
The 2EF Antibody Targets a Unique N-Terminal Epitope of Trop-2 and Enhances the In Vivo Activity of the Cancer-Selective 2G10 Antibody

 , , ,
, , ,  , ,
, ,  , , and
, , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Generation of the Anti-Trop-2 Murine mAb
2.2. DNA Transfection
2.3. ELISA
2.4. Flow Cytometry
2.5. Immunofluorescence and Confocal Microscopy
2.6. Cloning of the VH and VL Regions of Mouse 2EF
- 2EF VH
- 5′ primer 2EF-H5: 5′-TACACCTTCACTAACTACTGG-3′
- 3′ primers 2EF-H3: 5′-CCCAGTTCCTCTGCACAG-3′
- MCG2b: 5′-GCCAGTGGATAGACTGATGG-3’
- 2EF VL
- 5′ primer 2EF-L5: 5′-AGCCAAAGTGTCAGTACATC-3′
- 3′ primers JNT319: 5′-CTCCCTCTAACACTCATTCCTGTTGAAGC-3′
- 2EF-L3: 5′-GAATCTCCCGACTGTGCTG-3′
2.7. Construction of the Chimeric 2EF IgG1/k Ab
2.8. Strategy for 2EF Humanization
2.9. Generation of NS-0 and YB2/0 Producer Cell Lines
2.10. Expression and Purification of Hu2EF-7
2.11. Ab-dependent Cellular Cytotoxicity (ADCC) Assay
2.12. Experimental Tumors
2.13. Study Approval
2.14. Statistical Analysis
3. Results
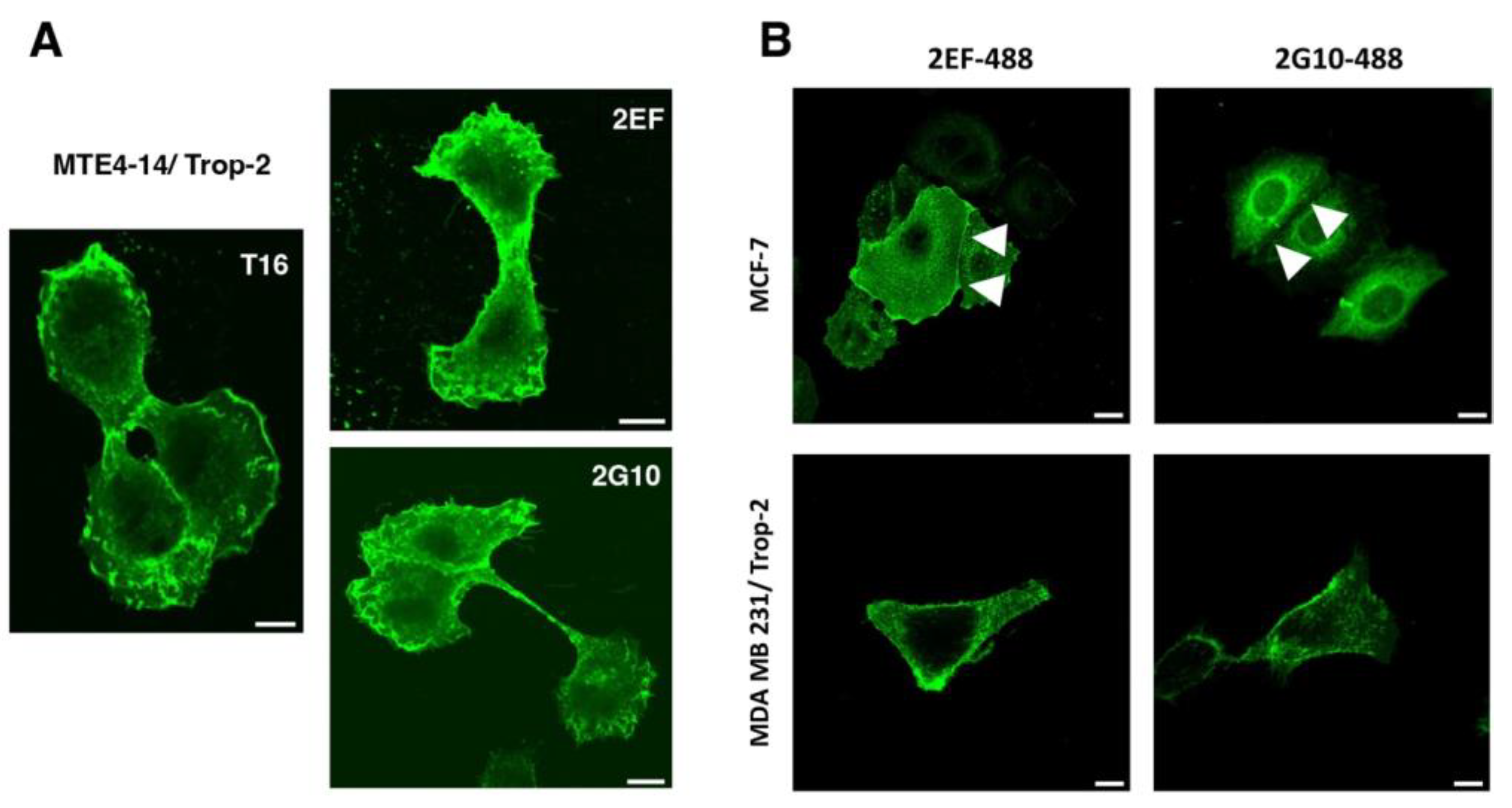
3.1. Generation of mAbs Directed against Accessible Trop-2 Sites in Tightly Packed Tumor Cells
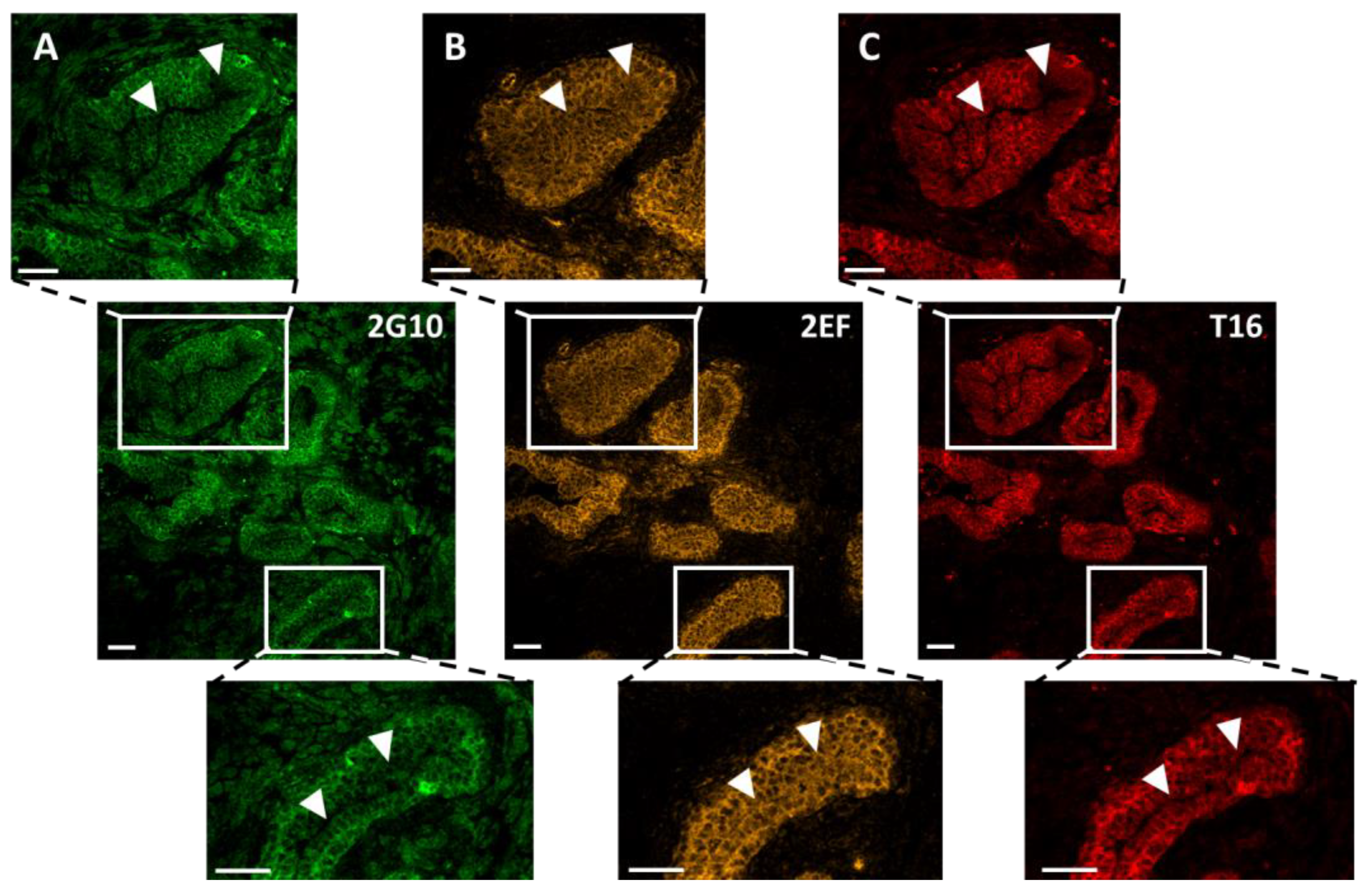
3.2. Effective Binding of 2EF to Tumor Cells at Deeply Seated Cancer Sites
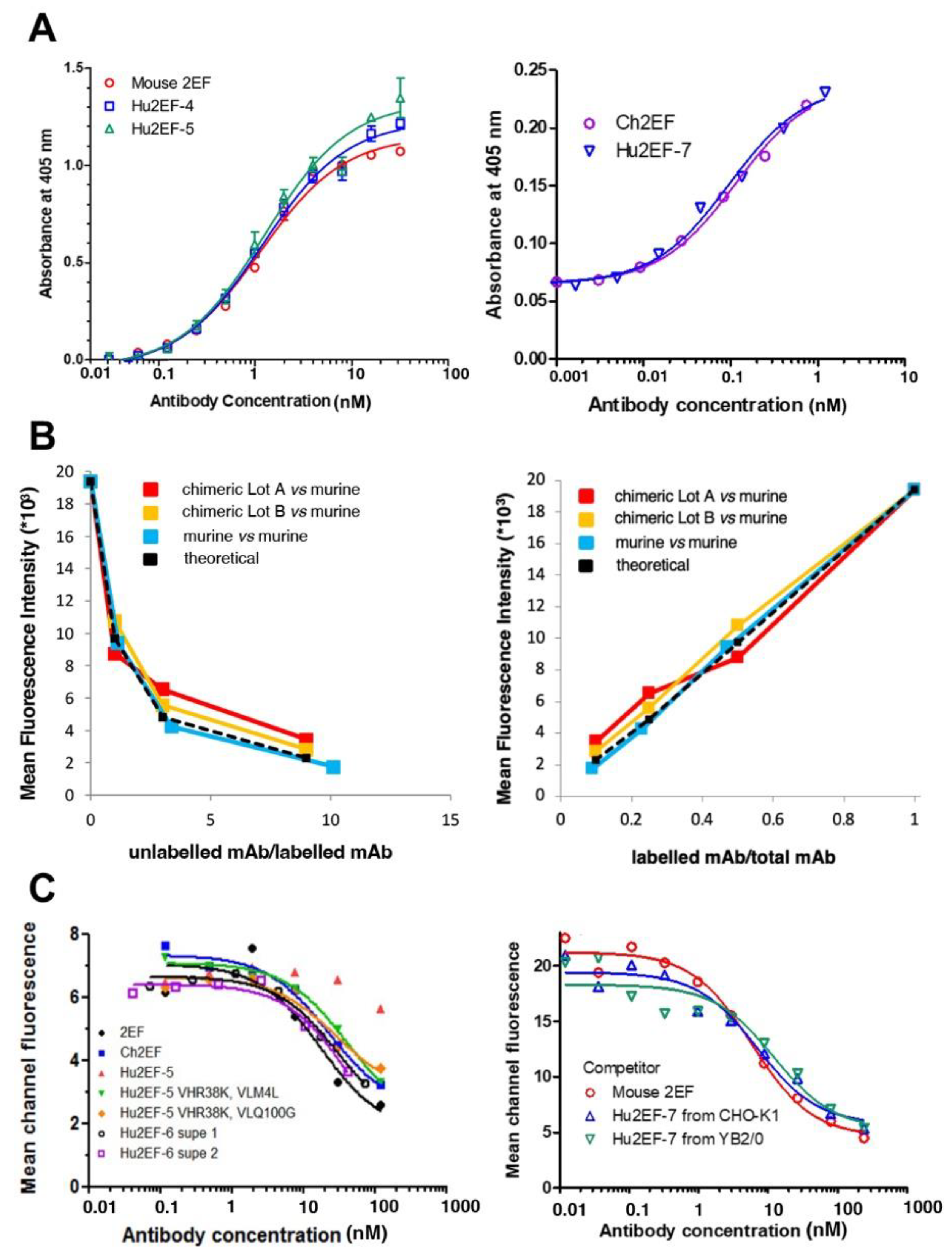
3.3. Engineering of Chimeric and Humanized 2EF
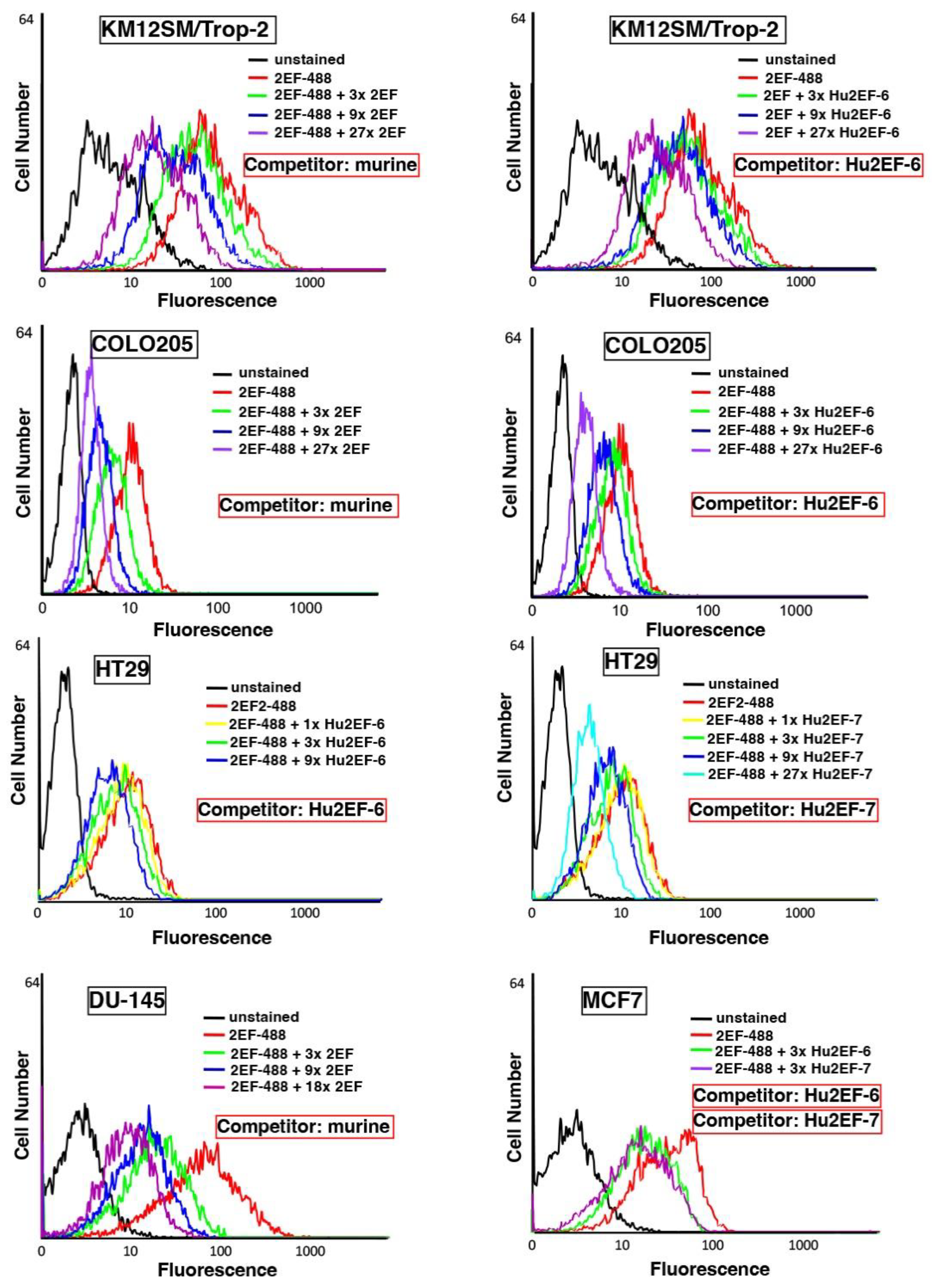
3.4. Hu2EF-7 Binding to Trop-2
3.5. Ab-Dependent Cell-Mediated Cytotoxicity by Hu2EF
3.6. Inhibition of Cancer Cell Growth by 2EF
3.7. Xenograft Growth Inhibition by 2EF
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Fornaro, M.; Dell′Arciprete, R.; Stella, M.; Bucci, C.; Nutini, M.; Capri, M.G.; Alberti, S. Cloning of the gene encoding TROP-2, a cell-surface glycoprotein expressed by human carcinomas. Int. J. Cancer 1995, 62, 610–618. [Google Scholar] [CrossRef]
- Linnenbach, A.J.; Seng, B.A.; Wu, S.; Robbins, S.; Scollon, M.; Pyrc, J.J.; Druck, T.; Huebner, K. Retroposition in a family of carcinoma-associated antigen genes. Mol. Cell Biol. 1993, 13, 1507–1515. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, G.; Crescenzi, C.; Morizio, E.; Palka, G.; Guerra, E.; Alberti, S. Assignment of TACSTD1 (alias TROP1, M4S1) to human chromosome 2p21 and refinement of mapping of TACSTD2 (alias TROP2, M1S1) to human chromosome 1p32 by in situ hybridization. Cytogenet. Cell Genet. 2001, 92, 164–165. [Google Scholar] [CrossRef]
- Zanna, P.; Trerotola, M.; Vacca, G.; Bonasera, V.; Palombo, B.; Guerra, E.; Rossi, C.; Lattanzio, R.; Piantelli, M.; Alberti, S. Trop-1 Are Conserved Growth Stimulatory Molecules That Mark Early Stages of Tumor Progression. Cancer 2007, 110, 452–464. [Google Scholar] [CrossRef]
- Ripani, E.; Sacchetti, A.; Corda, D.; Alberti, S. The human Trop-2 is a tumor-associated calcium signal transducer. Int. J. Cancer 1998, 76, 671–676. [Google Scholar] [CrossRef]
- Guerra, E.; Relli, V.; Ceci, M.; Tripaldi, R.; Simeone, P.; Aloisi, A.L.; Pantalone, L.; La Sorda, R.; Lattanzio, R.; Sacchetti, A.; et al. Trop-2, Na+/K+ ATPase, CD9, PKCα, cofilin assemble a membrane signaling super-complex that drives colorectal cancer growth and invasion. Oncogene 2022, 41, 1795–1808. [Google Scholar] [CrossRef] [PubMed]
- Guerra, E.; Trerotola, M.; Tripaldi, R.; Aloisi, A.L.; Simeone, P.; Sacchetti, A.; Relli, V.; D’Amore, A.; La Sorda, R.; Lattanzio, R.; et al. Trop-2 induces tumor growth through Akt and determines sensitivity to Akt inhibitors. Clin. Cancer Res. 2016, 22, 4197–4205. [Google Scholar] [CrossRef] [PubMed]
- Trerotola, M.; Cantanelli, P.; Guerra, E.; Tripaldi, R.; Aloisi, A.L.; Bonasera, V.; Lattanzio, R.; de Lange, R.; Weidle, U.H.; Piantelli, M.; et al. Up-regulation of Trop-2 quantitatively stimulates human cancer growth. Oncogene 2013, 32, 222–233. [Google Scholar] [CrossRef]
- Trerotola, M.; Guerra, E.; Ali, Z.; Aloisi, A.L.; Ceci, M.; Simeone, P.; Acciarito, A.; Zanna, P.; Vacca, G.; D′Amore, A.; et al. Trop-2 cleavage by ADAM10 is an activator switch for cancer growth and metastasis. Neoplasia 2021, 23, 415–428. [Google Scholar] [CrossRef]
- Guerra, E.; Trerotola, M.; Relli, V.; Lattanzio, R.; Tripaldi, R.; Vacca, G.; Ceci, M.; Boujnah, K.; Garbo, V.; Moschella, A.; et al. Trop-2 induces ADAM10-mediated cleavage of E-cadherin and drives EMT-less metastasis in colon cancer. Neoplasia 2021, 23, 898–911. [Google Scholar] [CrossRef]
- Relli, V.; Trerotola, M.; Guerra, E.; Alberti, S. Distinct lung cancer subtypes associate to distinct drivers of tumor progression. Oncotarget 2018, 9, 35528–35540. [Google Scholar] [CrossRef]
- Trerotola, M.; Jernigan, D.; Liu, Q.; Siddiqui, J.; Fatatis, A.; Languino, L. Trop-2 promotes prostate cancer metastasis by modulating β1 integrin functions. Cancer Res. 2013, 73, 3155–3167. [Google Scholar] [CrossRef] [PubMed]
- Bardia, A.; Mayer, I.A.; Vahdat, L.T.; Tolaney, S.M.; Isakoff, S.J.; Diamond, J.R.; O′Shaughnessy, J.; Moroose, R.L.; Santin, A.D.; Abramson, V.G.; et al. Sacituzumab Govitecan-hziy in Refractory Metastatic Triple-Negative Breast Cancer. N. Engl. J. Med. 2019, 380, 741–751. [Google Scholar] [CrossRef]
- Bardia, A.; Hurvitz, S.A.; Tolaney, S.M.; Loirat, D.; Punie, K.; Oliveira, M.; Brufsky, A.; Sardesai, S.D.; Kalinsky, K.; Zelnak, A.B.; et al. Sacituzumab Govitecan in Metastatic Triple-Negative Breast Cancer. N. Engl. J. Med. 2021, 384, 1529–1541. [Google Scholar] [CrossRef]
- Tagawa, S.T. TROPHY-U-01: A phase II open-label study of sacituzumab govitecan in patients with metastatic urothelial carcinoma progressing after platinum-based chemotherapy and checkpoint inhibitors. J. Clin. Oncol. 2021, 26, 66–675. [Google Scholar] [CrossRef] [PubMed]
- Trerotola, M.; Guerra, E.; Alberti, S. Letter to the editor: Efficacy and safety of anti-Trop antibodies. Biochim. Biophys. Acta 2010, 1805, 119–120. [Google Scholar]
- Okajima, D.; Yasuda, S.; Maejima, T.; Karibe, T.; Sakurai, K.; Aida, T.; Toki, T.; Yamaguchi, J.; Kitamura, M.; Kamei, R.; et al. Datopotamab Deruxtecan, a Novel TROP2-directed Antibody–drug Conjugate, Demonstrates Potent Antitumor Activity by Efficient Drug Delivery to Tumor Cells. Mol. Cancer Ther. 2021, 20, 2329. [Google Scholar] [CrossRef]
- King, G.T.; Eaton, K.D.; Beagle, B.R.; Zopf, C.J.; Wong, G.Y.; Krupka, H.I.; Hua, S.Y.; Messersmith, W.A.; El-Khoueiry, A.B. A phase 1, dose-escalation study of PF-06664178, an anti-Trop-2/Aur0101 antibody-drug conjugate in patients with advanced or metastatic solid tumors. Investig. New Drugs 2018, 36, 836–847. [Google Scholar] [CrossRef]
- Guerra, E.; Alberti, S. The anti-Trop-2 antibody-drug conjugate Sacituzumab Govitecan—Effectiveness, pitfalls and promises. Ann. Transl. Med. 2022, 10, 501–505. [Google Scholar] [CrossRef] [PubMed]
- Guerra, E.; Trerotola, M.; Relli, V.; Lattanzio, R.; Tripaldi, R.; Ceci, M.; Boujnah, K.; Pantalone, L.; Sacchetti, A.; Havas, K.M.; et al. 3D-informed targeting of the Trop-2 signal-activation site drives selective cancer vulnerability. Mol. Cancer Ther. 2023, 22, 790–804. [Google Scholar] [CrossRef]
- Alberti, S.; Trerotola, M.; Guerra, E. The Hu2G10 mAb targets the cleaved-activated form of Trop-2 and exploits vulnerability of multiple human cancers. J. Clin. Oncol. 2022, 40, e14548. [Google Scholar] [CrossRef]
- Alberti, S.; Trerotola, M.; Guerra, E. Abstract 340: The Hu2G10 tumor-selective anti-Trop-2 monoclonal antibody targets the cleaved-activated Trop-2 and shows therapeutic efficacy against multiple human cancers. Cancer Res. 2022, 82, 340. [Google Scholar] [CrossRef]
- Kim, H.; Guerra, E.; Baek, E.; Jeong, Y.; You, H.; Yu, B.; Jang, T.; Saverio, A.; Chung, C.-W.; Park, C. Abstract 328: LCB84, a TROP2-targeted ADC, for treatment of solid tumors that express TROP-2 using the hu2G10 tumor-selective anti-TROP2 monoclonal antibody, a proprietary site-directed conjugation technology and plasma-stable tumor-selective linker chemistry. Cancer Res. 2022, 82, 328. [Google Scholar] [CrossRef]
- Pavšič, M. Trop2 Forms a Stable Dimer with Significant Structural Differences within the Membrane-Distal Region as Compared to EpCAM. Int. J. Mol. Sci. 2021, 22, 640. [Google Scholar] [CrossRef]
- Sun, M.; Zhang, H.; Jiang, M.; Chai, Y.; Qi, J.; Gao, G.F.; Tan, S. Structural insights into the cis and trans assembly of human trophoblast cell surface antigen 2. iScience 2021, 24, 103190. [Google Scholar] [CrossRef]
- Balzar, M.; Briaire-de Bruijn, I.H.; Rees-Bakker, H.A.M.; Prins, F.A.; Helfrich, W.; Leij, L.D.; Riethmuller, G.; Alberti, S.; Warnaar, S.O.; Fleuren, G.J.; et al. Epidermal growth factor-like repeats mediate lateral and reciprocal interactions of Ep-CAM molecules in homophilic adhesions. Mol. Cell. Biol. 2001, 21, 2570–2580. [Google Scholar] [CrossRef] [PubMed]
- Fradet, Y.; Cordon-Cardo, C.; Thomson, T.; Daly, M.E.; Whitmore, W.F., Jr.; Lloyd, K.O.; Melamed, M.R.; Old, L.G. Cell-surface antigens of human bladder cancer defined by mouse monoclonal antibodies. Proc. Natl. Acad. Sci. USA 1984, 81, 224–228. [Google Scholar] [CrossRef]
- Alberti, S.; Miotti, S.; Stella, M.; Klein, C.E.; Fornaro, M.; Ménard, S.; Colnaghi, M.I. Biochemical characterization of Trop-2, a cell surface molecule expressed by human carcinomas: Formal proof that the monoclonal antibodies T16 and MOv-16 recognize Trop-2. Hybridoma 1992, 11, 539–545. [Google Scholar] [CrossRef] [PubMed]
- Alberti, S.; Nutini, M.; Herzenberg, L.A. DNA methylation prevents the amplification of TROP1, a tumor associated cell surface antigen gene. Proc. Natl. Acad. Sci. USA 1994, 91, 5833–5837. [Google Scholar] [CrossRef]
- Maguire, T.M.; Shering, S.G.; McDermott, E.W.; O’Higgins, N.; Fennelly, J.J.; Crown, J.; Duffy, M.J. Assay of E-cadherin by ELISA in human breast cancers. Eur. J. Cancer 1997, 33, 404–408. [Google Scholar] [CrossRef]
- Dell’Arciprete, R.; Stella, M.; Fornaro, M.; Ciccocioppo, R.; Capri, M.G.; Naglieri, A.M.; Alberti, S. High-efficiency expression gene cloning by flow cytometry. J. Histochem. Cytochem. 1996, 44, 629–640. [Google Scholar] [CrossRef]
- Alberti, S.; Bucci, C.; Fornaro, M.; Robotti, A.; Stella, M. Immunofluorescence analysis in flow cytometry: Better selection of antibody-labeled cells after fluorescence overcompensation in the red channel. J. Histochem. Cytochem. 1991, 39, 701–706. [Google Scholar] [CrossRef] [PubMed]
- Alberti, S.; Parks, D.R.; Herzenberg, L.A. A single laser method for subtraction of cell autofluorescence in flow cytometry. Cytometry 1987, 8, 114–119. [Google Scholar] [CrossRef]
- Kabat, E.A.; Wu, T.T.; Perry, H.M.; Gottesman, K.S.; Foeller, C. Sequences of Proteins of Immunological Interests, 5th ed.; NIH Publication No 91-3242; Diane Pub. Co.: Darby, PA, USA, 1991. [Google Scholar]
- Dersimonian, H.; Schwartz, R.S.; Barrett, K.J.; Stollar, B.D. Relationship of human variable region heavy chain germ-line genes to genes encoding anti-DNA autoantibodies. J. Immunol. 1987, 139, 2496–2501. [Google Scholar] [CrossRef]
- Weng, N.P.; Yu-Lee, L.Y.; Sanz, I.; Patten, B.M.; Marcus, D.M. Structure and specificities of anti-ganglioside autoantibodies associated with motor neuropathies. J. Immunol. 1992, 149, 2518–2529. [Google Scholar] [CrossRef]
- Teylaert, B.; Meurice, E.; Bobowski, M.; Harduin-Lepers, A.; Gaucher, C.; Fontayne, A.; Jorieux, S.; Delannoy, P. Molecular cloning, characterization, genomic organization and promoter analysis of the α1,6-fucosyltransferase gene (fut8) expressed in the rat hybridoma cell line YB2/0. BMC Biotechnol. 2011, 11, 1. [Google Scholar] [CrossRef]
- Ammons, W.S.; Bauer, R.J.; Horwitz, A.H.; Chen, Z.J.; Bautista, E.; Ruan, H.H.; Abramova, M.; Scott, K.R.; Dedrick, R.L. In vitro and in vivo pharmacology and pharmacokinetics of a human engineered monoclonal antibody to epithelial cell adhesion molecule. Neoplasia 2003, 5, 146–154. [Google Scholar] [CrossRef]
- Rossi, C.; Di Lena, A.; La Sorda, R.; Lattanzio, R.; Antolini, L.; Patassini, C.; Piantelli, M.; Alberti, S. Intestinal tumour chemoprevention with the antioxidant lipoic acid stimulates the growth of breast cancer. Eur. J. Cancer 2008, 44, 2696–2704. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, M.; Yamaguchi, M.; Kato, K.; Nakamura, K.; Shiina, S.; Ichikawa-Ando, T.; Misaka, H.; Myojo, K.; Sugimoto, Y.; Hamada, H. Pr1E11, a novel anti-TROP-2 antibody isolated by adenovirus-based antibody screening, recognizes a unique epitope. Biochem. Biophys. Res. Commun. 2015, 458, 877–882. [Google Scholar] [CrossRef] [PubMed]
- Pietras, R.J.; Pegram, M.D.; Finn, R.S.; Maneval, D.A.; Slamon, D.J. Remission of human breast cancer xenografts on therapy with humanized monoclonal antibody to HER-2 receptor and DNA-reactive drugs. Oncogene 1998, 17, 2235–2249. [Google Scholar] [CrossRef] [PubMed]
- Jefferis, R. Glycosylation as a strategy to improve antibody-based therapeutics. Nat. Rev. Drug Discov. 2009, 8, 226–234. [Google Scholar] [CrossRef] [PubMed]
- Niwa, R.; Hatanaka, S.; Shoji-Hosaka, E.; Sakurada, M.; Kobayashi, Y.; Uehara, A.; Yokoi, H.; Nakamura, K.; Shitara, K. Enhancement of the antibody-dependent cellular cytotoxicity of low-fucose IgG1 Is independent of FcgammaRIIIa functional polymorphism. Clin. Cancer Res. 2004, 10, 6248–6255. [Google Scholar] [CrossRef] [PubMed]
- Jonjic, N.; Alberti, S.; Bernasconi, S.; Peri, G.; Jìlek, P.; Anichini, A.; Parmiani, G.; Mantovani, A. Heterogeneous susceptibility of human melanoma clones to monocyte cytotoxicity: Role of ICAM-1 defined by antibody blocking and gene transfer. Eur. J. Immunol. 1992, 22, 2255–2260. [Google Scholar] [CrossRef] [PubMed]
- Truong, A.H.L.; Feng, N.; Sayegh, D.; Mak, B.C.; O’Reilly, K.; Fung, S.W.; Ceric, N.; Hahn, S.H.; Pereira, D.; Findlay, H. AR47A6.4.2, a functional naked monoclonal antibody targeting Trop-2, demonstrates in vivo efficacy in human pancreatic, colon, breast and prostate cancer models. Mol. Cancer Ther. 2007, 6, 3334s. [Google Scholar]
- Stepan, L.P.; Trueblood, E.S.; Hale, K.; Babcook, J.; Borges, L.; Sutherland, C.L. Expression of Trop2 Cell Surface Glycoprotein in Normal and Tumor Tissues: Potential Implications as a Cancer Therapeutic Target. J. Histochem. Cytochem. 2011, 59, 701–710. [Google Scholar] [CrossRef]
- Shields, R.L.; Lai, J.; Keck, R.; O’Connell, L.Y.; Hong, K.; Meng, Y.G.; Weikert, S.H.; Presta, L.G. Lack of fucose on human IgG1 N-linked oligosaccharide improves binding to human Fcgamma RIII and antibody-dependent cellular toxicity. J. Biol. Chem. 2002, 277, 26733–26740. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guerra, E.; Trerotola, M.; Relli, V.; Lattanzio, R.; Ceci, M.; Boujnah, K.; Pantalone, L.; Di Pietro, R.; Iezzi, M.; Tinari, N.; et al. The 2EF Antibody Targets a Unique N-Terminal Epitope of Trop-2 and Enhances the In Vivo Activity of the Cancer-Selective 2G10 Antibody. Cancers 2023, 15, 3721. https://doi.org/10.3390/cancers15143721
Guerra E, Trerotola M, Relli V, Lattanzio R, Ceci M, Boujnah K, Pantalone L, Di Pietro R, Iezzi M, Tinari N, et al. The 2EF Antibody Targets a Unique N-Terminal Epitope of Trop-2 and Enhances the In Vivo Activity of the Cancer-Selective 2G10 Antibody. Cancers. 2023; 15(14):3721. https://doi.org/10.3390/cancers15143721
Chicago/Turabian StyleGuerra, Emanuela, Marco Trerotola, Valeria Relli, Rossano Lattanzio, Martina Ceci, Khouloud Boujnah, Ludovica Pantalone, Roberta Di Pietro, Manuela Iezzi, Nicola Tinari, and et al. 2023. "The 2EF Antibody Targets a Unique N-Terminal Epitope of Trop-2 and Enhances the In Vivo Activity of the Cancer-Selective 2G10 Antibody" Cancers 15, no. 14: 3721. https://doi.org/10.3390/cancers15143721
APA StyleGuerra, E., Trerotola, M., Relli, V., Lattanzio, R., Ceci, M., Boujnah, K., Pantalone, L., Di Pietro, R., Iezzi, M., Tinari, N., & Alberti, S. (2023). The 2EF Antibody Targets a Unique N-Terminal Epitope of Trop-2 and Enhances the In Vivo Activity of the Cancer-Selective 2G10 Antibody. Cancers, 15(14), 3721. https://doi.org/10.3390/cancers15143721