
Development and Validation of Blood-Based Predictive Biomarkers for Response to PD-1/PD-L1 Checkpoint Inhibitors: Evidence of a Universal Systemic Core of 3D Immunogenetic Profiling across Multiple Oncological Indications
Abstract
Simple Summary
Abstract
1. Background
2. Materials and Methods
2.1. Patient Characteristics
2.2. Preparation of 3D Genomic Templates
2.3. Array Design
2.4. Translation of Array-Based 3D Genomic Markers to PCR Readouts
2.5. EpiSwitch® PCR
2.6. Genomic Mapping
2.7. Statistical Analysis
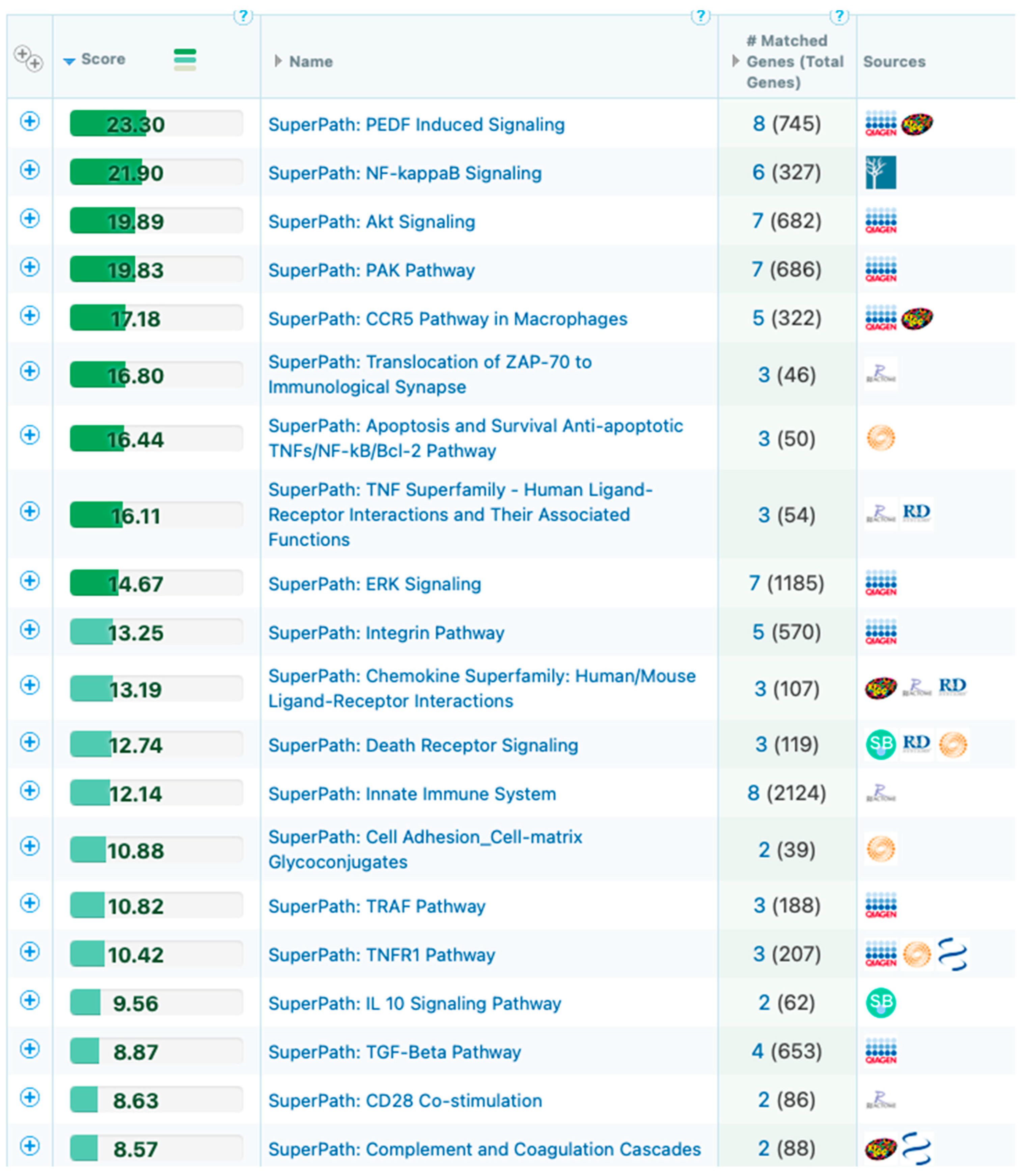
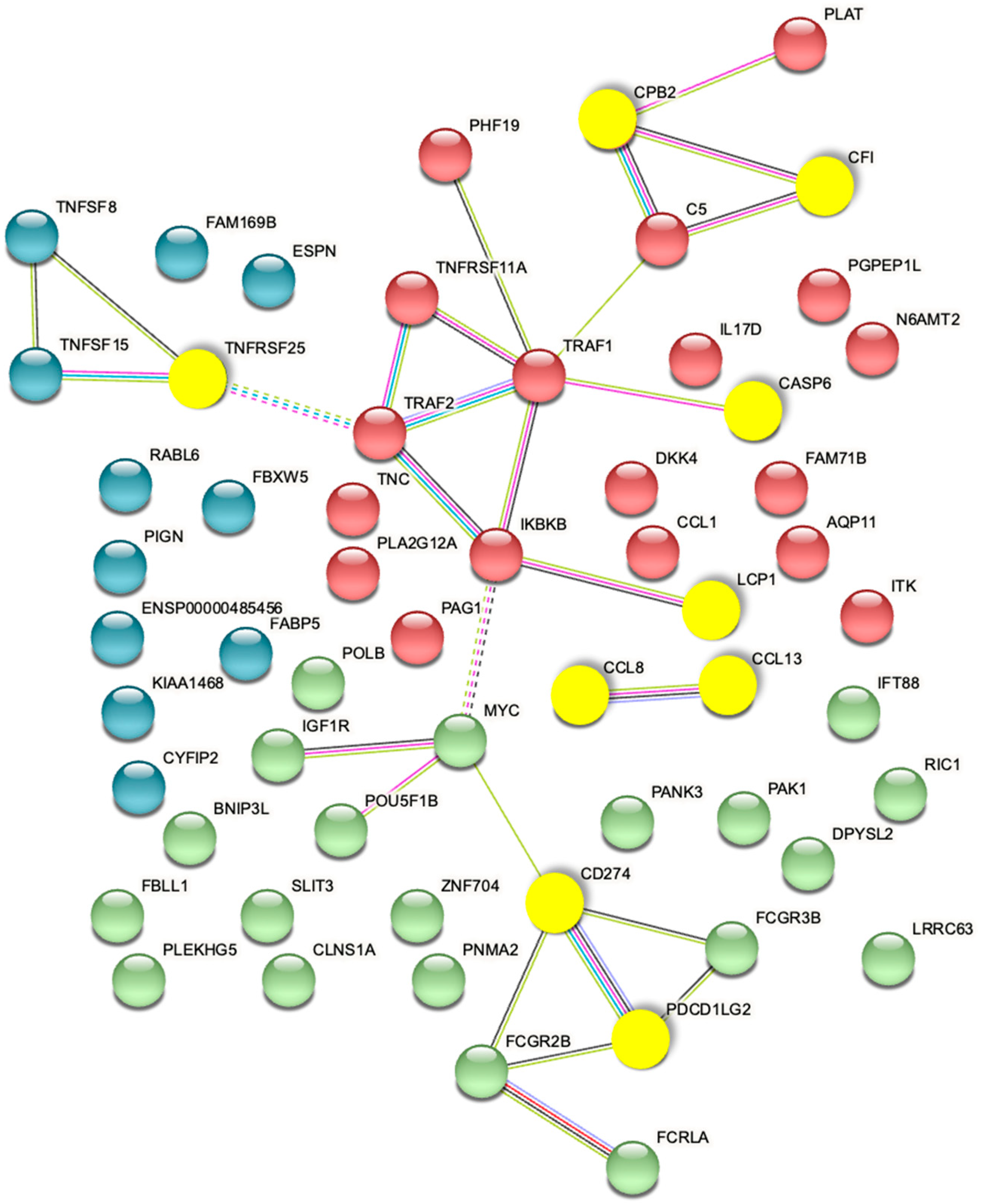
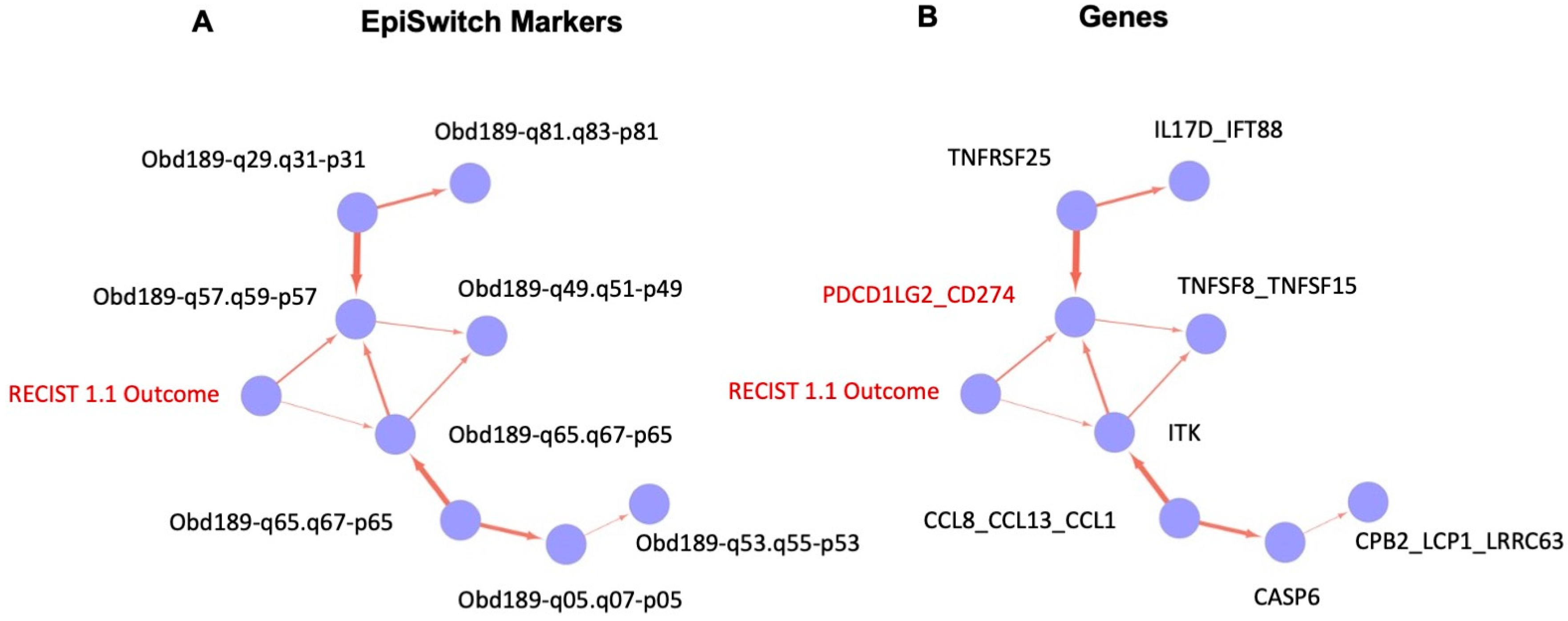
2.8. Biological Network/Pathway Analysis
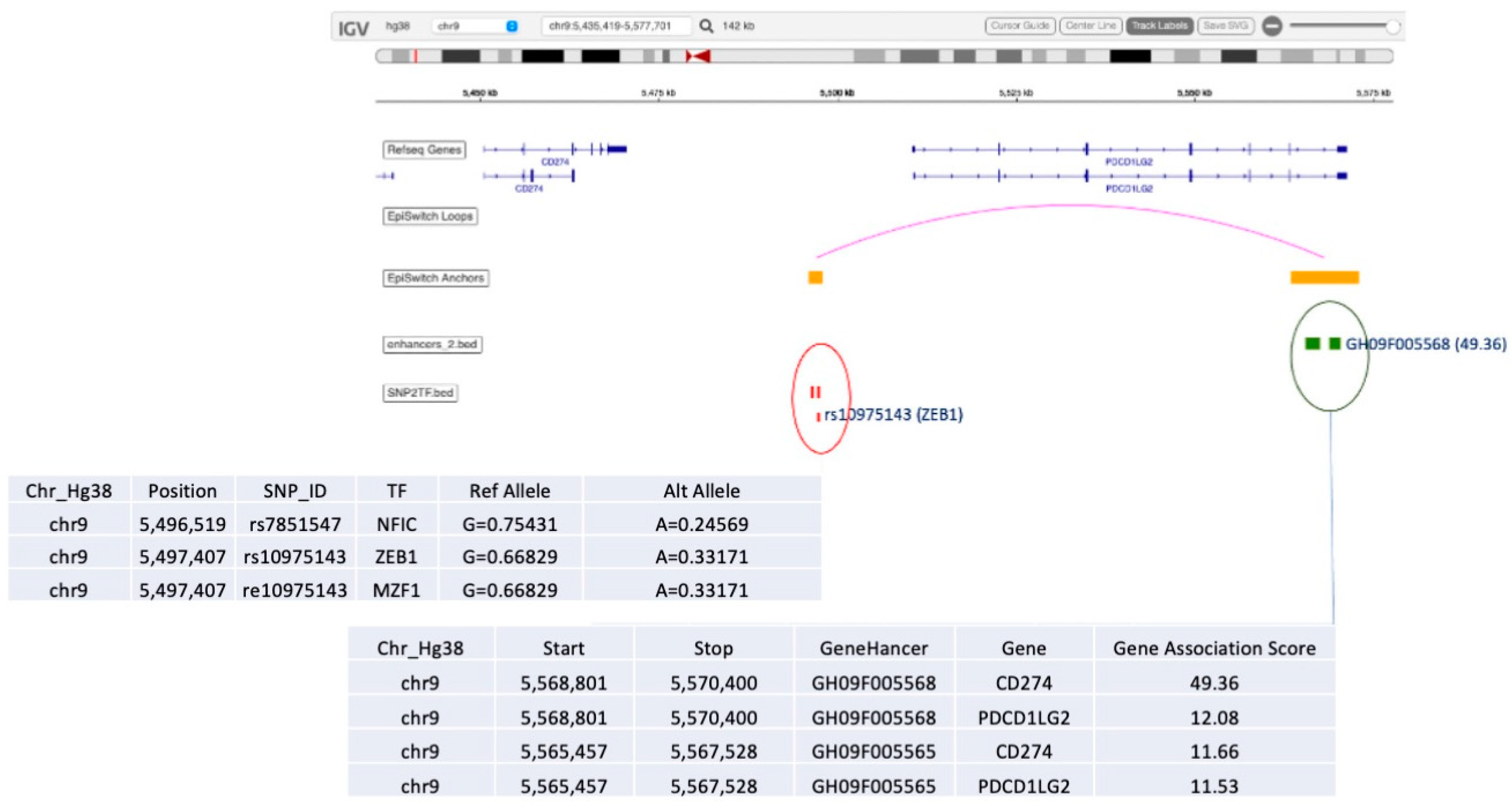
2.9. Causal Graph Analysis
3. Results
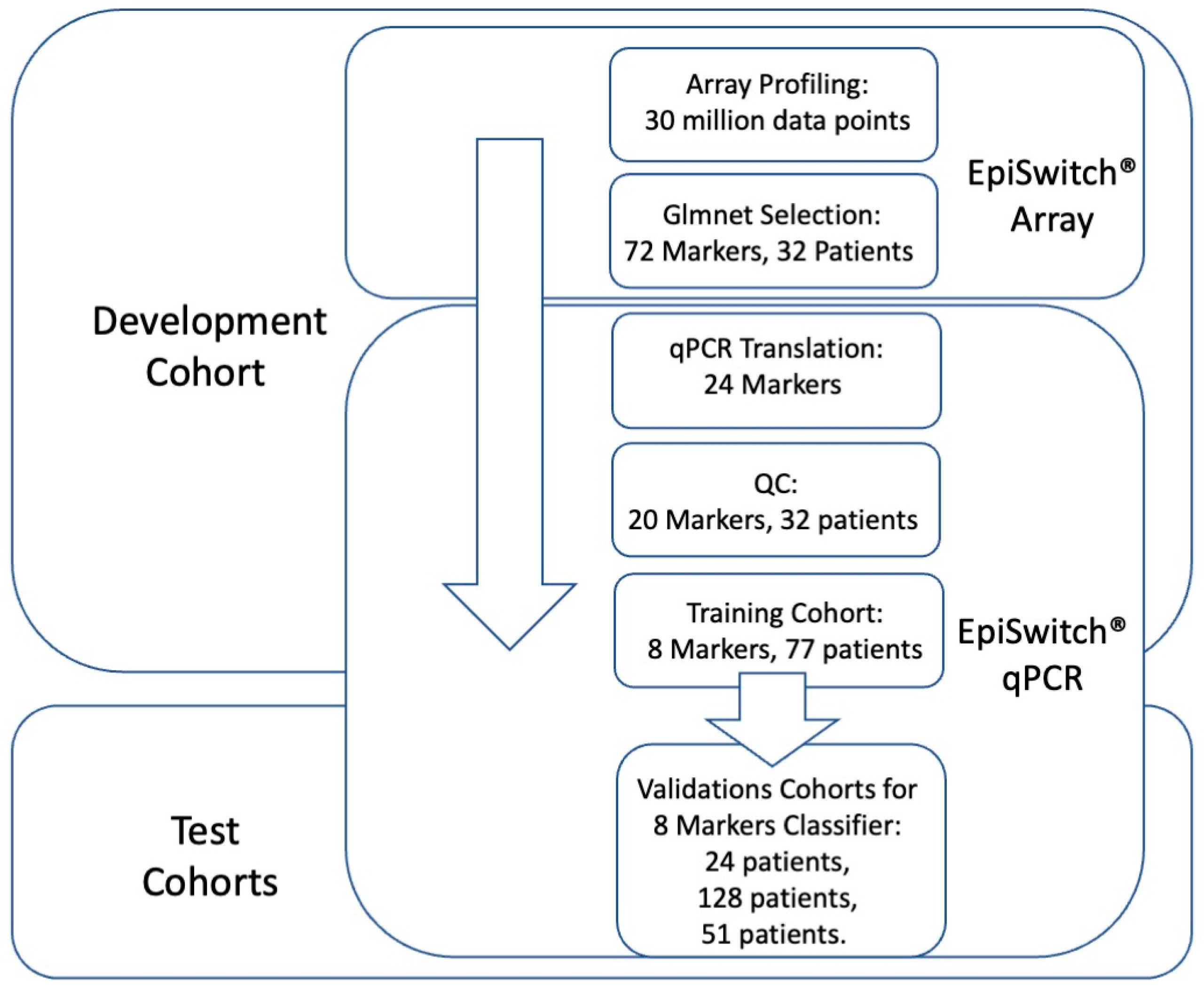
3.1. Whole Genome Array Profiling for Discovery of Predictive 3D Genomic Marker Leads in Baseline Immuno-Oncology (IO) Patients at Baseline
3.2. Identification of the Top Predictive 3D Genomic Markers for ICI Treatment Outcomes
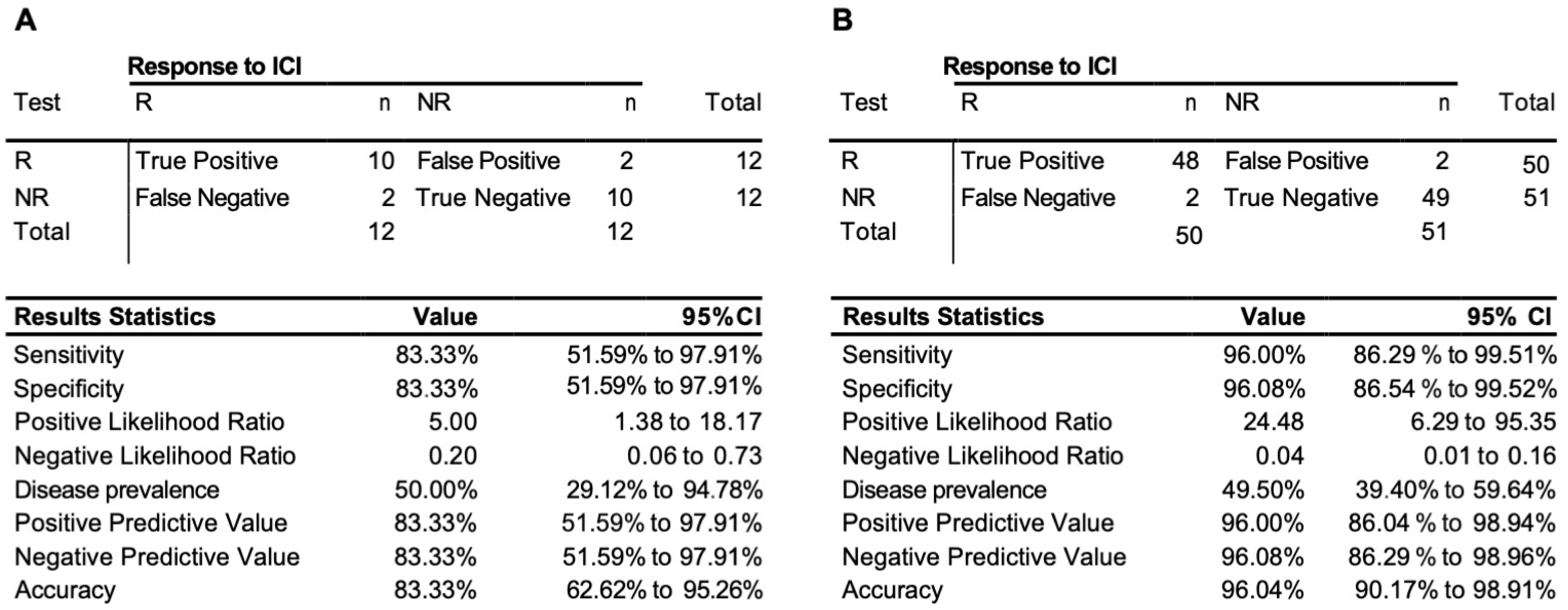
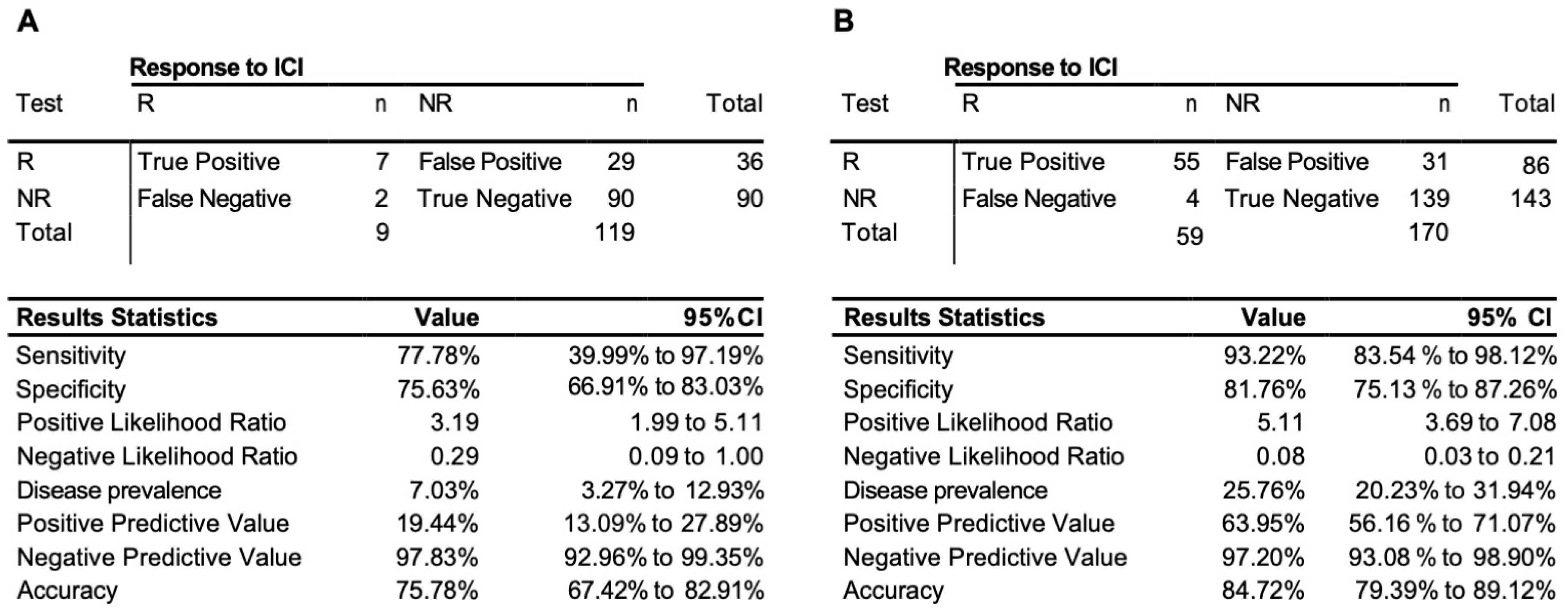
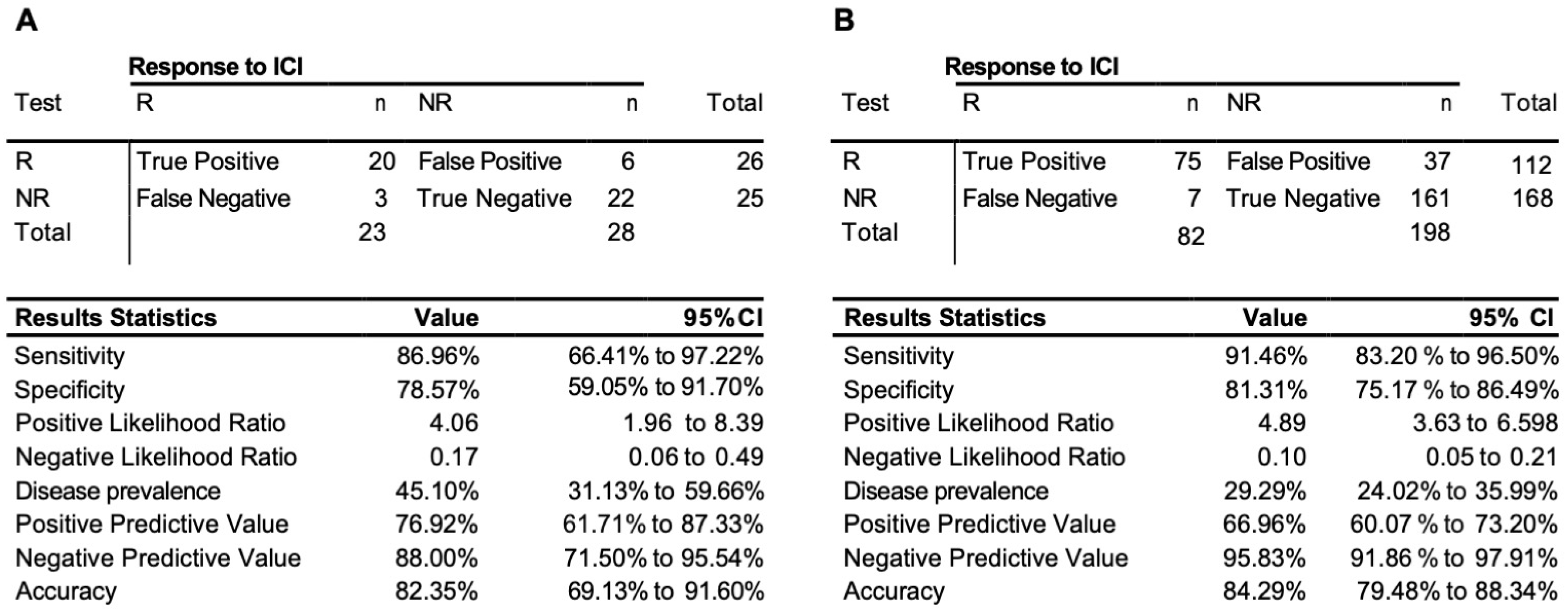
3.3. Testing of the Predictive 3D Genomic Biomarker Panel for Response to ICI Treatments on Independent Patient Cohorts
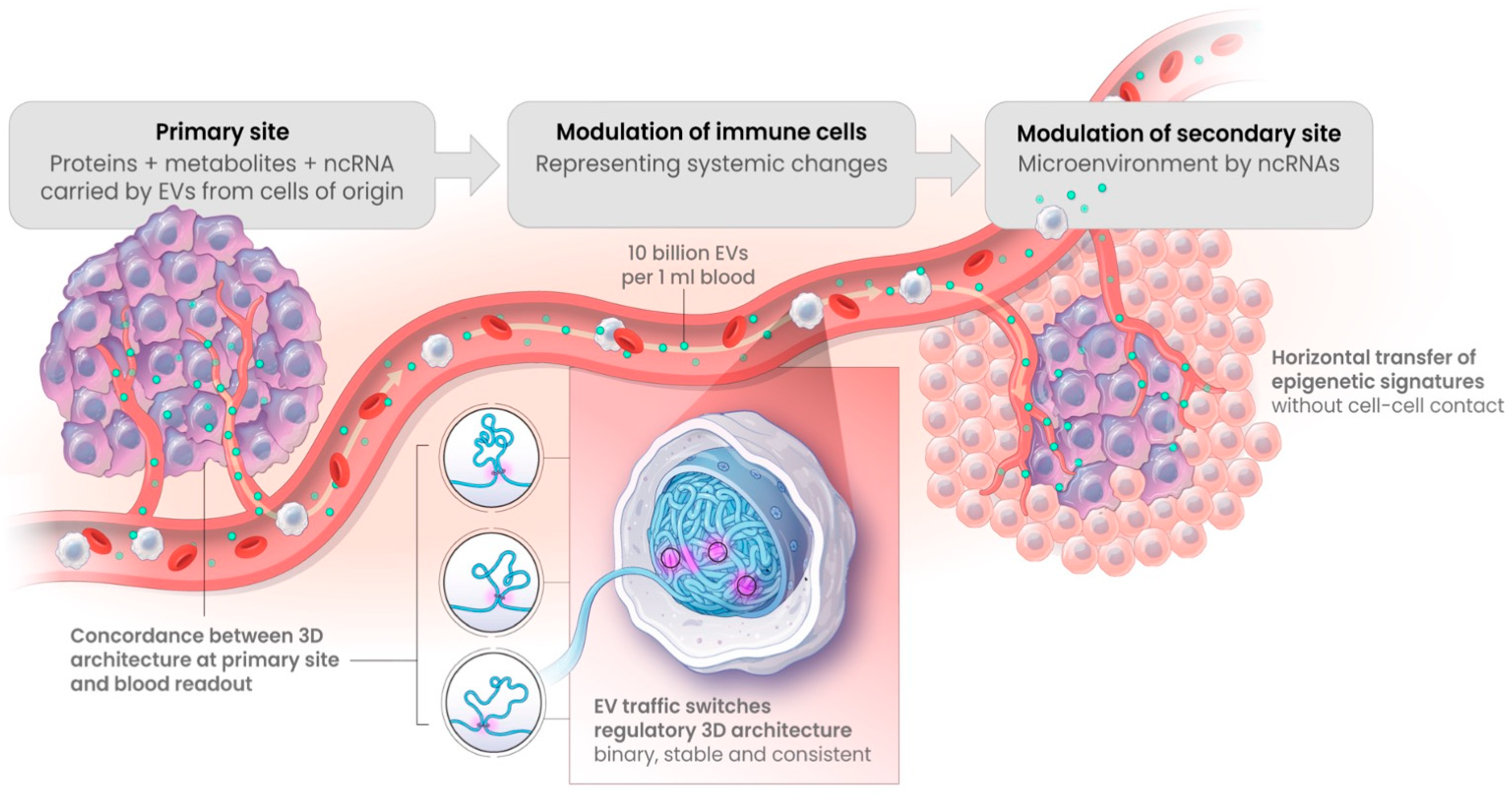
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pardoll, D.M. The blockade of immune checkpoints in cancer immunotherapy. Nat. Rev. Cancer 2012, 12, 252–264. [Google Scholar] [CrossRef] [PubMed]
- Wykes, M.N.; Lewin, S.R. Immune checkpoint blockade in infectious diseases. Nat. Rev. Immunol. 2018, 18, 91–104. [Google Scholar] [CrossRef] [PubMed]
- Morad, G.; Helmink, B.A.; Sharma, P.; Wargo, J.A. Hallmarks of response, resistance, and toxicity to immune checkpoint blockade. Cell 2021, 184, 5309–5337. [Google Scholar] [CrossRef] [PubMed]
- Doyle, A.M.; Mullen, A.C.; Villarino, A.V.; Hutchins, A.S.; High, F.A.; Lee, H.W. Induction of Cytotoxic T Lymphocyte Antigen 4 (Ctla-4) Restricts Clonal Expansion of Helper T Cells. J. Exp. Med. 2001, 194, 893–902. [Google Scholar] [CrossRef] [PubMed]
- Wing, K.; Onishi, Y.; Prieto-Martin, P.; Yamaguchi, T.; Miyara, M.; Fehervari, Z.; Nomura, T.; Sakaguchi, S. CTLA-4 control over Foxp3+ regulatory T cell function. Sci. N. Y. 2008, 322, 271–275. [Google Scholar] [CrossRef]
- O’Day, S.J.; Maio, M.; Chiarion-Sileni, V.; Gajewski, T.F.; Pehamberger, H.; Bondarenko, I.N.; Queirolo, P.; Lundgren, L.; Mikhailov, S.; Roman, L.; et al. Efficacy and safety of ipilimumab monotherapy in patients with pretreated advanced melanoma: A multicenter single-arm phase II study. Ann. Oncol. 2010, 21, 1712–1717. [Google Scholar] [CrossRef]
- Leach, D.R.; Krummel, M.F.; Allison, J.P. Enhancement of Antitumor Immunity by CTLA-4 Blockade. Science 1996, 271, 1734–1736. [Google Scholar] [CrossRef]
- Tivol, E.A.; Borriello, F.; Schweitzer, A.N.; Lynch, W.P.; Bluestone, J.A.; Sharpe, A.H. Loss of CTLA-4 leads to massive lymphoproliferation and fatal multiorgan tissue destruction, revealing a critical negative regulatory role of CTLA-4. Immunity 1995, 3, 541–547. [Google Scholar] [CrossRef]
- Hodi, F.S.; O’Day, S.J.; McDermott, D.F.; Weber, R.W.; Sosman, J.A.; Haanen, J.B.; Gonzales, R.; Robert, C.; Schadendorf, D.; Hassel, J.C.; et al. Improved Survival with Ipilimumab in Patients with Metastatic Melanoma. N. Engl. J. Med. 2010, 363, 711–723. [Google Scholar] [CrossRef]
- Brahmer, J.; Reckamp, K.L.; Baas, P.; Crinò, L.; Eberhardt, W.E.E.; Poddubskaya, E.; Antonia, S.; Pluzanski, A.; Vokes, E.E.; Holdago, E.; et al. Nivolumab versus Docetaxel in Advanced Squamous-Cell Non–Small-Cell Lung Cancer. N. Engl. J. Med. 2015, 373, 123–135. [Google Scholar] [CrossRef]
- Robert, C.; Long, G.V.; Brady, B.; Dutriaux, C.; Maio, M.; Mortier, L.; Hassel, J.C.; Rutkowski, P.; McNeil, C.; Kalinka-Warocha, E.; et al. Nivolumab in Previously Untreated Melanoma without BRAF Mutation. N. Engl. J. Med. 2015, 372, 320–330. [Google Scholar] [CrossRef] [PubMed]
- Garon, E.B.; Rizvi, N.A.; Hui, R.; Leighl, N.; Balmanoukian, A.S.; Eder, J.P.; Patnaik, A.; Aggarwal, C.; Gubens, M.; Horn, L.; et al. Pembrolizumab for the Treatment of Non–Small-Cell Lung Cancer. N. Engl. J. Med. 2015, 372, 2018–2028. [Google Scholar] [CrossRef] [PubMed]
- Patnaik, A.; Kang, S.P.; Rasco, D.; Papadopoulos, K.P.; Elassaiss-Schaap, J.; Beeram, M.; Drengler, R.; Chen, C.; Smith, L.; Espino, G.; et al. Phase I Study of Pembrolizumab (MK-3475; Anti–PD-1 Monoclonal Antibody) in Patients with Advanced Solid Tumors. Clin. Cancer Res. 2015, 21, 4286–4293. [Google Scholar] [CrossRef]
- Topalian, S.L.; Sznol, M.; McDermott, D.F.; Kluger, H.M.; Carvajal, R.D.; Sharfman, W.H.; Brahmer, J.R.; Lawrence, D.P.; Atkins, M.B.; Powderly, J.D.; et al. Survival, Durable Tumor Remission, and Long-Term Safety in Patients With Advanced Melanoma Receiving Nivolumab. J. Clin. Oncol. 2014, 32, 1020–1030. [Google Scholar] [CrossRef] [PubMed]
- Iwata, H.; Inoue, K.; Kaneko, K.; Ito, Y.; Tsugawa, K.; Hasegawa, A.; Nakagawa, S.; Kuratomi, H.; Tamura, K. Subgroup analysis of Japanese patients in a Phase 3 study of atezolizumab in advanced triple-negative breast cancer (IMpassion130). Jpn. J. Clin. Oncol. 2019, 49, 1083–1091. [Google Scholar] [CrossRef]
- Kaufman, H.L.; Russell, J.S.; Hamid, O.; Bhatia, S.; Terheyden, P.; D’Angelo, S.P.; Shih, K.C.; Lebbe, C.; Milella, M.; Brownell, I.; et al. Updated efficacy of avelumab in patients with previously treated metastatic Merkel cell carcinoma after ≥1 year of follow-up: JAVELIN Merkel 200, a phase 2 clinical trial. J. Immunother. Cancer 2018, 6, 7. [Google Scholar] [CrossRef]
- Powles, T.; Durán, I.; Heijden, M.S.; van der Loriot, Y.; Vogelzang, N.J.; Giorgi, U.D.; OUdard, S.; Retz, M.M.; Castellano, D.; Bamias, A.; et al. Atezolizumab versus chemotherapy in patients with platinum-treated locally advanced or metastatic urothelial carcinoma (IMvigor211): A multicentre, open-label, phase 3 randomised controlled trial. Lancet 2018, 391, 748–757. [Google Scholar] [CrossRef]
- Rittmeyer, A.; Barlesi, F.; Waterkamp, D.; Park, K.; Ciardiello, F.; von Pawel, J.; Gadgeel, S.M.; Hida, T.; Kowalski, D.M.; Dols, M.C.; et al. Atezolizumab versus docetaxel in patients with previously treated non-small-cell lung cancer (OAK): A phase 3, open-label, multicentre randomised controlled trial. Lancet 2017, 389, 255–265. [Google Scholar] [CrossRef]
- Ribas, A.; Hamid, O.; Daud, A.; Hodi, F.S.; Wolchok, J.D.; Kefford, R.; Joshua, M.A.; Patnaik, A.; Hwu, W.J.; Weber, J.S.; et al. Association of Pembrolizumab With Tumor Response and Survival among Patients With Advanced Melanoma. JAMA 2016, 315, 1600–1609. [Google Scholar] [CrossRef]
- Patel, M.R.; Ellerton, J.; Infante, J.R.; Agrawal, M.; Gordon, M.; Aljumaily, R.; Britten, C.D.; Dirix, L.; Lee, K.W.; Taylor, M.; et al. Avelumab in metastatic urothelial carcinoma after platinum failure (JAVELIN Solid Tumor): Pooled results from two expansion cohorts of an open-label, phase 1 trial. Lancet Oncol. 2018, 19, 51–64. [Google Scholar] [CrossRef]
- Snyder, A.; Makarov, V.; Merghoub, T.; Yuan, J.; Zaretsky, J.M.; Desrichard, A.; Walsh, L.A.; Postow, M.A.; Wong, P.; Ho, T.S.; et al. Genetic basis for clinical response to CTLA-4 blockade in melanoma. N. Engl. J. Med. 2014, 371, 2189–2199. [Google Scholar] [CrossRef] [PubMed]
- Rizvi, N.A.; Hellmann, M.D.; Snyder, A.; Kvistborg, P.; Makarov, V.; Havel, J.J.; Lee, W.; Yuan, J.; Wong, P.; Ho, T.S.; et al. Mutational landscape determines sensitivity to PD-1 blockade in non–small cell lung cancer. Science 2015, 348, 124–128. [Google Scholar] [CrossRef] [PubMed]
- Le, D.T.; Durham, J.N.; Smith, K.N.; Wang, H.; Bartlett, B.R.; Aulakh, L.K.; Lu, S.; Kemberling, H.; Wilt, C.; Luber, B.S.; et al. Mismatch repair deficiency predicts response of solid tumors to PD-1 blockade. Science 2017, 357, 409–413. [Google Scholar] [CrossRef] [PubMed]
- Riaz, N.; Havel, J.J.; Makarov, V.; Desrichard, A.; Urba, W.J.; Sims, J.S.; Hodi, F.S.; Martin-Algarra, S.; Mandal, R.; Sharfman, W.H.; et al. Tumor and Microenvironment Evolution during Immunotherapy with Nivolumab. Cell 2017, 171, 934–949.e16. [Google Scholar] [CrossRef] [PubMed]
- Tumeh, P.C.; Harview, C.L.; Yearley, J.H.; Shintaku, I.P.; Taylor, E.J.M.; Robert, L.; Chmielowski, B.; Spasic, M.; Henry, G.; Ciobanu, V.; et al. PD-1 blockade induces responses by inhibiting adaptive immune resistance. Nature 2014, 515, 568–571. [Google Scholar] [CrossRef]
- Hendry, S.; Salgado, R.; Gevaert, T.; Russell, P.A.; John, T.; Thapa, B.; Christie, M.; van de Vijver, K.; Estrada, M.V.; Gonzales-Ericsson, P.I.; et al. Assessing Tumor-Infiltrating Lymphocytes in Solid Tumors: A Practical Review for Pathologists and Proposal for a Standardized Method from the International Immuno-Oncology Biomarkers Working Group: Part 2: TILs in Melanoma, Gastrointestinal Tract Carcinomas, Non-Small Cell Lung Carcinoma and Mesothelioma, Endometrial and Ovarian Carcinomas, Squamous Cell Carcinoma of the Head and Neck, Genitourinary Carcinomas, and Primary Brain Tumors. Adv. Anat. Pathol. 2017, 24, 311–335. [Google Scholar]
- Herbst, R.S.; Soria, J.C.; Kowanetz, M.; Fine, G.D.; Hamid, O.; Gordon, M.S.; Sosman, J.A.; McDermott, D.F.; Powderly, J.D.; Gettinger, S.N.; et al. Predictive correlates of response to the anti-PD-L1 antibody MPDL3280A in cancer patients. Nature 2014, 515, 563–567. [Google Scholar] [CrossRef]
- Reck, M.; Rodríguez-Abreu, D.; Robinson, A.G.; Hui, R.; Csőszi, T.; Fülöp, A.; Gottfried, M.; Peled, N.; Tafreshi, A.; Cuffe, S.; et al. Pembrolizumab versus Chemotherapy for PD-L1–Positive Non–Small-Cell Lung Cancer. N. Engl. J. Med. 2016, 375, 1823–1833. [Google Scholar] [CrossRef]
- Daud, A.I.; Wolchok, J.D.; Robert, C.; Hwu, W.J.; Weber, J.S.; Ribas, A.; Hodi, F.S.; Joshua, M.A.; Kefford, R.; Hersey, P.; et al. Programmed Death-Ligand 1 Expression and Response to the Anti–Programmed Death 1 Antibody Pembrolizumab in Melanoma. J. Clin. Oncol. 2016, 34, 4102–4109. [Google Scholar] [CrossRef]
- Tordini, F.; Aldinucci, M.; Milanesi, L.; Liò, P.; Merelli, I. The Genome Conformation As an Integrator of Multi-Omic Data: The Example of Damage Spreading in Cancer. Front. Genet. 2016, 7, 194. [Google Scholar] [CrossRef]
- Bonev, B.; Cavalli, G. Organization and function of the 3D genome. Nat. Rev. Genet. 2016, 17, 661–678. [Google Scholar] [CrossRef] [PubMed]
- Stadhouders, R.; Filion, G.J.; Graf, T. Transcription factors and 3D genome conformation in cell-fate decisions. Nature 2019, 569, 345–354. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Li, K.; Cai, W.; Liu, X.; Zhang, Y.; Orkin, S.H.; Xu, J.; Yuan, G.C. Dissecting super-enhancer hierarchy based on chromatin interactions. Nat. Commun. 2018, 9, 943. [Google Scholar] [CrossRef] [PubMed]
- Di Iulio, J.; Bartha, I.; Wong, E.H.M.; Yu, H.C.; Lavrenko, V.; Yang, D.; Jung, I.; Hicks, M.A.; Shah, N.; Kirkness, E.F.; et al. The human noncoding genome defined by genetic diversity. Nat. Genet. 2018, 50, 333–337. [Google Scholar] [CrossRef]
- Kempfer, R.; Pombo, A. Methods for mapping 3D chromosome architecture. Nat. Rev. Genet. 2019, 21, 207–226. [Google Scholar] [CrossRef]
- Dekker, J.; Rippe, K.; Dekker, M.; Kleckner, N. Capturing Chromosome Conformation. Science 2002, 295, 1306–1311. [Google Scholar] [CrossRef]
- Jakub, J.W.; Grotz, T.E.; Jordan, P.; Hunter, E.; Pittelkow, M.; Ramadass, A.; Akoulitchev, A.; Markovic, S. A pilot study of chromosomal aberrations and epigenetic changes in peripheral blood samples to identify patients with melanoma. Melanoma Res. 2015, 25, 406–411. [Google Scholar] [CrossRef]
- Carini, C.; Hunter, E.; Porter, D.; McInnes, I.; Reid, D.; Ralston, S.H. Chromosome conformation signatures define predictive markers of inadequate response to methotrexate in early rheumatoid arthritis. J. Transl. Med. 2018, 16, 18. [Google Scholar] [CrossRef]
- Bastonini, E.; Jeznach, M.; Field, M.; Juszczyk, K.; Corfield, E.; Dezfouli, M.; Ahmat, N.; Smith, A.; Womersley, H.; Jordan, P.; et al. Chromatin barcodes as biomarkers for melanoma. Pigm. Cell Melanoma R 2014, 27, 788–800. Available online: http://www.ncbi.nlm.nih.gov/pubmed/24807349 (accessed on 2 April 2023). [CrossRef]
- Salter, M.; Corfield, E.; Ramadass, A.; Grand, F.; Green, J.; Westra, J.; Liim, C.R.; Farrimond, L.; Feneberg, E.; Scaber, J.; et al. Initial Identification of a Blood-Based Chromosome Conformation Signature for Aiding in the Diagnosis of Amyotrophic Lateral Sclerosis. Ebiomedicine 2018, 33, 169–184. [Google Scholar] [CrossRef]
- Yan, H.; Hunter, E.; Akoulitchev, A.; Park, P.; Winchester, D.J.; Moo-Young, T.A.; Prinz, R.A. Epigenetic chromatin conformation changes in peripheral blood can detect thyroid cancer. Surgery 2019, 165, 44–49. [Google Scholar] [CrossRef]
- Hunter, E.; McCord, R.; Ramadass, A.S.; Green, J.; Westra, J.W.; Mundt, K.; Akoulitchev, A. Comparative molecular cell-of-origin classification of diffuse large B-cell lymphoma based on liquid and tissue biopsies. Transl. Med. Commun. 2020, 5, 5. [Google Scholar] [CrossRef]
- Salter, M.; Powell, R.; Back, J.; Grand, F.; Koutsothanasi, C.; Green, J.; Hunter, E.; Ramadass, A.; Westra, J.; Akoulitchev, A. Genomic architecture differences at the HTT locus associated with symptomatic and pre-symptomatic cases of Huntington’s disease in a pilot study. F1000research 2019, 7, 1757. [Google Scholar]
- Alshaker, H.; Mills, R.; Hunter, E.; Salter, M.; Ramadass, A.; Skinner, B.M.; Westra, W.; Green, J.; Akoulitchev, A.; Winkler, M.; et al. Chromatin conformation changes in peripheral blood can detect prostate cancer and stratify disease risk groups. J. Transl. Med. 2021, 19, 46. [Google Scholar] [CrossRef]
- Hunter, E.; Koutsothanasi, C.; Wilson, A.; Santos, F.C.; Salter, M.; Powell, R.; Dring, A.; Brajer, P.; Egan, B.; Westra, J.W.; et al. 3D genomic capture of regulatory immuno-genetic profiles in COVID-19 patients for prognosis of severe COVID disease outcome. Biorxiv 2021. [Google Scholar] [CrossRef]
- Hunter, E.; Koutsothanasi, C.; Wilson, A.; Santos, F.C.; Salter, M.; Westra, J.W.; Powel, R.; Dring, A.S.; Brajer, P.; Egan, B.; et al. Development and validation of blood-based prognostic biomarkers for severity of COVID disease outcome using EpiSwitch 3D genomic regulatory immuno-genetic profiling. Medrxiv 2021. Available online: http://medrxiv.org/content/early/2021/06/28/2021.06.21.21259145.abstract (accessed on 28 June 2021).
- Cao, F.; Fang, Y.; Tan, H.K.; Goh, Y.; Choy, J.Y.H.; Koh, B.T.H. Super-Enhancers and Broad H3K4me3 Domains Form Complex Gene Regulatory Circuits Involving Chromatin Interactions. Sci. Rep. 2017, 7, 2186. Available online: http://www.nature.com/articles/s41598-017-02257-3 (accessed on 19 May 2017). [CrossRef] [PubMed]
- Mukhopadhyay, S.; Ramadass, A.S.; Akouloitchev, A.; Gordon, S. Formation of distinct chromatin conformation signatures epigenetically regulate macrophage activation. Int. Immunopharmacol. 2014, 18, 7–11. Available online: http://www.ncbi.nlm.nih.gov/pubmed/24211766 (accessed on 20 January 2014). [CrossRef] [PubMed]
- Hunter, E.; Akoulitchev, A. Clinical and Statistical Considerations in Personalized Medicine; Carini, C., Menon, S.M., Chang, M., Eds.; Current Advances in Epigenetics; CRC Press: Boca Raton, FL, USA, 2014. [Google Scholar]
- Hunter, E.; Potluri, S.; Zhang, S.; Dezfouli, M.; Back, J.; James, L.; Jandor, N.; Powell, R.; Salter, M.; Ramadass, A.S.; et al. Development and validation of baseline predictive biomarkers for response to avelumab in second-line (2L) non-small cell lung cancer (NSCLC) using EpiSwitch epigenetic profiling. J. Immunother. Cancer 2019, 7, 143. [Google Scholar]
- Pchejetski, D.; Hunter, E.; Dezfouli, M.; Salter, M.; Powell, R.; Green, J.; Naithani, T.; Koutsothanasi, C.; Alshaker, H.; Jaipura, J.; et al. Circulating Chromosome Conformation Signatures Significantly Enhance PSA Positive Predicting Value and Overall Accuracy for Prostate Cancer Detection. Cancers 2023, 15, 821. [Google Scholar] [CrossRef]
- Aykan, N.F.; Özatlı, T. Objective response rate assessment in oncology: Current situation and future expectations. World J. Clin. Oncol. 2020, 11, 53–73. [Google Scholar] [CrossRef]
- Levy, B.P.; Giaccone, G.; Besse, B.; Felip, E.; Garassino, M.C.; Gomez, M.D.; Garrido, P.; Piperdi, B.; Ponce-Aix, S.; Menezes, D.; et al. Randomised phase 2 study of pembrolizumab plus CC-486 versus pembrolizumab plus placebo in patients with previously treated advanced non-small cell lung cancer. Eur. J. Cancer 2019, 108, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Tsourkas, A.; Behlke, M.A.; Xu, Y.; Bao, G. Spectroscopic Features of Dual Fluorescence/Luminescence Resonance Energy-Transfer Molecular Beacons. Anal. Chem. 2003, 75, 3697–3703. [Google Scholar] [CrossRef] [PubMed]
- Hancock, J.T.; Khoshgoftaar, T.M. CatBoost for big data: An interdisciplinary review. J. Big Data 2020, 7, 94. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Morris, J.H.; Cook, H.; Kuhn, M.; Wyder, S.; Simonovic, M.; Santos, A.; Doncheva, N.T.; Roth, A.; Bork, P. The STRING database in 2017: Quality-controlled protein–protein association networks, made broadly accessible. Nucleic Acids Res. 2017, 45, D362–D368. [Google Scholar] [CrossRef] [PubMed]
- Fabregat, A.; Sidiropoulos, K.; Viteri, G.; Forner, O.; Marin-Garcia, P.; Arnau, V.; D’Eustachio, P.; Stein, L.; Hermjakob, H. Reactome pathway analysis: A high-performance in-memory approach. BMC Bioinform. 2017, 18, 142. [Google Scholar] [CrossRef] [PubMed]
- Fabregat, A.; Korninger, F.; Viteri, G.; Sidiropoulos, K.; Marin-Garcia, P.; Ping, P.; Wu, G.; Stein, L.; D’Eustachio, P.; Hermjakob, H. Reactome graph database: Efficient access to complex pathway data. PLoS Comput. Biol. 2018, 14, e1005968. [Google Scholar] [CrossRef]
- Scutari, M. Learning Bayesian Networks with the bnlearn R Package. J. Stat. Softw. 2010, 35, 1–22. [Google Scholar] [CrossRef]
- Betzler, A.C.; Theodoraki, M.N.; Schuler, P.J.; Döscher, J.; Laban, S.; Hoffmann, T.K.; Brunner, C. NF-κB and Its Role in Checkpoint Control. Int. J. Mol. Sci. 2020, 21, 3949. [Google Scholar] [CrossRef] [PubMed]
- Ganesh, K.; Massagué, J. TGF-β Inhibition and Immunotherapy: Checkmate. Immunity 2018, 48, 626–628. [Google Scholar] [CrossRef]
- Ravi, R.; Noonan, K.A.; Pham, V.; Bedi, R.; Zhavoronkov, A.; Ozerov, I.V.; Makarev, E.; Artemov, A.V.; Wysotcki, P.T.; Mehra, R.; et al. Bifunctional immune checkpoint-targeted antibody-ligand traps that simultaneously disable TGFβ enhance the efficacy of cancer immunotherapy. Nat. Commun. 2018, 9, 741. [Google Scholar] [CrossRef]
- Guo, Y.; Lu, X.; Chen, Y.; Rendon, B.; Mitchell, R.A.; Cuatrecasas, M.; Cortes, M.; Postigo, A.; Lu, Y.; Dean, D.C. Zeb1 induces immune checkpoints to form an immunosuppressive envelope around invading cancer cells. Sci Adv. 2021, 7, eabd7455. [Google Scholar] [CrossRef] [PubMed]
- Korb, K.B.; Hope, L.R.; Nicholson, A.E.; Axnick, K. PRICAI 2004: Trends in Artificial Intelligence. In Proceedings of the 8th Pacific Rim International Conference on Artificial Intelligence, Auckland, New Zealand, 9–13 August 2004; Springer: Berlin/Heidelberg, Germany, 2004; pp. 322–331. [Google Scholar]
- Man, J.; Millican, J.; Mulvey, A.; Gebski, V.; Hui, R. Response Rate and Survival at Key Timepoints With PD-1 Blockade vs Chemotherapy in PD-L1 Subgroups: Meta-Analysis of Metastatic NSCLC Trials. Jnci. Cancer Spectr. 2021, 5, pkab012. [Google Scholar] [CrossRef] [PubMed]
- Topalian, S.L.; Hodi, F.S.; Brahmer, J.R.; Gettinger, S.N.; Smith, D.C.; McDermott, D.F.; Powderly, J.D.; Carvajal, R.D.; Sosman, J.A.; Atkins, M.B.; et al. Safety, Activity, and Immune Correlates of Anti–PD-1 Antibody in Cancer. N. Engl. J. Med. 2012, 366, 2443–2454. [Google Scholar] [CrossRef] [PubMed]
- Adashek, J.J.; Subbiah, I.M.; Matos, I.; Garralda, E.; Menta, A.K.; Ganeshan, D.M.; Subbiah, V. Hyperprogression and Immunotherapy: Fact, Fiction, or Alternative Fact? Trends Cancer 2020, 6, 181–191. [Google Scholar] [CrossRef] [PubMed]
- Nesline, M.K.; Knight, T.; Colman, S.; Patel, K. Economic Burden of Checkpoint Inhibitor Immunotherapy for the Treatment of Non–Small Cell Lung Cancer in US Clinical Practice. Clin. Ther. 2020, 42, 1682–1698.e7. [Google Scholar] [CrossRef] [PubMed]
- Powles, T.B.; Sridhar, S.; Bellmunt, J.; Sternberg, C.; Grivas, P.; Hunter, E.; Dezfouli, M.; Salter, M.; Powell, R.; Dring, A.; et al. LBA74 Genomic biomarkers in peripheral blood (PB) from patients (pts) enrolled in the JAVELIN Bladder 100 trial of avelumab first-line (1L) maintenance in advanced urothelial carcinoma (aUC). Ann. Oncol. 2022, 33, S1442. [Google Scholar] [CrossRef]
- Kosaka, N.; Ochiya, T. Unraveling the Mystery of Cancer by Secretory microRNA: Horizontal microRNA Transfer between Living Cells. Front. Genet. 2012, 2, 97. [Google Scholar] [CrossRef]
- Melo, S.A.; Sugimoto, H.; O’Connell, J.T.; Kato, N.; Villanueva, A.; Vidal, A.; Qiu, L.; Vikin, E.; Perelman, L.T.; Melo, C.A.; et al. Cancer Exosomes Perform Cell-Independent MicroRNA Biogenesis and Promote Tumorigenesis. Cancer Cell 2014, 26, 707–721. [Google Scholar] [CrossRef]
- Lin, D.; Chen, J.; Perrone-Bizzozero, N.; Bustillo, J.R.; Du, Y.; Calhoun, V.D.; Liu, J. Characterization of cross-tissue genetic-epigenetic effects and their patterns in schizophrenia. Genome Med. 2018, 10, 13. [Google Scholar] [CrossRef]
- Alshaker, H.; Hunter, E.; Salter, M.; Ramadass, A.; Westra, W.; Winkler, M.; Green, J.; Akoulitchev, A.; Pchejetski, D. Monocytes acquire prostate cancer specific chromatin conformations upon indirect co-culture with prostate cancer cells. Front. Oncol. 2022, 12, 990842. [Google Scholar] [CrossRef]
- Serratì, S.; Guida, M.; Fonte, R.D.; Summa, S.D.; Strippoli, S.; Iacobazzi, R.M.; Quarta, A.; De Risis, I.; Guida, G.; Paradiso, A.; et al. Circulating extracellular vesicles expressing PD1 and PD-L1 predict response and mediate resistance to checkpoint inhibitors immunotherapy in metastatic melanoma. Mol. Cancer 2022, 21, 20. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Huang, A.C.; Zhang, W.; Zhang, G.; Wu, M.; Xu, W.; Yu, Z.; Yang, J.; Wang, B.; Sun, H.; et al. Exosomal PD-L1 Contributes to Immunosuppression and is Associated with anti-PD-1 Response. Nature 2018, 560, 382–386. [Google Scholar] [CrossRef]
- Vaddepally, R.K.; Kharel, P.; Pandey, R.; Garje, R.; Chandra, A.B. Review of Indications of FDA-Approved Immune Checkpoint Inhibitors per NCCN Guidelines with the Level of Evidence. Cancers 2020, 12, 738. [Google Scholar] [CrossRef] [PubMed]
- Twomey, J.D.; Zhang, B. Cancer Immunotherapy Update: FDA-Approved Checkpoint Inhibitors and Companion Diagnostics. Aaps J. 2021, 23, 39. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
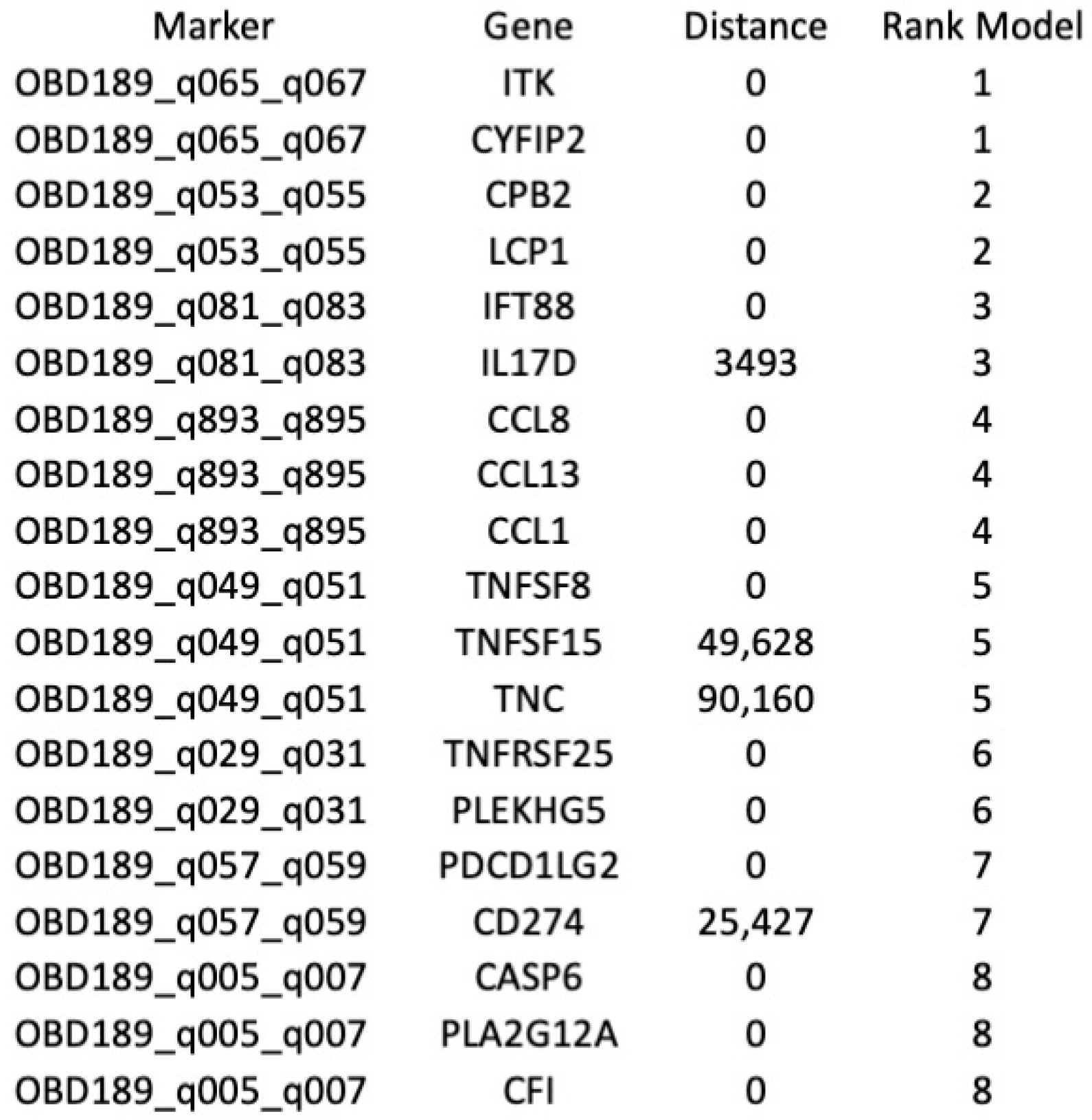
| Probe a | Glmnet_Coef b | p. Value | adj.p.Val c | FC d | Primers Design ID e | Primer1 f | Primer2 f | Gene g |
|---|---|---|---|---|---|---|---|---|
| Hg38_9_5495992_5572986_RR | 0.247101522 | 0.001213344 | 0.039106564 | 1.18337794 | OBD189-q0s57/q059 | GAGGGTCACTCACTGCCCAACAGGC | GACTGTAAGGTAGAAATCCTGCCTGGGT | PDCD1LG2, CD274 |
| Hg38_9_5495992_5572986_RR | 0.247101522 | 0.001213344 | 0.039106564 | 1.18337794 | OBD189-q057/q059 | GAGGGTCACTCACTGCCCAACAGGC | GACTGTAAGGTAGAAATCCTGCCTGGGT | CD274 |
| Hg38_9_5495992_5572986_RR | 0.247101522 | 0.001213344 | 0.039106564 | 1.18337794 | OBD189-q057/q059 | GAGGGTCACTCACTGCCCAACAGGC | GACTGTAAGGTAGAAATCCTGCCTGGGT | RIC1 |
| Hg38_13_20664875_20698635_FF | 0.111815729 | 0.039447494 | 0.258498898 | 1.165419467 | OBD189-q081/q083 | GAAGTGCCACGAGAAGGAGGATGGTCC | GGGCTGTGTCCTGATAAACCCATTGTTA | IFT88 |
| Hg38_13_20664875_20698635_FF | 0.111815729 | 0.039447494 | 0.258498898 | 1.165419467 | OBD189-q081/q083 | GAAGTGCCACGAGAAGGAGGATGGTCC | GGGCTGTGTCCTGATAAACCCATTGTTA | IL17D |
| Hg38_13_20664875_20698635_FF | 0.111815729 | 0.039447494 | 0.258498898 | 1.165419467 | OBD189-q081/q083 | GAAGTGCCACGAGAAGGAGGATGGTCC | GGGCTGTGTCCTGATAAACCCATTGTTA | N6AMT2 |
| Hg38_13_46087370_46193039_RF | 0.132758731 | 0.097937296 | 0.391249484 | 1.09303819 | OBD189-q053/q055 | TAGAAGCAGGGAGTAGTTGAGCAATGGG | TCTTCACTTGTGCTATTGGCTTTCCAGC | CPB2 |
| Hg38_13_46087370_46193039_RF | 0.132758731 | 0.097937296 | 0.391249484 | 1.09303819 | OBD189-q053/q055 | TAGAAGCAGGGAGTAGTTGAGCAATGGG | TCTTCACTTGTGCTATTGGCTTTCCAGC | LCP1 |
| Hg38_13_46087370_46193039_RF | 0.132758731 | 0.097937296 | 0.391249484 | 1.09303819 | OBD189-q053/q055 | TAGAAGCAGGGAGTAGTTGAGCAATGGG | TCTTCACTTGTGCTATTGGCTTTCCAGC | LRRC63 |
| Hg38_15_98731539_98790114_FF | 0.165726908 | 0.108961414 | 0.410037537 | 1.08563184 | OBD189-q001/q003 | GGCTGGTGGGAGTATTTTCAAAGAGAAC | GCTCTGTTCAAGTGGCTCTGTTCCA | IGF1R |
| Hg38_15_98731539_98790114_FF | 0.165726908 | 0.108961414 | 0.410037537 | 1.08563184 | OBD189-q001/q003 | GGCTGGTGGGAGTATTTTCAAAGAGAAC | GCTCTGTTCAAGTGGCTCTGTTCCA | PGPEP1L |
| Hg38_15_98731539_98790114_FF | 0.165726908 | 0.108961414 | 0.410037537 | 1.08563184 | OBD189-q001/q003 | GGCTGGTGGGAGTATTTTCAAAGAGAAC | GCTCTGTTCAAGTGGCTCTGTTCCA | FAM169B |
| Hg38_8_42264241_42332799_FR | 0.10211307 | 0.160069169 | 0.484452574 | 1.079808853 | OBD148_261/263 | CGGTGAGCACGGTCTGTCTACTT | GTCCTGGGTCCTGGGTGAAAGTC | IKBKB |
| Hg38_8_42264241_42332799_FR | 0.10211307 | 0.160069169 | 0.484452574 | 1.079808853 | OBD148_261/263 | CGGTGAGCACGGTCTGTCTACTT | GTCCTGGGTCCTGGGTGAAAGTC | POLB |
| Hg38_8_42264241_42332799_FR | 0.10211307 | 0.160069169 | 0.484452574 | 1.079808853 | OBD148_261/263 | CGGTGAGCACGGTCTGTCTACTT | GTCCTGGGTCCTGGGTGAAAGTC | DKK4 |
| Hg38_1_6461604_6515315_FR | 0.049660907 | 0.171178937 | 0.498342453 | 1.125005092 | OBD189-q029/q031 | TGCCCGTCGTGGTTCCGCCTTCA | AGAGACCCACCCCAGCCTCCTGA | TNFRSF25 |
| Hg38_1_6461604_6515315_FR | 0.049660907 | 0.171178937 | 0.498342453 | 1.125005092 | OBD189-q029/q031 | TGCCCGTCGTGGTTCCGCCTTCA | AGAGACCCACCCCAGCCTCCTGA | PLEKHG5 |
| Hg38_1_6461604_6515315_FR | 0.049660907 | 0.171178937 | 0.498342453 | 1.125005092 | OBD189-q029/q031 | TGCCCGTCGTGGTTCCGCCTTCA | AGAGACCCACCCCAGCCTCCTGA | ESPN |
| Hg38_4_109703339_109741090_RF | 0.050864625 | 0.218283899 | 0.551427137 | 1.109210269 | OBD189-q005/q007 | CCCCAACTCACAACACCCCAGAC | AGAGGAGGGCAAGGTGTCTGGCT | CASP6 |
| Hg38_4_109703339_109741090_RF | 0.050864625 | 0.218283899 | 0.551427137 | 1.109210269 | OBD189-q005/q007 | CCCCAACTCACAACACCCCAGAC | AGAGGAGGGCAAGGTGTCTGGCT | PLA2G12A |
| Hg38_4_109703339_109741090_RF | 0.050864625 | 0.218283899 | 0.551427137 | 1.109210269 | OBD189-q005/q007 | CCCCAACTCACAACACCCCAGAC | AGAGGAGGGCAAGGTGTCTGGCT | CFI |
| Hg38_8_81007411_81099880_FR | 0.115763362 | 0.224881246 | 0.55802192 | 1.062648413 | OBD189-q061/q063 | TGGACAGCCACTACTCAACCTTTTCCTA | CAAACCCAGATTGGACCTCACAGCCCC | PAG1 |
| Hg38_8_81007411_81099880_FR | 0.115763362 | 0.224881246 | 0.55802192 | 1.062648413 | OBD189-q061/q063 | TGGACAGCCACTACTCAACCTTTTCCTA | CAAACCCAGATTGGACCTCACAGCCCC | ZNF704 |
| Hg38_8_81007411_81099880_FR | 0.115763362 | 0.224881246 | 0.55802192 | 1.062648413 | OBD189-q061/q063 | TGGACAGCCACTACTCAACCTTTTCCTA | CAAACCCAGATTGGACCTCACAGCCCC | FABP5 |
| Hg38_1_161633494_161661864_RF | 0.027579594 | 0.247573207 | 0.581047746 | 1.148972903 | OBD189-q041/q043 | TTGCCACCTGTCTCAGATACCCTTGGTT | GCTGCTCCTCTTGCCTGGAATGCCTATT | FCGR2B |
| Hg38_1_161633494_161661864_RF | 0.027579594 | 0.247573207 | 0.581047746 | 1.148972903 | OBD189-q041/q043 | TTGCCACCTGTCTCAGATACCCTTGGTT | GCTGCTCCTCTTGCCTGGAATGCCTATT | FCGR3B |
| Hg38_1_161633494_161661864_RF | 0.027579594 | 0.247573207 | 0.581047746 | 1.148972903 | OBD189-q041/q043 | TTGCCACCTGTCTCAGATACCCTTGGTT | GCTGCTCCTCTTGCCTGGAATGCCTATT | FCRLA |
| Hg38_5_157178319_157271762_RR | 0.026456597 | 0.298498007 | 0.628568025 | 1.13173441 | OBD189-q065/q067 | TGTATGTCTCCTGAGGTGAAGCAAGAGG | CTTCCACCGTGCCCGCAGCCAGC | ITK |
| Hg38_5_157178319_157271762_RR | 0.026456597 | 0.298498007 | 0.628568025 | 1.13173441 | OBD189-q065/q067 | TGTATGTCTCCTGAGGTGAAGCAAGAGG | CTTCCACCGTGCCCGCAGCCAGC | CYFIP2 |
| Hg38_5_157178319_157271762_RR | 0.026456597 | 0.298498007 | 0.628568025 | 1.13173441 | OBD189-q065/q067 | TGTATGTCTCCTGAGGTGAAGCAAGAGG | CTTCCACCGTGCCCGCAGCCAGC | FAM71B |
| Hg38_9_114957908_114977746_RF | −0.026269829 | 0.301172707 | 0.630926161 | −1.124920787 | OBD148-q917/q919 | TTGCTTGTGAGTTTGATGCAG | AAGCCAAATGGGCCTAGCCA | TNFSF8 |
| Hg38_9_114957908_114977746_RF | −0.026269829 | 0.301172707 | 0.630926161 | −1.124920787 | OBD148-q917/q919 | TTGCTTGTGAGTTTGATGCAG | AAGCCAAATGGGCCTAGCCA | TNC |
| Hg38_9_114957908_114977746_RF | −0.026269829 | 0.301172707 | 0.630926161 | −1.124920787 | OBD148-q917/q919 | TTGCTTGTGAGTTTGATGCAG | AAGCCAAATGGGCCTAGCCA | TNFSF15 |
| Hg38_18_62330039_62362521_FR | −0.102570352 | 0.301374789 | 0.631092221 | −1.070331563 | OBD189-q037/q039 | CCTACTGGCACCACTGTGTTGGCTGG | TATCATAATCAGGCAACTGGCTGGTGC | TNFRSF11A |
| Hg38_18_62330039_62362521_FR | −0.102570352 | 0.301374789 | 0.631092221 | −1.070331563 | OBD189-q037/q039 | CCTACTGGCACCACTGTGTTGGCTGG | TATCATAATCAGGCAACTGGCTGGTGC | KIAA1468 |
| Hg38_18_62330039_62362521_FR | −0.102570352 | 0.301374789 | 0.631092221 | −1.070331563 | OBD189-q037/q039 | CCTACTGGCACCACTGTGTTGGCTGG | TATCATAATCAGGCAACTGGCTGGTGC | PIGN |
| Hg38_9_136904007_136941363_RF | 0.021876797 | 0.302188532 | 0.631834649 | 1.115259064 | OBD189-q009/q011 | AGCACTCGTCGTTGGGCGTGTAG | CGGCACACCTCTACTCTCAGCCT | RABL6 |
| Hg38_9_136904007_136941363_RF | 0.021876797 | 0.302188532 | 0.631834649 | 1.115259064 | OBD189-q009/q011 | AGCACTCGTCGTTGGGCGTGTAG | CGGCACACCTCTACTCTCAGCCT | TRAF2 |
| Hg38_9_136904007_136941363_RF | 0.021876797 | 0.302188532 | 0.631834649 | 1.115259064 | OBD189-q009/q011 | AGCACTCGTCGTTGGGCGTGTAG | CGGCACACCTCTACTCTCAGCCT | FBXW5 |
| Hg38_17_34316073_34373948_RF | −0.243856053 | 0.359171973 | 0.67918219 | −1.036704719 | OBD148-q893/q895 | ACTTGTGGCTTCCTTAGCCC | TCCTTTGCAGGTATGGACATC | CCL8 |
| Hg38_17_34316073_34373948_RF | −0.243856053 | 0.359171973 | 0.67918219 | −1.036704719 | OBD148-q893/q895 | ACTTGTGGCTTCCTTAGCCC | TCCTTTGCAGGTATGGACATC | CCL13 |
| Hg38_17_34316073_34373948_RF | −0.243856053 | 0.359171973 | 0.67918219 | −1.036704719 | OBD148-q893/q895 | ACTTGTGGCTTCCTTAGCCC | TCCTTTGCAGGTATGGACATC | CCL1 |
| Hg38_13_20664875_20744490_FR | 0.080919132 | 0.460746676 | 0.752684253 | 1.036210668 | OBD189-q073/q075 | GGAAGTGCCACGAGAAGGAGGATGGTCC | GGTAAGATGAGGCTGTGGGCAAGGAGC | IFT88 |
| Hg38_13_20664875_20744490_FR | 0.080919132 | 0.460746676 | 0.752684253 | 1.036210668 | OBD189-q073/q075 | GGAAGTGCCACGAGAAGGAGGATGGTCC | GGTAAGATGAGGCTGTGGGCAAGGAGC | IL17D |
| Hg38_13_20664875_20744490_FR | 0.080919132 | 0.460746676 | 0.752684253 | 1.036210668 | OBD189-q073/q075 | GGAAGTGCCACGAGAAGGAGGATGGTCC | GGTAAGATGAGGCTGTGGGCAAGGAGC | N6AMT2 |
| Hg38_11_77430379_77519103_RF | 0.150008419 | 0.482939086 | 0.766977325 | 1.027489411 | OBD189-q033/q035 | CATAACCACACTGCTACCAACACACCTA | CTGGTTATTCGGACACTCATAGGACTGG | PAK1 |
| Hg38_11_77430379_77519103_RF | 0.150008419 | 0.482939086 | 0.766977325 | 1.027489411 | OBD189-q033/q035 | CATAACCACACTGCTACCAACACACCTA | CTGGTTATTCGGACACTCATAGGACTGG | CLNS1A |
| Hg38_11_77430379_77519103_RF | 0.150008419 | 0.482939086 | 0.766977325 | 1.027489411 | OBD189-q033/q035 | CATAACCACACTGCTACCAACACACCTA | CTGGTTATTCGGACACTCATAGGACTGG | AQP11 |
| Hg38_8_42264241_42292124_FR | 0.028350069 | 0.486591863 | 0.769305711 | 1.0425716 | OBD189-q025/q027 | GGTGAGCACGGTCTGTCTACTTTCCC | GGACCCAGGCTCTGCTGCTACAG | IKBKB |
| Hg38_8_42264241_42292124_FR | 0.028350069 | 0.486591863 | 0.769305711 | 1.0425716 | OBD189-q025/q027 | GGTGAGCACGGTCTGTCTACTTTCCC | GGACCCAGGCTCTGCTGCTACAG | POLB |
| Hg38_8_42264241_42292124_FR | 0.028350069 | 0.486591863 | 0.769305711 | 1.0425716 | OBD189-q025/q027 | GGTGAGCACGGTCTGTCTACTTTCCC | GGACCCAGGCTCTGCTGCTACAG | PLAT |
| Hg38_18_62296384_62386748_FF | 0.055316179 | 0.499574836 | 0.777354809 | 1.031115956 | OBD189-q045/q047 | CATAGACCCAGGTGTGCTCCGTGGCAGC | GAGCACTGGTTCCCCGCAAATACTGGG | KIAA1468 |
| Hg38_18_62296384_62386748_FF | 0.055316179 | 0.499574836 | 0.777354809 | 1.031115956 | OBD189-q045/q047 | CATAGACCCAGGTGTGCTCCGTGGCAGC | GAGCACTGGTTCCCCGCAAATACTGGG | TNFRSF11A |
| Hg38_18_62296384_62386748_FF | 0.055316179 | 0.499574836 | 0.777354809 | 1.031115956 | OBD189-q045/q047 | CATAGACCCAGGTGTGCTCCGTGGCAGC | GAGCACTGGTTCCCCGCAAATACTGGG | PIGN |
| Hg38_9_114855753_114929419_FR | 0.134189513 | 0.52698868 | 0.793876125 | 1.033468601 | OBD189-q049/q051 | CCATTGTTGCTCAGGCTGCCCTCTTGC | GCATTCAAGTGACAGAGAGAAAAGAGGC | TNFSF8 |
| Hg38_9_114855753_114929419_FR | 0.134189513 | 0.52698868 | 0.793876125 | 1.033468601 | OBD189-q049/q051 | CCATTGTTGCTCAGGCTGCCCTCTTGC | GCATTCAAGTGACAGAGAGAAAAGAGGC | TNFSF15 |
| Hg38_9_114855753_114929419_FR | 0.134189513 | 0.52698868 | 0.793876125 | 1.033468601 | OBD189-q049/q051 | CCATTGTTGCTCAGGCTGCCCTCTTGC | GCATTCAAGTGACAGAGAGAAAAGAGGC | TNC |
| Hg38_8_26561792_26644530_FR | 0.055843287 | 0.625162161 | 0.848039488 | 1.042292141 | OBD189-q069/q071 | CAGTATGAGTGTTCTGTGGCTGCTCCCA | GCGTGTCTCTCAGGGAAGGCAGGATGC | DPYSL2 |
| Hg38_8_26561792_26644530_FR | 0.055843287 | 0.625162161 | 0.848039488 | 1.042292141 | OBD189-q069/q071 | CAGTATGAGTGTTCTGTGGCTGCTCCCA | GCGTGTCTCTCAGGGAAGGCAGGATGC | PNMA2 |
| Hg38_8_26561792_26644530_FR | 0.055843287 | 0.625162161 | 0.848039488 | 1.042292141 | OBD189-q069/q071 | CAGTATGAGTGTTCTGTGGCTGCTCCCA | GCGTGTCTCTCAGGGAAGGCAGGATGC | BNIP3L |
| Hg38_8_127691489_127740424_FR | −0.000580983 | 0.676617738 | 0.873913467 | 1.034088403 | OBD189-q013/q015 | GTCACCTTCATCTCCTTCTCACAGCAG | GCTTCGCTTACCAGAGTCGCTGC | MYC |
| Hg38_8_127691489_127740424_FR | −0.000580983 | 0.676617738 | 0.873913467 | 1.034088403 | OBD189-q013/q015 | GTCACCTTCATCTCCTTCTCACAGCAG | GCTTCGCTTACCAGAGTCGCTGC | AC108925.1 |
| Hg38_8_127691489_127740424_FR | −0.000580983 | 0.676617738 | 0.873913467 | 1.034088403 | OBD189-q013/q015 | GTCACCTTCATCTCCTTCTCACAGCAG | GCTTCGCTTACCAGAGTCGCTGC | POU5F1B |
| Hg38_9_120888366_120919710_RR | 0.018753385 | 0.784545989 | 0.922028968 | 1.010214644 | OBD189-q017/q019 | CCCAGTTGTCCAGGTTGCTGCCT | CCTGGAGCAGAACCTGTCAGACC | PHF19 |
| Hg38_9_120888366_120919710_RR | 0.018753385 | 0.784545989 | 0.922028968 | 1.010214644 | OBD189-q017/q019 | CCCAGTTGTCCAGGTTGCTGCCT | CCTGGAGCAGAACCTGTCAGACC | TRAF1 |
| Hg38_9_120888366_120919710_RR | 0.018753385 | 0.784545989 | 0.922028968 | 1.010214644 | OBD189-q017/q019 | CCCAGTTGTCCAGGTTGCTGCCT | CCTGGAGCAGAACCTGTCAGACC | C5 |
| Hg38_13_20664875_20691044_FF | −0.023912747 | 0.80365095 | 0.929901824 | −1.012741717 | OBD189-q077/q079 | GGAAGTGCCACGAGAAGGAGGATGGTCC | CCACCCAGTTCCTCCAGGCATAGCAGG | IFT88 |
| Hg38_13_20664875_20691044_FF | −0.023912747 | 0.80365095 | 0.929901824 | −1.012741717 | OBD189-q077/q079 | GGAAGTGCCACGAGAAGGAGGATGGTCC | CCACCCAGTTCCTCCAGGCATAGCAGG | IL17D |
| Hg38_13_20664875_20691044_FF | −0.023912747 | 0.80365095 | 0.929901824 | −1.012741717 | OBD189-q077/q079 | GGAAGTGCCACGAGAAGGAGGATGGTCC | CCACCCAGTTCCTCCAGGCATAGCAGG | N6AMT2 |
| Hg38_5_168579937_168620163_RR | 0.019685283 | 0.8953453 | 0.964751362 | −1.005369684 | OBD189-q021/q023 | CCGACCCTAACATTCAAGGTGTCTCTAT | GAGTCAGCGTGTAGTGCTCCCAC | PANK3 |
| Hg38_5_168579937_168620163_RR | 0.019685283 | 0.8953453 | 0.964751362 | −1.005369684 | OBD189-q021/q023 | CCGACCCTAACATTCAAGGTGTCTCTAT | GAGTCAGCGTGTAGTGCTCCCAC | SLIT3 |
| Hg38_5_168579937_168620163_RR | 0.019685283 | 0.8953453 | 0.964751362 | −1.005369684 | OBD189-q021/q023 | CCGACCCTAACATTCAAGGTGTCTCTAT | GAGTCAGCGTGTAGTGCTCCCAC | FBLL1 |
| Episwitch Interaction a | Primers Design ID b | Probe Used | Opt Ann Tm c | QC Optimisation d | Screen 1 | Screen 2 |
|---|---|---|---|---|---|---|
| CASP6_4_109703339_109741090_RF | OBD189-q005/q007 | OBD189-p005 | 68 | Passed | Passed | Passed |
| IGF1R_15_98731539_98790114_FF | OBD189-q001/q003 | OBD189-p003 | 64.4 | Passed | Failed | |
| IKBKB_8_42264241_42292124_FR | OBD189-q025/q027 | OBD189-p025 | 66.4 | Passed | Failed | |
| IKBKB_8_42264241_42332799_FR | OBD148_261/263 | OBD189-p261 | 68 | Passed | Failed | |
| IL17D_13_20664875_20691044_FF | OBD189-q077/q079 | OBD189-p077 | 67.5 | Passed | Failed | |
| ITK_5_157178319_157271762_RR | OBD189-q065/q067 | OBD189-p065 | 66.4 | Passed | Passed | Passed |
| MYC_8_127691489_127740424_FR | OBD189-q013/q015 | OBD189-p013 | 66.4 | Passed | Failed | |
| ORF102_17_34316073_34373948_RF | OBD148-q893/q895 | OBD148-p893 | 62 | Passed | Passed | Passed |
| ORF197_8_26561792_26644530_FR | OBD189-q069/q071 | N/A e | N/A | Failed | Failed | |
| ORF243_1_161633494_161661864_RF | OBD189-q041/q043 | OBD189-p043 | 62 | Passed | Failed | |
| ORF313_13_20664875_20698635_FF | OBD189-q081/q083 | OBD189-p081 | 64.4 | Passed | Passed | Passed |
| ORF313_13_20664875_20744490_FR | OBD189-q073/q075 | N/A | N/A | Failed | Failed | |
| ORF369_13_46087370_46193039_RF | OBD189-q053/q055 | OBD189-p053 | 67.5 | Passed | Passed | Passed |
| ORF479_8_81007411_81099880_FR | OBD189-q061/q063 | OBD189-p061 | 64.4 | Passed | Failed | |
| ORF480_11_77430379_77519103_RF | OBD189-q033/q035 | OBD189-p033 | 66.4 | Passed | Failed | |
| ORF482_5_168579937_168620163_RR | OBD189-q021/q023 | OBD189-p021 | 66.4 | Passed | Failed | |
| ORF698_18_62296384_62386748_FF | OBD189-q045/q047 | OBD189-p045 | 66.4 | Passed | Failed | |
| ORF698_18_62330039_62362521_FR | OBD189-q037/q039 | N/A | N/A | Failed | Failed | |
| ORF703_1_6461604_6515315_FR | OBD189-q029/q031 | OBD189-p031 | 67.5 | Passed | Passed | Passed |
| ORF705_9_114855753_114929419_FR | OBD189-q049/q051 | OBD189-p049 | 66.4 | Passed | Passed | Passed |
| ORF712_9_120888366_120919710_RR | OBD189-q017/q019 | OBD189-p017 | 68 | Passed | Failed | |
| PDCD1LG2_9_5495992_5572986_RR | OBD189-q057/q059 | OBD189-p057 | 67.5 | Passed | Passed | Passed |
| TNFSF8_9_114957908_114977746_RF | OBD148-q917/q919 | N/A | N/A | Failed | Failed | |
| TRAF2_9_136904007_136941363_RF | OBD189-q009/q011 | OBD189-p009 | 64.4 | Passed | Failed |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hunter, E.; Salter, M.; Powell, R.; Dring, A.; Naithani, T.; Chatziioannou, M.E.; Gebregzabhar, A.; Issa, M.; Green, J.; Ng, S.; et al. Development and Validation of Blood-Based Predictive Biomarkers for Response to PD-1/PD-L1 Checkpoint Inhibitors: Evidence of a Universal Systemic Core of 3D Immunogenetic Profiling across Multiple Oncological Indications. Cancers 2023, 15, 2696. https://doi.org/10.3390/cancers15102696
Hunter E, Salter M, Powell R, Dring A, Naithani T, Chatziioannou ME, Gebregzabhar A, Issa M, Green J, Ng S, et al. Development and Validation of Blood-Based Predictive Biomarkers for Response to PD-1/PD-L1 Checkpoint Inhibitors: Evidence of a Universal Systemic Core of 3D Immunogenetic Profiling across Multiple Oncological Indications. Cancers. 2023; 15(10):2696. https://doi.org/10.3390/cancers15102696
Chicago/Turabian StyleHunter, Ewan, Matthew Salter, Ryan Powell, Ann Dring, Tarun Naithani, Maria Eleni Chatziioannou, Abel Gebregzabhar, Mutaz Issa, Jayne Green, Serene Ng, and et al. 2023. "Development and Validation of Blood-Based Predictive Biomarkers for Response to PD-1/PD-L1 Checkpoint Inhibitors: Evidence of a Universal Systemic Core of 3D Immunogenetic Profiling across Multiple Oncological Indications" Cancers 15, no. 10: 2696. https://doi.org/10.3390/cancers15102696
APA StyleHunter, E., Salter, M., Powell, R., Dring, A., Naithani, T., Chatziioannou, M. E., Gebregzabhar, A., Issa, M., Green, J., Ng, S., Lim, C. R., Keat, C. S., Suan, A. T., Raman, R., Fatt, H. K., Luen, F. L. W., Alshaker, H., Pchejetski, D., Blum, D., ... Akoulitchev, A. (2023). Development and Validation of Blood-Based Predictive Biomarkers for Response to PD-1/PD-L1 Checkpoint Inhibitors: Evidence of a Universal Systemic Core of 3D Immunogenetic Profiling across Multiple Oncological Indications. Cancers, 15(10), 2696. https://doi.org/10.3390/cancers15102696