
Deletions of CDKN2A and MTAP Detected by Copy-Number Variation Array Are Associated with Loss of p16 and MTAP Protein in Pleural Mesothelioma
 , , , , and
, , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Tissue Samples and Tissue Microarray Construction
2.2. Copy-Number Variation Arrays (Oncoscan)
2.3. Immunohistochemistry for p16 and MTAP
2.4. Statistical Analysis
3. Results
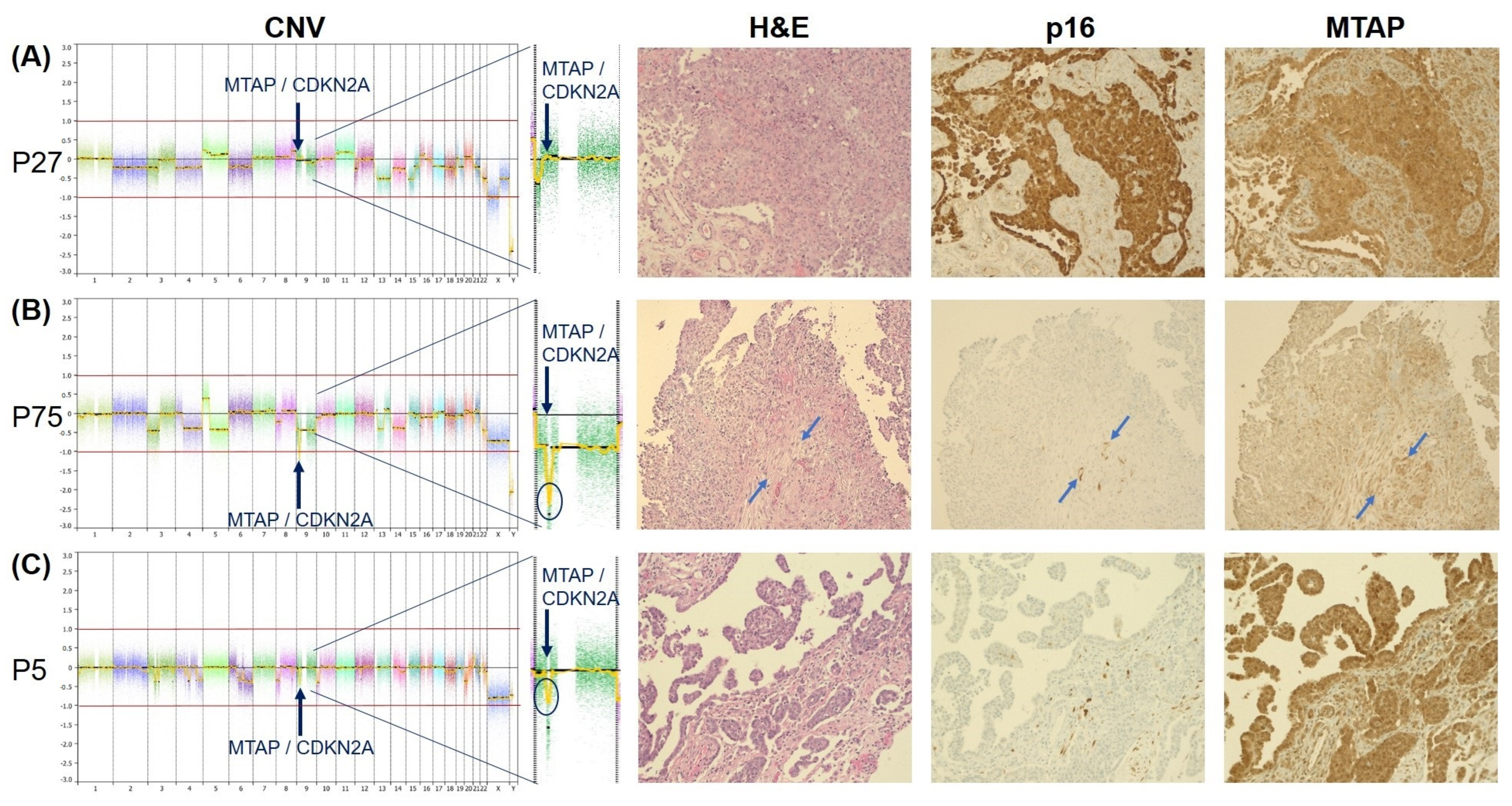
3.1. Copy-Number Variation Array (Oncoscan)
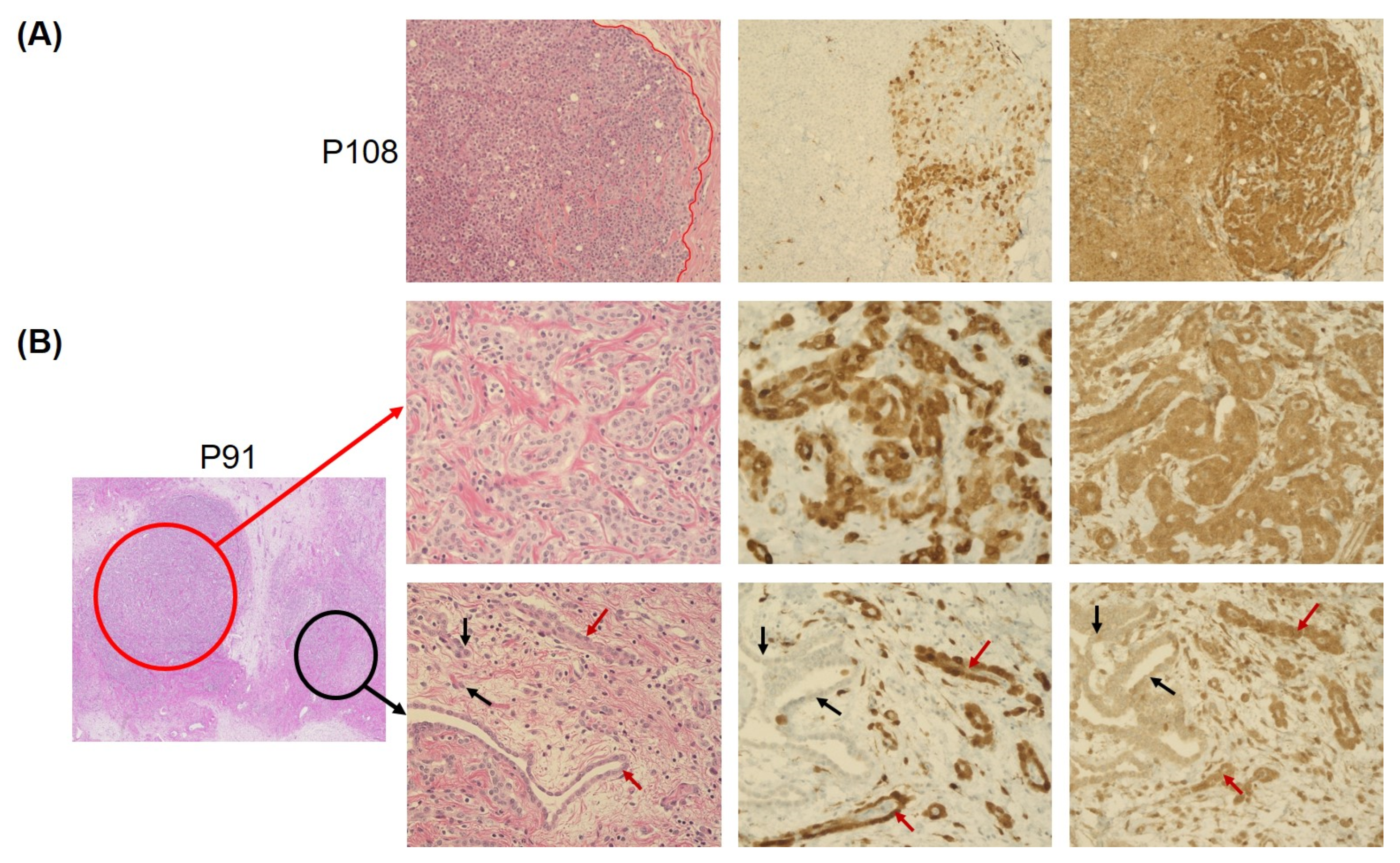
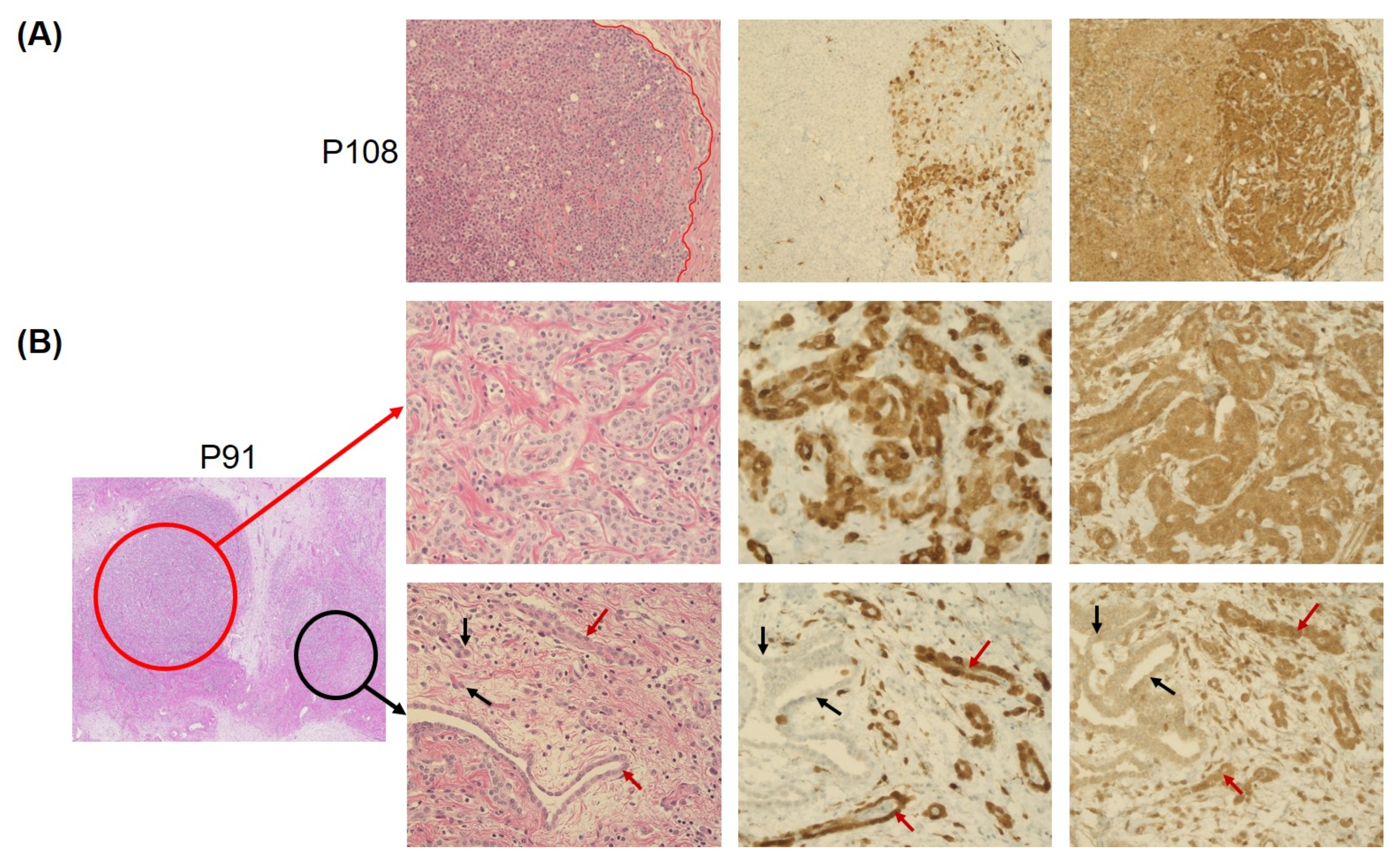
3.2. Concordance of Immunohistochemistry and Copy-Number Variation Array
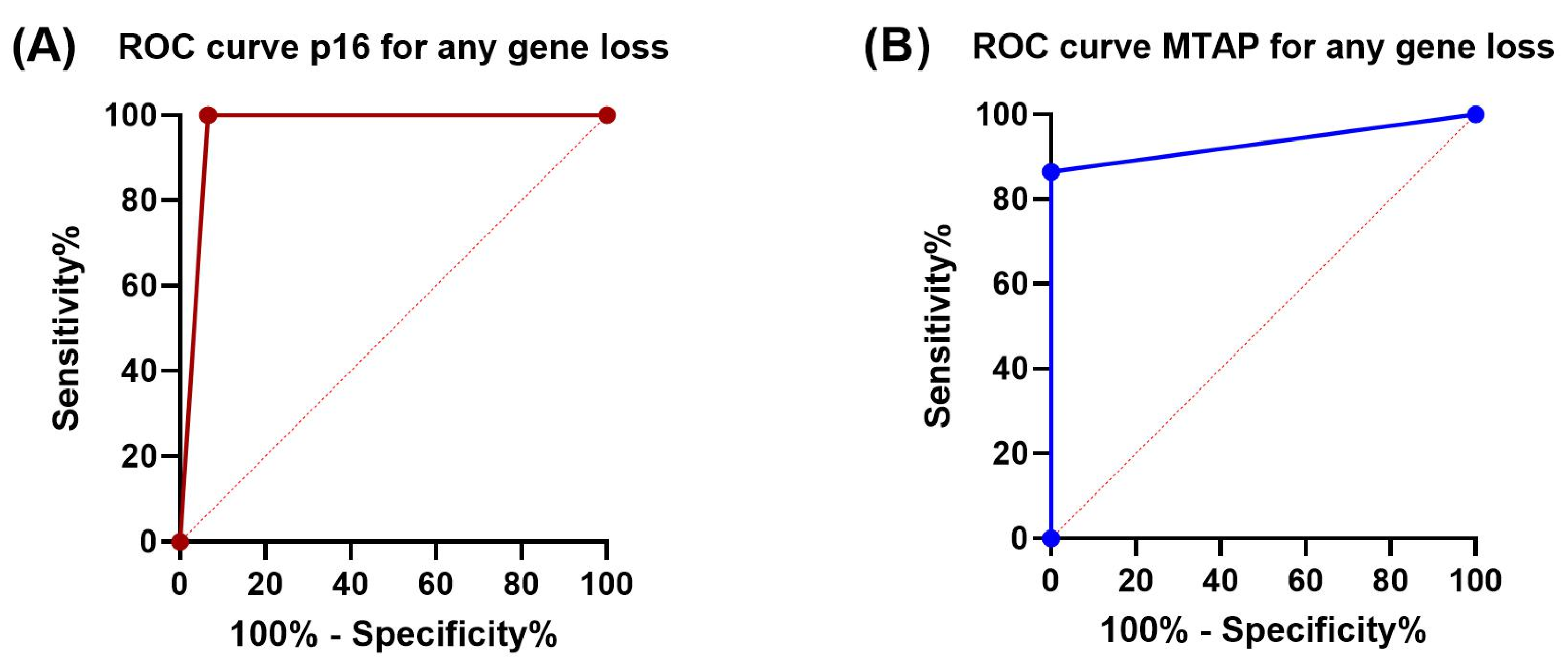
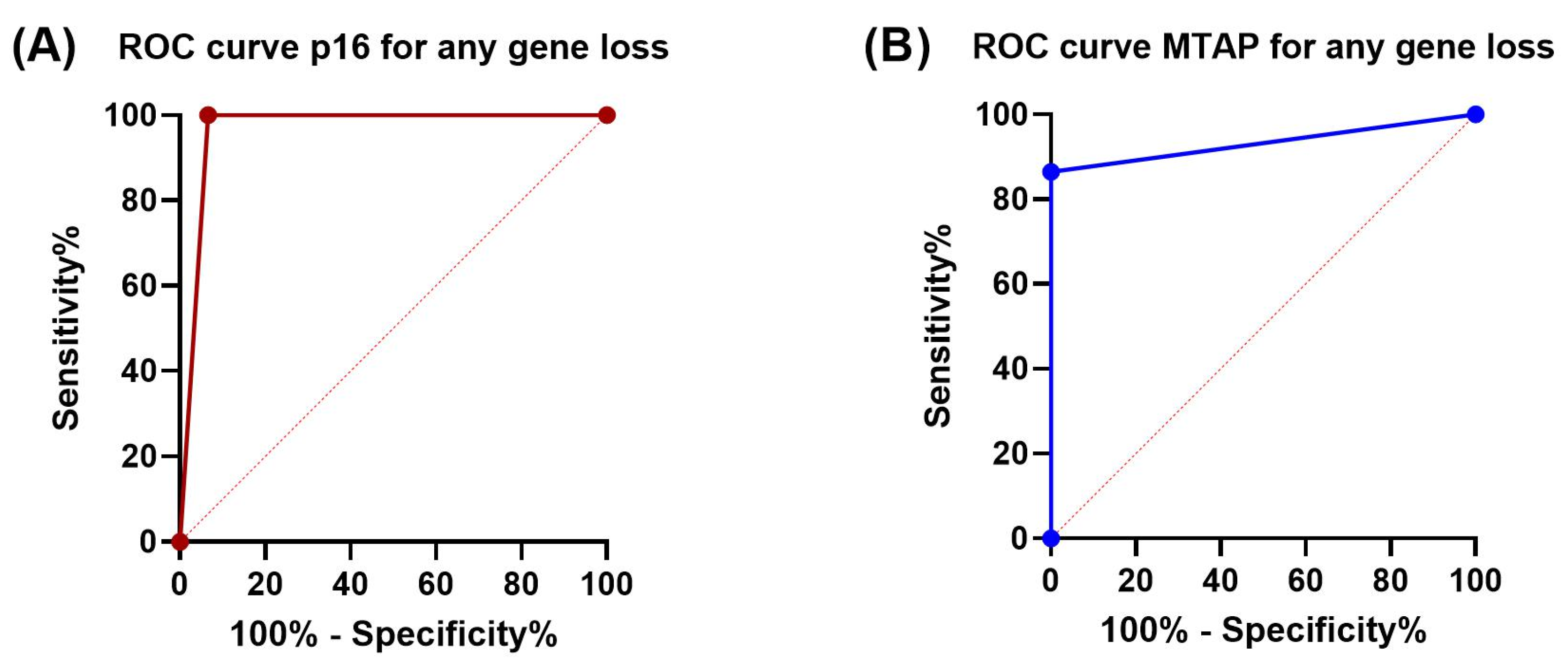
3.3. Sensitivity and Specificity of p16 and MTAP Immunohistochemistry
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Spirtas, R.; Heineman, E.F.; Bernstein, L.; Beebe, G.W.; Keehn, R.J.; Stark, A.; Harlow, B.L.; Benichou, J. Malignant mesothelioma: Attributable risk of asbestos exposure. Occup. Environ. Med. 1994, 51, 804–811. [Google Scholar] [CrossRef]
- Husain, A.N.; Colby, T.V.; Ordóñez, N.G.; Allen, T.C.; Attanoos, R.L.; Beasley, M.B.; Butnor, K.J.; Chirieac, L.R.; Churg, A.M.; Dacic, S.; et al. Guidelines for Pathologic Diagnosis of Malignant Mesothelioma 2017 Update of the Consensus Statement From the International Mesothelioma Interest Group. Arch. Pathol. Lab. Med. 2018, 142, 89–108. [Google Scholar] [CrossRef] [PubMed]
- Oehl, K.; Vrugt, B.; Wagner, U.; Kirschner, M.B.; Meerang, M.; Weder, W.; Felley-Bosco, E.; Wollscheid, B.; Bankov, K.; Demes, M.C.; et al. Alterations in BAP1 Are Associated with Cisplatin Resistance through Inhibition of Apoptosis in Malignant Pleural Mesothelioma. Clin. Cancer Res. 2021, 27, 2277–2291. [Google Scholar] [CrossRef]
- Nasu, M.; Emi, M.; Pastorino, S.; Tanji, M.; Powers, A.; Luk, H.; Baumann, F.; Zhang, Y.A.; Gazdar, A.; Kanodia, S.; et al. High Incidence of Somatic BAP1 alterations in sporadic malignant mesothelioma. J. Thorac. Oncol. 2015, 10, 565–576. [Google Scholar] [CrossRef] [PubMed]
- Berg, K.B.; Dacic, S.; Miller, C.; Cheung, S.; Churg, A. Utility of Methylthioadenosine Phosphorylase Compared With BAP1 Immunohistochemistry, and CDKN2A and NF2 Fluorescence In Situ Hybridization in Separating Reactive Mesothelial Proliferations From Epithelioid Malignant Mesotheliomas. Arch. Pathol. Lab. Med. 2018, 142, 1549–1553. [Google Scholar] [CrossRef] [PubMed]
- Chiosea, S.; Krasinskas, A.; Cagle, P.T.; Mitchell, K.A.; Zander, D.S.; Dacic, S. Diagnostic importance of 9p21 homozygous deletion in malignant mesotheliomas. Mod. Pathol. 2008, 21, 742–747. [Google Scholar] [CrossRef] [PubMed]
- Hida, T.; Hamasaki, M.; Matsumoto, S.; Sato, A.; Tsujimura, T.; Kawahara, K.; Iwasaki, A.; Okamoto, T.; Oda, Y.; Honda, H.; et al. BAP1 immunohistochemistry and p16 FISH results in combination provide higher confidence in malignant pleural mesothelioma diagnosis: ROC analysis of the two tests. Pathol. Int. 2016, 66, 563–570. [Google Scholar] [CrossRef]
- Hwang, H.C.; Pyott, S.; Rodriguez, S.; Cindric, A.; Carr, A.; Michelsen, C.; Thompson, K.; Tse, C.H.; Gown, A.M.; Churg, A. BAP1 Immunohistochemistry and p16 FISH in the Diagnosis of Sarcomatous and Desmoplastic Mesotheliomas. Am. J. Surg. Pathol. 2016, 40, 714–718. [Google Scholar] [CrossRef]
- Sheffield, B.S.; Hwang, H.C.; Lee, A.F.; Thompson, K.; Rodriguez, S.; Tse, C.H.; Gown, A.M.; Churg, A. BAP1 immunohistochemistry and p16 FISH to separate benign from malignant mesothelial proliferations. Am. J. Surg. Pathol. 2015, 39, 977–982. [Google Scholar] [CrossRef]
- Liu, J.; Liao, X.; Gu, Y.; Fu, L.; Zhao, J.; Li, L.; Chen, Z.; Jiang, J. Role of p16 deletion and BAP1 loss in the diagnosis of malignant mesothelioma. J. Thorac. Dis. 2018, 10, 5522–5530. [Google Scholar] [CrossRef]
- Kryukov, G.V.; Wilson, F.H.; Ruth, J.R.; Paulk, J.; Tsherniak, A.; Marlow, S.E.; Vazquez, F.; Weir, B.A.; Fitzgerald, M.E.; Tanaka, M.; et al. MTAP deletion confers enhanced dependency on the PRMT5 arginine methyltransferase in cancer cells. Science 2016, 351, 1214–1218. [Google Scholar] [CrossRef] [PubMed]
- Illei, P.B.; Rusch, V.W.; Zakowski, M.F.; Ladanyi, M. Homozygous deletion of CDKN2A and codeletion of the methylthioadenosine phosphorylase gene in the majority of pleural mesotheliomas. Clin. Cancer Res. 2003, 9, 2108–2113. [Google Scholar] [PubMed]
- Krasinskas, A.M.; Bartlett, D.L.; Cieply, K.; Dacic, S. CDKN2A and MTAP deletions in peritoneal mesotheliomas are correlated with loss of p16 protein expression and poor survival. Mod. Pathol. 2010, 23, 531–538. [Google Scholar] [CrossRef] [PubMed]
- Hustinx, S.R.; Leoni, L.M.; Yeo, C.J.; Brown, P.N.; Goggins, M.; Kern, S.E.; Hruban, R.H.; Maitra, A. Concordant loss of MTAP and p16/CDKN2A expression in pancreatic intraepithelial neoplasia: Evidence of homozygous deletion in a noninvasive precursor lesion. Mod. Pathol. 2005, 18, 959–963. [Google Scholar] [CrossRef]
- Zimling, Z.G.; Jørgensen, A.; Santoni-Rugiu, E. The diagnostic value of immunohistochemically detected methylthioadenosine phosphorylase deficiency in malignant pleural mesotheliomas. Histopathology 2012, 60, E96–E105. [Google Scholar] [CrossRef] [PubMed]
- Hida, T.; Hamasaki, M.; Matsumoto, S.; Sato, A.; Tsujimura, T.; Kawahara, K.; Iwasaki, A.; Okamoto, T.; Oda, Y.; Honda, H.; et al. Immunohistochemical detection of MTAP and BAP1 protein loss for mesothelioma diagnosis: Comparison with 9p21 FISH and BAP1 immunohistochemistry. Lung Cancer 2017, 104, 98–105. [Google Scholar] [CrossRef]
- Churg, A.; Nabeshima, K.; Ali, G.; Bruno, R.; Fernandez-Cuesta, L.; Galateau-Salle, F. Highlights of the 14th international mesothelioma interest group meeting: Pathologic separation of benign from malignant mesothelial proliferations and histologic/molecular analysis of malignant mesothelioma subtypes. Lung Cancer 2018, 124, 95–101. [Google Scholar] [CrossRef]
- Chapel, D.B.; Schulte, J.J.; Berg, K.; Churg, A.; Dacic, S.; Fitzpatrick, C.; Galateau-Salle, F.; Hiroshima, K.; Krausz, T.; Le Stang, N.; et al. MTAP immunohistochemistry is an accurate and reproducible surrogate for CDKN2A fluorescence in situ hybridization in diagnosis of malignant pleural mesothelioma. Mod. Pathol. 2020, 33, 245–254. [Google Scholar] [CrossRef]
- Brcic, L.; Le Stang, N.; Gallob, F.; Pissaloux, D.; Sequeiros, R.; Paindavoine, S.; Pairon, J.C.; Karanian, M.; Dacic, S.; Girard, N.; et al. A Combination of MTAP and p16 Immunohistochemistry Can Substitute for CDKN2A Fluorescence In Situ Hybridization in Diagnosis and Prognosis of Pleural Mesotheliomas. Arch. Pathol. Lab. Med. 2023, 147, 313–322. [Google Scholar] [CrossRef]
- Sauter, J.L.; Dacic, S.; Galateau-Salle, F.; Attanoos, R.L.; Butnor, K.J.; Churg, A.; Husain, A.N.; Kadota, K.; Khoor, A.; Nicholson, A.G.; et al. The 2021 WHO Classification of Tumors of the Pleura: Advances Since the 2015 Classification. J. Thorac. Oncol. 2022, 17, 608–622. [Google Scholar] [CrossRef]
- Hinterberger, M.; Reineke, T.; Storz, M.; Weder, W.; Vogt, P.; Moch, H. D2-40 and calretinin—A tissue microarray analysis of 341 malignant mesotheliomas with emphasis on sarcomatoid differentiation. Mod. Pathol. 2007, 20, 248–255. [Google Scholar] [CrossRef] [PubMed]
- Chapel, D.B.; Dubuc, A.M.; Hornick, J.L.; Sholl, L.M. Correlation of methylthioadenosine phosphorylase (MTAP) protein expression with MTAP and CDKN2A copy number in malignant pleural mesothelioma. Histopathology 2021, 78, 1032–1042. [Google Scholar] [CrossRef] [PubMed]
- Chung, C.T.; Gda, C.S.; Hwang, D.M.; Ludkovski, O.; Pintilie, M.; Squire, J.A.; Tsao, M.S. FISH assay development for the detection of p16/CDKN2A deletion in malignant pleural mesothelioma. J. Clin. Pathol. 2010, 63, 630–634. [Google Scholar] [CrossRef]
- Oehl, K.; Vrugt, B.; Opitz, I.; Meerang, M. Heterogeneity in Malignant Pleural Mesothelioma. Int. J. Mol. Sci. 2018, 19, 1603. [Google Scholar] [CrossRef]
- Wu, D.; Hiroshima, K.; Matsumoto, S.; Nabeshima, K.; Yusa, T.; Ozaki, D.; Fujino, M.; Yamakawa, H.; Nakatani, Y.; Tada, Y.; et al. Diagnostic usefullness of p16/CDKN2A FISH in distinguishing sarcomatoid mesotheliomaand fibrous pleuritis. Am. J. Clin. Pathol. 2013, 139, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Destro, A.; Ceresoli, G.L.; Baryshnikova, E.; Garassino, I.; Zucali, P.A.; De Vincenzo, F.; Bianchi, P.; Morenghi, E.; Testori, A.; Alloisio, M.; et al. Gene methylation in pleural mesothelioma: Correlations with clinico-pathological features and patient’s follow-up. Lung Cancer 2008, 59, 369–376. [Google Scholar] [CrossRef]
- Kobayashi, N.; Toyooka, S.; Yanai, H.; Soh, J.; Fujimoto, N.; Yamamoto, H.; Ichihara, S.; Kimura, K.; Ichimura, K.; Sano, Y.; et al. Frequent p16 inactivation by homozygous deletion or methylation is associated with a poor prognosis in Japanese patients with pleural mesothelioma. Lung Cancer 2008, 62, 120–125. [Google Scholar] [CrossRef]
- Behrmann, I.; Wallner, S.; Komyod, W.; Heinrich, P.C.; Schuierer, M.; Buettner, R.; Bosserhoff, A.K. Characterization of methylthioadenosin phosphorylase (MTAP) expression in malignant melanoma. Am. J. Pathol. 2003, 163, 683–690. [Google Scholar] [CrossRef]
- Kadariya, Y.; Yin, B.; Tang, B.; Shinton, S.A.; Quinlivan, E.P.; Hua, X.; Klein-Szanto, A.; Al-Saleem, T.I.; Bassing, C.H.; Hardy, R.R.; et al. Mice heterozygous for germ-line mutations in methylthioadenosine phosphorylase (MTAP) die prematurely of T-cell lymphoma. Cancer Res. 2009, 69, 5961–5969. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Median Age (Range) | 60 (35–73) |
|---|---|
| Gender | |
| Male | 45 (86.5%) |
| Female | 7 (13.5%) |
| Histotype | |
| Epithelioid | 43 (82.7%) |
| Biphasic | 6 (11.5%) |
| Sarcomatoid | 3 (5.8%) |
| Pathological Stage (IMIG, 8th edition) | |
| IA | 2 (3.8%) |
| IB | 23 (44.2%) |
| II | 0 (0.0%) |
| IIIA | 8 (15.4%) |
| IIIB | 15 (28.8%) |
| unknown | 4 (7.7%) |
| Median OS from diagnosis (range) | 14.95 months (3.94–79.86) |
| Any Loss | Homozygous Loss | Heterozygous Loss | ||||
|---|---|---|---|---|---|---|
| p16 | MTAP | p16 | MTAP | p16 | MTAP | |
| Cohen’s kappa | 0.952 | 0.787 | 0.944 | 1.0 | 0.931 | 0.692 |
| Sensitivity | 100% | 86.5% | 100% | 100% | 100% | 70.6% |
| Specificity | 93.3% | 100% | 93.3% | 100% | 93.3% | 100% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vrugt, B.; Kirschner, M.B.; Meerang, M.; Oehl, K.; Wagner, U.; Soltermann, A.; Moch, H.; Opitz, I.; Wild, P.J. Deletions of CDKN2A and MTAP Detected by Copy-Number Variation Array Are Associated with Loss of p16 and MTAP Protein in Pleural Mesothelioma. Cancers 2023, 15, 4978. https://doi.org/10.3390/cancers15204978
Vrugt B, Kirschner MB, Meerang M, Oehl K, Wagner U, Soltermann A, Moch H, Opitz I, Wild PJ. Deletions of CDKN2A and MTAP Detected by Copy-Number Variation Array Are Associated with Loss of p16 and MTAP Protein in Pleural Mesothelioma. Cancers. 2023; 15(20):4978. https://doi.org/10.3390/cancers15204978
Chicago/Turabian StyleVrugt, Bart, Michaela B. Kirschner, Mayura Meerang, Kathrin Oehl, Ulrich Wagner, Alex Soltermann, Holger Moch, Isabelle Opitz, and Peter J. Wild. 2023. "Deletions of CDKN2A and MTAP Detected by Copy-Number Variation Array Are Associated with Loss of p16 and MTAP Protein in Pleural Mesothelioma" Cancers 15, no. 20: 4978. https://doi.org/10.3390/cancers15204978
APA StyleVrugt, B., Kirschner, M. B., Meerang, M., Oehl, K., Wagner, U., Soltermann, A., Moch, H., Opitz, I., & Wild, P. J. (2023). Deletions of CDKN2A and MTAP Detected by Copy-Number Variation Array Are Associated with Loss of p16 and MTAP Protein in Pleural Mesothelioma. Cancers, 15(20), 4978. https://doi.org/10.3390/cancers15204978