TIM-3 Is a Potential Immune Checkpoint Target in Cats with Mammary Carcinoma

 ,
,  and
and 
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Immunohistochemical Staining and Analysis
2.3. Measurement of Serum TIM-3
2.4. DNA Amplification and Sequence Analysis of Feline TIM-3 Gene
2.5. Statistical Analysis
3. Results
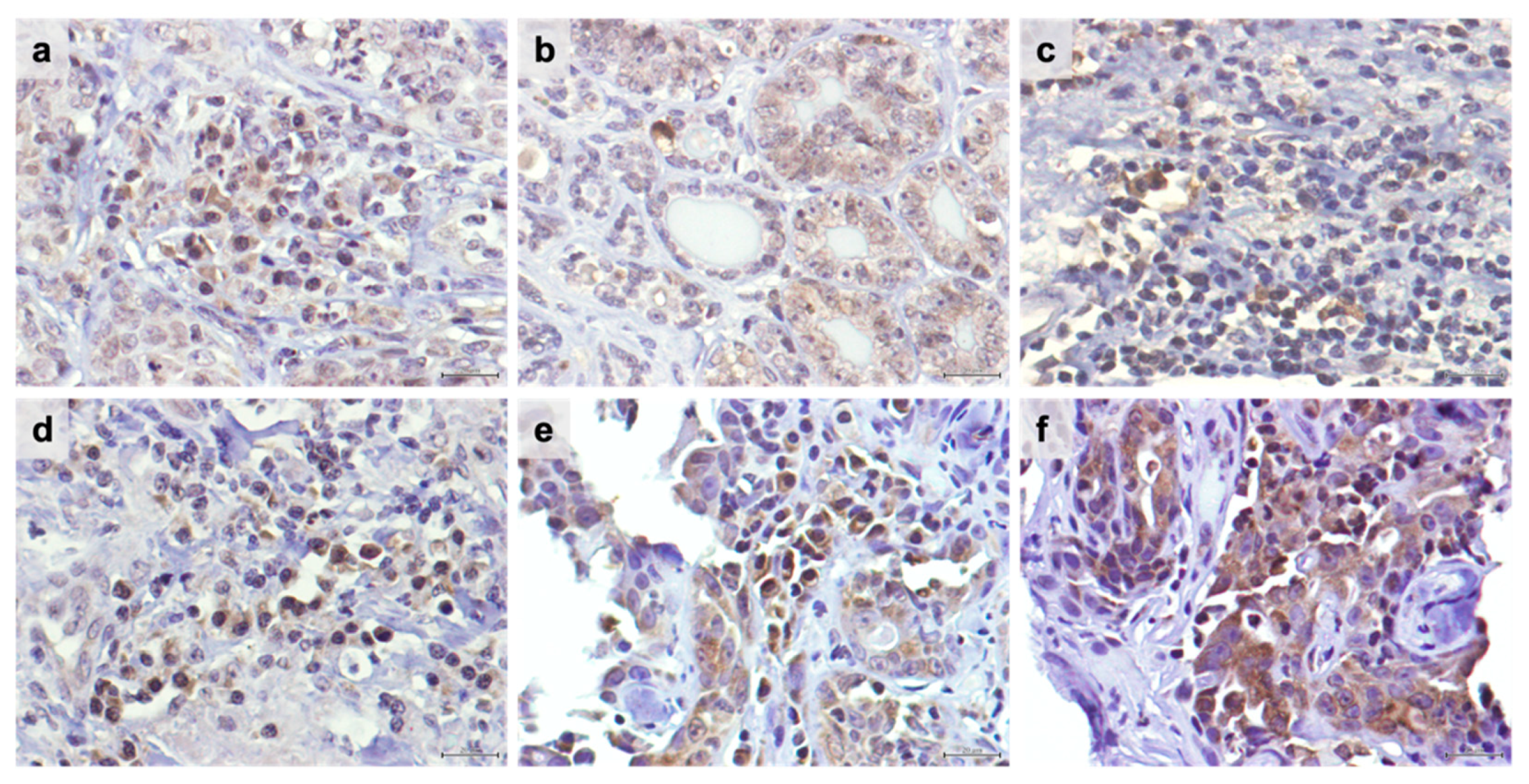
3.1. TIM-3 Is Highly Expressed in TILs and in Cancer Cells in Feline Mammary Carcinoma, with sTILs-TIM-3+ Being More Frequent Than iTILs-TIM-3+
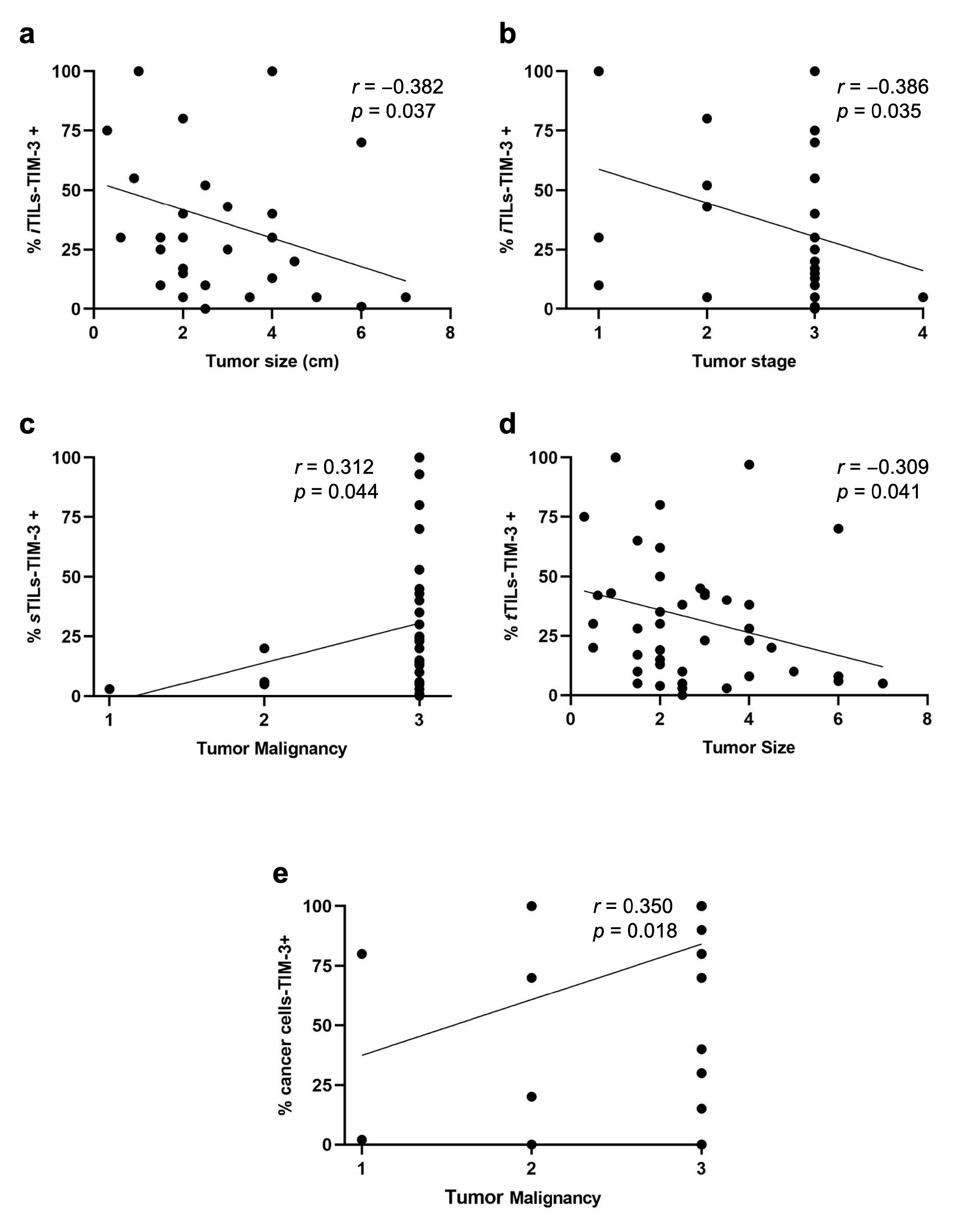
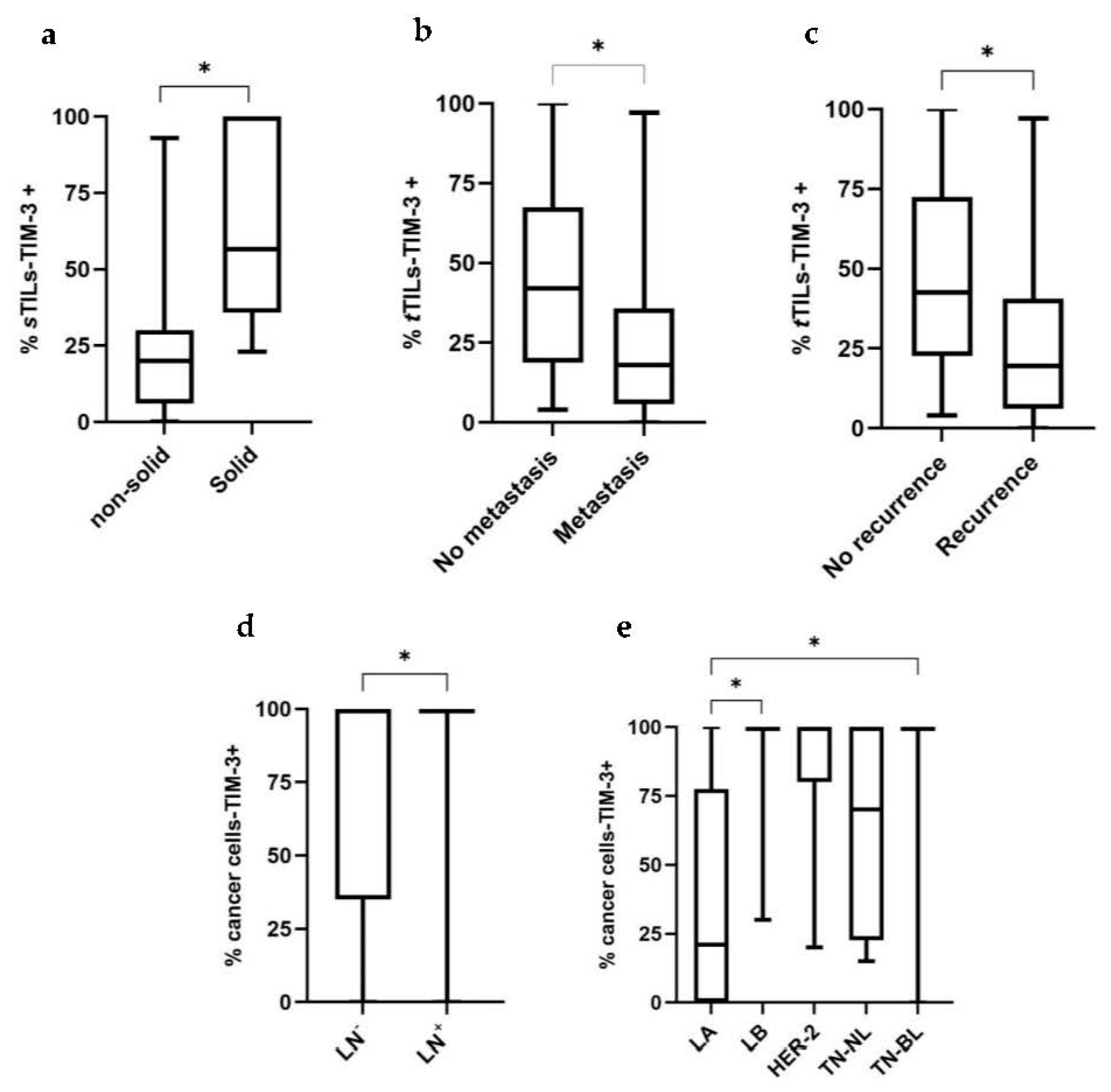
3.2. Higher TIM-3 Expression in iTILs and in tTILs Is Associated with a More Benign Tumor Behavior, whereas sTILs-TIM-3+ Are Associated with Aggressive Clinicopathological Features
3.3. Higher TIM-3 Expression in Cancer Cells Is Associated with Unfavorable Clinicopathological Features
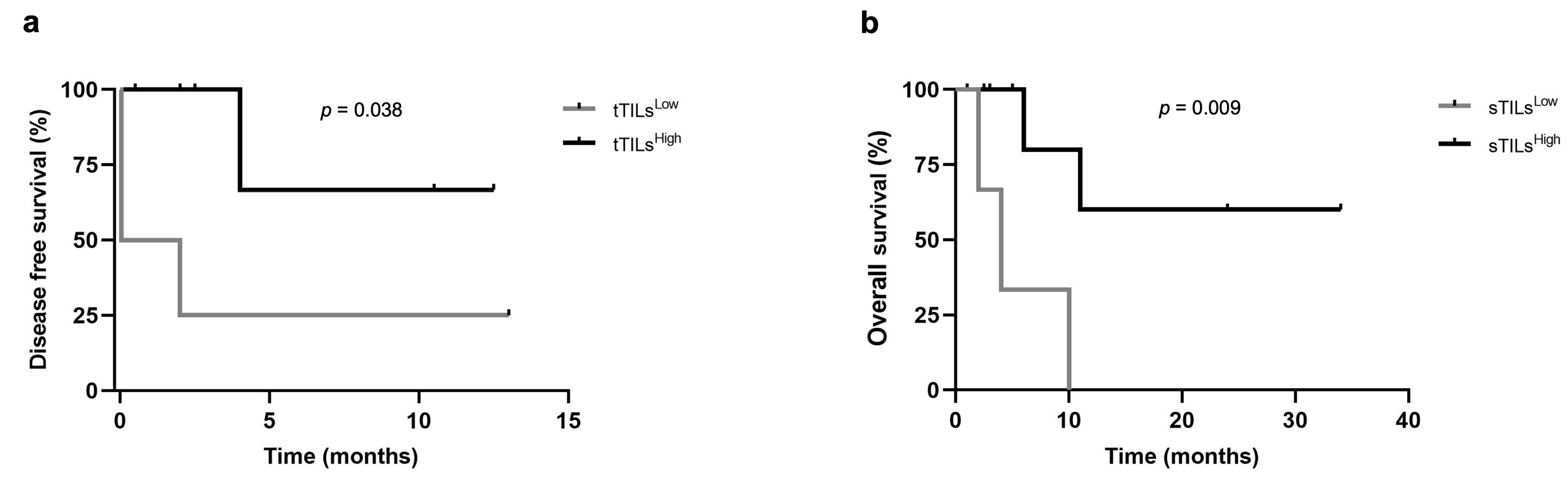
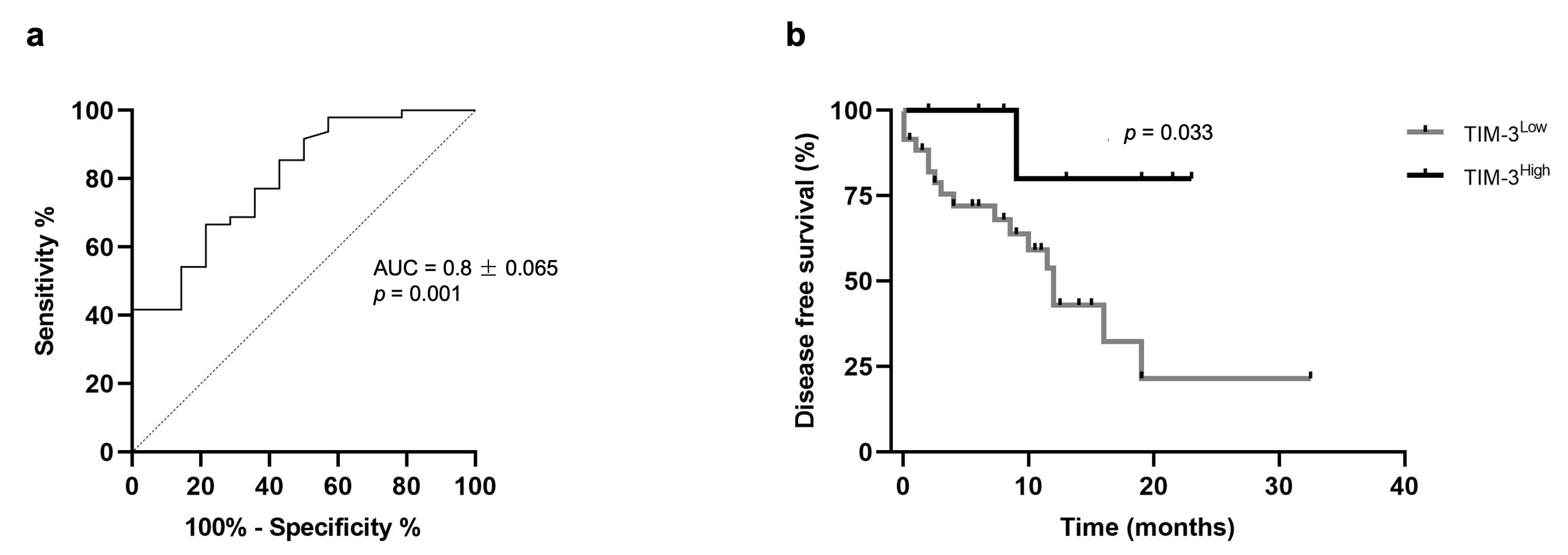
3.4. Higher Percentages of tTILs-TIM-3+ and sTILs-TIM-3+ Showed Prognostic Value in Triple-Negative FMC Subtype
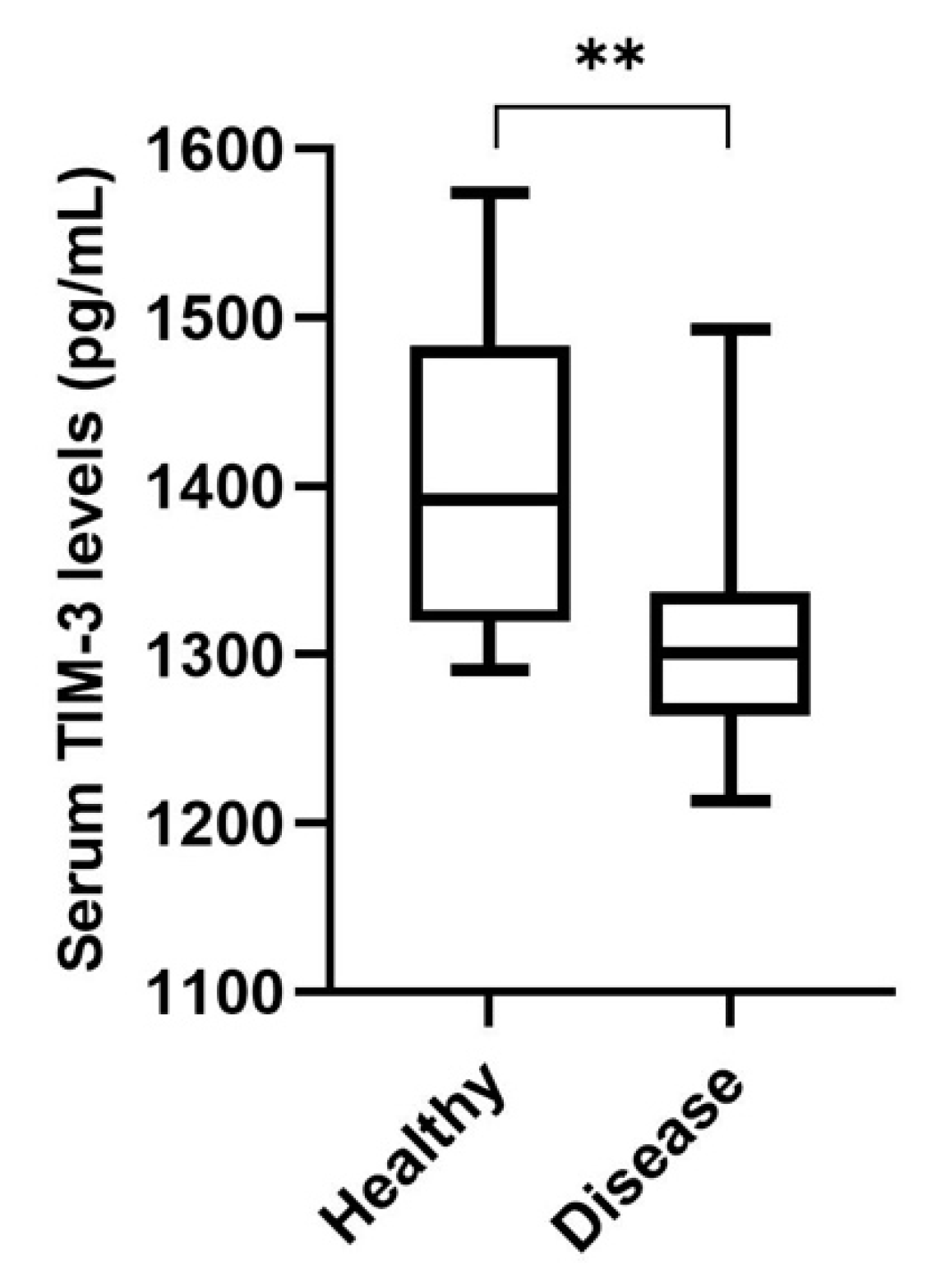
3.5. Cats with Mammary Carcinoma Showed Decreased Serum TIM-3 Levels, with Higher Levels being Associated with a Better Disease-Free Survival
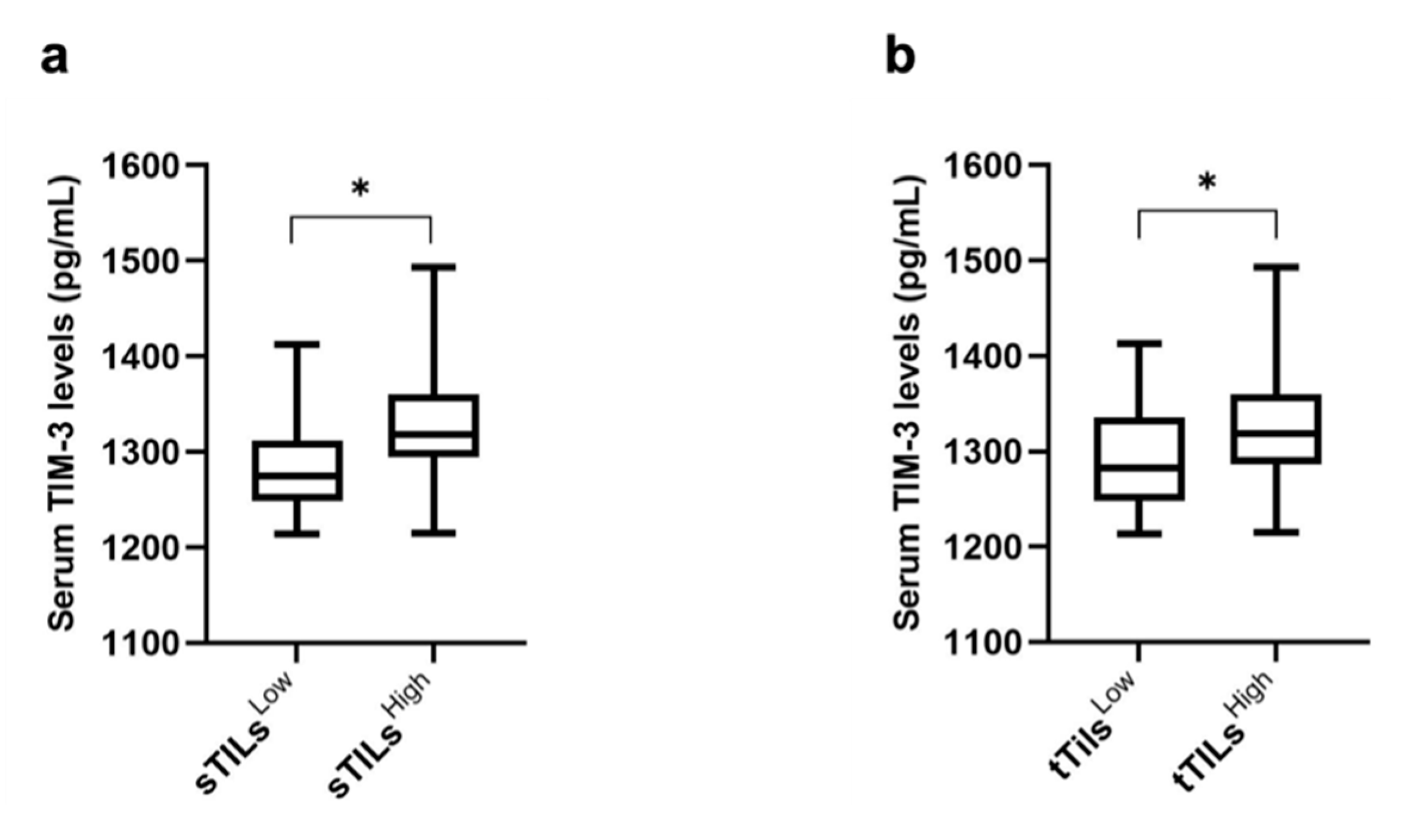
3.6. Higher Serum TIM-3 Levels Are Associated with Higher TIM-3 Expression in TILs
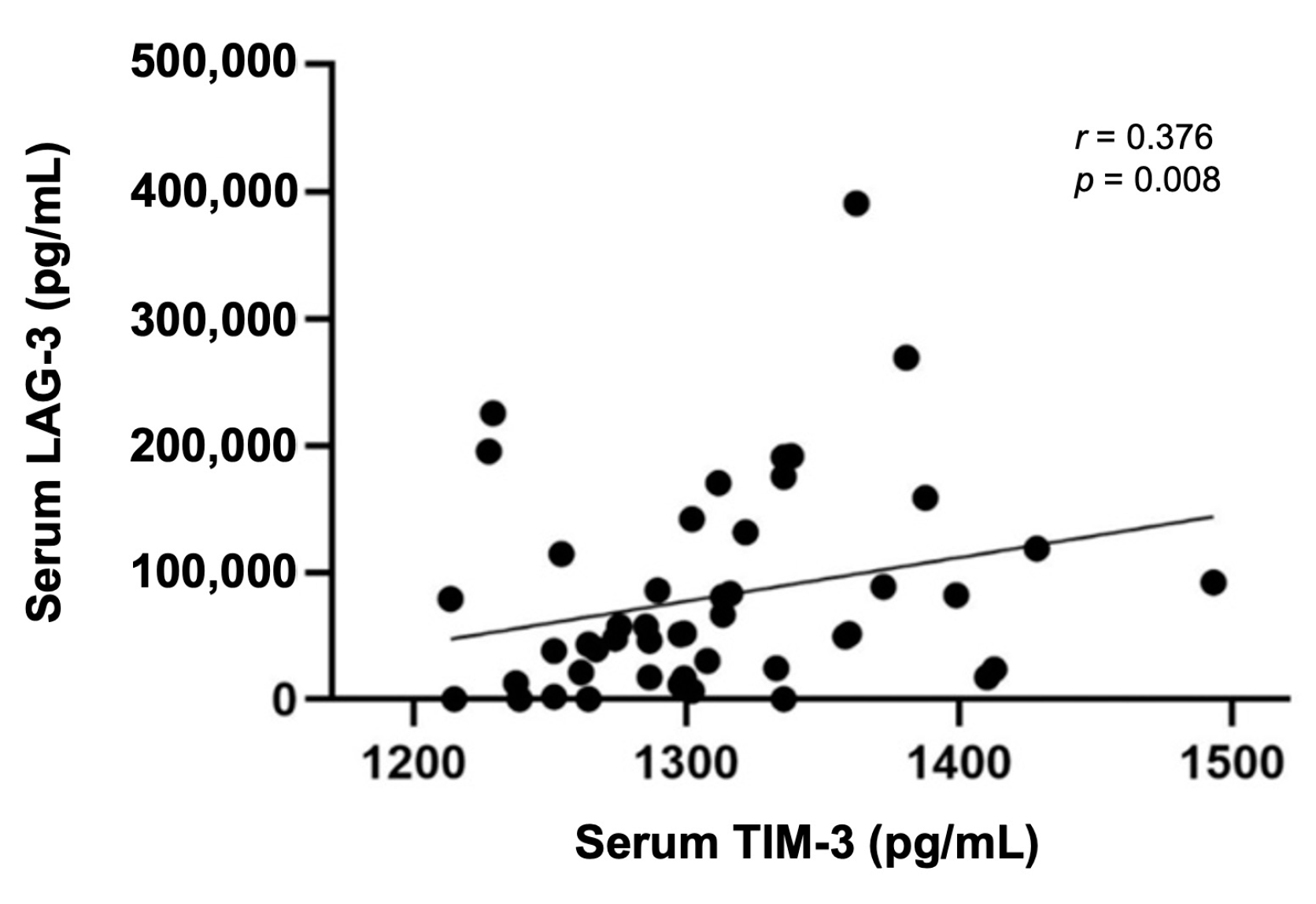
3.7. Serum TIM-3 Levels Are Positively Correlated with Serum LAG-3 Levels
3.8. Cats with Mammary Carcinoma Showed No Mutations in the Exon 3 of the TIM-3 Gene
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cannon, C.M. Cats, cancer and comparative oncology. Vet. Sci. 2015, 2, 111–126. [Google Scholar] [CrossRef]
- Goldschmidt, M.H.; Peña, L.; Zappulli, V. Tumors of the mammary gland. In Tumors in Domestic Animals, 5th ed.; Meuten, D.J., Ed.; Wiley Blackwell: Ames, IA, USA, 2017; pp. 723–765. [Google Scholar]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: Globocan estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Adega, F.; Borges, A.; Chaves, R. Cat mammary tumors: Genetic models for the human counterpart. Vet. Sci. 2016, 3, 17. [Google Scholar] [CrossRef]
- Santos, S.; Bastos, E.; Baptista, C.S.; Sá, D.; Caloustian, C.; Guedes-Pinto, H.; Gärtner, F.; Gut, I.G.; Chaves, R. Sequence variants and haplotype analysis of cat ERBB2 gene: A survey on spontaneous cat mammary neoplastic and non-neoplastic lesions. Int. J. Mol. Sci. 2012, 13, 2783–2800. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.T. Breast carcinoma: Pattern of metastasis at autopsy. J. Clin. Oncol. 1983, 23, 175–180. [Google Scholar] [CrossRef] [PubMed]
- Giménez, F.; Hecht, S.; Craig, L.E.; Legendre, A.M. Early detection, aggressive therapy. J. Feline Med. Surg. 2010, 12, 214–224. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, C.; Ferreira, F. Tumor microenvironment of human breast cancer, and feline mammary carcinoma as a potential study model. Biochim. Biophys. Acta-Rev. Cancer 2021, 1876, 188587. [Google Scholar] [CrossRef]
- Gameiro, A.; Urbano, A.C.; Ferreira, F. Emerging biomarkers and targeted therapies in feline mammary carcinoma. Vet. Sci. 2021, 8, 164. [Google Scholar] [CrossRef]
- Esfahani, K.; Roudaia, L.; Buhlaiga, N.; Del Rincon, S.V.; Papneja, N.; Miller, W.H. A review of cancer immunotherapy: From the past, to the present, to the future. Curr. Oncol. 2020, 27, 87–97. [Google Scholar] [CrossRef]
- Ribas, A.; Wolchok, J.D. Cancer immunotherapy using checkpoint blockade. Science 2018, 359, 1350–1355. [Google Scholar] [CrossRef]
- Abdou, Y.; Goudarzi, A.; Yu, J.X.; Upadhaya, S.; Vincent, B.; Carey, L.A. Immunotherapy in triple negative breast cancer: Beyond checkpoint inhibitors. NPJ Breast Cancer 2022, 8, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Haslam, A.; Prasad, V. Estimation of the percentage of US patients with cancer who are eligible for and respond to checkpoint inhibitor immunotherapy drugs. JAMA Netw. Open 2019, 2, e192535. [Google Scholar] [CrossRef] [PubMed]
- Marin-Acevedo, J.A.; Kimbrough, E.O.; Lou, Y. Next generation of immune checkpoint inhibitors and beyond. J. Hematol. Oncol. 2021, 14, 1–29. [Google Scholar] [CrossRef] [PubMed]
- Andrews, L.P.; Yano, H.; Vignali, D.A.A. Inhibitory receptors and ligands beyond PD-1, PD-L1 and CTLA-4: Breakthroughs or backups. Nat. Immunol. 2019, 20, 1425–1434. [Google Scholar] [CrossRef]
- Cong, Y.; Liu, J.; Chen, G.; Qiao, G. The emerging role of T-cell immunoglobulin mucin-3 in breast cancer: A promising target for immunotherapy. Front. Oncol. 2021, 11, 723238. [Google Scholar] [CrossRef]
- Zhao, L.; Cheng, S.; Fan, L.; Zhang, B.; Xu, S. TIM-3: An update on immunotherapy. Int. Immunopharmacol. 2021, 99, 107933. [Google Scholar] [CrossRef]
- Acharya, N.; Sabatos-Peyton, C.; Anderson, A.C. Tim-3 finds its place in the cancer immunotherapy landscape. J. Immuno. Therap. Cancer 2020, 8, e000911. [Google Scholar] [CrossRef]
- Van de Weyer, P.S.; Muehlfeit, M.; Klose, C.; Bonventre, J.V.; Walz, G.; Kuehn, E.W. A highly conserved tyrosine of Tim-3 is phosphorylated upon stimulation by its ligand galectin-9. Biochem. Biophys. Res. Commun. 2006, 351, 571–576. [Google Scholar] [CrossRef]
- Huang, Y.-H.; Zhu, C.; Kondo, Y.; Anderson, A.C.; Gandhi, A.; Russell, A.; Dougan, S.K.; Petersen, B.-S.; Melum, E.; Pertel, T.; et al. CEACAM1 regulates TIM-3-mediated tolerance and exhaustion. Nature 2015, 517, 386–390. [Google Scholar] [CrossRef]
- Cong, Y.; Cui, Y.; Zhu, S.; Cao, J.; Zou, H.A.; Martin, T.; Qiao, G.; Jiang, W.; Yu, Z. Tim-3 promotes cell aggressiveness and paclitaxel resistance through NF-κB/STAT3 signalling pathway in breast cancer cells. Chin. J. Cancer Res. 2020, 32, 564–579. [Google Scholar] [CrossRef]
- Wolf, Y.; Anderson, A.C.; Kuchroo, V.K. TIM3 comes of age as an inhibitory receptor. Nat. Rev. Immunol. 2020, 20, 173–185. [Google Scholar] [CrossRef]
- Yang, Z.-Z.; Grote, D.M.; Ziesmer, S.C.; Niki, T.; Hirashima, M.; Novak, A.J.; Witzig, T.E.; Ansell, S.M. IL-12 upregulates TIM-3 expression and induces T cell exhaustion in patients with follicular B cell non-Hodgkin lymphoma. J. Clin. Invest. 2012, 122, 1271–1282. [Google Scholar] [CrossRef]
- Granier, C.; Dariane, C.; Combe, P.; Verkarre, V.; Urien, S.; Badoual, C.; Roussel, H.; Mandavit, M.; Ravel, P.; Sibony, M.; et al. Tim-3 Expression on Tumor-Infiltrating PD-1+CD8+ T Cells Correlates with Poor Clinical Outcome in Renal Cell Carcinoma. Cancer Res. 2016, 77, 1075–1082. [Google Scholar] [CrossRef] [PubMed]
- Cao, B.; Zhu, L.; Zhu, S.; Li, D.; Zhang, C.; Xu, C.; Zhang, S. Genetic variations and haplotypes in TIM-3 gene and the risk of gastric cancer. Cancer Immunol. Immunother. 2010, 59, 1851–1857. [Google Scholar] [CrossRef]
- Cao, Y.; Zhou, X.; Huang, X.; Li, Q.; Gao, L.; Jiang, L.; Huang, M.; Zhou, J. Correction: Tim-3 expression in cervical cancer promotes tumor metastasis. PLoS ONE 2016, 11, e0152830. [Google Scholar] [CrossRef] [PubMed]
- Piao, Y.-R.; Piao, L.-Z.; Zhu, L.-H.; Jin, Z.-H.; Dong, X.-Z. Prognostic value of T cell immunoglobulin mucin-3 in prostate cancer. Asian Pac. J. Cancer Prev. 2013, 14, 3897–3901. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Liu, Y.; Chen, Z. Tim-3 expression and its role in hepatocellular carcinoma. J. Hematol. Oncol. 2018, 11, 126. [Google Scholar] [CrossRef]
- Cheng, S.; Han, F.; Xu, Y.; Qu, T.; Ju, Y. Expression of Tim-3 in breast cancer tissue promotes tumor progression. Int. J. Clin. Exp. Pathol. 2018, 11, 1157–1166. [Google Scholar]
- Zang, K.; Hui, L.; Wang, M.; Huang, Y.; Zhu, X.; Yao, B. TIM-3 as a prognostic marker and a potential immunotherapy target in human malignant tumors: A meta-analysis and bioinformatics validation. Front Oncol. 2021, 11, 579351. [Google Scholar] [CrossRef]
- Gameiro, A.; Nascimento, C.; Correia, J.; Ferreira, F. HER2-targeted immunotherapy and combined protocols showed promising antiproliferative effects in feline mammary carcinoma cell-based models. Cancers 2021, 13, 2007. [Google Scholar] [CrossRef]
- Wang, Z.; Liu, X.; Wang, X.; Chong, T.; Lin, S.; Wang, M.; Ma, X.; Liu, K.; Xu, P.; Feng, Y.; et al. Polymorphisms in TIM-3 and breast cancer susceptibility in Chinese women: A case-control study. Oncotarget 2016, 7, 43703–43712. [Google Scholar] [CrossRef] [PubMed]
- Urbano, A.C.; Nascimento, C.; Soares, M.; Correia, J.; Ferreira, F. Clinical relevance of the serum CTLA-4 in cats with mammary carcinoma. Sci. Rep. 2020, 10, 3822. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, C.; Urbano, A.C.; Gameiro, A.; Ferreira, J.; Correia, J.; Ferreira, F. Serum PD-1/PD-L1 levels, tumor expression and PD-L1 somatic mutations in HER2-positive and triple negative normal-Like feline mammary carcinoma subtypes. Cancers 2020, 12, 1386. [Google Scholar] [CrossRef]
- Nascimento, C.; Gameiro, A.; Ferreira, J.; Correia, J.; Ferreira, F. Diagnostic value of VEGF-A, VEGFR-1 and VEGFR-2 in feline mammary carcinoma. Cancers 2021, 13, 117. [Google Scholar] [CrossRef]
- Cheng, S.; Ju, Y.; Han, F.; Wang, Y.; Xu, Y.; Qu, T.; Lu, Z. 2017. T cell immunoglobulin- and mucin-domain-containing molecule 3 gene polymorphisms and susceptibility to invasive breast cancer. Ann. Clin. Lab. Sci. 2017, 47, 668–675. [Google Scholar]
- Burugu, S.; Gao, D.; Leung, S.; Chia, S.K.; Nielsen, T.O. TIM-3 expression in breast cancer. OncoImmunology 2018, 7, e1502128. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.-W.; Ryu, H.S.; Jin, M.-S.; Lee, K.-H.; Suh, K.J.; Youk, J.; Kim, J.Y.; Min, A.; Lee, H.-B.; Moon, H.-G.; et al. Immune recurrence score using 7 immunoregulatory protein expressions can predict recurrence in stage I–III breast cancer patients. Br. J. Cancer 2019, 121, 230–236. [Google Scholar] [CrossRef]
- Zhang, H.; Xiang, R.; Wu, B.; Li, J.; Luo, G. T-cell immunoglobulin mucin-3 expression in invasive ductal breast carcinoma: Clinicopathological correlations and association with tumor infiltration by cytotoxic lymphocytes. Mol. Clin. Oncol. 2017, 7, 557–563. [Google Scholar] [CrossRef]
- Koyama, S.; Akbay, E.A.; Li, Y.Y.; Herter-Sprie, G.S.; Buczkowski, K.A.; Richards, W.G.; Gandhi, L.; Redig, A.J.; Rodig, S.J.; Asahina, H.; et al. Adaptive resistance to therapeutic PD-1 blockade is associated with upregulation of alternative immune checkpoints. Nat. Commun. 2016, 7, 10501. [Google Scholar] [CrossRef]
- Yasinska, I.M.; Sakhnevych, S.S.; Pavlova, L.; Teo Hansen Selnø, A.; Teuscher Abeleira, A.M.; Benlaouer, O.; Gonçalves Silva, I.; Mosimann, M.; Varani, L.; Bardelli, M.; et al. The Tim-3-galectin-9 pathway and its regulatory mechanisms in human breast cancer. Front. Immunol. 2019, 10, 1594. [Google Scholar] [CrossRef]
- Rapoport, B.L.; Steel, H.; Smit, T.; Heyman, L.; Theron, A.; Hlatswayo, N.N.; Kwofie, L.; Jooste, L.; Benn, C.A.; Nayler, S.; et al. 37P Dysregulation of soluble immune checkpoint proteins in newly diagnosed early breast cancer patients. Ann. Oncol. 2020, 31, S1228. [Google Scholar] [CrossRef]
- Byun, K.D.; Hwang, H.J.; Park, K.J.; Kim, M.C.; Cho, S.H.; Ju, M.H.; Lee, J.H.; Jeong, J.S. T-cell immunoglobulin mucin 3 expression on tumor infiltrating lymphocytes as a positive prognosticator in triple-negative breast cancer. J. Breast Cancer 2018, 21, 406–414. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, K.; Ishida, M.; Yanai, H.; Tsuta, K.; Sekimoto, M.; Sugie, T. Prognostic significance of the expression levels of T-cell immunoglobulin mucin-3 and its ligand galectin-9 for relapse-free survival in triple-negative breast cancer. Oncol. Lett. 2022, 23, 197. [Google Scholar] [CrossRef] [PubMed]
- Rapoport, B.L.; Steel, H.C.; Hlatshwayo, N.; Theron, A.J.; Meyer, P.W.A.; Nayler, S.; Benn, C.-A.; Smit, T.; Kwofie, L.L.I.; Heyman, L.; et al. Systemic immune dysregulation in early breast cancer is associated with decreased plasma levels of both soluble co-inhibitory and co-stimulatory immune checkpoint molecules. Front. Immunol. 2022, 13, 823842. [Google Scholar] [CrossRef]
- Tawbi, H.A.; Schadendorf, D.; Lipson, E.J.; Ascierto, P.A.; Matamala, L.; Castillo, G.E.; Rutkowski, P.; Gogas, H.J.; Lao, C.D.; De Menezes, J.J.; et al. Relatlimab and Nivolumab versus Nivolumab in Untreated Advanced Melanoma. N. Engl. J. Med. 2022, 386, 24–34. [Google Scholar] [CrossRef]
- Fang, H.; Yuan, C.; Gu, X.; Chen, Q.; Huang, D.; Li, H.; Sun, M. Association between TIM-3 polymorphisms and cancer risk: A meta-analysis. Ann. Transl. Med. 2019, 7, 550. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clinicopathological Features | Number of Animals (%) | Clinicopathological Features | Number of Animals (%) |
|---|---|---|---|
| Age | LVI | ||
| <8 years old | 4 (8.3%) | No | 41 (85.4%) |
| 8–12 years old | 24 (50%) | Yes | 7 (14.6%) |
| >12 years old | 31.3 (41.7%) | ||
| Breed | Tumor ulceration | ||
| Undifferentiated | 35 (72.9%) | No | 41 (85.4%) |
| Pure | 13 (27.1%) | Yes | 7 (14.6%) |
| Contraceptive administration: 6 unknown | TNM classification | ||
| No | 20 (41.7%) | I | 11 (22.9%) |
| Yes | 22 (45.8%) | II | 6 (12.5%) |
| Spayed: 1 unknown | III | 27 (56.3%) | |
| No | 23 (47.9%) | IV | 4 (8.3%) |
| Yes | 24 (50.0%) | ||
| Tumor burden | Lymph node status: 3 unknown | ||
| Single tumor | 19 (39.6%) | Negative | 29 (60.4%) |
| Multiple tumors | 29 (60.4%) | Positive | 16 (33.3%) |
| Tumor size | ER status | ||
| <2 cm | 15 (31.3%) | Negative | 36 (75%) |
| ≥2 cm | 33 (68.8%) | Positive | 12 (25%) |
| HP classification | PR status | ||
| Tubular carcinoma | 3 (6.3%) | Negative | 20 (41.7) |
| Tubulopapillary carcinoma | 6 (12.5%) | Positive | 28 (58.3) |
| Papillary-cystic carcinoma | 6 (12.5%) | HER-2 status | |
| Cribriform carcinoma | 19 (39.6%) | Negative | 38 (79.2%) |
| Solid carcinoma | 11 (22.9%) | Positive | 10 (20.8%) |
| Mucinous carcinoma | 3 (6.3%) | ||
| Malignancy Grade | Ki-67 index | ||
| 1 | 2 (4.2%) | Low | 14 (29.2%) |
| 2 | 6 (12.5%) | High | 34 (70.8%) |
| 3 | 40 (83.3%) | ||
| Tumor Necrosis | |||
| No | 11 (22.9%) | ||
| Yes | 37 (77.1%) | ||
| Exon | Forward (5′–3′) | Reverse (5′–3′) |
|---|---|---|
| 3 | ACTAGACTGTGATGACGATGGC | AGGAACATTCACACCTCCACTC |
| Occurrence of TIM-3+ TILs | Tumors with iTILs-TIM-3+ (%) | |||||
|---|---|---|---|---|---|---|
| LA | LB | HER-2+ | TN-NL | TN-BL | Total | |
| Present | 83.3% (n = 5) | 70.6% (n = 12) | 66.7% (n = 6) | 60.0% (n = 3) | 37.5% (n = 3) | 64.4% (n = 29) |
| Absent | 16.7% (n = 1) | 29.4% (n = 5) | 33.3% (n = 3) | 40.0% (n = 2) | 62.5% (n = 5) | 35.6% (n = 16) |
| Total | 100% (n = 6) | 100% (n = 17) | 100% (n = 9) | 100% (n = 5) | 100% (n = 8) | 100% (n = 45) |
| Occurrence of TIM-3+ TILs | Tumors with sTILs-TIM-3+ (%) | |||||
|---|---|---|---|---|---|---|
| LA | LB | HER-2+ | TN-NL | TN-BL | Total | |
| Present | 100% (n = 6) | 94.1% (n = 16) | 77.8% (n = 7) | 100% (n = 5) | 100% (n = 8) | 93.3% (n = 42) |
| Absent | 0% (n = 0) | 5.9% (n = 1) | 22.2% (n = 2) | 0% (n = 0) | 0% (n = 0) | 6.7% (n = 3) |
| Total | 100% (n = 6) | 100% (n = 17) | 100% (n = 9) | 100% (n = 5) | 100% (n = 8) | 100% (n = 45) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valente, S.; Nascimento, C.; Gameiro, A.; Ferreira, J.; Correia, J.; Ferreira, F. TIM-3 Is a Potential Immune Checkpoint Target in Cats with Mammary Carcinoma. Cancers 2023, 15, 384. https://doi.org/10.3390/cancers15020384
Valente S, Nascimento C, Gameiro A, Ferreira J, Correia J, Ferreira F. TIM-3 Is a Potential Immune Checkpoint Target in Cats with Mammary Carcinoma. Cancers. 2023; 15(2):384. https://doi.org/10.3390/cancers15020384
Chicago/Turabian StyleValente, Sofia, Catarina Nascimento, Andreia Gameiro, João Ferreira, Jorge Correia, and Fernando Ferreira. 2023. "TIM-3 Is a Potential Immune Checkpoint Target in Cats with Mammary Carcinoma" Cancers 15, no. 2: 384. https://doi.org/10.3390/cancers15020384
APA StyleValente, S., Nascimento, C., Gameiro, A., Ferreira, J., Correia, J., & Ferreira, F. (2023). TIM-3 Is a Potential Immune Checkpoint Target in Cats with Mammary Carcinoma. Cancers, 15(2), 384. https://doi.org/10.3390/cancers15020384