Liu, S.; Xiao, D.; et al. Role of Non-Coding RNAs and RNA Modifiers in Cancer Therapy Resistance. Mol. Cancer 2020, 19, 47. [Google Scholar] [CrossRef]
- Vo, J.N.; Cieslik, M.; Zhang, Y.; Shukla, S.; Xiao, L.; Zhang, Y.; Wu, Y.-M.; Dhanasekaran, S.M.; Engelke, C.G.; Cao, X.; et al. The Landscape of Circular RNA in Cancer. Cell 2019, 176, 869–881.e13. [Google Scholar] [CrossRef] [PubMed]
- Kristensen, L.S.; Jakobsen, T.; Hager, H.; Kjems, J. The Emerging Roles of circRNAs in Cancer and Oncology. Nat. Rev. Clin. Oncol. 2022, 19, 188–206. [Google Scholar] [CrossRef] [PubMed]
- Goodall, G.J.; Wickramasinghe, V.O. RNA in Cancer. Nat. Rev. Cancer 2021, 21, 22–36. [Google Scholar] [CrossRef] [PubMed]
- Kristensen, L.S.; Andersen, M.S.; Stagsted, L.V.W.; Ebbesen, K.K.; Hansen, T.B.; Kjems, J. The Biogenesis, Biology and Characterization of Circular RNAs. Nat. Rev. Genet. 2019, 20, 675–691. [Google Scholar] [CrossRef] [PubMed]
- Du, L.; Guo, D.; Sun, C.; Yan, X.; Lin, S.; Xu, S. CircPIM3 Regulates Taxol Resistance in Non-Small Cell Lung Cancer via miR-338-3p/TNFAIP8 Axis. Anticancer. Drugs 2023, 34, 115–125. [Google Scholar] [CrossRef]
- Wang, F.; Yu, C.; Chen, L.; Xu, S. Landscape of Circular RNAs in Different Types of Lung Cancer and an Emerging Role in Therapeutic Resistance. Int. J. Oncol. 2023, 62, 21. [Google Scholar] [CrossRef]
- Chen, Z.-W.; Hu, J.-F.; Wang, Z.-W.; Liao, C.-Y.; Kang, F.-P.; Lin, C.-F.; Huang, Y.; Huang, L.; Tian, Y.-F.; Chen, S. Circular RNA Circ-MTHFD1L Induces HR Repair to Promote Gemcitabine Resistance via the miR-615-3p/RPN6 Axis in Pancreatic Ductal Adenocarcinoma. J. Exp. Clin. Cancer Res. 2022, 41, 153. [Google Scholar] [CrossRef]
- Cui, Y.; Fan, J.; Shi, W.; Zhou, Z. Circ_0001667 Knockdown Blocks Cancer Progression and Attenuates Adriamycin Resistance by Depleting NCOA3 via Releasing miR-4458 in Breast Cancer. Drug Dev. Res. 2022, 83, 75–87. [Google Scholar] [CrossRef]
- Hu, C.; Xia, R.; Zhang, X.; Li, T.; Ye, Y.; Li, G.; He, R.; Li, Z.; Lin, Q.; Zheng, S. circFARP1 Enables Cancer-Associated Fibroblasts to Promote Gemcitabine Resistance in Pancreatic Cancer via the LIF/STAT3 Axis. Mol. Cancer 2022, 21, 24. [Google Scholar] [CrossRef]
- Li, P.; Song, R.; Yin, F.; Liu, M.; Liu, H.; Ma, S.; Jia, X.; Lu, X.; Zhong, Y.; Yu, L. circMRPS35 Promotes Malignant Progression and Cisplatin Resistance in Hepatocellular Carcinoma. Mol. Ther. 2022, 30, 431–447. [Google Scholar] [CrossRef]
- Li, H.; Luo, F.; Jiang, X.; Zhang, W.; Xiang, T.; Pan, Q.; Cai, L.; Zhao, J.; Weng, D.; Li, Y. CircITGB6 Promotes Ovarian Cancer Cisplatin Resistance by Resetting Tumor-Associated Macrophage Polarization toward the M2 Phenotype. J. Immunother. Cancer 2022, 10, e004029. [Google Scholar] [CrossRef] [PubMed]
- Ling, Y.; Liang, G.; Lin, Q.; Fang, X.; Luo, Q.; Cen, Y.; Mehrpour, M.; Hamai, A.; Liu, Z.; Shi, Y. circCDYL2 Promotes Trastuzumab Resistance via Sustaining HER2 Downstream Signaling in Breast Cancer. Mol. Cancer 2022, 21, 8. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.-X.; Gao, J.; Long, X.; Zhang, P.-F.; Yang, X.; Zhu, S.-Q.; Pei, X.; Qiu, B.-Q.; Chen, S.-W.; Lu, F. The Circular RNA circHMGB2 Drives Immunosuppression and Anti-PD-1 Resistance in Lung Adenocarcinomas and Squamous Cell Carcinomas via the miR-181a-5p/CARM1 Axis. Mol. Cancer 2022, 21, 110. [Google Scholar] [CrossRef] [PubMed]
- Jahani, S.; Nazeri, E.; Majidzadeh-A, K.; Jahani, M.; Esmaeili, R. Circular RNA; a New Biomarker for Breast Cancer: A Systematic Review. J. Cell. Physiol. 2020, 235, 5501–5510. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Cui, Z.; Guo, X. Comprehensive Analysis of Circular RNA Expression Profiles in Cisplatin-Resistant Non-Small Cell Lung Cancer Cell Lines. Acta Biochim. Biophys. Sin. 2020, 52, 944–953. [Google Scholar] [CrossRef]
- Xin, C.; Huang, F.; Wang, J.; Li, J.; Chen, Q. Roles of circRNAs in Cancer Chemoresistance. Oncol. Rep. 2021, 46, 225. [Google Scholar] [CrossRef]
- Wei, D.; Zeng, J.; Rong, F.; Xu, Y.; Wei, R.; Zou, C. Circ_0020123 Enhances the Cisplatin Resistance in Non-Small Cell Lung Cancer Cells Partly by Sponging miR-140-3p to Regulate Homeobox B5 (HOXB5). Bioengineered 2022, 13, 5126–5140. [Google Scholar] [CrossRef]
- Solassol, I.; Pinguet, F.; Quantin, X. FDA-and EMA-Approved Tyrosine Kinase Inhibitors in Advanced EGFR-Mutated Non-Small Cell Lung Cancer: Safety, Tolerability, Plasma Concentration Monitoring, and Management. Biomolecules 2019, 9, 668. [Google Scholar] [CrossRef]
- Zhu, M.; Yang, L.; Wang, X. NEAT1 Knockdown Suppresses the Cisplatin Resistance in Ovarian Cancer by Regulating miR-770-5p/PARP1 Axis. Cancer Manag. Res. 2020, 12, 7277–7289. [Google Scholar] [CrossRef]
- Zhang, D.; Li, Y.; Sun, P. miR-770-5p Modulates Resistance to Methotrexate in Human Colorectal Adenocarcinoma Cells by Downregulating HIPK. Exp. Ther. Med. 2020, 19, 339–346. [Google Scholar]
- Chen, Z.; Xu, X. Roles of Nucleolin: Focus on Cancer and Anti-Cancer Therapy. Saudi Med. J. 2016, 37, 1312–1318. [Google Scholar] [CrossRef] [PubMed]
- Tominaga, K.; Srikantan, S.; Lee, E.K.; Subaran, S.S.; Martindale, J.L.; Abdelmohsen, K.; Gorospe, M. Competitive Regulation of Nucleolin Expression by HuR and miR-494. Mol. Cell. Biol. 2011, 31, 4219–4231. [Google Scholar] [CrossRef] [PubMed]
- Ke, J.; Gu, C.; Zhang, H.; Liu, Y.; Zhang, W.; Rao, H.; Li, S.; Wu, F. Nucleolin Promotes Cisplatin Resistance in Cervical Cancer by the YB1-MDR1 Pathway. J. Oncol. 2021, 2021, 9992218. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, T.; Xin, C.; Zhang, S.; Tian, X.; Hu, Y.; Wang, Y.; Wang, J.; Ji, N.; Zeng, X.; Li, J. Circular RNA from Tyrosylprotein Sulfotransferase 2 Gene Inhibits Cisplatin Sensitivity in Head and Neck Squamous Cell Carcinoma by Sponging miR-770-5p and Interacting with Nucleolin. Cancers 2023, 15, 5351. https://doi.org/10.3390/cancers15225351
Wang T, Xin C, Zhang S, Tian X, Hu Y, Wang Y, Wang J, Ji N, Zeng X, Li J. Circular RNA from Tyrosylprotein Sulfotransferase 2 Gene Inhibits Cisplatin Sensitivity in Head and Neck Squamous Cell Carcinoma by Sponging miR-770-5p and Interacting with Nucleolin. Cancers. 2023; 15(22):5351. https://doi.org/10.3390/cancers15225351
Chicago/Turabian StyleWang, Tianqing, Chuan Xin, Shiyu Zhang, Xin Tian, Yuting Hu, Ying Wang, Jiongke Wang, Ning Ji, Xin Zeng, and Jing Li. 2023. "Circular RNA from Tyrosylprotein Sulfotransferase 2 Gene Inhibits Cisplatin Sensitivity in Head and Neck Squamous Cell Carcinoma by Sponging miR-770-5p and Interacting with Nucleolin" Cancers 15, no. 22: 5351. https://doi.org/10.3390/cancers15225351
APA StyleWang, T., Xin, C., Zhang, S., Tian, X., Hu, Y., Wang, Y., Wang, J., Ji, N., Zeng, X., & Li, J. (2023). Circular RNA from Tyrosylprotein Sulfotransferase 2 Gene Inhibits Cisplatin Sensitivity in Head and Neck Squamous Cell Carcinoma by Sponging miR-770-5p and Interacting with Nucleolin. Cancers, 15(22), 5351. https://doi.org/10.3390/cancers15225351