
Carbonic Anhydrase Inhibitors Induce Ferroptosis through Inhibition of AKT/FTH1 Signaling in Ewing Sarcoma Tumor Cells

 , , , , , ,
, , , , , ,
Abstract
:Simple Summary
Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Methods
2.1. Cells and Reagents
2.2. Single-Cell RNA-Seq and Data Analysis
2.3. Bioinformatic Analysis
2.4. Alamar Blue Cell Viability Assay
2.5. Clonogenic Assay
2.6. Scratch Assay
2.7. MTT
2.8. LDH Assay
2.9. Immunofluorescence
2.10. Flow Cytometry
2.11. Western Blotting
2.12. RNAi Knockdown
2.13. H&E and Immunohistochemistry of Patient Tumor Sections
2.14. Statistical Analysis
3. Results
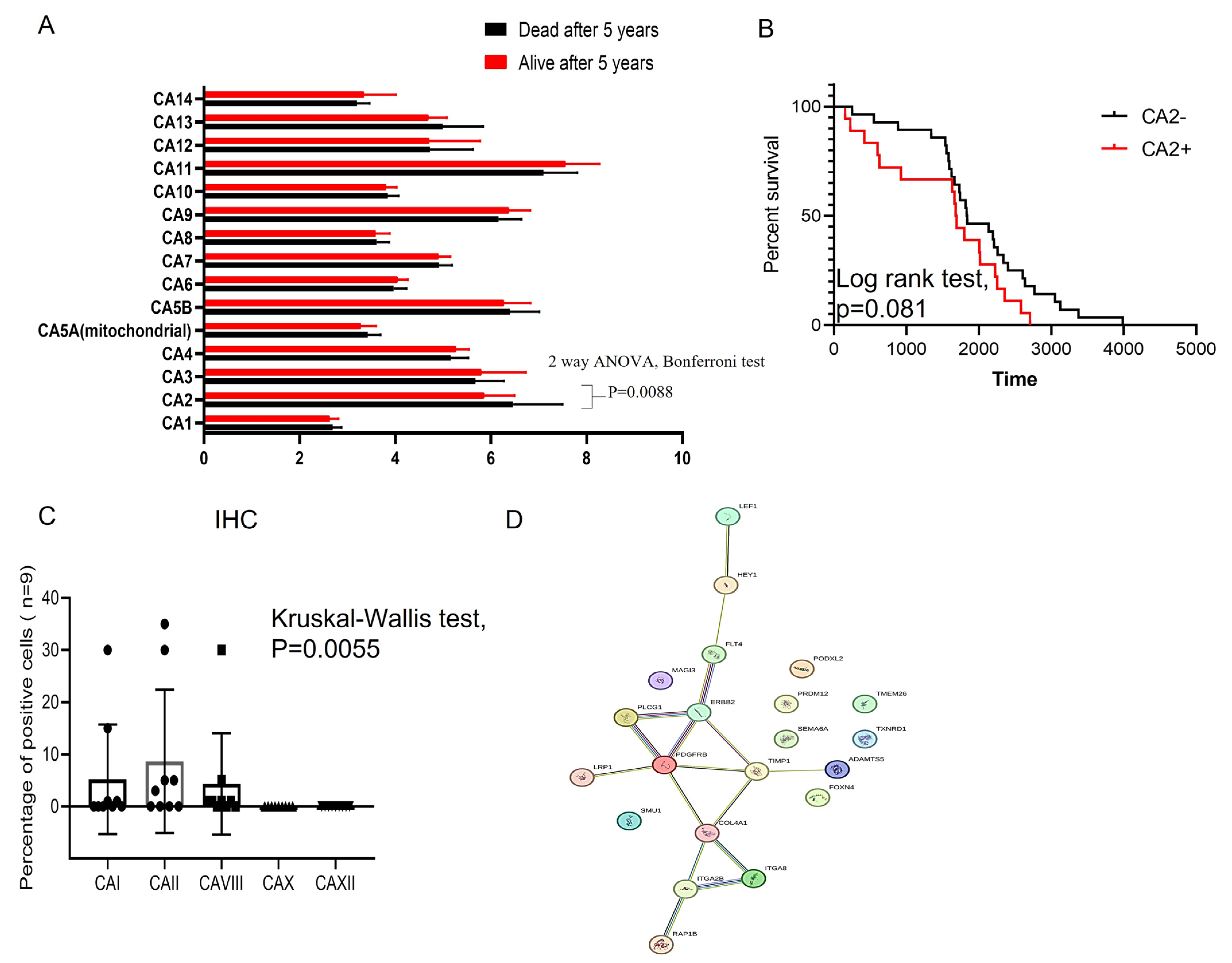
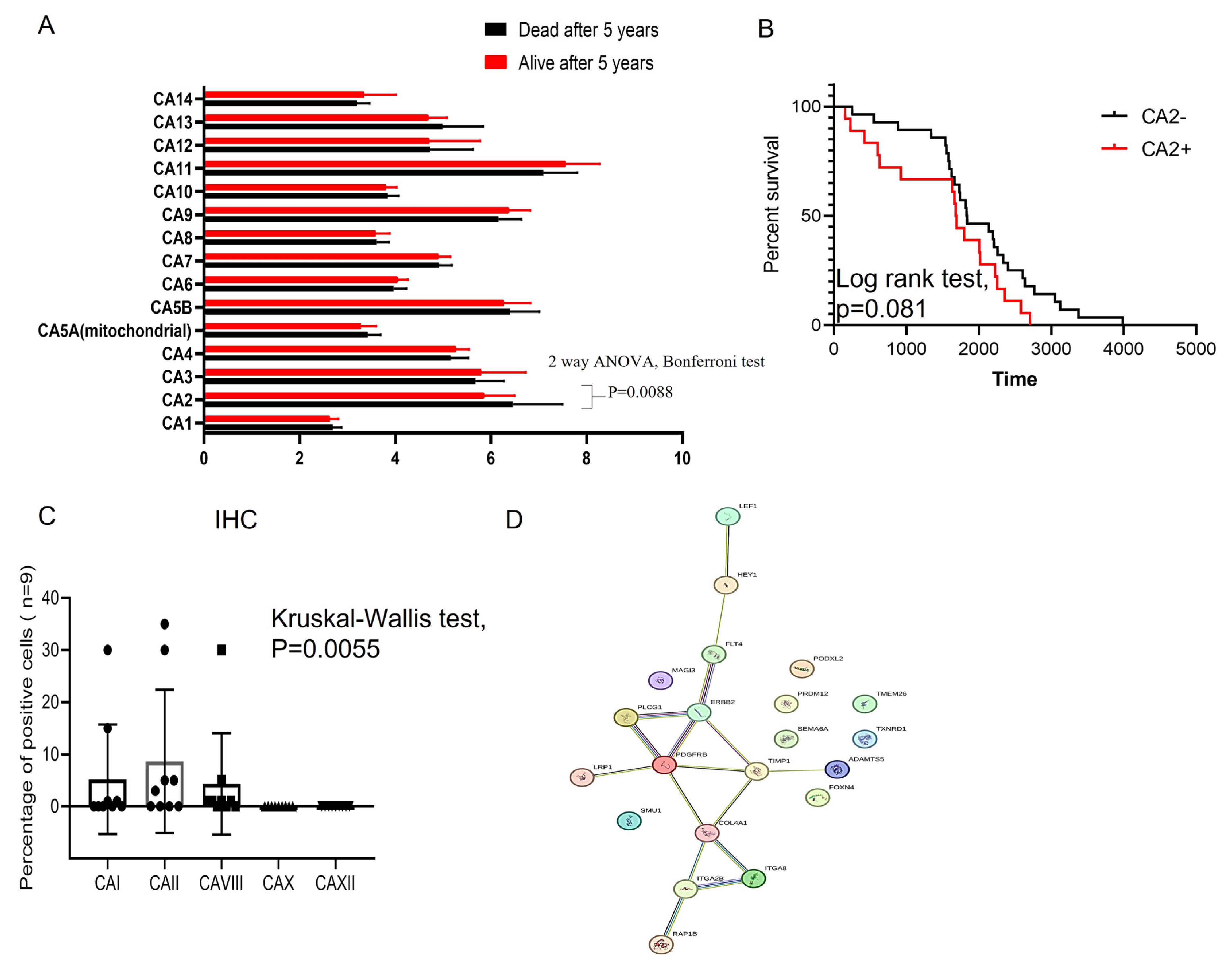
3.1. CA2 Gene Elevates Expression in Primary ES Cells
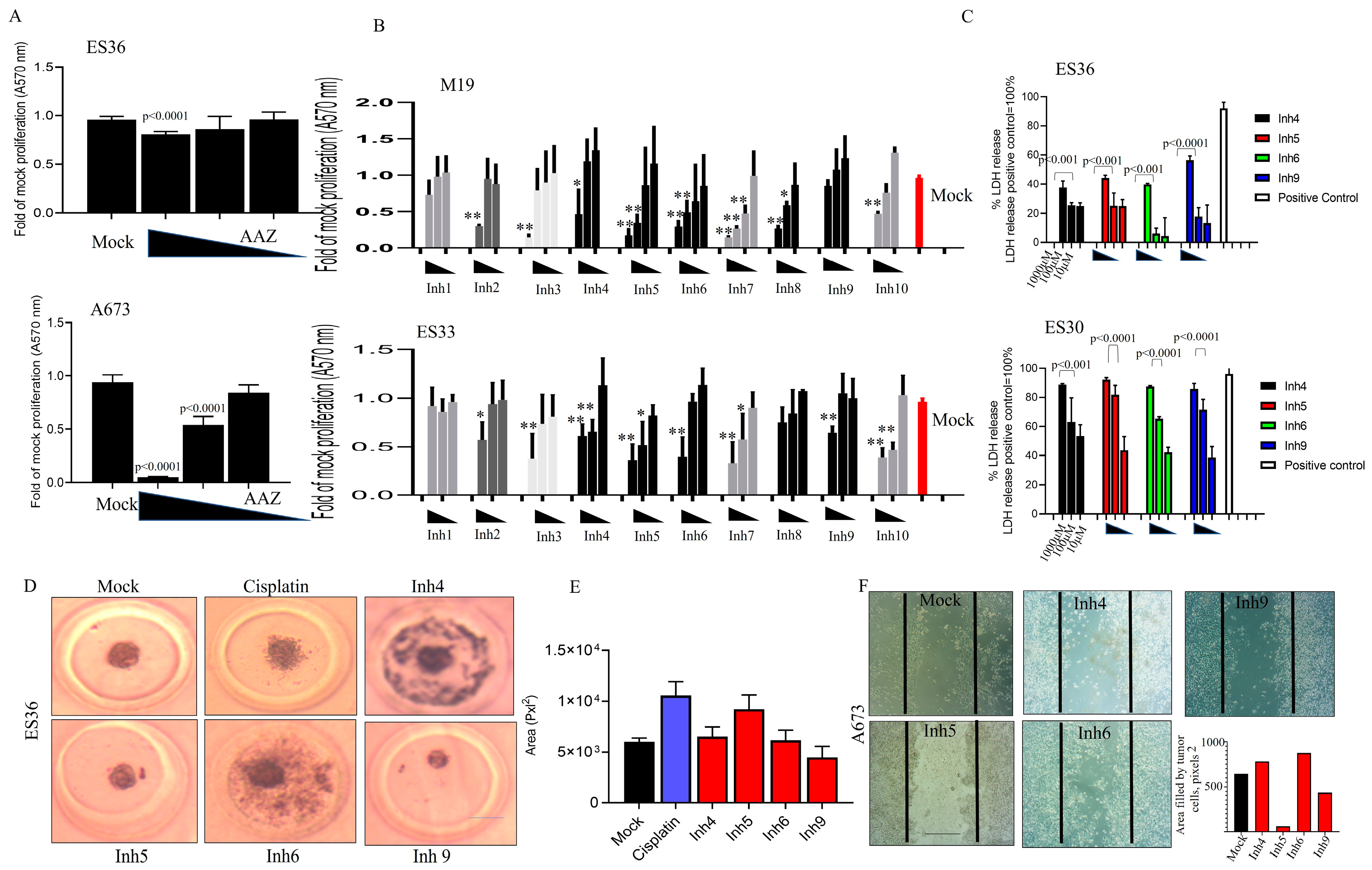
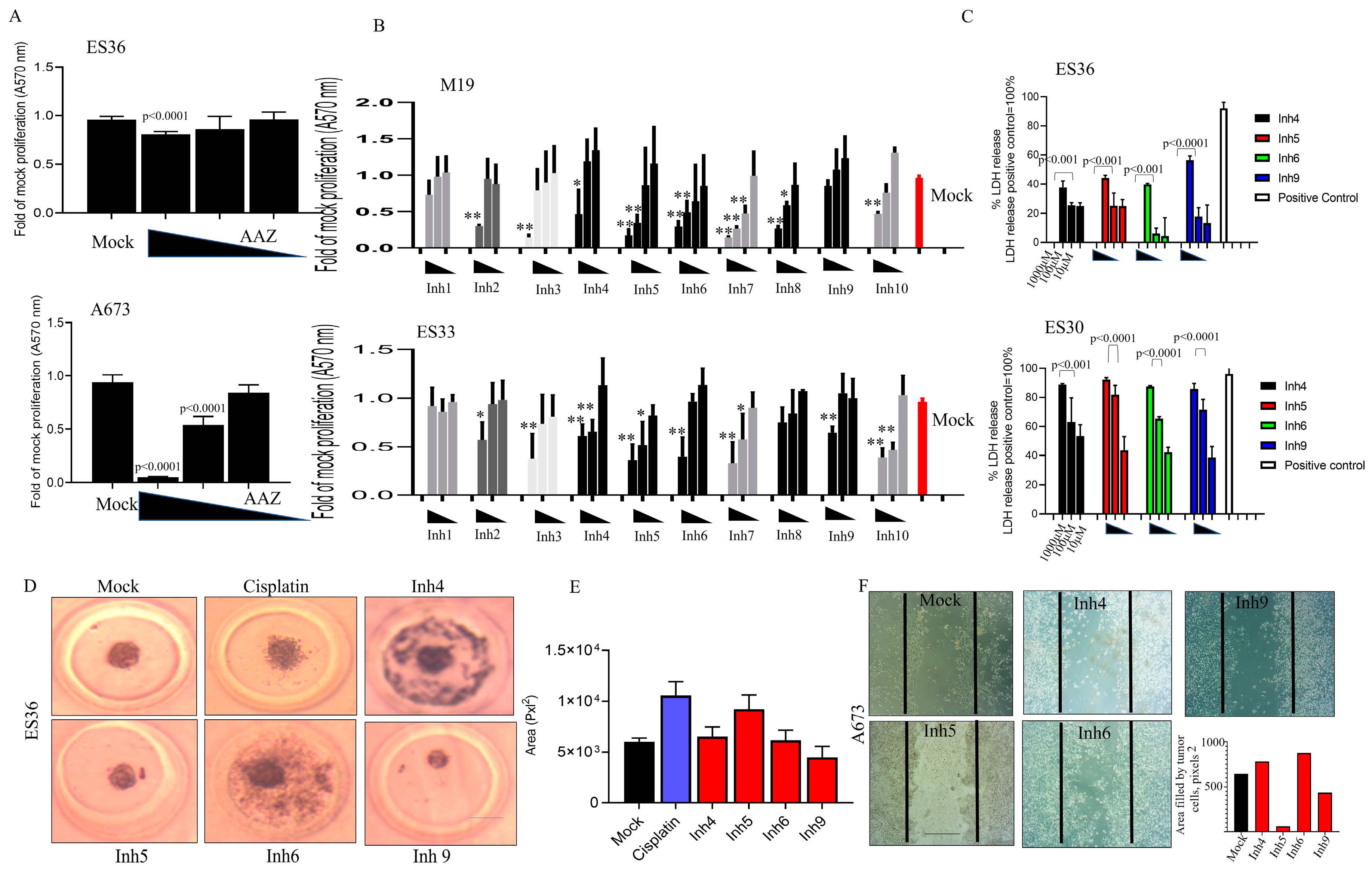
3.2. Inhibition of CAII Decreases ES Tumor Cell Proliferation
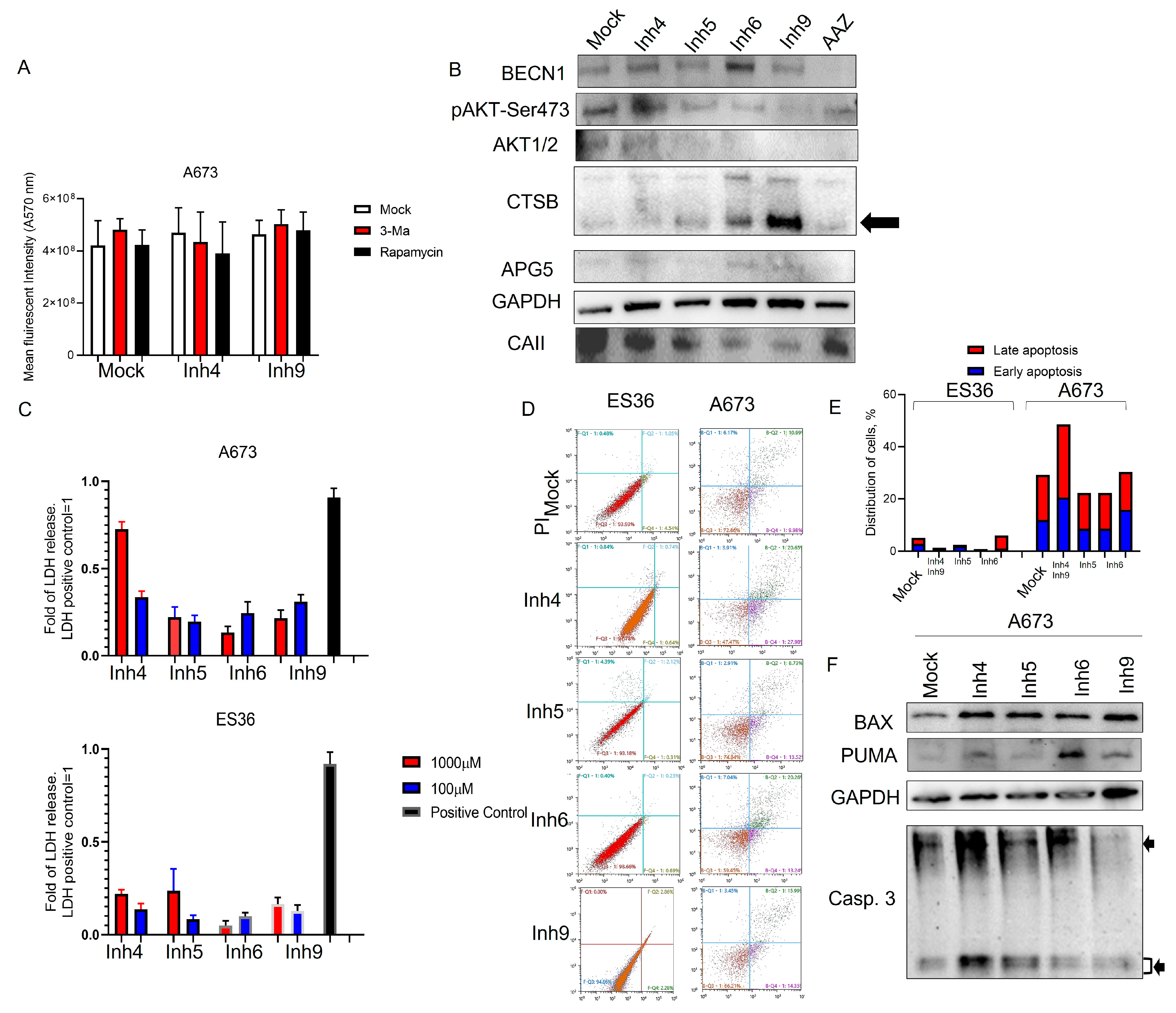
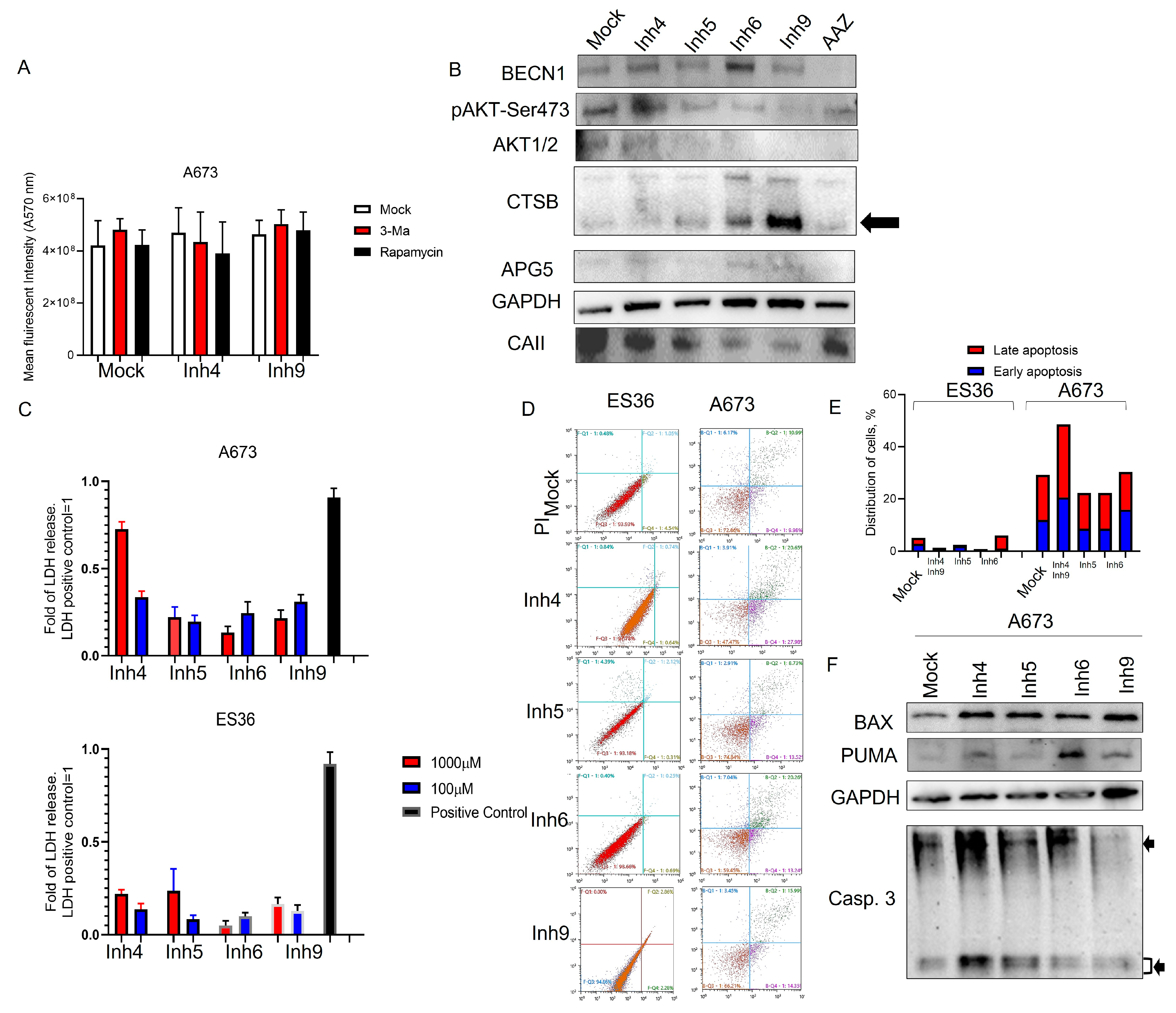
3.3. Inhibition of CA Expression Leads to Activation of Apoptotic and Autophagic Protein Expressions
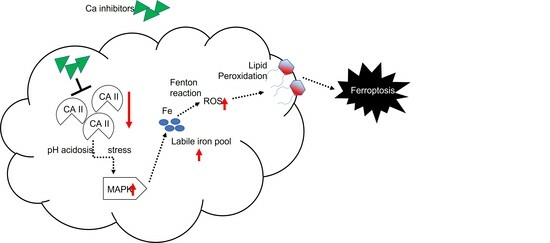
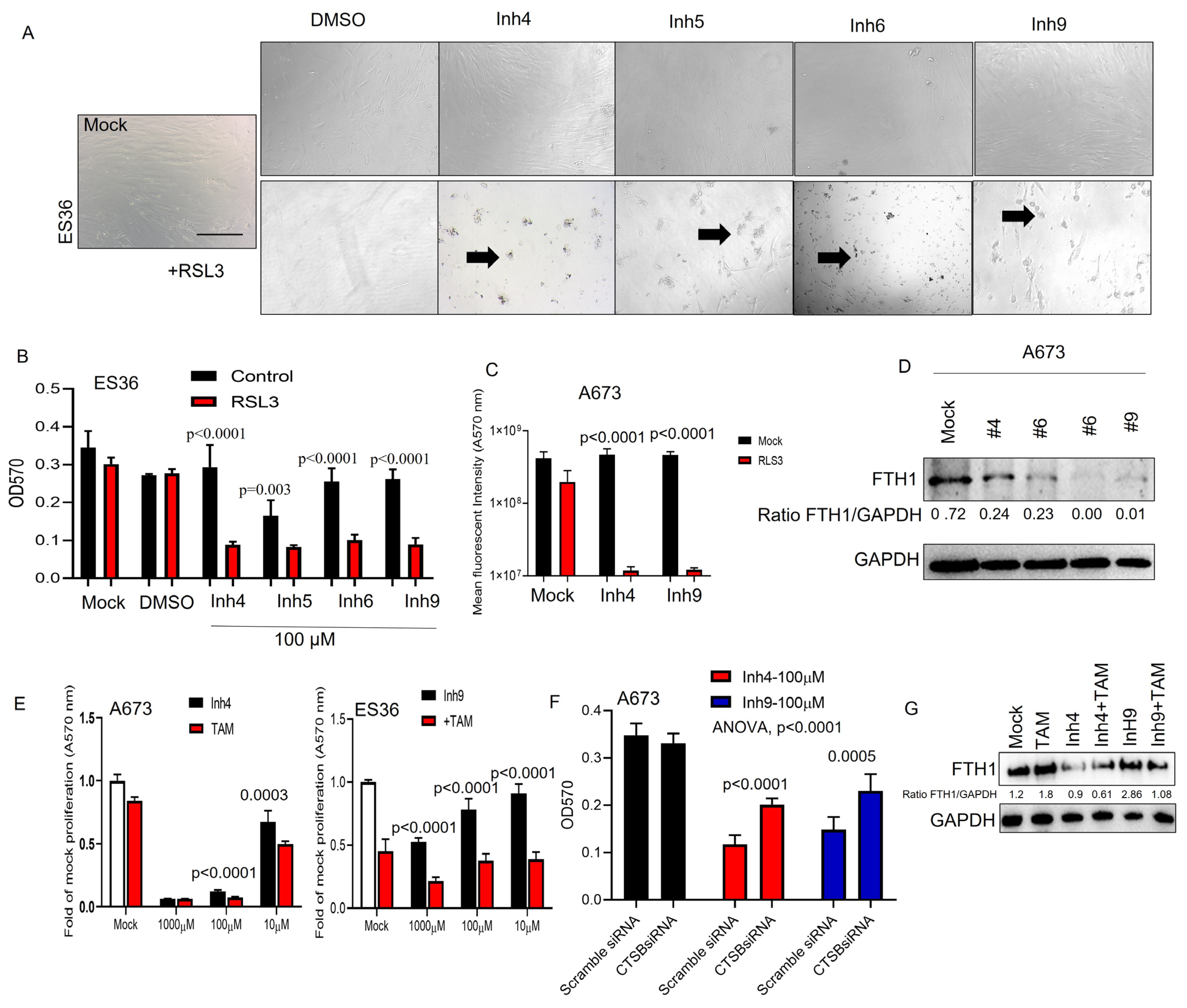
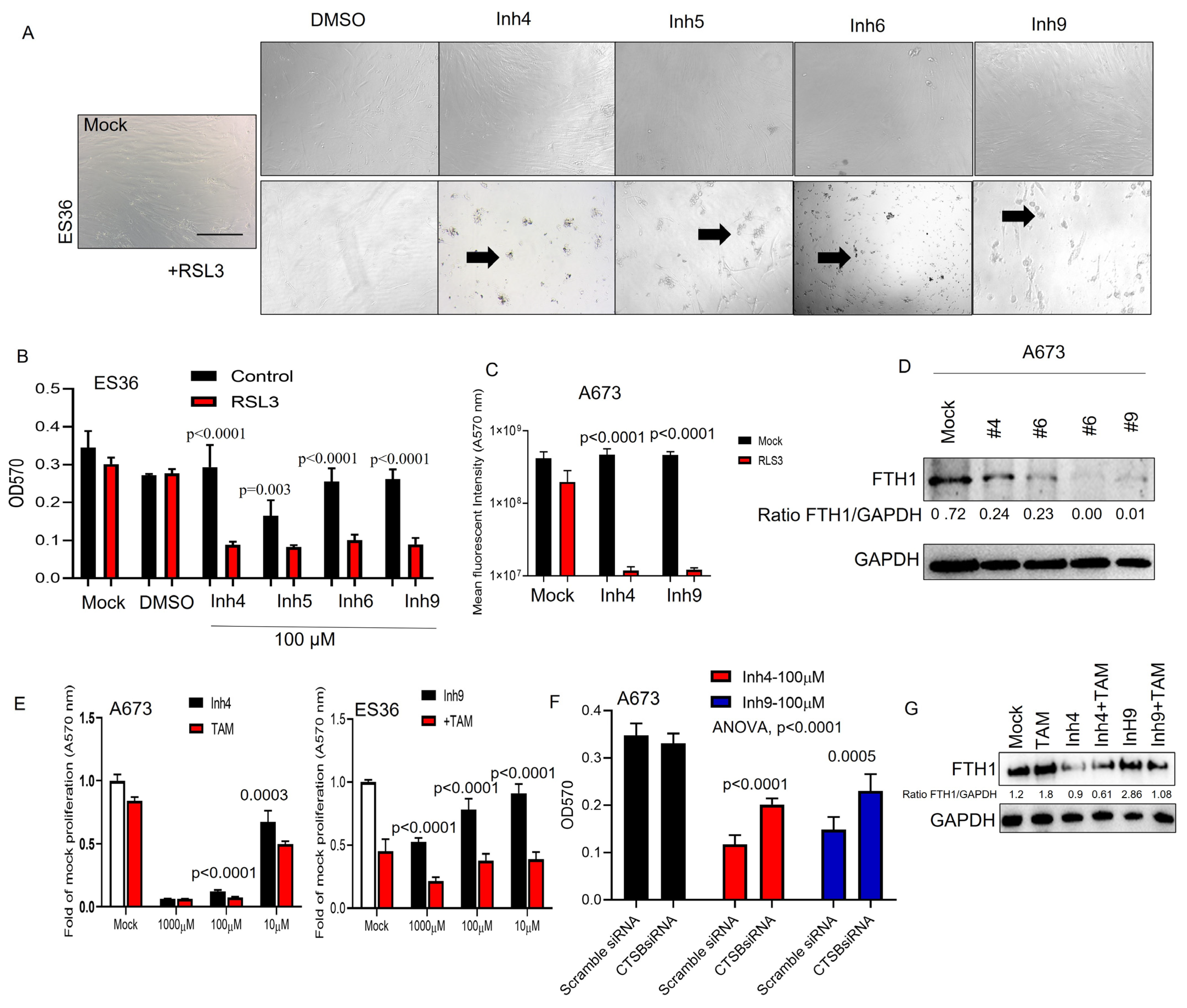
3.4. CAII Suppression Promotes Ferroptosis at ES Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AAZ | Acetazolamide |
| APBA1 | Amyloid beta precursor protein-binding family A member 1 |
| AKT1 | AKT serine/threonine kinase 1 |
| BAX | BCL2-associated X, apoptosis regulator |
| CAII | Carbonic anhydrase isoform II |
| DMSO | Dimethyl sulfoxide |
| GAPDH | Glyceraldehyde-3-Phosphate Dehydrogenase |
| IGF-1R | Insulin-like growth factor 1 receptor |
| IGF-1 | Insulin-like growth factor 1 |
| IGFBP-1 | Insulin-like growth factor binding protein 1 |
| IGFBP-3 | Insulin-like growth factor binding protein 3 |
| FTH1 | Ferritin heavy chain 1 |
| LDH | Lactate dehydrogenase A |
| NLGN4X | Neuroligin 4 X-linked |
| ES | Ewing sarcoma |
| PI3K | Phosphoinositide 3-kinase |
| PUMA | BH3-containing protein |
| SOX2 | SRY-box transcription factor 2 |
| TAM | Tamoxifen |
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Durer, S.; Shaikh, H. Ewing Sarcoma. In StatPearls; StatPearls: Treasure Island, FL, USA, 2023. [Google Scholar]
- Esiashvili, N. Radiation Oncology in the Developing Economies of Central and Eastern Europe. Semin. Radiat. Oncol. 2017, 27, 150–157. [Google Scholar] [CrossRef] [PubMed]
- Albergo, J.I.; Gaston, C.L.; Laitinen, M.; Darbyshire, A.; Jeys, L.M.; Sumathi, V.; Parry, M.; Peake, D.; Carter, S.R.; Tillman, R.; et al. Ewing’s sarcoma: Only patients with 100% of necrosis after chemotherapy should be classified as having a good response. Bone Joint J. 2016, 98B, 1138–1144. [Google Scholar] [CrossRef]
- Fayzullina, D.; Tsibulnikov, S.; Stempen, M.; Schroeder, B.A.; Kumar, N.; Kharwar, R.K.; Acharya, A.; Timashev, P.; Ulasov, I. Novel Targeted Therapeutic Strategies for Ewing Sarcoma. Cancers 2022, 14, 1988. [Google Scholar] [CrossRef]
- Vaezi, M.A.; Eghtedari, A.R.; Safizadeh, B.; Babaheidarian, P.; Salimi, V.; Adjaminezhad-Fard, F.; Yarahmadi, S.; Mirzaei, A.; Rahbar, M.; Tavakoli-Yaraki, M. Evaluating the local expression pattern of IGF-1R in tumor tissues and the circulating levels of IGF-1, IGFBP-1, and IGFBP-3 in the blood of patients with different primary bone tumors. Front. Oncol. 2022, 12, 1096438. [Google Scholar] [CrossRef]
- Roundhill, E.A.; Chicon-Bosch, M.; Jeys, L.; Parry, M.; Rankin, K.S.; Droop, A.; Burchill, S.A. RNA sequencing and functional studies of patient-derived cells reveal that neurexin-1 and regulators of this pathway are associated with poor outcomes in Ewing sarcoma. Cell Oncol. 2021, 44, 1065–1085. [Google Scholar] [CrossRef]
- Sannino, G.; Marchetto, A.; Ranft, A.; Jabar, S.; Zacherl, C.; Alba-Rubio, R.; Stein, S.; Wehweck, F.S.; Kiran, M.M.; Holting, T.L.B.; et al. Gene expression and immunohistochemical analyses identify SOX2 as major risk factor for overall survival and relapse in Ewing sarcoma patients. EBioMedicine 2019, 47, 156–162. [Google Scholar] [CrossRef]
- Benassi, M.S.; Rimondi, E.; Balladelli, A.; Ghinelli, C.; Magagnoli, G.; Vanel, D. The role of imaging for translational research in bone tumors. Eur. J. Radiol. 2013, 82, 2115–2123. [Google Scholar] [CrossRef]
- Yakushov, S.; Menyailo, M.; Denisov, E.; Karlina, I.; Zainullina, V.; Kirgizov, K.; Romantsova, O.; Timashev, P.; Ulasov, I. Identification of Factors Driving Doxorubicin-Resistant Ewing Tumor Cells to Survival. Cancers 2022, 14, 5498. [Google Scholar] [CrossRef]
- Vasileva, E.; Warren, M.; Triche, T.J.; Amatruda, J.F. Dysregulated heparan sulfate proteoglycan metabolism promotes Ewing sarcoma tumor growth. Elife 2022, 11, e69734. [Google Scholar] [CrossRef]
- Ruiz-Pinto, S.; Pita, G.; Patino-Garcia, A.; Garcia-Miguel, P.; Alonso, J.; Perez-Martinez, A.; Sastre, A.; Gomez-Mariano, G.; Lissat, A.; Scotlandi, K.; et al. Identification of genetic variants in pharmacokinetic genes associated with Ewing Sarcoma treatment outcome. Ann. Oncol. 2016, 27, 1788–1793. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.K.; Lin, Y.M.; Lo, C.M.; Tang, C.H.; Teng, C.L.; Chao, W.T.; Wu, M.H.; Liu, C.S.; Hsieh, M. Oncogenic roles of carbonic anhydrase 8 in human osteosarcoma cells. Tumour Biol. 2016, 37, 7989–8005. [Google Scholar] [CrossRef] [PubMed]
- Nazon, C.; Pierrevelcin, M.; Willaume, T.; Lhermitte, B.; Weingertner, N.; Marco, A.D.; Bund, L.; Vincent, F.; Bierry, G.; Gomez-Brouchet, A.; et al. Together Intra-Tumor Hypoxia and Macrophagic Immunity Are Driven Worst Outcome in Pediatric High-Grade Osteosarcomas. Cancers 2022, 14, 1482. [Google Scholar] [CrossRef]
- Zhang, D.; Cui, G.; Sun, C.; Lei, L.; Lei, L.; Williamson, R.A.; Wang, Y.; Zhang, J.; Chen, P.; Wang, A.; et al. Hypoxia promotes osteosarcoma cell proliferation and migration through enhancing platelet-derived growth factor-BB/platelet-derived growth factor receptor-beta axis. Biochem. Biophys. Res. Commun. 2019, 512, 360–366. [Google Scholar] [CrossRef] [PubMed]
- Forker, L.; Gaunt, P.; Sioletic, S.; Shenjere, P.; Potter, R.; Roberts, D.; Irlam, J.; Valentine, H.; Hughes, D.; Hughes, A.; et al. The hypoxia marker CAIX is prognostic in the UK phase III VorteX-Biobank cohort: An important resource for translational research in soft tissue sarcoma. Br. J. Cancer 2018, 118, 698–704. [Google Scholar] [CrossRef]
- Kim, J.I.; Choi, K.U.; Lee, I.S.; Choi, Y.J.; Kim, W.T.; Shin, D.H.; Kim, K.; Lee, J.H.; Kim, J.Y.; Sol, M.Y. Expression of hypoxic markers and their prognostic significance in soft tissue sarcoma. Oncol. Lett. 2015, 9, 1699–1706. [Google Scholar] [CrossRef]
- Parkkila, S.; Lasota, J.; Fletcher, J.A.; Ou, W.B.; Kivela, A.J.; Nuorva, K.; Parkkila, A.K.; Ollikainen, J.; Sly, W.S.; Waheed, A.; et al. Carbonic anhydrase II. A novel biomarker for gastrointestinal stromal tumors. Mod. Pathol. 2010, 23, 743–750. [Google Scholar] [CrossRef]
- Krasavin, M.; Shetnev, A.; Sharonova, T.; Baykov, S.; Tuccinardi, T.; Kalinin, S.; Angeli, A.; Supuran, C.T. Heterocyclic periphery in the design of carbonic anhydrase inhibitors: 1,2,4-Oxadiazol-5-yl benzenesulfonamides as potent and selective inhibitors of cytosolic hCA II and membrane-bound hCA IX isoforms. Bioorg. Chem. 2018, 76, 88–97. [Google Scholar] [CrossRef]
- Krasavin, M.; Shetnev, A.; Sharonova, T.; Baykov, S.; Kalinin, S.; Nocentini, A.; Sharoyko, V.; Poli, G.; Tuccinardi, T.; Presnukhina, S.; et al. Continued exploration of 1,2,4-oxadiazole periphery for carbonic anhydrase-targeting primary arene sulfonamides: Discovery of subnanomolar inhibitors of membrane-bound hCA IX isoform that selectively kill cancer cells in hypoxic environment. Eur. J. Med. Chem. 2019, 164, 92–105. [Google Scholar] [CrossRef]
- Krasavin, M.; Shetnev, A.; Baykov, S.; Kalinin, S.; Nocentini, A.; Sharoyko, V.; Poli, G.; Tuccinardi, T.; Korsakov, M.; Tennikova, T.B.; et al. Pyridazinone-substituted benzenesulfonamides display potent inhibition of membrane-bound human carbonic anhydrase IX and promising antiproliferative activity against cancer cell lines. Eur. J. Med. Chem. 2019, 168, 301–314. [Google Scholar] [CrossRef]
- Sharonova, T.; Paramonova, P.; Kalinin, S.; Bunev, A.; Gasanov Rcapital Ie, C.; Nocentini, A.; Sharoyko, V.; Tennikova, T.B.; Dar’in, D.; Supuran, C.T.; et al. Insertion of metal carbenes into the anilinic N-H bond of unprotected aminobenzenesulfonamides delivers low nanomolar inhibitors of human carbonic anhydrase IX and XII isoforms. Eur. J. Med. Chem. 2021, 218, 113352. [Google Scholar] [CrossRef]
- Ulasov, I.; Borovjagin, A.; Fares, J.; Yakushov, S.; Malin, D.; Timashev, P.; Lesniak, M.S. MicroRNA 345 (miR345) regulates KISS1-E-cadherin functional interaction in breast cancer brain metastases. Cancer Lett. 2020, 481, 24–31. [Google Scholar] [CrossRef]
- Chakraborty, S.; Bhat, A.M.; Mushtaq, I.; Luan, H.; Kalluchi, A.; Mirza, S.; Storck, M.D.; Chaturvedi, N.; Lopez-Guerrero, J.A.; Llombart-Bosch, A.; et al. EHD1-dependent traffic of IGF-1 receptor to the cell surface is essential for Ewing sarcoma tumorigenesis and metastasis. Commun. Biol. 2023, 6, 758. [Google Scholar] [CrossRef]
- Wang, Z.; Sun, L.; Liu, S.; Jiang, H. TIPE1 inhibits the growth of Ewing’s sarcoma cells by suppressing Wnt/beta-catenin signaling. Clin. Transl. Oncol. 2023, 25, 1332–1339. [Google Scholar] [CrossRef]
- Palombo, R.; Passacantilli, I.; Terracciano, F.; Capone, A.; Matteocci, A.; Tournier, S.; Alberdi, A.; Chiurchiu, V.; Volpe, E.; Paronetto, M.P. Inhibition of the PI3K/AKT/mTOR signaling promotes an M1 macrophage switch by repressing the ATF3-CXCL8 axis in Ewing sarcoma. Cancer Lett. 2023, 555, 216042. [Google Scholar] [CrossRef]
- Supuran, C.T. Acetazolamide for the treatment of idiopathic intracranial hypertension. Expert. Rev. Neurother. 2015, 15, 851–856. [Google Scholar] [CrossRef]
- Li, H.B.; Chen, J.K.; Su, Z.X.; Jin, Q.L.; Deng, L.W.; Huang, G.; Shen, J.N. Cordycepin augments the chemosensitivity of osteosarcoma to cisplatin by activating AMPK and suppressing the AKT signaling pathway. Cancer Cell Int. 2021, 21, 706. [Google Scholar] [CrossRef]
- Yu, H.; Hou, Z.; Tian, Y.; Mou, Y.; Guo, C. Design, synthesis, cytotoxicity and mechanism of novel dihydroartemisinin-coumarin hybrids as potential anti-cancer agents. Eur. J. Med. Chem. 2018, 151, 434–449. [Google Scholar] [CrossRef]
- Li, Z.; Jiang, L.; Chew, S.H.; Hirayama, T.; Sekido, Y.; Toyokuni, S. Carbonic anhydrase 9 confers resistance to ferroptosis/apoptosis in malignant mesothelioma under hypoxia. Redox Biol. 2019, 26, 101297. [Google Scholar] [CrossRef]
- Liu, N.; Lin, Q.; Zuo, W.; Chen, W.; Huang, S.; Han, Y.; Liang, X.J.; Zhu, X.; Huo, S. Carbonic anhydrase IX-targeted nanovesicles potentiated ferroptosis by remodeling the intracellular environment for synergetic cancer therapy. Nanoscale Horiz. 2023, 8, 783–793. [Google Scholar] [CrossRef]
- Venkateswaran, G.; McDonald, P.C.; Chafe, S.C.; Brown, W.S.; Gerbec, Z.J.; Awrey, S.J.; Parker, S.J.; Dedhar, S. A Carbonic Anhydrase IX/SLC1A5 axis regulates glutamine metabolism dependent ferroptosis in hypoxic tumor cells. Mol. Cancer Ther. 2023, 22, 1228–1242. [Google Scholar] [CrossRef]
- Huang, W.; He, Y.; Yang, S.; Xue, X.; Qin, H.; Sun, T.; Yang, W. CA9 knockdown enhanced ionizing radiation-induced ferroptosis and radiosensitivity of hypoxic glioma cells. Int. J. Radiat. Biol. 2023, 1–17. [Google Scholar] [CrossRef]
- Huang, L.; Zhu, J.; Xiong, W.; Feng, J.; Yang, J.; Lu, X.; Lu, Y.; Zhang, Q.; Yi, P.; Feng, Y.; et al. Tumor-Generated Reactive Oxygen Species Storm for High-Performance Ferroptosis Therapy. ACS Nano 2023, 17, 11492–11506. [Google Scholar] [CrossRef] [PubMed]
- Teicher, B.A.; Liu, S.D.; Liu, J.T.; Holden, S.A.; Herman, T.S. A carbonic anhydrase inhibitor as a potential modulator of cancer therapies. Anticancer Res. 1993, 13, 1549–1556. [Google Scholar]
- Kaluz, S.; Kaluzova, M.; Chrastina, A.; Olive, P.L.; Pastorekova, S.; Pastorek, J.; Lerman, M.I.; Stanbridge, E.J. Lowered oxygen tension induces expression of the hypoxia marker MN/carbonic anhydrase IX in the absence of hypoxia-inducible factor 1 alpha stabilization: A role for phosphatidylinositol 3′-kinase. Cancer Res. 2002, 62, 4469–4477. [Google Scholar]
- Supuran, C.T. Structure and function of carbonic anhydrases. Biochem. J. 2016, 473, 2023–2032. [Google Scholar] [CrossRef]
- Aspatwar, A.; Tolvanen, M.E.; Parkkila, S. An update on carbonic anhydrase-related proteins VIII, X and XI. J. Enzym. Inhib. Med. Chem. 2013, 28, 1129–1142. [Google Scholar] [CrossRef]
- Abdou, A.G.; Kandil, M.; Elshakhs, S.; El-Dien, M.S.; Abdallah, R. Renal cell carcinoma with rhabdoid and sarcomatoid features presented as a metastatic thigh mass with an unusual immunohistochemical profile. Rare Tumors 2014, 6, 5037. [Google Scholar] [CrossRef]
- Dong, X.; Wang, Y.; Zhuang, H.; An, G. Hydroxygenkwanin suppresses proliferation, invasion and migration of osteosarcoma cells via the miR-320a/SOX9 axis. Mol. Med. Rep. 2022, 26, 299. [Google Scholar] [CrossRef]
- Zhang, C.; Lu, X.; Liu, X.; Xu, J.; Li, J.; Qu, T.; Dai, J.; Guo, R. Carbonic Anhydrase IX Controls Vulnerability to Ferroptosis in Gefitinib-Resistant Lung Cancer. Oxid. Med. Cell Longev. 2023, 2023, 1367938. [Google Scholar] [CrossRef] [PubMed]
- Kalinin, S.; Malkova, A.; Sharonova, T.; Sharoyko, V.; Bunev, A.; Supuran, C.T.; Krasavin, M. Carbonic Anhydrase IX Inhibitors as Candidates for Combination Therapy of Solid Tumors. Int. J. Mol. Sci. 2021, 22, 13405. [Google Scholar] [CrossRef]
- Demuynck, R.; Efimova, I.; Naessens, F.; Krysko, D.V. Immunogenic ferroptosis and where to find it? J. Immunother. Cancer 2021, 9, e003430. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fayzullina, D.; Yakushov, S.; Kantserova, K.; Belyaeva, E.; Aniskin, D.; Tsibulnikov, S.; Fayzullina, N.; Kalinin, S.; Romantsova, O.; Timashev, P.S.; et al. Carbonic Anhydrase Inhibitors Induce Ferroptosis through Inhibition of AKT/FTH1 Signaling in Ewing Sarcoma Tumor Cells. Cancers 2023, 15, 5225. https://doi.org/10.3390/cancers15215225
Fayzullina D, Yakushov S, Kantserova K, Belyaeva E, Aniskin D, Tsibulnikov S, Fayzullina N, Kalinin S, Romantsova O, Timashev PS, et al. Carbonic Anhydrase Inhibitors Induce Ferroptosis through Inhibition of AKT/FTH1 Signaling in Ewing Sarcoma Tumor Cells. Cancers. 2023; 15(21):5225. https://doi.org/10.3390/cancers15215225
Chicago/Turabian StyleFayzullina, Darya, Semyon Yakushov, Kamilla Kantserova, Elizaveta Belyaeva, Denis Aniskin, Sergey Tsibulnikov, Nafisa Fayzullina, Stanislav Kalinin, Olga Romantsova, Peter S. Timashev, and et al. 2023. "Carbonic Anhydrase Inhibitors Induce Ferroptosis through Inhibition of AKT/FTH1 Signaling in Ewing Sarcoma Tumor Cells" Cancers 15, no. 21: 5225. https://doi.org/10.3390/cancers15215225
APA StyleFayzullina, D., Yakushov, S., Kantserova, K., Belyaeva, E., Aniskin, D., Tsibulnikov, S., Fayzullina, N., Kalinin, S., Romantsova, O., Timashev, P. S., Schroeder, B. A., & Ulasov, I. V. (2023). Carbonic Anhydrase Inhibitors Induce Ferroptosis through Inhibition of AKT/FTH1 Signaling in Ewing Sarcoma Tumor Cells. Cancers, 15(21), 5225. https://doi.org/10.3390/cancers15215225