
Exploring Cell Migration Mechanisms in Cancer: From Wound Healing Assays to Cellular Automata Models


 , and
, and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. In Vitro Experiments
2.2. Model Development
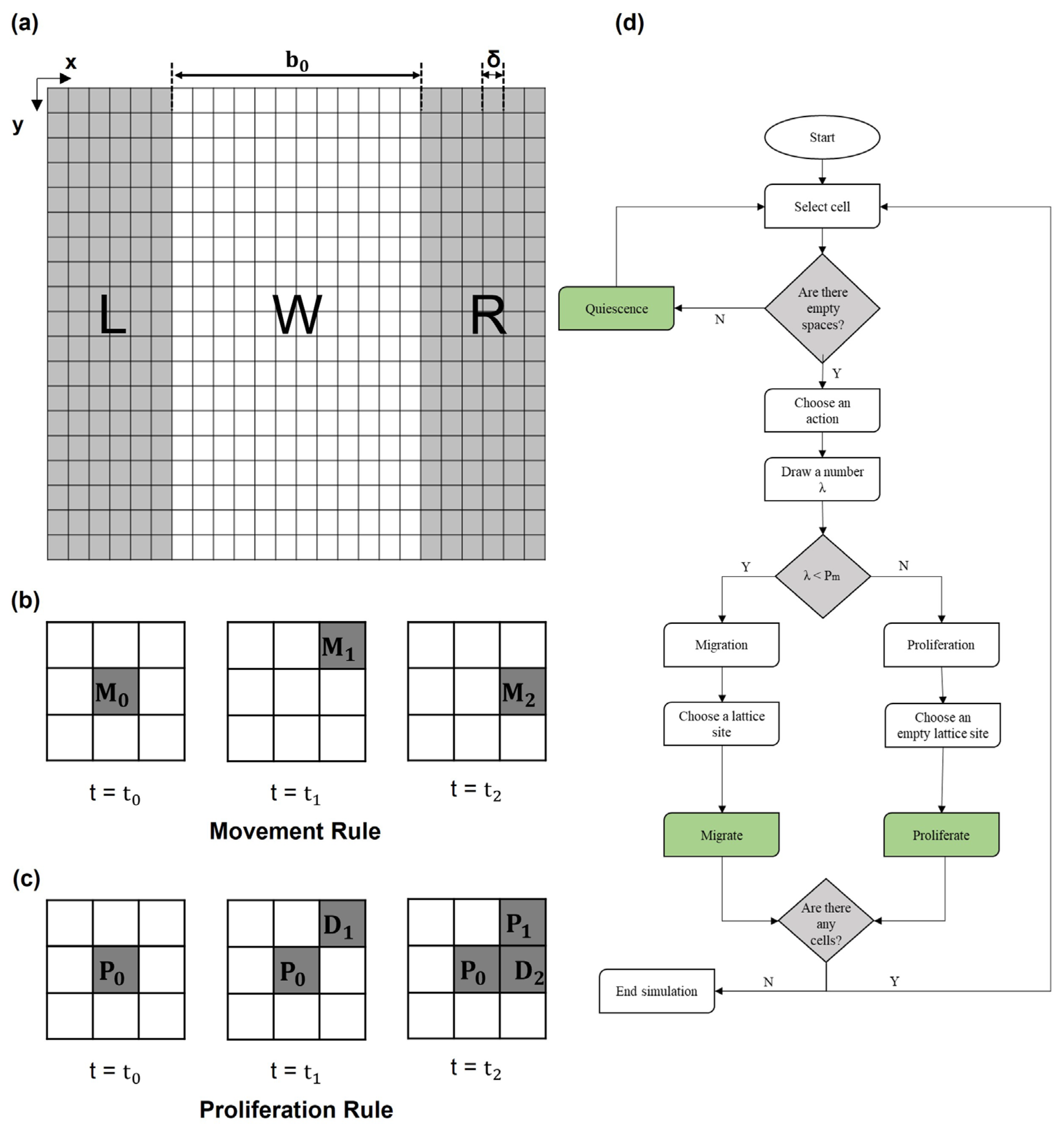
2.2.1. Domain Building
2.2.2. Rules Governing Cellular Dynamics
- Migration and proliferation
- 2.
- Quiescence
- (1)
- Spatial domain discretization and initialization of cell positions.
- (2)
- Testing for empty neighbors for every occupied element.
- (3)
- Random number (λ) assignment to the occupied CA elements to decide the actions of cells:
- If λ > , the actual site of the cell of interest will remain occupied by the cell, and a daughter cell will be placed in a randomly chosen empty site among the neighbors.
- If λ < , the site of the cell of interest will become empty, and a neighboring empty site will become occupied.
- (4)
- Lattice updates according to the selected actions based on probabilities.
- (5)
- Stop if the wound was healed; otherwise, proceed to next time step and return to (1).
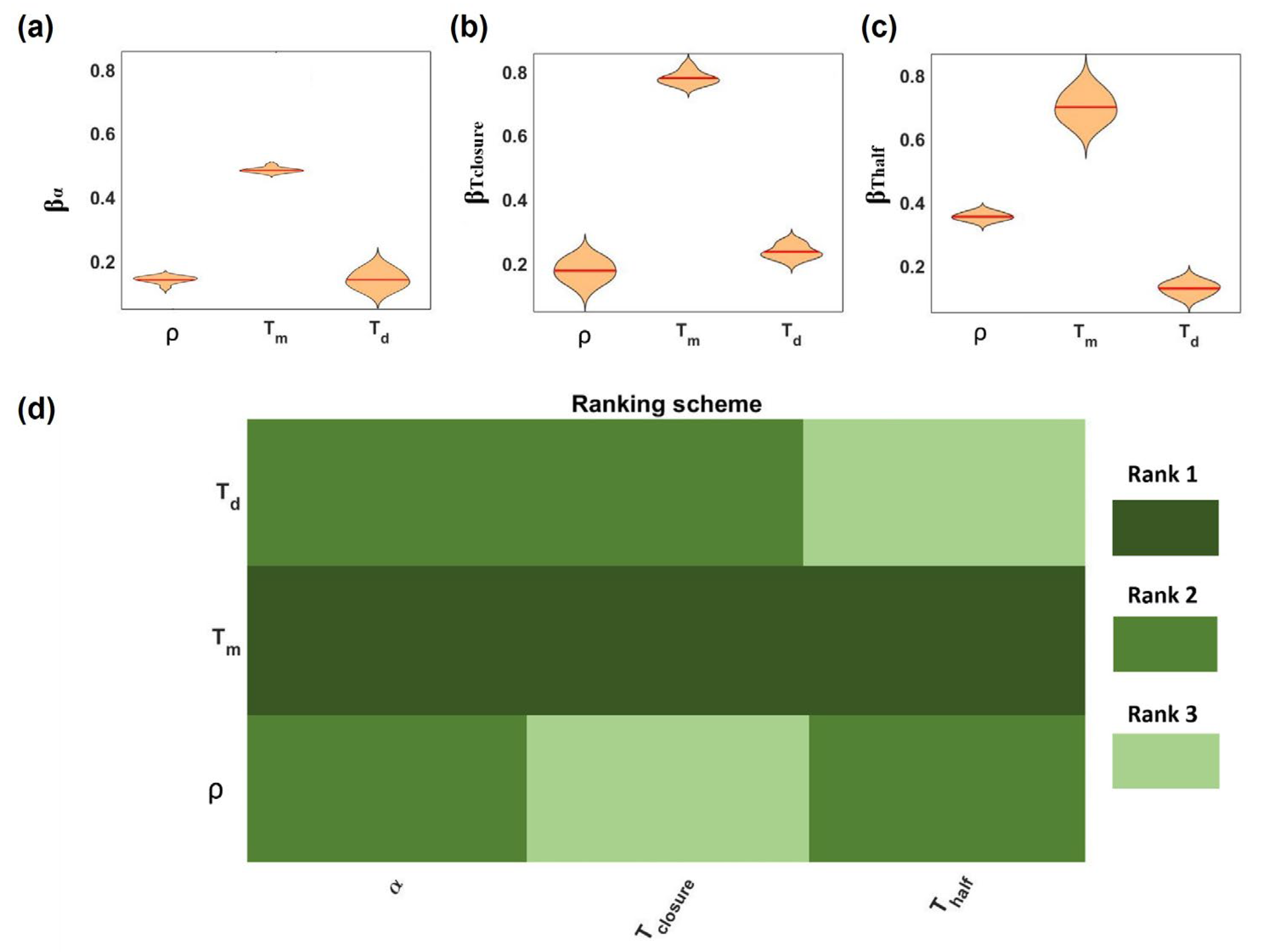
2.3. Parameter Sensitivity Analysis
Global Sensitivity Analysis
3. Results
3.1. Baseline Model Behavior and Model Calibration
3.2. Model Predictions
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dillekås, H.; Rogers, M.S.; Straume, O. Are 90% of deaths from cancer caused by metastases? Cancer Med. 2019, 8, 5574–5576. [Google Scholar] [CrossRef]
- Zhang, Z.; Chen, L.; Humphries, B.; Brien, R.; Wicha, M.S.; Luker, K.E.; Luker, G.D.; Chen, Y.-C.; Yoon, E. Morphology-based prediction of cancer cell migration using an artificial neural network and a random decision forest. Integr. Biol. 2018, 10, 758–767. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, H.; Wyckoff, J.; Condeelis, J. Cell migration in tumors. Curr. Opin. Cell Biol. 2005, 17, 559–564. [Google Scholar] [CrossRef] [PubMed]
- Vedula, S.R.K.; Ravasio, A.; Lim, C.T.; Ladoux, B. Collective cell migration: A mechanistic perspective. Physiology 2013, 28, 370–379. [Google Scholar] [CrossRef] [PubMed]
- Ascione, F.; Caserta, S.; Guido, S. The wound healing assay revisited: A transport phenomena approach. Chem. Eng. Sci. 2017, 160, 200–209. [Google Scholar] [CrossRef]
- Grada, A.; Otero-Vinas, M.; Prieto-Castrillo, F.; Obagi, Z.; Falanga, V. Research techniques made simple: Analysis of collective cell migration using the wound healing assay. J. Investig. Dermatol. 2017, 137, e11–e16. [Google Scholar] [CrossRef]
- Martinotti, S.; Ranzato, E. Scratch wound healing assay. Epidermal Cells Methods Protoc. 2020, 225–229. [Google Scholar]
- Chen, H.-C. Boyden Chamber Assay. In Cell Migration; Springer: Berlin/Heidelberg, Germany, 2005; pp. 15–22. [Google Scholar]
- Woo, M.M.M.; Salamanca, C.M.; Minor, A.; Auersperg, N. An improved assay to quantitate the invasiveness of cells in modified Boyden chambers. Vitr. Cell. Dev. Biol. Anim. 2007, 43, 7–9. [Google Scholar] [CrossRef]
- Prasad, M.; Montell, D.J. Cellular and molecular mechanisms of border cell migration analyzed using time-lapse live-cell imaging. Dev. Cell 2007, 12, 997–1005. [Google Scholar] [CrossRef]
- Kijanka, G.S.; Dimov, I.K.; Burger, R.; Ducrée, J. Real-time monitoring of cell migration, phagocytosis and cell surface receptor dynamics using a novel, live-cell opto-microfluidic technique. Anal. Chim. Acta 2015, 872, 95–99. [Google Scholar] [CrossRef]
- Huang, Y.; Agrawal, B.; Clark, P.A.; Williams, J.C.; Kuo, J.S. Evaluation of cancer stem cell migration using compartmentalizing microfluidic devices and live cell imaging. J. Vis. Exp. 2011, 58, e3297. [Google Scholar]
- Ascione, F.; Vasaturo, A.; Caserta, S.; D’Esposito, V.; Formisano, P.; Guido, S. Comparison between fibroblast wound healing and cell random migration assays in vitro. Exp. Cell Res. 2016, 347, 123–132. [Google Scholar] [CrossRef]
- Blockhuys, S.; Zhang, X.; Wittung-Stafshede, P. Single-cell tracking demonstrates copper chaperone Atox1 to be required for breast cancer cell migration. Proc. Natl. Acad. Sci. USA 2020, 117, 2014–2019. [Google Scholar] [CrossRef] [PubMed]
- Brand, S.; Beigel, F.; Olszak, T.; Zitzmann, K.; Eichhorst, S.R.T.; Otte, J.-M.; Diepolder, H.; Marquardt, A.; Jagla, W.; Popp, A. IL-22 is increased in active Crohn’s disease and promotes proinflammatory gene expression and intestinal epithelial cell migration. Am. J. Physiol. Gastrointest. Liver Physiol. 2006, 290, G827–G838. [Google Scholar] [CrossRef] [PubMed]
- Willier, S.; Butt, E.; Grunewald, T.G.P. Lysophosphatidic acid (LPA) signalling in cell migration and cancer invasion: A focussed review and analysis of LPA receptor gene expression on the basis of more than 1700 cancer microarrays. Biol. Cell 2013, 105, 317–333. [Google Scholar] [CrossRef]
- Feng, Y.; Walsh, C.A. Protein–protein interactions, cytoskeletal regulation and neuronal migration. Nat. Rev. Neurosci. 2001, 2, 408–416. [Google Scholar] [CrossRef]
- Jonkman, J.E.N.; Cathcart, J.A.; Xu, F.; Bartolini, M.E.; Amon, J.E.; Stevens, K.M.; Colarusso, P. An introduction to the wound healing assay using live-cell microscopy. Cell Adhes. Migr. 2014, 8, 440–451. [Google Scholar] [CrossRef]
- Tremel, A.; Cai, A.; Tirtaatmadja, N.; Hughes, B.D.; Stevens, G.W.; Landman, K.A.; Connor, A.J.O. Cell migration and proliferation during monolayer formation and wound healing. Chem. Eng. Sci. 2009, 64, 247–253. [Google Scholar] [CrossRef]
- Cao, D.; Chu, W.K.; Ng, T.K.; Yip, Y.W.Y.; Young, A.L.; Pang, C.P.; Jhanji, V. Cellular proliferation and migration of human pterygium cells: Mitomycin versus small-molecule inhibitors. Cornea 2018, 37, 760–766. [Google Scholar] [CrossRef]
- Lorie, E.P.; Stricker, N.; Plitta-Michalak, B.; Chen, I.P.; Volkmer, B.; Greinert, R.; Jauch, A.; Boukamp, P.; Rapp, A. Characterisation of the novel spontaneously immortalized and invasively growing human skin keratinocyte line HaSKpw. Sci. Rep. 2020, 10, 15196. [Google Scholar] [CrossRef]
- Savla, U.; Olson, L.E.; Waters, C.M. Mathematical modeling of airway epithelial wound closure during cyclic mechanical strain. J. Appl. Physiol. 2004, 96, 566–574. [Google Scholar] [CrossRef] [PubMed]
- Ledzewicz, U.; Naghnaeian, M.; Schättler, H. Optimal response to chemotherapy for a mathematical model of tumor–immune dynamics. J. Math. Biol. 2012, 64, 557–577. [Google Scholar] [CrossRef] [PubMed]
- Ledzewicz, U.; Schättler, H. Optimal and suboptimal protocols for a class of mathematical models of tumor anti-angiogenesis. J. Theor. Biol. 2008, 252, 295–312. [Google Scholar] [CrossRef]
- Bartocci, E.; Lió, P. Computational modeling, formal analysis, and tools for systems biology. PLoS Comput. Biol. 2016, 12, e1004591. [Google Scholar] [CrossRef] [PubMed]
- Baratchart, E.; Lo, C.H.; Lynch, C.C.; Basanta, D. Integrated computational and in vivo models reveal Key Insights into macrophage behavior during bone healing. PLoS Comput. Biol. 2022, 18, e1009839. [Google Scholar] [CrossRef]
- Fisher, R.A. The wave of advance of advantageous genes. Ann. Eugen. 1937, 7, 355–369. [Google Scholar] [CrossRef]
- Maini, P.K.; McElwain, D.L.S.; Leavesley, D. Travelling waves in a wound healing assay. Appl. Math. Lett. 2004, 17, 575–580. [Google Scholar] [CrossRef]
- Zahm, J.M.; Kaplan, H.; Hérard, A.L.; Doriot, F.; Pierrot, D.; Somelette, P.; Puchelle, E. Cell migration and proliferation during the in vitro wound repair of the respiratory epithelium. Cell Motil. Cytoskelet. 1997, 37, 33–43. [Google Scholar] [CrossRef]
- Cai, A.Q.; Landman, K.A.; Hughes, B.D. Multi-scale modeling of a wound-healing cell migration assay. J. Theor. Biol. 2007, 245, 576–594. [Google Scholar] [CrossRef]
- Ascione, F.; Guarino, A.M.; Calabrò, V.; Guido, S.; Caserta, S. A novel approach to quantify the wound closure dynamic. Exp. Cell Res. 2017, 352, 175–183. [Google Scholar] [CrossRef]
- Dallon, J.C.; Sherratt, J.A.; Maini, P.K. Modeling the effects of transforming growth factor-β on extracellular matrix alignment in dermal wound repair. Wound Repair Regen. 2001, 9, 278–286. [Google Scholar] [CrossRef] [PubMed]
- Dolak, Y.; Schmeiser, C. Kinetic models for chemotaxis: Hydrodynamic limits and spatio-temporal mechanisms. J. Math. Biol. 2005, 51, 595–615. [Google Scholar] [CrossRef]
- Chauviere, A.; Hillen, T.; Preziosi, L. Modeling cell movement in anisotropic and heterogeneous network tissues. Netw. Heterog. Media 2007, 2, 333. [Google Scholar] [CrossRef]
- Dickinson, R.B.; Tranquillo, R.T. Optimal estimation of cell movement indices from the statistical analysis of cell tracking data. AIChE J. 1993, 39, 1995–2010. [Google Scholar] [CrossRef]
- Santé, I.; García, A.M.; Miranda, D.; Crecente, R. Cellular automata models for the simulation of real-world urban processes: A review and analysis. Landsc. Urban Plan. 2010, 96, 108–122. [Google Scholar] [CrossRef]
- Farrelly, T. A review of quantum cellular automata. Quantum 2020, 4, 368. [Google Scholar] [CrossRef]
- Ermentrout, G.B.; Edelstein-Keshet, L. Cellular automata approaches to biological modeling. J. Theor. Biol. 1993, 160, 97–133. [Google Scholar] [CrossRef]
- Metzcar, J.; Wang, Y.; Heiland, R.; Macklin, P. A review of cell-based computational modeling in cancer biology. JCO Clin. Cancer Inform. 2019, 2, 1–13. [Google Scholar] [CrossRef]
- Deutsch, A.; Nava-Sedeño, J.M.; Syga, S.; Hatzikirou, H. BIO-LGCA: A cellular automaton modelling class for analysing collective cell migration. PLoS Comput. Biol. 2021, 17, e1009066. [Google Scholar] [CrossRef]
- Walker, D.C.; Hill, G.; Wood, S.M.; Smallwood, R.H.; Southgate, J. Agent-based computational modeling of wounded epithelial cell monolayers. IEEE Trans. Nanobiosci. 2004, 3, 153–163. [Google Scholar] [CrossRef]
- Khain, E.; Leonard, M.S.; Schneider-Mizell, a.C.M. The Role of Cell-Cell Adhesion in Wound Healing. J. Stat. Phys. 2007, 128, 209–218. [Google Scholar] [CrossRef]
- Beauchemin, C.; Samuel, J.; Tuszynski, J. A simple cellular automaton model for influenza A viral infections. J. Theor. Biol. 2005, 232, 223–234. [Google Scholar] [CrossRef] [PubMed]
- Pourhasanzade, F.; Sabzpoushan, S.H. A cellular automata model of chemotherapy effects on tumour growth: Targeting cancer and immune cells. Math. Comput. Model. Dyn. Syst. 2019, 25, 63–89. [Google Scholar] [CrossRef]
- Brüningk, S.C.; Ziegenhein, P.; Rivens, I.; Oelfke, U.; Haar, G.t. A cellular automaton model for spheroid response to radiation and hyperthermia treatments. Sci. Rep. 2019, 9, 17674. [Google Scholar] [CrossRef] [PubMed]
- Robertson-Tessi, M.; Gillies, R.J.; Gatenby, R.A.; Anderson, A.R.A. Impact of metabolic heterogeneity on tumor growth, invasion, and treatment outcomes. Cancer Res. 2015, 75, 1567–1579. [Google Scholar] [CrossRef]
- Alber, M.S.; Kiskowski, M.A.; Glazier, J.A.; Jiang, Y. On cellular automaton approaches to modeling biological cells. In Mathematical Systems Theory in Biology, Communications, Computation, and Finance; Springer: Berlin/Heidelberg, Germany, 2003; pp. 1–39. [Google Scholar]
- Krzyszczyk, P.; Acevedo, A.; Davidoff, E.J.; Timmins, L.M.; Marrero-Berrios, I.; Patel, M.; White, C.; Lowe, C.; Sherba, J.J.; Hartmanshenn, C. The growing role of precision and personalized medicine for cancer treatment. Technology 2018, 6, 79–100. [Google Scholar] [CrossRef]
- Gaglione, R.; Dell’Olmo, E.; Bosso, A.; Chino, M.; Pane, K.; Ascione, F.; Itri, F.; Caserta, S.; Amoresano, A.; Lombardi, A.; et al. Novel human bioactive peptides identified in Apolipoprotein B: Evaluation of their therapeutic potential. Biochem. Pharmacol. 2017, 130, 34–50. [Google Scholar] [CrossRef]
- Montano, E.; Vivo, M.; Guarino, A.M.; di Martino, O.; Di Luccia, B.; Calabrò, V.; Caserta, S.; Pollice, A. Colloidal Silver Induces Cytoskeleton Reorganization and E-Cadherin Recruitment at Cell-Cell Contacts in HaCaT Cells. Pharmaceuticals 2019, 12, 72. [Google Scholar] [CrossRef]
- Vuoso, D.C.; D’Angelo, S.; Ferraro, R.; Caserta, S.; Guido, S.; Cammarota, M.; Porcelli, M.; Cacciapuoti, G. Annurca apple polyphenol extract promotes mesenchymal-to-epithelial transition and inhibits migration in triple-negative breast cancer cells through ROS/JNK signaling. Sci. Rep. 2020, 10, 15921. [Google Scholar] [CrossRef]
- D’Argenio, G.; Mazzone, G.; Tuccillo, C.; Ribecco, M.T.; Graziani, G.; Gravina, A.G.; Caserta, S.; Guido, S.; Fogliano, V.; Caporaso, N.; et al. Apple polyphenols extract (APE) improves colon damage in a rat model of colitis. Dig. Liver Dis. 2012, 44, 555–562. [Google Scholar] [CrossRef]
- Mayor, R.; Carmona-Fontaine, C. Keeping in touch with contact inhibition of locomotion. Trends Cell Biol. 2010, 20, 319–328. [Google Scholar] [CrossRef]
- Abercrombie, M. Contact inhibition in tissue culture. Vitr. Cell. Dev. Biol. Plant 1970, 6, 128–142. [Google Scholar] [CrossRef]
- Wang, Z.; Deisboeck, T.S.; Cristini, V. Development of a sampling-based global sensitivity analysis workflow for multiscale computational cancer models. IET Syst. Biol. 2014, 8, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Seaholm, S.K.; Ackerman, E.; Wu, S.C. Latin hypercube sampling and the sensitivity analysis of a Monte Carlo epidemic model. Int. J. Biomed. Comput. 1988, 23, 97–112. [Google Scholar] [CrossRef] [PubMed]
- Ottino, J.M. Chemical engineering in a complex world: Grand challenges, vast opportunities. AIChE J. 2011, 57, 1654–1668. [Google Scholar] [CrossRef]
- Gayan, S.; Teli, A.; Dey, T. Inherent aggressive character of invasive and non-invasive cells dictates the in vitro migration pattern of multicellular spheroid. Sci. Rep. 2017, 7, 11527. [Google Scholar] [CrossRef]
- Tschon, M.; Incerti-Parenti, S.; Cepollaro, S.; Checchi, L.; Fini, M. Photobiomodulation with low-level diode laser promotes osteoblast migration in an in vitro micro wound model. J. Biomed. Opt. 2015, 20, 078002. [Google Scholar] [CrossRef]
- Rodan, S.B.; Imai, Y.; Thiede, M.A.; Wesolowski, G.; Thompson, D.; Bar-Shavit, Z.; Shull, S.; Mann, K.; Rodan, G.A. Characterization of a human osteosarcoma cell line (Saos-2) with osteoblastic properties. Cancer Res. 1987, 47, 4961–4966. [Google Scholar]
- Meir, M.; Flemming, S.; Burkard, N.; Bergauer, L.; Metzger, M.; Germer, C.-T.; Schlegel, N. Glial cell line-derived neurotrophic factor promotes barrier maturation and wound healing in intestinal epithelial cells in vitro. Am. J. Physiol. Gastrointest. Liver Physiol. 2015, 309, G613–G624. [Google Scholar] [CrossRef]
- Michael, W. (Ed.) An Introduction to Molecular Biotechnology: Fundamentals, Methods and Applications; John Wiley & Sons: Hoboken, NJ, USA, 2020; Volume 27, pp. 1043–1149. [Google Scholar]
- Pautke, C.; Schieker, M.; Tischer, T.; Kolk, A.; Neth, P.; Mutschler, W.; Milz, S. Characterization of osteosarcoma cell lines MG-63, Saos-2 and U-2 OS in comparison to human osteoblasts. Anticancer Res. 2004, 24, 3743–3748. [Google Scholar]
- Connolly, S.; McGourty, K.; Newport, D. The in vitro inertial positions and viability of cells in suspension under different in vivo flow conditions. Sci. Rep. 2020, 10, 1711. [Google Scholar] [CrossRef] [PubMed]
- Sixt, M.; Lämmermann, T. In Vitro Analysis of Chemotactic Leukocyte Migration in 3D Environments. In Cell Migration; Springer: Berlin/Heidelberg, Germany, 2011; pp. 149–165. [Google Scholar]
- Lämmermann, T.; Bader, B.L.; Monkley, S.J.; Worbs, T.; Wedlich-Söldner, R.; Hirsch, K.; Keller, M.; Förster, R.; Critchley, D.R.; Fässler, R. Rapid leukocyte migration by integrin-independent flowing and squeezing. Nature 2008, 453, 51–55. [Google Scholar] [CrossRef] [PubMed]
- Friedl, P.; Bröcker, E.B. The biology of cell locomotion within three-dimensional extracellular matrix. Cell. Mol. Life Sci. CMLS 2000, 57, 41–64. [Google Scholar] [CrossRef] [PubMed]
- Sant, S.; Johnston, P.A. The production of 3D tumor spheroids for cancer drug discovery. Drug Discov. Today Technol. 2017, 23, 27–36. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Description | Range of Variation | Dimension |
|---|---|---|---|
| Characteristic time of migration | 0.005; 0.5 | h | |
| Characteristic time of proliferation | 12; 40 | h | |
| Density: number of cells in unit area | 10−6; 10−3 | cells/µm2 | |
| Characteristic dimension of the cell | 15; 25 | µm | |
| b0 | Initial length of the wound | 370; 900 | µm |
| Motility: the time necessary to travel a length equal to delta | 103; 104 | µm2/h | |
| v | Velocity of the fronts of cells | 5; 60 | µm/h |
| Velocity of wound area variation | 0.02; 0.13 | 1/h |
| Cell line | Id | [#cells/μm2] | α [1/h] | b0 [μm] | Td [h] | Tm [h] | References |
|---|---|---|---|---|---|---|---|
| HT-1080 | 1 | 2.7 × 10−3 | 0.012 | 468 * | 24 | 0.063 | [5] |
| 2 | 2.9 × 10−3 | 0.128 | 371 * | 24 | 0.075 | ||
| 3 | 1.6 × 10−3 | 0.078 | 532 * | 24 | 0.107 | ||
| 4 | 2.1 × 10−3 | 0.078 | 638 * | 24 | 0.075 | ||
| 5 | 1.5 × 10−3 | 0.069 | 548 * | 24 | 0.129 | ||
| 6 | 1.2 × 10−3 | 0.069 | 687 * | 24 | 0.082 | ||
| 7 | N/A | 0.110 | 288 | 24 | 0.110 | [58] | |
| MDA-MB-231 | 8 | 1.0 × 10−3 | 0.023 | 800 | 38 | 0.338 | [31,51] |
| 9 | 1.2 × 10−3 | 0.042 | 930 | 38 | 0.075 | ||
| 10 | 2.3 × 10−3 | 0.044 | 800 | 38 | 0.095 | ||
| 11 | N/A | 0.040 | 288 | 38 | 0.476 | [58] | |
| MDA-MB468 | 12 | 1.2 × 10−3 | 0.031 | 800 | 47 | 0.154 | [51] |
| HaCaT | 13 | 1.2 × 10−3 | 0.043 | 900 | 19 | 0.156 | [49,50] |
| 14 | 1.7 × 10−3 | 0.132 | 900 | 19 | 0.017 | ||
| 15 | 2.5 × 10−2 * | 0.029 | N/A | 19 | 0.078 | [21] | |
| Saos-2: HTB 85 | 16 | N/A | 0.010 | 800 | 37 | 5.851 | [59,60] |
| Caco-2 | 17 | 1.2 × 10−3 * | 0.014 | 882 * | 80 | 0.385 | [61] |
| BEAS | 18 | N/A | 0.054 | 500 | 26 | 0.188 | [18] |
| MCF-7 | 19 | N/A | 0.031 | 500 | 38 | 0.741 | |
| 20 | N/A | 0.040 | 287 | 38 | 0.54 | [58] | |
| NIH/3T3 | 21 | 1.3 × 10−4 | 0.062 | 933 | 20 | 0.002 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Migliaccio, G.; Ferraro, R.; Wang, Z.; Cristini, V.; Dogra, P.; Caserta, S. Exploring Cell Migration Mechanisms in Cancer: From Wound Healing Assays to Cellular Automata Models. Cancers 2023, 15, 5284. https://doi.org/10.3390/cancers15215284
Migliaccio G, Ferraro R, Wang Z, Cristini V, Dogra P, Caserta S. Exploring Cell Migration Mechanisms in Cancer: From Wound Healing Assays to Cellular Automata Models. Cancers. 2023; 15(21):5284. https://doi.org/10.3390/cancers15215284
Chicago/Turabian StyleMigliaccio, Giorgia, Rosalia Ferraro, Zhihui Wang, Vittorio Cristini, Prashant Dogra, and Sergio Caserta. 2023. "Exploring Cell Migration Mechanisms in Cancer: From Wound Healing Assays to Cellular Automata Models" Cancers 15, no. 21: 5284. https://doi.org/10.3390/cancers15215284
APA StyleMigliaccio, G., Ferraro, R., Wang, Z., Cristini, V., Dogra, P., & Caserta, S. (2023). Exploring Cell Migration Mechanisms in Cancer: From Wound Healing Assays to Cellular Automata Models. Cancers, 15(21), 5284. https://doi.org/10.3390/cancers15215284