
Preclinical Models of Neuroblastoma—Current Status and Perspectives

Abstract
Simple Summary
Abstract
1. Introduction
2. In Vitro Models of Neuroblastoma
2.1. Conventional 2D Cell Cultures

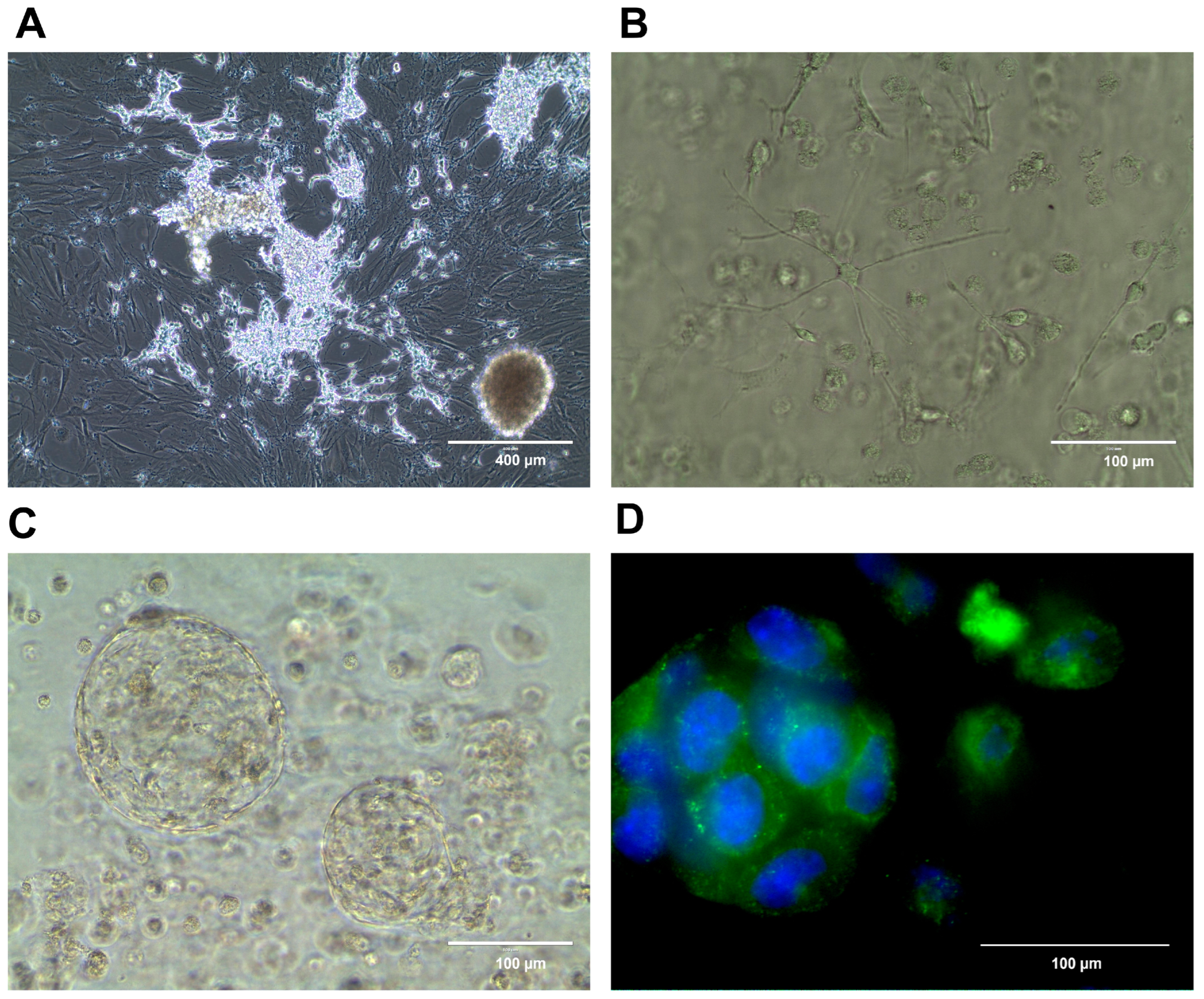{kind=link}
| Name | Origin | Stage (INSS *) | Treatment | MYCN Status | ALK Status | P53 Status | Differentiation Status | References |
|---|---|---|---|---|---|---|---|---|
| IMR-32 | Human | none | amplified | wt | wt | adrenergic | [49,54,85,120,121] | |
| SK-N-SH | Human | 4 | CT/RT | non-amplified | mut | wt | adrenergic | [54,85,105,122,123,124] |
| SK-SY5Y | Human; thrice cloned (SK-N-SH -> SH-SY -> SH-SY5 -> SH-SY5Y) subline of SK-N-SH | 4 | CT/RT | non-amplified | mut | wt | adrenergic | [50,54,85,124,125,126] |
| SK-N-BE(2) | Human | 4 | CT/RT | amplified | wt | wt | [49,85,125,127] | |
| BE(2)-C | Human; clone of SK-N-BE(2) | 4 | amplified | adrenergic | [54,125,128] | |||
| BE(2)-M17 | Human; clone of SK-N-BE(2) | 4 | [125] | |||||
| Neuro-2a | Mouse | - | [79] | |||||
| SK-N-FI | Human | 4 | CT | non-amplified | wt | mut | adrenergic | [54,77,85,105] |
| SK-N-DZ | Human | 4 | amplified | wt | wt | [78,85,123] | ||
| B35 | Rat | - | [66] | |||||
| SK-N-AS | Human | 4 | non-amplified | wt | wt | mesenchymal | [50,54,77,123,124] | |
| N1E-115 | Mouse | - | [65] | |||||
| NBFL | Human | 4 | [64] | |||||
| CHP-212 | Human | amplified | [49,128] | |||||
| NB41A3 | Mouse | - | [62] |
2.2. Conditionally Reprogrammed Cells
2.3. 3D Cell Cultures
2.4. Complex In Vitro Cultures
2.5. Models of Neuroblastoma Initiation
3. Animal Models of Neuroblastoma
3.1. Genetically Engineered Mouse Models
3.2. Syngeneic Murine Models
3.3. Xenograft Murine Models
3.4. Humanized Mice and Other Future Murine Neuroblastoma Models
| Model | Strengths | Limitations | Human Relevance | References |
|---|---|---|---|---|
| Syngeneic mouse models |
|
| low | [16,117] |
| Transgenic mouse models |
|
| intermediate | [186,193,195,199,209] |
| Human xenograft murine models |
|
| low | [219,220,221] |
| PDX models |
|
| high | [223,224,225,228] |
| Zebrafish models |
|
| low | [117,236] |
3.5. Other Animal Models Proposed for Neuroblastoma Research
3.6. Models of Neuroblastoma Metastasis
4. Conclusions and Future Directions
Author Contributions
Funding
Conflicts of Interest
References
- Kamb, A. What’s Wrong with Our Cancer Models? Nat. Rev. Drug Discov. 2005, 4, 161–165. [Google Scholar] [CrossRef]
- Krüger, M.; Kopp, S. Tumor Models and Drug Targeting In Vitro—Where Are We Today? Where Do We Go from Here? Cancers 2023, 15, 1768. [Google Scholar] [CrossRef]
- He, Z.; Wilson, A.; Rich, F.; Kenwright, D.; Stevens, A.; Low, Y.S.; Thunders, M. Chromosomal Instability and Its Effect on Cell Lines. Cancer Rep. 2023, 6, e1822. [Google Scholar] [CrossRef] [PubMed]
- Movia, D.; Prina-Mello, A. Cancer Cell Culture Methods and Protocols Methods in Molecular Biology; Movia, D., Prina-Mello, A., Eds.; Humana Press: Totowa, NJ, USA, 2023. [Google Scholar]
- Wilding, J.L.; Bodmer, W.F. Cancer Cell Lines for Drug Discovery and Development. Cancer Res. 2014, 74, 2377–2384. [Google Scholar] [CrossRef] [PubMed]
- Voskoglou-Nomikos, T.; Pater, J.L.; Seymour, L. Clinical Predictive Value of the In Vitro Cell Line, Human Xenograft, and Mouse Allograft Preclinical Cancer Models. Clin. Cancer Res. 2003, 9, 4227–4239. [Google Scholar] [PubMed]
- Biju, T.S.; Priya, V.V.; Francis, A.P. Role of Three-Dimensional Cell Culture in Therapeutics and Diagnostics: An Updated Review. Drug Deliv. Transl. Res. 2023. online ahead of print. [Google Scholar] [CrossRef]
- Ye, F.; Chen, C.; Qin, J.; Liu, J.; Zheng, A.C. Genetic Profiling Reveals an Alarming Rate of Cross-Contamination among Human Cell Lines Used in China. FASEB J. 2015, 29, 4268–4272. [Google Scholar] [CrossRef]
- Drexler, H.G.; Dirks, W.G.; Matsuo, Y.; MacLeod, R.A.F. False Leukemia-Lymphoma Cell Lines: An Update on over 500 Cell Lines. Leukemia 2003, 17, 416–426. [Google Scholar] [CrossRef]
- Lacroix, M. Persistent Use of “False” Cell Lines. Int. J. Cancer 2008, 122, 1–4. [Google Scholar] [CrossRef]
- Abbot, A. Cell Culture: Biology’s New Dimension. Nature 2003, 424, 870–872. [Google Scholar] [CrossRef]
- Shay, J.; Wright, W. Hayflick, His Limit, and Cellular Ageing. Nat. Rev. Mol. Cell Biol. 2000, 1, 72–76. [Google Scholar] [CrossRef] [PubMed]
- Balmain, A.; Harris, C.C. Carcinogenesis in Mouse and Human Cells: Parallels and Paradoxes. Carcinogenesis 2000, 21, 371–377. [Google Scholar] [CrossRef] [PubMed]
- Hann, B.; Balmain, A. Building “validated” Mouse Models of Human Cancer. Curr. Opin. Cell Biol. 2001, 13, 778–784. [Google Scholar] [CrossRef]
- Jonkers, J.; Berns, A. Conditional Mouse Models of Sporadic Cancer. Nat. Rev. Cancer 2002, 2, 251–265. [Google Scholar] [CrossRef]
- Seitz, G.; Armeanu-Ebinger, S.; Warmann, S.; Fuchs, J. Animal Models of Extracranial Pediatric Solid Tumors (Review). Oncol. Lett. 2012, 4, 859–864. [Google Scholar] [CrossRef]
- Dhiman, H.K.; Ray, A.R.; Panda, A.K. Three-Dimensional Chitosan Scaffold-Based MCF-7 Cell Culture for the Determination of the Cytotoxicity of Tamoxifen. Biomaterials 2005, 26, 979–986. [Google Scholar] [CrossRef]
- Weigelt, B.; Ghajar, C.M.; Bissell, M.J. The Need for Complex 3D Culture Models to Unravel Novel Pathways and Identify Accurate Biomarkers in Breast Cancer. Adv. Drug Deliv. Rev 2014, 69–70, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Gao, Y.; Hao, Y.; Li, E.; Wang, Y.; Zhang, J.; Wang, W.; Gao, Z.; Wang, Q. Application of a Microfluidic Chip-Based 3D Co-Culture to Test Drug Sensitivity for Individualized Treatment of Lung Cancer. Biomaterials 2013, 34, 4109–4117. [Google Scholar] [CrossRef]
- Lv, D.; Hu, Z.; Lu, L.; Lu, H.; Xu, X. Three-Dimensional Cell Culture: A Powerful Tool in Tumor Research and Drug Discovery. Oncol. Lett. 2017, 14, 6999–7010. [Google Scholar] [CrossRef]
- Ito, T.; Kawai, Y.; Yasui, Y.; Iriguchi, S.; Minagawa, A.; Ishii, T.; Miyoshi, H.; Taketo, M.M.; Kawada, K.; Obama, K.; et al. The Therapeutic Potential of Multiclonal Tumoricidal T Cells Derived from Tumor Infiltrating Lymphocyte-1derived IPS Cells. Commun. Biol. 2021, 4, 694. [Google Scholar] [CrossRef]
- Grskovic, M.; Javaherian, A.; Strulovici, B.; Daley, G.Q. Induced Pluripotent Stem Cells-Opportunities for Disease Modelling and Drug Discovery. Nat. Rev. Drug Discov. 2011, 10, 915–929. [Google Scholar] [CrossRef] [PubMed]
- Yuan, H.; Myers, S.; Wang, J.; Zhou, D.; Woo, J.A.; Kallakury, B.; Ju, A.; Bazylewicz, M.; Carter, Y.M.; Albanese, C.; et al. Use of Reprogrammed Cells to Identify Therapy for Respiratory Papillomatosis. N. Engl. J. Med. 2012, 367, 1220–1227. [Google Scholar] [CrossRef] [PubMed]
- Yuan, H.; Krawczyk, E.; Blancato, J.; Albanese, C.; Zhou, D.; Wang, N.; Paul, S.; Alkhilaiwi, F.; Palechor-Ceron, N.; Dakic, A.; et al. HPV Positive Neuroendocrine Cervical Cancer Cells Are Dependent on Myc but Not E6/E7 Viral Oncogenes. Sci. Rep. 2017, 7, srep45617. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Krawczyk, E.; Suprynowicz, F.A.; Palechor-Ceron, N.; Yuan, H.; Dakic, A.; Simic, V.; Zheng, Y.L.; Sripadhan, P.; Chen, C.; et al. Conditional Reprogramming and Long-Term Expansion of Normal and Tumor Cells from Human Biospecimens. Nat. Protoc. 2017, 12, 439–451. [Google Scholar] [CrossRef]
- Palechor-Ceron, N.; Krawczyk, E.; Dakic, A.; Simic, V.; Yuan, H.; Blancato, J.; Wang, W.; Hubbard, F.; Zheng, Y.L.; Dan, H.; et al. Conditional Reprogramming for Patient-Derived Cancer Models and next-Generation Living Biobanks. Cells 2019, 8, 1327. [Google Scholar] [CrossRef]
- Timofeeva, O.A.; Palechor-Ceron, N.; Li, G.; Yuan, H.; Krawczyk, E.; Zhong, X.; Liu, G.; Upadhyay, G.; Dakic, A.; Yu, S.; et al. Conditionally Reprogrammed Normal and Primary Tumor Prostate Epithelial Cells: A Novel Patient-Derived Cell Model for Studies of Human Prostate Cancer. Oncotarget 2017, 8, 22741–22758. [Google Scholar] [CrossRef]
- De La Rochere, P.; Guil-Luna, S.; Decaudin, D.; Azar, G.; Sidhu, S.S.; Piaggio, E. Humanized Mice for the Study of Immuno-Oncology. Trends Immunol. 2018, 39, 748–763. [Google Scholar] [CrossRef]
- Zhou, Y.; Xia, J.; Xu, S.; She, T.; Zhang, Y.; Sun, Y.; Wen, M.; Jiang, T.; Xiong, Y.; Lei, J. Experimental Mouse Models for Translational Human Cancer Research. Front. Immunol. 2023, 14, 1095388. [Google Scholar] [CrossRef]
- Mondal, A.M.; Ma, A.H.; Li, G.; Krawczyk, E.; Yuan, R.; Lu, J.; Schlegel, R.; Stamatakis, L.; Kowalczyk, K.J.; Philips, G.K.; et al. Fidelity of a PDX-CR Model for Bladder Cancer. Biochem. Biophys. Res. Commun. 2019, 517, 49–56. [Google Scholar] [CrossRef]
- Khamis, Z.I.; Sarker, D.B.; Xue, Y.; Al-Akkary, N.; James, V.D.; Zeng, C.; Li, Y.; Sang, Q.X.A. Modeling Human Brain Tumors and the Microenvironment Using Induced Pluripotent Stem Cells. Cancers 2023, 15, 1253. [Google Scholar] [CrossRef]
- Luo, J.; Li, P. Human Pluripotent Stem Cell-Derived Brain Organoids as In Vitro Models for Studying Neural Disorders and Cancer. Cell Biosci. 2021, 11, 99. [Google Scholar] [CrossRef] [PubMed]
- Novelli, G.; Spitalieri, P.; Murdocca, M.; Centanini, E.; Sangiuolo, F. Organoid Factory: The Recent Role of the Human Induced Pluripotent Stem Cells (HiPSCs) in Precision Medicine. Front. Cell Dev. Biol. 2023, 10, 1059579. [Google Scholar] [CrossRef] [PubMed]
- Jung, O.; Song, M.J.; Ferrer, M. Operationalizing the Use of Biofabricated Tissue Models as Preclinical Screening Platforms for Drug Discovery and Development. SLAS Discov. 2021, 26, 1164–1176. [Google Scholar] [CrossRef] [PubMed]
- Browning, J.R.; Derr, P.; Derr, K.; Doudican, N.; Michael, S.; Lish, S.R.; Taylor, N.A.; Krueger, J.G.; Ferrer, M.; Carucci, J.A.; et al. A 3D Biofabricated Cutaneous Squamous Cell Carcinoma Tissue Model with Multi-Channel Confocal Microscopy Imaging Biomarkers to Quantify Antitumor Effects of Chemotherapeutics in Tissue. Oncotarget 2020, 11, 2587–2596. [Google Scholar] [CrossRef]
- Pinto, N.R.; Applebaum, M.A.; Volchenboum, S.L.; Matthay, K.K.; London, W.B.; Ambros, P.F.; Nakagawara, A.; Berthold, F.; Schleiermacher, G.; Park, J.R.; et al. Advances in Risk Classification and Treatment Strategies for Neuroblastoma. J. Clin. Oncol. 2015, 33, 3008–3017. [Google Scholar] [CrossRef] [PubMed]
- Kuzyk, A.; Gartner, J.; Mai, S. Identification of Neuroblastoma Subgroups Based on Three-Dimensional Telomere Organization. Transl. Oncol. 2016, 9, 348–356. [Google Scholar] [CrossRef] [PubMed]
- Gheytanchi, E.; Mehrazma, M.; Madjd, Z. Expression of Ki-67, P53 and VEGF in Pediatric Neuroblastoma. Asian Pac. J. Cancer Prev. 2014, 15, 3065–3070. [Google Scholar] [CrossRef]
- Maris, J.M. Recent Advances in Neuroblastoma. N. Engl. J. Med. 2010, 362, 2202–2211. [Google Scholar] [CrossRef]
- Matthay, K.K.; Maris, J.M.; Schleiermacher, G.; Nakagawara, A.; Mackall, C.L.; Diller, L.; Weiss, W.A. Neuroblastoma. Nat. Rev. Dis. Primers 2016, 2, 16078. [Google Scholar] [CrossRef]
- Ackermann, S.; Cartolano, M.; Hero, B.; Welte, A.; Kahlert, Y.; Roderwieser, A.; Bartenhagen, C.; Walter, E.; Gecht, J.; Kerschke, L.; et al. A Mechanistic Classification of Clinical Phenotypes in Neuroblastoma. Science 2018, 362, 1165–1170. [Google Scholar] [CrossRef]
- DuBois, S.G.; Macy, M.E.; Henderson, T.O. High-Risk and Relapsed Neuroblastoma: Toward More Cures and Better Outcomes. Am. Soc. Clin. Oncol. Educ. Book 2022, 42, 768–780. [Google Scholar] [CrossRef]
- Cheung, N.K.V.; Dyer, M.A. Neuroblastoma: Developmental Biology, Cancer Genomics and Immunotherapy. Nat. Rev. Cancer 2013, 13, 397–411. [Google Scholar] [CrossRef]
- Brodeur, G.M. Neuroblastoma: Biological Insights into a Clinical Enigma. Nat. Rev. Cancer 2003, 3, 203–216. [Google Scholar] [CrossRef] [PubMed]
- Speleman, F.; Park, J.R.; Henderson, T.O. Neuroblastoma: A Tough Nut to Crack. ASCO Educ. Book 2016, 35, e548–e557. [Google Scholar] [CrossRef] [PubMed]
- Matthay, K.K.; George, R.E.; Yu, A.L. Promising Therapeutic Targets in Neuroblastoma. Clin. Cancer Res. 2012, 18, 2740–2753. [Google Scholar] [CrossRef]
- Castel, V.; Segura, V.; Berlanga, P. Emerging Drugs for Neuroblastoma. Expert Opin. Emerg. Drugs 2013, 18, 155–171. [Google Scholar] [CrossRef]
- Ornell, K.J.; Coburn, J.M. Developing Preclinical Models of Neuroblastoma: Driving Therapeutic Testing. BMC Biomed. Eng. 2019, 1, 33. [Google Scholar] [CrossRef]
- Thiele, C.J. Neuroblastoma Cell Lines. In Human Cell Culture Volume I: Cancer Cell Lines Part 1: Chapter 2 Neuroblastoma; Masters, J.R.W., Palsson, B.O., Eds.; Kluwer Academic Publishers: New York, NY, USA, 1999. [Google Scholar]
- Campos Cogo, S.; Gradowski Farias da Costa do Nascimento, T.; de Almeida Brehm Pinhatti, F.; de França Junior, N.; Santos Rodrigues, B.; Regina Cavalli, L.; Elifio-Esposito, S. An Overview of Neuroblastoma Cell Lineage Phenotypes and In Vitro Models. Exp. Biol. Med. 2020, 245, 1637–1647. [Google Scholar] [CrossRef]
- Paolini, L.; Hussain, S.; Galardy, P.J. Chromosome Instability in Neuroblastoma: A Pathway to Aggressive Disease. Front. Oncol. 2022, 12, 988972. [Google Scholar] [CrossRef]
- Attiyeh, E.F.; London, W.B.; Mossé, Y.P.; Wang, Q.; Winter, C.; Khazi, D.; McGrady, P.W.; Seeger, R.C.; Thomas Look, A.; Shimada, H.; et al. Chromosome 1p and 11q Deletions and Outcome in Neuroblastoma. N. Engl. J. Med. 2005, 24, 2243–2253. [Google Scholar] [CrossRef]
- Parvin, S.; Akter, J.; Takenobu, H.; Katai, Y.; Satoh, S.; Okada, R.; Haruta, M.; Mukae, K.; Wada, T.; Ohira, M.; et al. ATM Depletion Induces Proteasomal Degradation of FANCD2 and Sensitizes Neuroblastoma Cells to PARP Inhibitors. BMC Cancer 2023, 23, 313. [Google Scholar] [CrossRef]
- van Groningen, T.; Koster, J.; Valentijn, L.J.; Zwijnenburg, D.A.; Akogul, N.; Hasselt, N.E.; Broekmans, M.; Haneveld, F.; Nowakowska, N.E.; Bras, J.; et al. Neuroblastoma Is Composed of Two Super-Enhancer-Associated Differentiation States. Nat. Genet. 2017, 49, 1261–1266. [Google Scholar] [CrossRef]
- van Groningen, T.; Akogul, N.; Westerhout, E.M.; Chan, A.; Hasselt, N.E.; Zwijnenburg, D.A.; Broekmans, M.; Stroeken, P.; Haneveld, F.; Hooijer, G.K.J.; et al. A NOTCH Feed-Forward Loop Drives Reprogramming from Adrenergic to Mesenchymal State in Neuroblastoma. Nat. Commun. 2019, 10, 1530. [Google Scholar] [CrossRef] [PubMed]
- Shmakova, A.A.; Klimovich, P.S.; Rysenkova, K.D.; Popov, V.S.; Gorbunova, A.S.; Karpukhina, A.A.; Karagyaur, M.N.; Rubina, K.A.; Tkachuk, V.A.; Semina, E.V. Urokinase Receptor UPAR Downregulation in Neuroblastoma Leads to Dormancy, Chemoresistance and Metastasis. Cancers 2022, 14, 994. [Google Scholar] [CrossRef]
- Huertas-Castaño, C.; Gómez-Muñoz, M.A.; Pardal, R.; Vega, F.M. Hypoxia in the Initiation and Progression of Neuroblastoma Tumours. Int. J. Mol. Sci. 2020, 21, 39. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Cai, W.; Li, S.; Da, Z.; Sun, H.; Ma, L.; Lin, Y.; Zhi, D. Characterization of Neuroblastoma Bone Invasion/Metastasis in Established Bone Metastatic Model of SY5Y and KCNR Cell Lines. Child’s Nerv. Syst. 2013, 29, 1097–1105. [Google Scholar] [CrossRef] [PubMed]
- Cuperus, R.; Tytgat, G.A.M.; Leen, R.; Brites, P.; Bras, J.; Caron, H.N.; Van Kuilenburg, A.P.B. Pleiotropic Effects of Fenretinide in Neuroblastoma Cell Lines and Multicellular Tumor Spheroids. Int. J. Oncol. 2008, 32, 1011–1019. [Google Scholar] [CrossRef] [PubMed]
- Roomi, M.W.; Kalinovsky, T.; Roomi, N.W.; Niedzwiecki, A.; Rath, M. Inhibition of the SK-N-MC Human Neuroblastoma Cell Line In Vivo and In Vitro by a Novel Nutrient Mixture. Oncol. Rep. 2013, 29, 1714–1720. [Google Scholar] [CrossRef]
- Lu, C.; Everhart, L.; Tilan, J.; Kuo, L.; Sun, C.C.J.; Munivenkatappa, R.B.; Jönsson-Rylander, A.C.; Sun, J.; Kuan-Celarier, A.; Li, L.; et al. Neuropeptide y and Its Y2 Receptor: Potential Targets in Neuroblastoma Therapy. Oncogene 2010, 29, 5630–5642. [Google Scholar] [CrossRef]
- Thompson, J.M.; London, E.D.; Johnson, J.E. Ultrastructural, Functional and Biochemical Characteristics of Mouse and Human Neuroblastoma Cell Lines. Neuroscience 1982, 7, 1807–1815. [Google Scholar] [CrossRef]
- Gazitt, Y.; He, Y.J.; Chang, L.; Koza, S.; Fisk, D.; Graham-Pole, J. Expression of N-Myc, c-Myc, and MDR-1 Proteins in Newly Established Neuroblastoma Cell Lines: A Study by Immunofluorescence Staining and Flow Cytometry. Cancer Res. 1992, 52, 2957–2965. [Google Scholar]
- Symes, A.J.; Rao, M.S.; Lewis, S.E.; Landis, S.C.; Hyman, S.E.; Stephen Fink, J. Ciliary Neurotrophic Factor Coordinately Activates Transcription of Neuropeptide Genes in a Neuroblastoma Cell Line. Proc. Natl. Acad. Sci. USA 1993, 90, 572–576. [Google Scholar] [CrossRef]
- Amano, T.; Richelson, E.; Nirenberg, M. Neurotransmitter Synthesis by Neuroblastoma Clones (Neuroblast Differentiation/Cell Culture/Choline Acetyltransferase/Acetylcholinesterase/Tyrosine Hydroxylase/Axons-Dendrites). Proc. Natl. Acad. Sci. USA 1972, 69, 258–263. [Google Scholar] [CrossRef]
- Schubert, D.; Heinemann, S.; Carlisle, W.; Tarikas, H.; Kimes, B.; Patrick, J.; Steinbach, J.H.; Culp, W.; Brandt, B.L. Clonal Cell Lines from the Rat Central Nervous System. Nature 1974, 2, 224–227. [Google Scholar] [CrossRef] [PubMed]
- Abualsaud, N.; Caprio, L.; Galli, S.; Krawczyk, E.; Alamri, L.; Zhu, S.; Gallicano, G.I.; Kitlinska, J. Neuropeptide Y/Y5 Receptor Pathway Stimulates Neuroblastoma Cell Motility Through RhoA Activation. Front. Cell Dev. Biol. 2021, 8, 627090. [Google Scholar] [CrossRef] [PubMed]
- Rupniak, H.T.; Rein, G.; Powell, J.F.; Ryder, T.A.; Carson, S.; Povey, S.; Hill, B.T. Characteristics of a New Human Neuroblastoma Cell Line Which Differentiates in Response to Cyclic Adenosine 3′:5′-Monophosphate1. Cancer Res. 1984, 44, 2600–2607. [Google Scholar] [PubMed]
- West, G.J.; Uki, J.; Herschman, H.R.; Seeger Robert, C. Adrenergic, Cholinergic, and Inactive Human Neuroblastoma Cell Lines with the Action-Potential Na+ Ionophore. Cancer Res. 1977, 37, 1372–1376. [Google Scholar] [PubMed]
- Schlesinger, H.R.; Gerson, J.M.; Moorhead, P.S.; Maguire, H.; Hummeler, K. Establishment and Characterization of Human Neuroblastoma Cell Lines. Cancer Res. 1976, 36, 3094–3100. [Google Scholar]
- Gilbert, F.; Balaban, G.; Moorhead, P.; Bianchi, D.; Schlesinger, H. Abnormalities of Chromosome Lp in Human Neuroblastoma Tumors and Cell Lines. Cancer Genet. Cytogenet. 1982, 7, 33–42. [Google Scholar] [CrossRef]
- Naiditch, J.A.; Jie, C.; Lautz, T.B.; Yu, S.; Clark, S.; Voronov, D.; Chu, F.; Madonna, M.B. Mesenchymal Change and Drug Resistance in Neuroblastoma. J. Surg. Res. 2015, 193, 279–288. [Google Scholar] [CrossRef]
- Seeger, R.C.; Rayner, S.A.; Banerjee, A.; Chung, H.; Laug, E.; Neustein, H.B.; Benedict, W.F. Morphology, Growth, Chromosomal Pattern, and Fibrinolytic of Two New Human Neuroblastoma Cell Lines. Cancer Res. 1977, 37, 1364–1371. [Google Scholar]
- Pajtler, K.W.; Mahlow, E.; Odersky, A.; Lindner, S.; Stephan, H.; Bendix, I.; Eggert, A.; Schramm, A.; Schulte, J.H. Neuroblastoma in Dialog with Its Stroma: NTRK1 Is a Regulator of Cellular Cross-Talk with Schwann Cells. Oncotarget 2014, 5, 11180–11192. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Tsubota, S.; Nishio, N.; Takahashi, Y.; Kadomatsu, K. Combination of Tumor Necrosis Factor-α and Epidermal Growth Factor Induces the Adrenergic-to-Mesenchymal Transdifferentiation in SH-SY5Y Neuroblastoma Cells. Cancer Sci. 2021, 112, 715–724. [Google Scholar] [CrossRef]
- Wolpaw, A.J.; Grossmann, L.D.; Dessau, J.L.; Dong, M.M.; Aaron, B.J.; Brafford, P.A.; Volgina, D.; Pascual-Pasto, G.; Rodriguez-Garcia, A.; Uzun, Y.; et al. Epigenetic State Determines Inflammatory Sensing in Neuroblastoma. Proc. Natl. Acad. Sci. USA 2022, 119, e2102358119. [Google Scholar] [CrossRef]
- Iavarone, A.; Lasorella, A.; Servidei, T.; Riccardi, R.; Mastrangelo, R. Uptake and Storage of M-Lodobenzylguanidine Are Frequent Neuronal Functions of Human Neuroblastoma Cell Lines. Cancer Res. 1993, 53, 304–309. [Google Scholar] [PubMed]
- Matsumoto, M.; Akiyama, T.; Miyatake, S.; Kikuchi, H.; Hanaoka, M.; Namba, Y. Expression of Proto-Oncogene Products during Drug-Induced Differentiation of a Neuroblastoma Cell Line SK-N-DZ. Acta Neuropathol. 1989, 79, 217–221. [Google Scholar] [CrossRef]
- Olmsted, J.B.; Carlson, K.; Klebe, R.; Ruddle, F.; Rosenbaum, J. Isolation of Microtubule Protein from Cultured Mouse Neuroblastoma Cells. Proc. Natl. Acad. Sci. USA 1970, 65, 129–136. [Google Scholar] [CrossRef]
- Ross, R.A.; Walton, J.D.; Han, D.; Guo, H.F.; Cheung, N.K.V. A Distinct Gene Expression Signature Characterizes Human Neuroblastoma Cancer Stem Cells. Stem Cell Res. 2015, 15, 419–426. [Google Scholar] [CrossRef]
- Spengler, B.A.; Lazarova, D.L.; Ross, R.A.; Biedler, J.L. Cell Lineage and Differentiation State Are Primary Determinants of MYCN Gene Expression and Malignant Potential in Human Neuroblastoma Cells. Oncol. Res. 1997, 9, 467–476. [Google Scholar]
- Foley, J.; Cohn, S.L.; Salwen, H.R.; Parysek, L.M. Differential Expression of N-Myc in Phenotypically Distinct Subclones of a Human Neuroblastoma Cell Line. Cancer Res. 1991, 51, 6338–6345. [Google Scholar]
- Ciccarone, V.; Spengler, B.A.; Meyers, M.B.; Biedler, J.L.; Ross, R.A. Phenotypic Diversification in Human Neuroblastoma Cells: Expression of Distinct Neural Crest Lineages. Cancer Res. 1989, 49, 219–225. [Google Scholar]
- Chen, L.; Alexe, G.; Dharia, N.V.; Ross, L.; Iniguez, A.B.; Conway, A.S.; Wang, E.J.; Veschi, V.; Lam, N.; Qi, J.; et al. CRISPR-Cas9 Screen Reveals a MYCN-Amplified Neuroblastoma Dependency on EZH2. J. Clin. Investig. 2018, 128, 446–462. [Google Scholar] [CrossRef]
- Blanco-Luquin, I.; Lázcoz, P.; Celay, J.; Castresana, J.S.; Encío, I.J. In Vitro Assessment of the Role of P53 on Chemotherapy Treatments in Neuroblastoma Cell Lines. Pharmaceuticals 2021, 14, 1184. [Google Scholar] [CrossRef] [PubMed]
- Barretina, J.; Caponigro, G.; Stransky, N.; Venkatesan, K.; Margolin, A.A.; Kim, S.; Wilson, C.J.; Lehár, J.; Kryukov, G.V.; Sonkin, D.; et al. The Cancer Cell Line Encyclopedia Enables Predictive Modelling of Anticancer Drug Sensitivity. Nature 2012, 483, 603–607. [Google Scholar] [CrossRef]
- Park, J.H.; Szemes, M.; Vieira, G.C.; Melegh, Z.; Malik, S.; Heesom, K.J.; Von Wallwitz-Freitas, L.; Greenhough, A.; Brown, K.W.; Zheng, Y.G.; et al. Protein Arginine Methyltransferase 5 Is a Key Regulator of the MYCN Oncoprotein in Neuroblastoma Cells. Mol. Oncol. 2015, 9, 617–627. [Google Scholar] [CrossRef] [PubMed]
- Gupta, M.K.; Mallepalli, S.; Damu, A.; Vadde, R. Neuroblastoma and Stem Cell Therapy: An Updated Review. CNS Neurol. Disord. Drug Targets 2020, 20, 625–643. [Google Scholar] [CrossRef] [PubMed]
- Saintas, E.; Abrahams, L.; Ahmad, G.T.; Ajakaiye, A.O.M.; Alhumaidi, A.S.H.A.M.; Ashmore-Harris, C.; Clark, I.; Dura, U.K.; Fixmer, C.N.; Ike-Morris, C.; et al. Acquired Resistance to Oxaliplatin Is Not Directly Associated with Increased Resistance to DNA Damage in SK-N-ASrOXALI4000, a Newly Established Oxaliplatin-Resistant Sub-Line of the Neuroblastoma Cell Line SK-N-AS. PLoS ONE 2017, 12, e0172140. [Google Scholar] [CrossRef]
- Sitnikov, D.; Revkova, V.; Ilina, I.; Shatalova, R.; Komarov, P.; Struleva, E.; Konoplyannikov, M.; Kalsin, V.; Baklaushev, V. Sensitivity of Neuroblastoma and Induced Neural Progenitor Cells to High-Intensity THz Radiation. Int. J. Mol. Sci. 2023, 24, 6558. [Google Scholar] [CrossRef]
- Cornel, A.M.; Dunnebach, E.; Hofman, D.A.; Das, S.; Sengupta, S.; Van Den Ham, F.; Wienke, J.; Strijker, J.G.M.; Van Den Beemt, D.A.M.H.; Essing, A.H.W.; et al. Epigenetic Modulation of Neuroblastoma Enhances T Cell and NK Cell Immunogenicity by Inducing a Tumor-Cell Lineage Switch. J. Immunother. Cancer 2022, 10, e005002. [Google Scholar] [CrossRef]
- Jiménez, C.; Moreno, L.; Segura, M.F. Epigenetic Therapies for Neuroblastoma: Immunogenicity Awakens. Mol. Oncol. 2023, 17, 718–721. [Google Scholar] [CrossRef]
- Iguchi, M.; Yagyu, S.; Kambe, K.; Higashi, M.; Fumino, S.; Kishida, T.; Iehara, T.; Mazda, O.; Tajiri, T. Development of Anti-GD2 Antibody-Producing Mesenchymal Stem Cells as Cellular Immunotherapy. Anticancer Res. 2023, 43, 2417–2424. [Google Scholar] [CrossRef] [PubMed]
- Karapurkar, J.K.; Kim, M.S.; Colaco, J.C.; Suresh, B.; Sarodaya, N.; Kim, D.H.; Park, C.H.; Hong, S.H.; Kim, K.S.; Ramakrishna, S. CRISPR/Cas9-Based Genome-Wide Screening of the Deubiquitinase Subfamily Identifies USP3 as a Protein Stabilizer of REST Blocking Neuronal Differentiation and Promotes Neuroblastoma Tumorigenesis. J. Exp. Clin. Cancer Res. 2023, 42, 121. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Monleon, A.; Gaarder, J.; Djos, A.; Kogner, P.; Fransson, S. Identification of Recurrent 3q13.31 Chromosomal Rearrangement Indicates LSAMP as a Tumor Suppressor Gene in Neuroblastoma. Int. J. Oncol. 2023, 62, 27. [Google Scholar] [CrossRef]
- Eleveld, T.F.; Bakali, C.; Eijk, P.P.; Stathi, P.; Vriend, L.E.; Poddighe, P.J.; Ylstra, B. Engineering Large-Scale Chromosomal Deletions by CRISPR-Cas9. Nucleic Acids Res. 2021, 49, 12007–12016. [Google Scholar] [CrossRef]
- Sanmartín, E.; Muñoz, L.; Piqueras, M.; Sirerol, J.A.; Berlanga, P.; Cañete, A.; Castel, V.; Font De Mora, J. Deletion of 11q in Neuroblastomas Drives Sensitivity to PARP Inhibition. Clin. Cancer Res. 2017, 23, 6875–6887. [Google Scholar] [CrossRef]
- López-Carrasco, A.; Martín-Vañó, S.; Burgos-Panadero, R.; Monferrer, E.; Berbegall, A.P.; Fernández-Blanco, B.; Navarro, S.; Noguera, R. Impact of Extracellular Matrix Stiffness on Genomic Heterogeneity in MYCN-Amplified Neuroblastoma Cell Line. J. Exp. Clin. Cancer Res. 2020, 39, 226. [Google Scholar] [CrossRef] [PubMed]
- Kasemeier-Kulesa, J.C.; Schnell, S.; Woolley, T.; Spengler, J.A.; Morrison, J.A.; McKinney, M.C.; Pushel, I.; Wolfe, L.A.; Kulesa, P.M. Predicting Neuroblastoma Using Developmental Signals and a Logic-Based Model. Biophys. Chem. 2018, 238, 30–38. [Google Scholar] [CrossRef]
- de Medeiros, L.M.; De Bastiani, M.A.; Rico, E.P.; Schonhofen, P.; Pfaffenseller, B.; Wollenhaupt-Aguiar, B.; Grun, L.; Barbé-Tuana, F.; Zimmer, E.R.; Castro, M.A.A.; et al. Cholinergic Differentiation of Human Neuroblastoma SH-SY5Y Cell Line and Its Potential Use as an In Vitro Model for Alzheimer’s Disease Studies. Mol. Neurobiol. 2019, 56, 7355–7367. [Google Scholar] [CrossRef]
- Xie, H.-R.; Hu, L.-S.; Li, G.-Y. SH-SY5Y Human Neuroblastoma Cell Line: In Vitro Cell Model of Dopaminergic Neurons in Parkinson’s Disease. Chin. Med. J. 2010, 123, 1086–1092. [Google Scholar]
- Pandhare, J.; Dash, S.; Jones, B.; Villalta, F.; Dash, C. A Novel Role of Proline Oxidase in HIV-1 Envelope Glycoprotein-Induced Neuronal Autophagy. J. Biol. Chem. 2015, 290, 25439–25451. [Google Scholar] [CrossRef]
- Shastry, P.; Basu, A.; Rajadhyaksha, M.S. Neuroblastoma Cell Lines—A Versatile In Vitro Model in Neurobiology. Int. J. Neurosci. 2001, 108, 109–126. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, C.P.; Biedler, J.L.; Spengler, B.A.; Reynolds, D.A.; Ross, R.A.; Frenkel, E.P.; Smith, R.G. Characterization of Human Neuroblastoma Cell Lines Established Before and After Therapy. J. Natl. Cancer Inst. 1986, 76, 375–387. [Google Scholar] [PubMed]
- Harenza, J.L.; DIamond, M.A.; Adams, R.N.; Song, M.M.; Davidson, H.L.; Hart, L.S.; Dent, M.H.; Fortina, P.; Reynolds, C.P.; Maris, J.M. Transcriptomic Profiling of 39 Commonly-Used Neuroblastoma Cell Lines. Sci. Data 2017, 4, 170033. [Google Scholar] [CrossRef]
- Bate-Eya, L.T.; Ebus, M.E.; Koster, J.; Den Hartog, I.J.M.; Zwijnenburg, D.A.; Schild, L.; Van Der Ploeg, I.; Dolman, M.E.M.; Caron, H.N.; Versteeg, R.; et al. Newly-Derived Neuroblastoma Cell Lines Propagated in Serum-Free Media Recapitulate the Genotype and Phenotype of Primary Neuroblastoma Tumours. Eur. J. Cancer 2014, 50, 628–637. [Google Scholar] [CrossRef] [PubMed]
- Ben-David, U.; Siranosian, B.; Ha, G.; Tang, H.; Oren, Y.; Hinohara, K.; Strathdee, C.A.; Dempster, J.; Lyons, N.J.; Burns, R.; et al. Genetic and Transcriptional Evolution Alters Cancer Cell Line Drug Response. Nature 2018, 560, 325–330. [Google Scholar] [CrossRef]
- Nestor, C.E.; Ottaviano, R.; Reinhardt, D.; Cruickshanks, H.A.; Mjoseng, H.K.; McPherson, R.C.; Lentini, A.; Thomson, J.P.; Dunican, D.S.; Pennings, S.; et al. Rapid Reprogramming of Epigenetic and Transcriptional Profiles in Mammalian Culture Systems. Genome Biol. 2015, 16, 11. [Google Scholar] [CrossRef]
- Persson, C.U.; Von Stedingk, K.; Bexell, D.; Merselius, M.; Braekeveldt, N.; Gisselsson, D.; Arsenian-Henriksson, M.; Påhlman, S.; Wigerup, C. Neuroblastoma Patient-Derived Xenograft Cells Cultured in Stem-Cell Promoting Medium Retain Tumorigenic and Metastatic Capacities but Differentiate in Serum. Sci. Rep. 2017, 7, 10274. [Google Scholar] [CrossRef]
- McNerney, K.O.; Karageorgos, S.; Ferry, G.M.; Wolpaw, A.J.; Burudpakdee, C.; Khurana, P.; Toland, C.N.; Vemu, R.; Vu, A.; Hogarty, M.D.; et al. TH-MYCN Tumors, but Not Tumor-Derived Cell Lines, Are Adrenergic Lineage, GD2+, and Responsive to Anti-GD2 Antibody Therapy. Oncoimmunology 2022, 11, 2075204. [Google Scholar] [CrossRef]
- Santillo, S. Changes in Biophysical Properties of Undifferentiated SH-SY5Y Cells During Long-Term Cultures. Neuroscience 2022, 482, 143–158. [Google Scholar] [CrossRef]
- Kelner, M.J.; Diccianni, M.B.; Yu, A.L.; Rutherford, M.R.; Estes, L.A.; Morgenstern, R. Absence of MGST1 MRNA and Protein Expression in Human Neuroblastoma Cell Lines and Primary Tissue. Free Radic. Biol. Med. 2014, 69, 167–171. [Google Scholar] [CrossRef]
- Grainger, D.W. Cell-Based Drug Testing; This World Is Not Flat. Adv. Drug Deliv. Rev. 2014, 69–70, 7–11. [Google Scholar] [CrossRef]
- Proietto, M.; Crippa, M.; Damiani, C.; Pasquale, V.; Sacco, E.; Vanoni, M.; Gilardi, M. Tumor Heterogeneity: Preclinical Models, Emerging Technologies, and Future Applications. Front. Oncol. 2023, 13, 1164535. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.; Zhou, J.; Xiao, Q.; Fujiwara, K.; Zhang, M.; Mo, G.; Gong, W.; Zheng, L. Cancer-Associated Fibroblast Heterogeneity Is Associated with Organ-Specific Metastasis in Pancreatic Ductal Adenocarcinoma. J. Hematol. Oncol. 2021, 14, 184. [Google Scholar] [CrossRef] [PubMed]
- Hickman, J.A.; Graeser, R.; de Hoogt, R.; Vidic, S.; Brito, C.; Gutekunst, M.; van der Kuip, H. Imi Predect consortium Three-Dimensional Models of Cancer for Pharmacology and Cancer Cell Biology: Capturing Tumor Complexity In Vitro/Ex Vivo. Biotechnol. J. 2014, 9, 1115–1128. [Google Scholar] [CrossRef]
- Nolan, J.C.; Frawley, T.; Tighe, J.; Soh, H.; Curtin, C.; Piskareva, O. Preclinical Models for Neuroblastoma: Advances and Challenges. Cancer Lett. 2020, 474, 53–62. [Google Scholar] [CrossRef]
- Corallo, D.; Frabetti, S.; Candini, O.; Gregianin, E.; Dominici, M.; Fischer, H.; Aveic, S. Emerging Neuroblastoma 3D In Vitro Models for Pre-Clinical Assessments. Front. Immunol. 2020, 11, 584214. [Google Scholar] [CrossRef] [PubMed]
- Quinn, C.H.; Beierle, A.M.; Beierle, E.A. Artificial Tumor Microenvironments in Neuroblastoma. Cancers 2021, 13, 1629. [Google Scholar] [CrossRef]
- Tumilowicz, J.J.; Nichols, W.W.; Cholon, J.J.; Greene, A.E. Definition of a Continuous Human Cell Line Derived from Neuroblastoma. Cancer Res. 1970, 30, 2110–2118. [Google Scholar]
- Reynolds, C.P.; Regino Perez-Polo, J. Human neuroblastoma: Glial induced morphological differentiation. Neurosci. Lett. 1975, 1, 91–97. [Google Scholar] [CrossRef]
- Biedler, J.L.; Helson, L.; Spengler, B.A. Morphology and Growth, Tumorigenicity, and Cytogenetics of Human Neuroblastoma Cells in Continuous Culture. Cancer Res. 1973, 33, 2643–2652. [Google Scholar]
- Mossé, Y.P.; Laudenslager, M.; Longo, L.; Cole, K.A.; Wood, A.; Attiyeh, E.F.; Laquaglia, M.J.; Sennett, R.; Lynch, J.E.; Perri, P.; et al. Identification of ALK as a Major Familial Neuroblastoma Predisposition Gene. Nature 2008, 455, 930–935. [Google Scholar] [CrossRef] [PubMed]
- Levesque, A.A.; Pappalardo, R.M.; Puli, P.; Enzor, L.A.; Angeles, C. P53 Oligomerization Status as an Indicator of Sensitivity of P53-Wildtype Neuroblastomas to the Combination of DNA Damaging Agent and Chk1 Inhibitor. PLoS ONE 2022, 17, e0263463. [Google Scholar] [CrossRef] [PubMed]
- Biedler, J.L.; Roffler-Tarlov, S.; Schachner, M.; Freedman, L.S. Multiple Neurotransmitter Synthesis by Human Neuroblastoma Cell Lines and Clones. Cancer Res. 1978, 38, 3751–3757. [Google Scholar]
- Qin Wang, H.; Halilovic, E.; Li, X.; Liang, J.; Cao, Y.; Rakiec, D.P.; Ruddy, D.A.; Jeay, S.; Wuerthner, J.U.; Timple, N.; et al. Combined ALK and MDM2 Inhibition Increases Antitumor Activity and Overcomes Resistance in Human ALK Mutant Neuroblastoma Cell Lines and Xenograft Models. eLife 2017, 6, e17137. [Google Scholar] [CrossRef] [PubMed]
- Di Paolo, D.; Ambrogio, C.; Pastorino, F.; Brignole, C.; Martinengo, C.; Carosio, R.; Loi, M.; Pagnan, G.; Emionite, L.; Cilli, M.; et al. Selective Therapeutic Targeting of the Anaplastic Lymphoma Kinase with Liposomal SiRNA Induces Apoptosis and Inhibits Angiogenesis in Neuroblastoma. Mol. Ther. 2011, 19, 2201–2212. [Google Scholar] [CrossRef] [PubMed]
- Gu, L.; Chu, P.; Lingeman, R.; McDaniel, H.; Kechichian, S.; Hickey, R.J.; Liu, Z.; Yuan, Y.C.; Sandoval, J.A.; Fields, G.B.; et al. The Mechanism by Which MYCN Amplification Confers an Enhanced Sensitivity to a PCNA-Derived Cell Permeable Peptide in Neuroblastoma Cells. Ebiomedicine 2015, 2, 1923–1931. [Google Scholar] [CrossRef]
- Brodeur, G.M.; Pritchard, J.; Berthold, F.; Carlsen, N.L.T.; Castel, V.; Castleberry, R.P.; De Bernardi, B.; Evans, A.E.; Favrot, M.; Hedborg, F.; et al. Revisions of the International Criteria for Neuroblastoma Diagnosis, Staging, and Response to Treatment Purpose and Methods: Based on Preliminary Expe-Rience, There Was a Need for Modifications and Clarifi-Cations in the International Neuroblastoma Staging Sys-Tem (INSS) and International Neuroblastoma Response Criteria (INRC). J. Clin. Oncol. 1993, 11, 1466–1477. [Google Scholar] [PubMed]
- Liu, X.; Ory, V.; Chapman, S.; Yuan, H.; Albanese, C.; Kallakury, B.; Timofeeva, O.A.; Nealon, C.; Dakic, A.; Simic, V.; et al. ROCK Inhibitor and Feeder Cells Induce the Conditional Reprogramming of Epithelial Cells. Am. J. Pathol. 2012, 180, 599–607. [Google Scholar] [CrossRef] [PubMed]
- Suprynowicz, F.A.; Upadhyay, G.; Krawczyk, E.; Kramer, S.C.; Hebert, J.D.; Liu, X.; Yuan, H.; Cheluvaraju, C.; Clapp, P.W.; Boucher, R.C.; et al. Conditionally Reprogrammed Cells Represent a Stem-like State of Adult Epithelial Cells. Proc. Natl. Acad. Sci. USA 2012, 109, 20035–20040. [Google Scholar] [CrossRef]
- Saeed, K.; Rahkama, V.; Eldfors, S.; Bychkov, D.; Mpindi, J.P.; Yadav, B.; Paavolainen, L.; Aittokallio, T.; Heckman, C.; Wennerberg, K.; et al. Comprehensive Drug Testing of Patient-Derived Conditionally Reprogrammed Cells from Castration-Resistant Prostate Cancer. Eur. Urol. 2017, 71, 319–327. [Google Scholar] [CrossRef]
- Alamri, A.M.; Liu, X.; Blancato, J.K.; Haddad, B.R.; Wang, W.; Zhong, X.; Choudhary, S.; Krawczyk, E.; Kallakury, B.V.; Davidson, B.J.; et al. Expanding Primary Cells from Mucoepidermoid and Other Salivary Gland Neoplasms for Genetic and Chemosensitivity Testing. DMM Dis. Model. Mech. 2018, 11, dmm031716. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Thomas, C.; Shrivastava, N.; Gersten, A.; Gadsden, N.; Schlecht, N.; Kawachi, N.; Schiff, B.A.; Smith, R.V.; Rosenblatt, G.; et al. Establishment of a Diverse Head and Neck Squamous Cancer Cell Bank Using Conditional Reprogramming Culture Methods. J. Med. Virol. 2023, 95, e28388. [Google Scholar] [CrossRef] [PubMed]
- Alothman, S.J.; Kang, K.; Liu, X.; Krawczyk, E.; Azhar, R.I.; Hu, R.; Goerlitz, D.; Kallakury, B.V.; Furth, P.A. Characterization of Transcriptome Diversity and In Vitro Behavior of Primary Human High-Risk Breast Cells. Sci. Rep. 2022, 12, 6159. [Google Scholar] [CrossRef]
- Su, S.; Di Poto, C.; Roy, R.; Liu, X.; Cui, W.; Kroemer, A.; Ressom, H.W. Highlight Article: Long-Term Culture and Characterization of Patient-Derived Primary Hepatocytes Using Conditional Reprogramming. Exp. Biol. Med. 2019, 244, 857–864. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Bai, Q.; Chen, Y.; Ye, L.; Wu, X.; Long, X.; Ye, L.; Liu, J.; Li, H. Conditionally Reprogrammed Human Normal Bronchial Epithelial Cells Express Comparable Levels of Cytochromes P450 and Are Sensitive to BaP Induction. Biochem. Biophys. Res. Commun. 2018, 503, 2132–2138. [Google Scholar] [CrossRef] [PubMed]
- Moorefield, E.C.; Blue, R.E.; Quinney, N.L.; Gentzsch, M.; Ding, S. Generation of Renewable Mouse Intestinal Epithelial Cell Monolayers and Organoids for Functional Analyses. BMC Cell Biol. 2018, 19, 15. [Google Scholar] [CrossRef]
- Mahajan, A.S.; Sugita, B.M.; Duttargi, A.N.; Saenz, F.; Krawczyk, E.; McCutcheon, J.N.; Fonseca, A.S.; Kallakury, B.; Pohlmann, P.; Gusev, Y.; et al. Genomic Comparison of Early-Passage Conditionally Reprogrammed Breast Cancer Cells to Their Corresponding Primary Tumors. PLoS ONE 2017, 12, e0186190. [Google Scholar] [CrossRef]
- Jin, L.; Qu, Y.; Gomez, L.J.; Chung, S.; Han, B.; Gao, B.; Yue, Y.; Gong, Y.; Liu, X.; Amersi, F.; et al. Characterization of Primary Human Mammary Epithelial Cells Isolated and Propagated by Conditional Reprogrammed Cell Culture. Oncotarget 2018, 9, 11503. [Google Scholar] [CrossRef]
- Krawczyk, E.; Hong, S.H.; Galli, S.; Trinh, E.; Wietlisbach, L.; Misiukiewicz, S.F.; Tilan, J.U.; Chen, Y.S.; Schlegel, R.; Kitlinska, J. Murine Neuroblastoma Cell Lines Developed by Conditional Reprogramming Preserve Heterogeneous Phenotypes Observed In Vivo. Lab. Investig. 2020, 100, 38–51. [Google Scholar] [CrossRef]
- Festing, S.; Wilkinson, R. The Ethics of Animal Research. Talking Point on the Use of Animals in Scientific Research. EMBO Rep. 2007, 8, 526–530. [Google Scholar] [CrossRef]
- Miserocchi, G.; Bocchini, M.; Cortesi, M.; Arienti, C.; De Vita, A.; Liverani, C.; Mercatali, L.; Bravaccini, S.; Ulivi, P.; Zanoni, M. Combining Preclinical Tools and Models to Unravel Tumor Complexity: Jump into the next Dimension. Front. Immunol. 2023, 14, 1171141. [Google Scholar] [CrossRef] [PubMed]
- Tosca, E.M.; Ronchi, D.; Facciolo, D.; Magni, P. Replacement, Reduction, and Refinement of Animal Experiments in Anticancer Drug Development: The Contribution of 3D In Vitro Cancer Models in the Drug Efficacy Assessment. Biomedicines 2023, 11, 1058. [Google Scholar] [CrossRef] [PubMed]
- Kimlin, L.C.; Casagrande, G.; Virador, V.M. In Vitro Three-Dimensional (3D) Models in Cancer Research: An Update. Mol. Carcinog. 2013, 52, 167–182. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.; Restan Perez, M.; da Silva, V.A.; Thomsen, J.; Bhardwaj, L.; Andrade, T.; Alhussan, A.; Willerth, S.M. 3D Bioprinting Complex Models of Cancer. Biomater. Sci. 2023, 11, 3414–3430. [Google Scholar] [CrossRef]
- Hirschhaeuser, F.; Menne, H.; Dittfeld, C.; West, J.; Mueller-Klieser, W.; Kunz-Schughart, L.A. Multicellular Tumor Spheroids: An Underestimated Tool Is Catching up Again. J. Biotechnol. 2010, 148, 3–15. [Google Scholar] [CrossRef]
- Zanoni, M.; Piccinini, F.; Arienti, C.; Zamagni, A.; Santi, S.; Polico, R.; Bevilacqua, A.; Tesei, A. 3D Tumor Spheroid Models for in Vitro Therapeutic Screening: A Systematic Approach to Enhance the Biological Relevance of Data Obtained. Sci. Rep. 2016, 6, srep19103. [Google Scholar] [CrossRef]
- Kaess, C.; Matthes, M.; Gross, J.; Waetzig, R.; Heise, T.; Corbacioglu, S.; Sommer, G. Evaluating the RIST Molecular-Targeted Regimen in a Three-Dimensional Neuroblastoma Spheroid Cell Culture Model. Cancers 2023, 15, 1749. [Google Scholar] [CrossRef]
- Nasehi, R.; Abdallah, A.T.; Pantile, M.; Zanon, C.; Vogt, M.; Rütten, S.; Fischer, H.; Aveic, S. 3D Geometry Orchestrates the Transcriptional Landscape of Metastatic Neuroblastoma Cells in a Multicellular In Vitro Bone Model. Mater. Today Bio. 2023, 19, 100596. [Google Scholar] [CrossRef]
- Kock, A.; Bergqvist, F.; Steinmetz, J.; Elfman, L.H.M.; Korotkova, M.; Johnsen, J.I.; Jakobsson, P.J.; Kogner, P.; Larsson, K. Establishment of an In Vitro 3D Model for Neuroblastoma Enables Preclinical Investigation of Combined Tumor-Stroma Drug Targeting. FASEB J. 2020, 34, 11101–11114. [Google Scholar] [CrossRef]
- Baek, N.H.; Seo, O.W.; Kim, M.S.; Hulme, J.; An, S.S.A. Monitoring the Effects of Doxorubicin on 3D-Spheroid Tumor Cells in Real-Time. Oncol. Targets Ther. 2016, 9, 7207–7218. [Google Scholar] [CrossRef]
- Besançon, O.G.; Tytgat, G.A.M.; Meinsma, R.; Leen, R.; Hoebink, J.; Kalayda, G.V.; Jaehde, U.; Caron, H.N.; van Kuilenburg, A.B.P. Synergistic Interaction between Cisplatin and Gemcitabine in Neuroblastoma Cell Lines and Multicellular Tumor Spheroids. Cancer Lett. 2012, 319, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Craig, B.T.; Rellinger, E.J.; Alvarez, A.L.; Dusek, H.L.; Qiao, J.; Chung, D.H. Induced Differentiation Inhibits Sphere Formation in Neuroblastoma. Biochem. Biophys. Res. Commun. 2016, 477, 255–259. [Google Scholar] [CrossRef] [PubMed]
- Tesei, A.; Sarnelli, A.; Arienti, C.; Menghi, E.; Medri, L.; Gabucci, E.; Pignatta, S.; Falconi, M.; Silvestrini, R.; Zoli, W.; et al. In Vitro Irradiation System for Radiobiological Experiments. Radiat. Oncol. 2013, 8, 257. [Google Scholar] [CrossRef] [PubMed]
- Thoma, C.R.; Zimmermann, M.; Agarkova, I.; Kelm, J.M.; Krek, W. 3D Cell Culture Systems Modeling Tumor Growth Determinants in Cancer Target Discovery. Adv. Drug Deliv. Rev. 2014, 69–70, 29–41. [Google Scholar] [CrossRef]
- Redden, R.A.; Doolin, E.J. Microgravity Assay of Neuroblastoma: In Vitro Aggregation Kinetics and Organoid Morphology Correlate with MYCN Expression. Vitr. Cell Dev. Biol. Anim. 2011, 47, 312–317. [Google Scholar] [CrossRef]
- Lu, P.; Weaver, V.M.; Werb, Z. The Extracellular Matrix: A Dynamic Niche in Cancer Progression. J. Cell Biol. 2012, 196, 395–406. [Google Scholar] [CrossRef]
- Aljitawi, O.S.; Li, D.; Xiao, Y.; Zhang, D.; Ramachandran, K.; Stehno-Bittel, L.; Van Veldhuizen, P.; Lin, T.L.; Kambhampati, S.; Garimella, R. A Novel Three-Dimensional Stromal-Based Model for In Vitro Chemotherapy Sensitivity Testing of Leukemia Cells. Leuk. Lymphoma 2014, 55, 378–391. [Google Scholar] [CrossRef]
- Innala, M.; Riebe, I.; Kuzmenko, V.; Sundberg, J.; Gatenholm, P.; Hanse, E.; Johannesson, S. 3D Culturing and Differentiation of SH-SY5Y Neuroblastoma Cells on Bacterial Nanocellulose Scaffolds. Artif. Cells Nanomed. Biotechnol. 2014, 42, 302–308. [Google Scholar] [CrossRef] [PubMed]
- Almici, E.; Caballero, D.; Montero, J.; Samitier, J. 3D Neuroblastoma In Vitro Models Using Engineered Cell-Derived Matrices. In Biomaterials for 3D Tumor Modeling; Elsevier: Amsterdam, The Netherlands, 2020; pp. 107–130. ISBN 9780128181287. [Google Scholar]
- Neuhaus, J.; Rabien, A.; Reinhold, A.; Koehler, L.; Berndt-Paetz, M. 3D Tumor Models in Urology. Int. J. Mol. Sci. 2023, 24, 6232. [Google Scholar] [CrossRef]
- Yokota, E.; Iwai, M.; Yukawa, T.; Yoshida, M.; Naomoto, Y.; Haisa, M.; Monobe, Y.; Takigawa, N.; Guo, M.; Maeda, Y.; et al. Clinical Application of a Lung Cancer Organoid (Tumoroid) Culture System. NPJ Precis. Oncol. 2021, 5, 29. [Google Scholar] [CrossRef]
- Lancaster, M.A.; Renner, M.; Martin, C.A.; Wenzel, D.; Bicknell, L.S.; Hurles, M.E.; Homfray, T.; Penninger, J.M.; Jackson, A.P.; Knoblich, J.A. Cerebral Organoids Model Human Brain Development and Microcephaly. Nature 2013, 501, 373–379. [Google Scholar] [CrossRef] [PubMed]
- Lancaster, M.A.; Huch, M. Disease Modelling in Human Organoids. DMM Dis. Model. Mech. 2019, 12, dmm039347. [Google Scholar] [CrossRef]
- Magré, L.; Verstegen, M.M.A.; Buschow, S.; van der Laan, L.J.W.; Peppelenbosch, M.; Desai, J. Emerging Organoid-Immune Co-Culture Models for Cancer Research: From Oncoimmunology to Personalized Immunotherapies. J. Immunother. Cancer 2023, 11, e006290. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Cong, L.; Cong, X. Patient-Derived Organoids in Precision Medicine: Drug Screening, Organoid-on-a-Chip and Living Organoid Biobank. Front. Oncol. 2021, 11, 762184. [Google Scholar] [CrossRef] [PubMed]
- Gallagher, C.; Murphy, C.; Kelly, G.; O’brien, F.J.; Piskareva, O. Three-Dimensional In Vitro Biomimetic Model of Neuroblastoma Using Collagen-Based Scaffolds. J. Vis. Exp. 2021, 2021, e62627. [Google Scholar] [CrossRef]
- Bordoni, M.; Karabulut, E.; Kuzmenko, V.; Fantini, V.; Pansarasa, O.; Cereda, C.; Gatenholm, P. 3D Printed Conductive Nanocellulose Scaffolds for the Differentiation of Human Neuroblastoma Cells. Cells 2020, 9, 682. [Google Scholar] [CrossRef]
- Desai, A.; Kisaalita, W.S.; Keith, C.; Wu, Z.Z. Human Neuroblastoma (SH-SY5Y) Cell Culture and Differentiation in 3-D Collagen Hydrogels for Cell-Based Biosensing. Biosens. Bioelectron. 2006, 21, 1483–1492. [Google Scholar] [CrossRef]
- Fabbri, R.; Cacopardo, L.; Ahluwalia, A.; Magliaro, C. Advanced 3D Models of Human Brain Tissue Using Neural Cell Lines: State-of-the-Art and Future Prospects. Cells 2023, 12, 1181. [Google Scholar] [CrossRef]
- Marrella, A.; Dondero, A.; Aiello, M.; Casu, B.; Olive, D.; Regis, S.; Bottino, C.; Pende, D.; Meazza, R.; Caluori, G.; et al. Cell-Laden Hydrogel as a Clinical-Relevant 3D Model for Analyzing Neuroblastoma Growth, Immunophenotype, and Susceptibility to Therapies. Front. Immunol. 2019, 10, 1876. [Google Scholar] [CrossRef]
- Curtin, C.; Nolan, J.C.; Conlon, R.; Deneweth, L.; Gallagher, C.; Tan, Y.J.; Cavanagh, B.L.; Asraf, A.Z.; Harvey, H.; Miller-Delaney, S.; et al. A Physiologically Relevant 3D Collagen-Based Scaffold–Neuroblastoma Cell System Exhibits Chemosensitivity Similar to Orthotopic Xenograft Models. Acta Biomater. 2018, 70, 84–97. [Google Scholar] [CrossRef]
- Monferrer, E.; Dobre, O.; Trujillo, S.; González Oliva, M.A.; Trubert-Paneli, A.; Acevedo-León, D.; Noguera, R.; Salmeron-Sanchez, M. Vitronectin-Based Hydrogels Recapitulate Neuroblastoma Growth Conditions. Front. Cell Dev. Biol. 2022, 10, 988699. [Google Scholar] [CrossRef]
- Nothdurfter, D.; Ploner, C.; Coraça-Huber, D.C.; Wilflingseder, D.; Müller, T.; Hermann, M.; Hagenbuchner, J.; Ausserlechner, M.J. 3D Bioprinted, Vascularized Neuroblastoma Tumor Environment in Fluidic Chip Devices for Precision Medicine Drug Testing. Biofabrication 2022, 14, 035002. [Google Scholar] [CrossRef] [PubMed]
- Villasante, A.; Sakaguchi, K.; Kim, J.; Cheung, N.K.; Nakayama, M.; Parsa1, H.; Okano, T.; Shimizu, T.; Vunjak-Novakovic, G. Vascularized Tissue-Engineered Model for Studying Drug Resistance in Neuroblastoma. Theranostics 2017, 7, 4099–4117. [Google Scholar] [CrossRef]
- Wan, X.; Wang, W.; Liang, Z. Epigallocatechin-3-Gallate Inhibits the Growth of Three-Dimensional in Vitro Models of Neuroblastoma Cell SH-SY5Y. Mol. Cell Biochem. 2021, 476, 3141–3148. [Google Scholar] [CrossRef] [PubMed]
- Ning, L.; Shim, J.; Tomov, M.L.; Liu, R.; Mehta, R.; Mingee, A.; Hwang, B.; Jin, L.; Mantalaris, A.; Xu, C.; et al. A 3D Bioprinted in Vitro Model of Neuroblastoma Recapitulates Dynamic Tumor-Endothelial Cell Interactions Contributing to Solid Tumor Aggressive Behavior. Adv. Sci. 2022, 9, 2200244. [Google Scholar] [CrossRef]
- Barberio, C.; Withers, A.; Mishra, Y.; Couraud, P.O.; Romero, I.A.; Weksler, B.; Owens, R.M. A Human-Derived Neurovascular Unit In Vitro Model to Study the Effects of Cellular Cross-Talk and Soluble Factors on Barrier Integrity. Front. Cell Neurosci. 2022, 16, 1065193. [Google Scholar] [CrossRef] [PubMed]
- Weng, Z.; Lin, J.; He, J.; Gao, L.; Lin, S.; Tsang, L.L.; Zhang, H.; He, X.; Wang, G.; Yang, X.; et al. Human Embryonic Stem Cell-Derived Neural Crest Model Unveils CD55 as a Cancer Stem Cell Regulator for Therapeutic Targeting in MYCN-Amplified Neuroblastoma. Neuro Oncol. 2022, 24, 872–885. [Google Scholar] [CrossRef] [PubMed]
- Saxena, S.; Wahl, J.; Huber-Lang, M.S.; Stadel, D.; Braubach, P.; Debatin, K.M.; Beltinger, C. Generation of Murine Sympathoadrenergic Progenitor-Like Cells from Embryonic Stem Cells and Postnatal Adrenal Glands. PLoS ONE 2013, 8, e64454. [Google Scholar] [CrossRef]
- Schloo, H.; Kutscher, L.M. Modeling Brain and Neural Crest Neoplasms with Human Pluripotent Stem Cells. Neuro Oncol. 2023. online ahead of print. [Google Scholar] [CrossRef]
- Carr-Wilkinson, J.; Prathalingam, N.; Pal, D.; Moad, M.; Lee, N.; Sundaresh, A.; Forgham, H.; James, P.; Herbert, M.; Lako, M.; et al. Differentiation of Human Embryonic Stem Cells to Sympathetic Neurons: A Potential Model for Understanding Neuroblastoma Pathogenesis. Stem Cells Int. 2018, 2018, 4391641. [Google Scholar] [CrossRef]
- Kramer, M.; Ribeiro, D.; Arsenian-Henriksson, M.; Deller, T.; Rohrer, H. Proliferation and Survival of Embryonic Sympathetic Neuroblasts by MYCN and Activated ALK Signaling. J. Neurosci. 2016, 36, 10425–10439. [Google Scholar] [CrossRef]
- Huang, M.; Weiss, W.A. Neuroblastoma and MYCN. Cold Spring Harb. Perspect. Med. 2013, 3, a014415. [Google Scholar] [CrossRef]
- Weiss, W.A.; Aldape, K.; Mohapatra, G.; Feuerstein, B.G.; Michael Bishop, J. Targeted Expression of MYCN Causes Neuroblastoma in Transgenic Mice. EMBO J. 1997, 16, 2985–2995. [Google Scholar] [CrossRef]
- Teitz, T.; Stanke, J.J.; Federico, S.; Bradley, C.L.; Brennan, R.; Zhang, J.; Johnson, M.D.; Sedlacik, J.; Inoue, M.; Zhang, Z.M.; et al. Preclinical Models for Neuroblastoma: Establishing a Baseline for Treatment. PLoS ONE 2011, 6, e19133. [Google Scholar] [CrossRef]
- Webb, E.R.; Lanati, S.; Wareham, C.; Easton, A.; Dunn, S.N.; Inzhelevskaya, T.; Sadler, F.M.; James, S.; Ashton-Key, M.; Cragg, M.S.; et al. Immune Characterization of Pre-Clinical Murine Models of Neuroblastoma. Sci. Rep. 2020, 10, 16695. [Google Scholar] [CrossRef]
- Hong, S.H.; Wietlisbach, L.; Galli, S.; Mahajan, A.; Zhu, S.; Tilan, J.; Lee, Y.; Rodriguez, O.; Albanese, C.; Kitlinska, J. Abstract 1940: Prenatal Stress Increases Malignancy of Neuroblastoma Tumors in TH-MYCN Animal Model. Cancer Res. 2017, 77, 1940. [Google Scholar] [CrossRef]
- Faisal, A.; Vaughan, L.; Bavetsias, V.; Sun, C.; Atrash, B.; Avery, S.; Jamin, Y.; Robinson, S.P.; Workman, P.; Blagg, J.; et al. The Aurora Kinase Inhibitor CCT137690 Downregulates MYCN and Sensitizes MYCN-Amplified Neuroblastoma In Vivo. Mol. Cancer Ther. 2011, 10, 2115–2123. [Google Scholar] [CrossRef]
- Morowitz, M.J.; Barr, R.; Wang, Q.; King, R.; Rhodin, N.; Pawel, B.; Zhao, H.; Erickson, S.A.; Sheppard, G.S.; Wang, J.; et al. Methionine Aminopeptidase 2 Inhibition Is an EffectiveTreatment Strategy for Neuroblastoma in Preclinical Models. Clin. Cancer Res. 2005, 11, 2680–2685. [Google Scholar] [CrossRef]
- Yogev, O.; Almeida, G.S.; Barker, K.T.; George, S.L.; Kwok, C.; Campbell, J.; Zarowiecki, M.; Kleftogiannis, D.; Smith, L.M.; Hallsworth, A.; et al. In Vivo Modeling of Chemoresistant Neuroblastoma Provides New Insights into Chemorefractory Disease and Metastasis. Cancer Res. 2019, 79, 5382–5393. [Google Scholar] [CrossRef] [PubMed]
- Chesler, L.; Weiss, W.A. Genetically Engineered Murine Models-Contribution to Our Understanding of the Genetics, Molecular Pathology and Therapeutic Targeting of Neuroblastoma. Semin. Cancer Biol. 2011, 21, 245–255. [Google Scholar] [CrossRef] [PubMed]
- Nijhuis, A.; Sikka, A.; Yogev, O.; Herendi, L.; Balcells, C.; Ma, Y.; Poon, E.; Eckold, C.; Valbuena, G.N.; Xu, Y.; et al. Indisulam Targets RNA Splicing and Metabolism to Serve as a Therapeutic Strategy for High-Risk Neuroblastoma. Nat. Commun. 2022, 13, 1380. [Google Scholar] [CrossRef] [PubMed]
- Kamili, A.; Atkinson, C.; Trahair, T.N.; Fletcher, J.I. Mouse Models of High-Risk Neuroblastoma. Cancer Metastasis Rev. 2020, 39, 261–274. [Google Scholar] [CrossRef]
- Yogev, O.; Barker, K.; Sikka, A.; Almeida, G.S.; Hallsworth, A.; Smith, L.M.; Jamin, Y.; Ruddle, R.; Koers, A.; Twebber, H.; et al. P53 Loss in MYC-Driven Neuroblastoma Leads to Metabolic Adaptations Supporting Radioresistance. Cancer Res. 2016, 76, 3025–3035. [Google Scholar] [CrossRef] [PubMed]
- Kim, P.Y.; Tan, O.; Diakiw, S.M.; Carter, D.; Sekerye, E.O.; Wasinger, V.C.; Liu, T.; Kavallaris, M.; Norris, M.D.; Haber, M.; et al. Identification of Plasma Complement C3 as a Potential Biomarker for Neuroblastoma Using a Quantitative Proteomic Approach. J. Proteom. 2014, 96, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Shawraba, F.; Hammoud, H.; Mrad, Y.; Saker, Z.; Fares, Y.; Harati, H.; Bahmad, H.F.; Nabha, S. Biomarkers in Neuroblastoma: An Insight into Their Potential Diagnostic and Prognostic Utilities. Curr. Treat Options Oncol. 2021, 22, 102. [Google Scholar] [CrossRef]
- Teitz, T.; Inoue, M.; Valentine, M.B.; Zhu, K.; Rehg, J.E.; Zhao, W.; Finkelstein, D.; Wang, Y.D.; Johnson, M.D.; Calabrese, C.; et al. Th-MYCN Mice with Caspase-8 Deficiency Develop Advanced Neuroblastoma with Bone Marrow Metastasis. Cancer Res. 2013, 73, 4086–4097. [Google Scholar] [CrossRef] [PubMed]
- Althoff, K.; Schramm, A. MYCN-Mediated Murine Cancer Models. Aging 2017, 9, 1084–1085. [Google Scholar] [CrossRef]
- Althoff, K.; Beckers, A.; Bell, E.; Nortmeyer, M.; Thor, T.; Sprüssel, A.; Lindner, S.; De Preter, K.; Florin, A.; Heukamp, L.C.; et al. A Cre-Conditional MYCN-Driven Neuroblastoma Mouse Model as an Improved Tool for Preclinical Studies. Oncogene 2015, 34, 3357–3368. [Google Scholar] [CrossRef]
- Chen, Z.; Lin, Y.; Barbieri, E.; Burlingame, S.; Hicks, J.; Ludwig, A.; Shohet, J.M. Mdm2 Deficiency Suppresses MYCN-Driven Neuroblastoma Tumorigenesis In Vivo. Neoplasia 2009, 11, 753–762. [Google Scholar] [CrossRef]
- Galli, S.; Naranjo, A.; Van Ryn, C.; Tilan, J.U.; Trinh, E.; Yang, C.; Tsuei, J.; Hong, S.H.; Wang, H.; Izycka-Swieszewska, E.; et al. Neuropeptide Y as a Biomarker and Therapeutic Target for Neuroblastoma. Am. J. Pathol. 2016, 186, 3040–3053. [Google Scholar] [CrossRef] [PubMed]
- Schleiermacher, G.; Janoueix-Lerosey, I.; Delattre, O. Recent Insights into the Biology of Neuroblastoma. Int. J. Cancer 2014, 135, 2249–2261. [Google Scholar] [CrossRef]
- Kiyonari, S.; Kadomatsu, K. Neuroblastoma Models for Insights into Tumorigenesis and New Therapies. Expert Opin. Drug Discov. 2015, 10, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Heukamp, L.C.; Thor, T.; Schramm, A.; De Preter, K.; Kumps, C.; De Wilde, B.; Odersky, A.; Peifer, M.; Lindner, S.; Spruessel, A.; et al. Targeted Expression of Mutated ALK Induces Neuroblastoma in Transgenic Mice. Sci. Transl. Med. 2012, 4, 141ra91. [Google Scholar] [CrossRef] [PubMed]
- Molenaar, J.J.; Domingo-Fernández, R.; Ebus, M.E.; Lindner, S.; Koster, J.; Drabek, K.; Mestdagh, P.; Van Sluis, P.; Valentijn, L.J.; Van Nes, J.; et al. LIN28B Induces Neuroblastoma and Enhances MYCN Levels via Let-7 Suppression. Nat. Genet. 2012, 44, 1199–1206. [Google Scholar] [CrossRef]
- Berry, T.; Luther, W.; Bhatnagar, N.; Jamin, Y.; Poon, E.; Sanda, T.; Pei, D.; Sharma, B.; Vetharoy, W.R.; Hallsworth, A.; et al. The ALKF1174L Mutation Potentiates the Oncogenic Activity of MYCN in Neuroblastoma. Cancer Cell 2012, 22, 117–130. [Google Scholar] [CrossRef] [PubMed]
- Ono, S.; Saito, T.; Terui, K.; Yoshida, H.; Enomoto, H. Generation of Conditional ALK F1174L Mutant Mouse Models for the Study of Neuroblastoma Pathogenesis. Genesis 2019, 57, e23323. [Google Scholar] [CrossRef]
- Tao, T.; Shi, H.; Mariani, L.; Abraham, B.J.; Durbin, A.D.; Zimmerman, M.W.; Powers, J.T.; Missios, P.; Ross, K.N.; Perez-Atayde, A.R.; et al. LIN28B Regulates Transcription and Potentiates MYCN-Induced Neuroblastoma through Binding to ZNF143 at Target Gene Promotors. Proc. Natl. Acad. Sci. USA 2020, 117, 16516–16526. [Google Scholar] [CrossRef]
- De Wilde, B.; Beckers, A.; Lindner, S.; Kristina, A.; De Preter, K.; Depuydt, P.; Mestdagh, P.; Sante, T.; Lefever, S.; Hertwig, F.; et al. The Mutational Landscape of MYCN, Lin28b and ALK F1174L Driven Murine Neuroblastoma Mimics Human Disease. Oncotarget 2018, 9, 8334. [Google Scholar] [CrossRef]
- Singh, S.; Quarni, W.; Goralski, M.; Wan, S.; Jin, H.; Van de Velde, L.-A.; Fang, J.; Wu, Q.; Abu-Zaid, A.; Wang, T.; et al. Targeting the Spliceosome through RBM39 Degradation Results in Exceptional Responses in High-Risk Neuroblastoma Models. Sci. Adv. 2021, 7, eabj5405. [Google Scholar] [CrossRef]
- Iwakura, H.; Akamizu, T. Neuroblastoma Mouse Model; Hayat, M.A., Ed.; Pediatric Cancer; Springer: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- Koike, K.; Jay, G.; Hartley, J.W.; Schrenzel, M.D.; Higgins, R.J.; Hinrichs, S.H. Activation of Retrovirus in Transgenic Mice: Association with Development of Olfactory Neuroblastoma. J. Virol. 1990, 64, 3988–3991. [Google Scholar] [CrossRef]
- Small, J.A.; Khoury, G.; Jayt, G.; Howley, P.M.; Scangos, G.A. Early Regions of JC Virus and BK Virus Induce Distinct and Tissue-Specific Tumors in Transgenic Mice. Proc. Natl. Acad. Sci. USA 1986, 83, 8288–8292. [Google Scholar] [CrossRef]
- Iwamoto, T.; Taniguchi, M.; Wajjwalku, W.; Nakashimal, I.; Takahashi, M. Neuroblastoma in a Transgenic Mouse Carrying a Metallothionein/Ret Fusion Gene. Br. J. Cancer 1993, 67, 504. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, C.; Sakata, S.; Kakinoki, S.; Takeyama, Y.; Ohyanagi, H.; Shiozaki, H. In Vivo Evaluation of Microspheres Containing the Angiogenesis Inhibitor, TNP-470, and the Metastasis Suppression with Liver Metastatic Model Implanted Neuroblastoma. Pathophysiology 2010, 17, 149–155. [Google Scholar] [CrossRef]
- Beltinger, C.; Debatin, K.M. Murine Models for Experimental Therapy of Pediatric Solid Tumors with Poor Prognosis. Int. J. Cancer 2001, 92, 313–318. [Google Scholar] [CrossRef]
- Khanna, C.; Jaboin, J.J.; Drakos, E.; Tsokos, M.; Thiele, C.J. Biologically Relevant Orthotopic Neuroblastoma Xenograft Models: Primary Adrenal Tumor Growth and Spontaneous Distant Metastasis. In Vivo 2002, 16, 77–86. [Google Scholar]
- Shultz, L.D.; Goodwin, N.; Ishikawa, F.; Hosur, V.; Lyons, B.L.; Greiner, D.L. Human Cancer Growth and Therapy in Immunodeficient Mouse Models. Cold Spring Harb. Protoc. 2014, 2014, 694–708. [Google Scholar] [CrossRef]
- Kaneko, Y.; Tsuchida, Y.; Maseki, N.; Takasaki, N.; Sakurai, M.; Saito, S. Chromosome findings in human neuroblastomas xenografted in nude mice. J. Cancer Res. GANN 1985, 76, 359–364. [Google Scholar]
- Hidalgo, M.; Bruckheimer, E.; Rajeshkumar, N.V.; Garrido-Laguna, I.; De Oliveira, E.; Rubio-Viqueira, B.; Strawn, S.; Wick, M.J.; Martell, J.; Sidransky, D. A Pilot Clinical Study of Treatment Guided by Personalized Tumorgrafts in Patients with Advanced Cancer. Mol. Cancer Ther. 2011, 10, 1311–1316. [Google Scholar] [CrossRef]
- Gao, Q.; Chen, C.F.; Dong, Q.; Hou, L.; Chen, X.; Zhi, Y.L.; Li, X.; Lu, H.T.; Zhang, H.Y. Establishment of a Neuroblastoma Mouse Model by Subcutaneous Xenograft Transplantation and Its Use to Study Metastatic Neuroblastoma. Genet. Mol. Res. 2015, 14, 16297–16307. [Google Scholar] [CrossRef]
- Braekeveldt, N.; Wigerup, C.; Gisselsson, D.; Mohlin, S.; Merselius, M.; Beckman, S.; Jonson, T.; Börjesson, A.; Backman, T.; Tadeo, I.; et al. Neuroblastoma Patient-Derived Orthotopic Xenografts Retain Metastatic Patterns and Geno-and Phenotypes of Patient Tumours. Int. J. Cancer 2015, 136, E252–E261. [Google Scholar] [CrossRef]
- Braekeveldt, N.; Bexell, D. Patient-Derived Xenografts as Preclinical Neuroblastoma Models. Cell Tissue Res. 2018, 372, 233–243. [Google Scholar] [CrossRef] [PubMed]
- Grant, C.N.; Wills, C.A.; Liu, X.; Spiegelman, V.S.; Wang, H.G. Thoracic Neuroblastoma: A Novel Surgical Model for the Study of Extra-Adrenal Neuroblastoma. In Vivo 2022, 36, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Stewart, E.; Shelat, A.; Bradley, C.; Chen, X.; Federico, S.; Thiagarajan, S.; Shirinifard, A.; Bahrami, A.; Pappo, A.; Qu, C.; et al. Development and Characterization of a Human Orthotopic Neuroblastoma Xenograft. Dev. Biol. 2015, 407, 344–355. [Google Scholar] [CrossRef]
- Braekeveldt, N.; Wigerup, C.; Tadeo, I.; Beckman, S.; Sandén, C.; Jönsson, J.; Erjefält, J.S.; Berbegall, A.P.; Börjesson, A.; Backman, T.; et al. Neuroblastoma Patient-Derived Orthotopic Xenografts Reflect the Microenvironmental Hallmarks of Aggressive Patient Tumours. Cancer Lett. 2016, 375, 384–389. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Cao, Y.; Okada, R.; Reyes-González, J.M.; Stack, H.G.; Qin, H.; Li, N.; Seibert, C.; Kelly, M.C.; Ruppin, E.; et al. Preclinical Optimization of a GPC2-Targeting CAR T-Cell Therapy for Neuroblastoma. J. Immunother. Cancer 2023, 11, e005881. [Google Scholar] [CrossRef]
- Quinn, C.H.; Beierle, A.M.; Hutchins, S.C.; Marayati, R.; Bownes, L.V.; Stewart, J.E.; Markert, H.R.; Erwin, M.H.; Aye, J.M.; Yoon, K.J.; et al. Targeting High-Risk Neuroblastoma Patient-Derived Xenografts with Oncolytic Virotherapy. Cancers 2022, 14, 762. [Google Scholar] [CrossRef]
- Labitzky, V.; Baranowsky, A.; Maar, H.; Hanika, S.; Starzonek, S.; Ahlers, A.K.; Stübke, K.; Koziolek, E.J.; Heine, M.; Schäfer, P.; et al. Modeling Spontaneous Bone Metastasis Formation of Solid Human Tumor Xenografts in Mice. Cancers 2020, 12, 385. [Google Scholar] [CrossRef]
- Rokita, J.L.; Rathi, K.S.; Cardenas, M.F.; Upton, K.A.; Jayaseelan, J.; Cross, K.L.; Pfeil, J.; Egolf, L.E.; Way, G.P.; Farrel, A.; et al. Genomic Profiling of Childhood Tumor Patient-Derived Xenograft Models to Enable Rational Clinical Trial Design. Cell Rep. 2019, 29, 1675–1689.e9. [Google Scholar] [CrossRef] [PubMed]
- Morton, J.J.; Bird, G.; Keysar, S.B.; Astling, D.P.; Lyons, T.R.; Anderson, R.T.; Glogowska, M.J.; Estes, P.; Eagles, J.R.; Le, P.N.; et al. XactMice: Humanizing Mouse Bone Marrow Enables Microenvironment Reconstitution in a Patient-Derived Xenograft Model of Head and Neck Cancer. Oncogene 2016, 35, 290–300. [Google Scholar] [CrossRef]
- Cohen, M.A.; Zhang, S.; Sengupta, S.; Ma, H.; Bell, G.W.; Horton, B.; Sharma, B.; George, R.E.; Spranger, S.; Jaenisch, R. Formation of Human Neuroblastoma in Mouse-Human Neural Crest Chimeras. Cell Stem Cell 2020, 26, 579–592.e6. [Google Scholar] [CrossRef]
- Thole, T.M.; Toedling, J.; Sprüssel, A.; Pfeil, S.; Savelyeva, L.; Capper, D.; Messerschmidt, C.; Beule, D.; Groeneveld-Krentz, S.; Eckert, C.; et al. Reflection of Neuroblastoma Intratumor Heterogeneity in the New OHC-NB1 Disease Model. Int. J. Cancer 2020, 146, 1031–1041. [Google Scholar] [CrossRef]
- Li, S.; Yeo, K.S.; Levee, T.M.; Howe, C.J.; Her, Z.P.; Zhu, S. Zebrafish as a Neuroblastoma Model: Progress Made, Promise for the Future. Cells 2021, 10, 580. [Google Scholar] [CrossRef]
- Martinez, D.A.; Kahwash, S.; O’Dorisio, M.S.; Lloyd, T.V.; McGhee, R.B.; Qualman, S.J. Disseminated Neuroblastoma in the Nude Rat: A Xenograft Model of Human Malignancy. Cancer 1996, 77, 409–419. [Google Scholar] [CrossRef]
- Medina-Cuadra, L.; Monsoro-Burq, A.H. Xenopus, an Emerging Model for Studying Pathologies of the Neural Crest. In Current Topics in Developmental Biology; Academic Press Inc.: Cambridge, MA, USA, 2021; Volume 145, pp. 313–348. [Google Scholar]
- Gonzalez Malagon, S.G.; Lopez Muñoz, A.M.; Doro, D.; Bolger, T.G.; Poon, E.; Tucker, E.R.; Adel Al-Lami, H.; Krause, M.; Phiel, C.J.; Chesler, L.; et al. Glycogen Synthase Kinase 3 Controls Migration of the Neural Crest Lineage in Mouse and Xenopus. Nat. Commun. 2018, 9, 1126. [Google Scholar] [CrossRef]
- Corallo, D.; Donadon, M.; Pantile, M.; Sidarovich, V.; Cocchi, S.; Ori, M.; De Sarlo, M.; Candiani, S.; Frasson, C.; Distel, M.; et al. LIN28B Increases Neural Crest Cell Migration and Leads to Transformation of Trunk Sympathoadrenal Precursors. Cell Death Differ. 2020, 27, 1225–1242. [Google Scholar] [CrossRef]
- Zhu, S.; Thomas Look, A. Neuroblastoma and Its Zebrafish Model. In Advances in Experimental Medicine and Biology; Springer: New York, NY, USA, 2016; Volume 916, pp. 451–478. [Google Scholar]
- Her, Z.P.; Yeo, K.S.; Howe, C.; Levee, T.; Zhu, S. Zebrafish Model of Neuroblastoma Metastasis. J. Vis. Exp. 2021, 2021, e62416. [Google Scholar] [CrossRef]
- Sarmiento, B.E.; Callegari, S.; Ghotme, K.A.; Akle, V. Patient-Derived Xenotransplant of CNS Neoplasms in Zebrafish: A Systematic Review. Cells 2022, 11, 1204. [Google Scholar] [CrossRef]
- Delloye-Bourgeois, C.; Bertin, L.; Thoinet, K.; Jarrosson, L.; Kindbeiter, K.; Buffet, T.; Tauszig-Delamasure, S.; Bozon, M.; Marabelle, A.; Combaret, V.; et al. Microenvironment-Driven Shift of Cohesion/Detachment Balance within Tumors Induces a Switch toward Metastasis in Neuroblastoma. Cancer Cell 2017, 32, 427–443.e8. [Google Scholar] [CrossRef]
- Ben Amar, D.; Thoinet, K.; Villalard, B.; Imbaud, O.; Costechareyre, C.; Jarrosson, L.; Reynaud, F.; Novion Ducassou, J.; Couté, Y.; Brunet, J.F.; et al. Environmental Cues from Neural Crest Derivatives Act as Metastatic Triggers in an Embryonic Neuroblastoma Model. Nat. Commun. 2022, 13, e16629. [Google Scholar] [CrossRef] [PubMed]
- Swadi, R.; Mather, G.; Pizer, B.L.; Losty, P.D.; See, V.; Moss, D. Optimising the Chick Chorioallantoic Membrane Xenograft Model of Neuroblastoma for Drug Delivery. BMC Cancer 2018, 18, 28. [Google Scholar] [CrossRef] [PubMed]
- Ribatti, D.; Tamma, R. The Chick Embryo Chorioallantoic Membrane as an In Vivo Experimental Model to Study Human Neuroblastoma. J. Cell Physiol. 2018, 234, 152–157. [Google Scholar] [CrossRef]
- Herrmann, A.; Rice, M.; Lévy, R.; Pizer, B.L.; Losty, P.D.; Moss, D.; Sée, V. Cellular Memory of Hypoxia Elicits Neuroblastoma Metastasis and Enables Invasion by Non-Aggressive Neighbouring Cells. Oncogenesis 2015, 4, e138. [Google Scholar] [CrossRef] [PubMed]
- Bogenmann, E. A Metastatic Neuroblastoma Model in SCID Mice. Int. J. Cancer 1996, 67, 379–385. [Google Scholar] [CrossRef]
- Chen, D.; Cox, J.; Annam, J.; Weingart, M.; Essien, G.; Rathi, K.S.; Rokita, J.L.; Khurana, P.; Cuya, S.M.; Bosse, K.R.; et al. LIN28B Promotes Neuroblastoma Metastasis and Regulates PDZ Binding Kinase. Neoplasia 2020, 22, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Seong, B.K.A.; Fathers, K.E.; Hallett, R.; Yung, C.K.; Stein, L.D.; Mouaaz, S.; Kee, L.; Hawkins, C.E.; Irwin, M.S.; Kaplan, D.R. A Metastatic Mouse Model Identifies Genes That Regulate Neuroblastoma Metastasis. Cancer Res. 2017, 77, 696–706. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.L.; Chan, S.; Fung, M.K.L.; Chan, G.C.F. Mesenchymal Stem Cells Accelerated Growth and Metastasis of Neuroblastoma and Preferentially Homed towards Both Primary and Metastatic Loci in Orthotopic Neuroblastoma Model. BMC Cancer 2021, 21, 393. [Google Scholar] [CrossRef] [PubMed]
- Segura, M.F.; Soriano, A.; Roma, J.; Piskareva, O.; Jiménez, C.; Boloix, A.; Fletcher, J.I.; Haber, M.; Gray, J.C.; Cerdá-Alberich, L.; et al. Methodological Advances in the Discovery of Novel Neuroblastoma Therapeutics. Expert Opin. Drug Discov. 2022, 17, 167–179. [Google Scholar] [CrossRef]
- Chilamakuri, R.; Rouse, D.C.; Agarwal, S. Inhibition of Polo-like Kinase 1 by HMN-214 Blocks Cell Cycle Progression and Inhibits Neuroblastoma Growth. Pharmaceuticals 2022, 15, 523. [Google Scholar] [CrossRef]
- Marzagalli, M.; Pelizzoni, G.; Fedi, A.; Vitale, C.; Fontana, F.; Bruno, S.; Poggi, A.; Dondero, A.; Aiello, M.; Castriconi, R.; et al. A Multi-Organ-on-Chip to Recapitulate the Infiltration and the Cytotoxic Activity of Circulating NK Cells in 3D Matrix-Based Tumor Model. Front. Bioeng. Biotechnol. 2022, 10, 945149. [Google Scholar] [CrossRef]
- Heinze, A.; Grebe, B.; Bremm, M.; Huenecke, S.; Munir, T.A.; Graafen, L.; Frueh, J.T.; Merker, M.; Rettinger, E.; Soerensen, J.; et al. The Synergistic Use of IL-15 and IL-21 for the Generation of NK Cells From CD3/CD19-Depleted Grafts Improves Their Ex Vivo Expansion and Cytotoxic Potential Against Neuroblastoma: Perspective for Optimized Immunotherapy Post Haploidentical Stem Cell Transplantation. Front. Immunol. 2019, 10, 2816. [Google Scholar] [CrossRef]
- Logan, J.A.; Kelly, M.E.; Ayers, D.; Shipillis, N.; Baier, G.; Day, P.J.R. Systems Biology and Modeling in Neuroblastoma: Practicalities and Perspectives. Expert Rev. Mol. Diagn. 2010, 10, 131–145. [Google Scholar] [CrossRef] [PubMed]
- Yan, Z.; Liu, Q.; Cao, Z.; Wang, J.; Zhang, H.; Liu, J.; Zou, L. Multi-Omics Integration Reveals a Six-Malignant Cell Maker Gene Signature for Predicting Prognosis in High-Risk Neuroblastoma. Front. Neuroinform. 2022, 16, 1034793. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krawczyk, E.; Kitlińska, J. Preclinical Models of Neuroblastoma—Current Status and Perspectives. Cancers 2023, 15, 3314. https://doi.org/10.3390/cancers15133314
Krawczyk E, Kitlińska J. Preclinical Models of Neuroblastoma—Current Status and Perspectives. Cancers. 2023; 15(13):3314. https://doi.org/10.3390/cancers15133314
Chicago/Turabian StyleKrawczyk, Ewa, and Joanna Kitlińska. 2023. "Preclinical Models of Neuroblastoma—Current Status and Perspectives" Cancers 15, no. 13: 3314. https://doi.org/10.3390/cancers15133314
APA StyleKrawczyk, E., & Kitlińska, J. (2023). Preclinical Models of Neuroblastoma—Current Status and Perspectives. Cancers, 15(13), 3314. https://doi.org/10.3390/cancers15133314