
The CAM Model—Q&A with Experts
 ,
,  , , , ,
, , , ,  ,
,  , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Simple Summary
Abstract
1. Introduction
We Start with a Simple Question—What Is the CAM?
2. Regularities
2.1. What Are the Major General Prerequisites for Alternative Animal Test System?
2.2. What Is Needed to Run a CAM Lab?
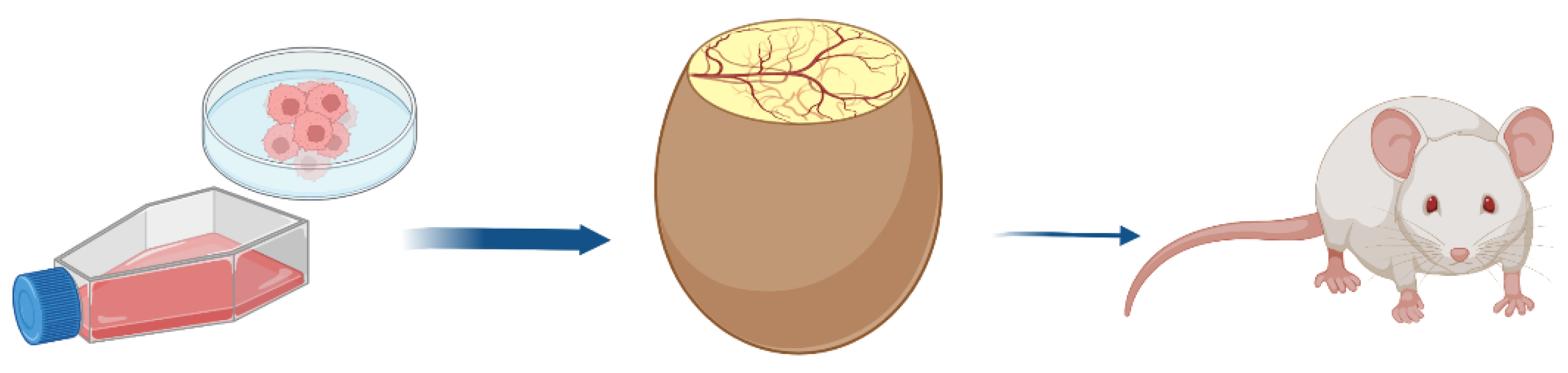
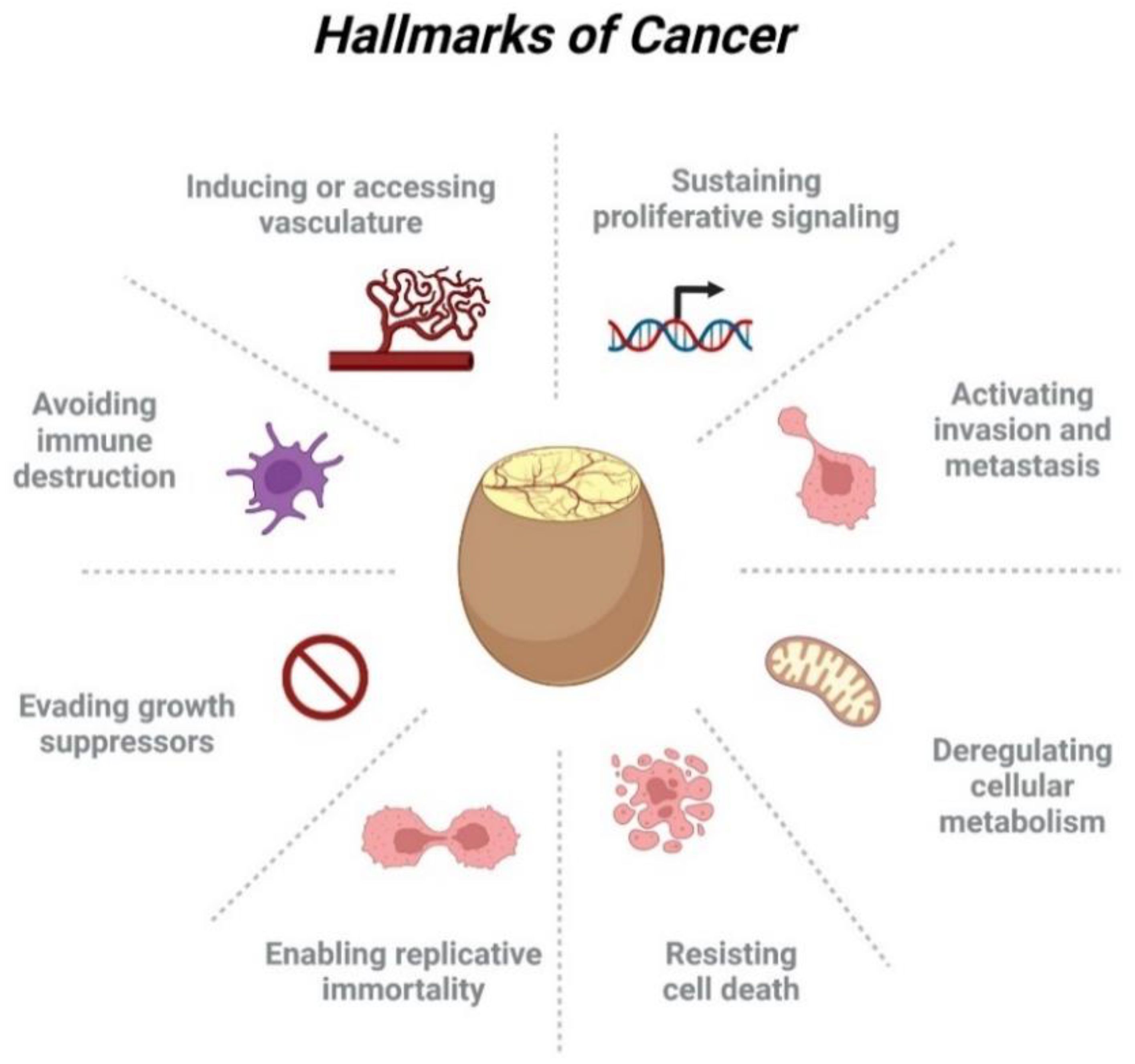
3. Tumor Growth Features on the CAM
3.1. How Can a CAM Tumor Graft Be Exploited?
3.2. How Can CAM Xenografts Be Used to Study Cell Death?
3.3. How Can the CAM Model Be Used to Study Cancer Cell Stemness?
4. Tumor Metastasis in the CAM Model
4.1. Is the CAM a Complete In Vivo Model for Studying Metastasis?
4.2. How Can the CAM Model Be Used to Study Tumor Invasion at the Primary Site?
4.3. Can the CAM Model Be Useful in Tumor Metastasis Research?
5. Angiogenesis on the CAM
5.1. Why the CAM Assay May Be Considered a Suitable Experimental Model to Study Tumor Angiogenesis?
5.2. Can Imaging Be Applied to Monitor Angiogenesis and Tumor Growth?
6. The Chicken Immune System
6.1. Is the Immunological Situation of the Eggs Comparable to the Human Situation?
6.2. Does Tumor-Induced Inflammation in the CAM Model Parallel the Typical Immune Cell Infiltration Observed in Human Tumors?
6.3. How Does the Developing Chicken Immune System Respond/Contribute to Human Tumor Xenografts?
7. The CAM Model in Immuno-Oncology
7.1. Can the CAM Model Be Used as a Powerful Model for Immuno-Oncology?
7.2. How Can the Model Be Used for Drug Discovery and Testing?
7.3. How Widely Can the CAM Assay Be Used for Oncology Drug Discovery?
7.4. Can Imaging Be Applied for Initial Testing of Pharmacokinetics and Bio-Distribution?
8. The CAM Model in Personalized Medicine
8.1. In What Ways Will the CAM Model Facilitate and Extend Possibilities for Developing Personalized/Precision Medicine Solutions in Oncology?
8.2. In Particular, What Are the Specific Benefits for Personalized/Precision Medicine That Can Only Be Achieved Using the CAM Model?
8.3. How to Leverage on the CAM Model as a Living Biobank to Facilitate the Precision and Personalized Medicine for Patients in the Real World?
8.4. Is the CAM Model an Animal Experiment and What Are the Consequences for Investigational New Drug (IND) Documentation?
9. Limitation and Advantages of the CAM Model
9.1. What Are the Advantages?
9.2. What Are the Limitations?
9.3. When Debate Is Ending in a Tie, Is It However a Win-Win for Researchers?
10. Conclusions/Outlook/Perspectives
11. Patents
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Weber, J.; Weber, M.; Steinle, H.; Schlensak, C.; Wendel, H.-P.; Avci-Adali, M. An alternative in vivo model to evaluate pluripotency of patient-specific iPSCs. ALTEX-Altern. Anim. Exp. 2021, 38, 442–450. [Google Scholar] [CrossRef] [PubMed]
- Miebach, L.; Berner, J.; Bekeschus, S. In ovo model in cancer research and tumor immunology. Front. Immunol. 2022, 13, 1006064. [Google Scholar] [CrossRef] [PubMed]
- Pion, E.; Karnosky, J.; Boscheck, S.; Wagner, B.J.; Schmidt, K.M.; Brunner, S.M.; Schlitt, H.J.; Aung, T.; Hackl, C.; Haerteis, S. 3D In Vivo Models for Translational Research on Pancreatic Cancer: The Chorioallantoic Membrane (CAM) Model. Cancers 2022, 14, 3733. [Google Scholar] [CrossRef] [PubMed]
- Nagai, H.; Tanoue, Y.; Nakamura, T.; Chan, C.J.J.; Yamada, S.; Saitou, M.; Fukuda, T.; Sheng, G. Mesothelial fusion mediates chorioallantoic membrane formation. Philos. Trans. R Soc. Lond. B Biol. Sci. 2022, 377, 20210263. [Google Scholar] [CrossRef]
- Schlatter, P.; König, M.F.; Karlsson, L.M.; Burri, P.H. Quantitative study of intussusceptive capillary growth in the chorioallantoic membrane (CAM) of the chicken embryo. Microvasc. Res. 1997, 54, 65–73. [Google Scholar] [CrossRef]
- Ausprunk, D.H.; Knighton, D.R.; Folkman, J. Differentiation of vascular endothelium in the chick chorioallantois: A structural and autoradiographic study. Dev. Biol. 1974, 38, 237–248. [Google Scholar] [CrossRef]
- Official Journal of the European Union. Directive 2010/63/EU of the European Parlament and of the Council of 22 September 2010. Available online: https://eur-lex.europa.eu/ (accessed on 2 November 2022).
- Hanahan, D. Hallmarks of Cancer: New Dimensions. Cancer Discov. 2022, 12, 31–46. [Google Scholar] [CrossRef]
- Chu, P.Y.; Koh, A.P.; Antony, J.; Huang, R.Y. Applications of the Chick Chorioallantoic Membrane as an Alternative Model for Cancer Studies. Cells Tissues Organs. 2022, 211, 222–237. [Google Scholar] [CrossRef]
- Mapanao, A.K.; Che, P.P.; Sarogni, P.; Sminia, P.; Giovannetti, E.; Voliani, V. Tumor grafted—Chick chorioallantoic membrane as an alternative model for biological cancer research and conventional/nanomaterial-based theranostics evaluation. Expert Opin. Drug Metab. Toxicol. 2021, 17, 947–968. [Google Scholar] [CrossRef]
- Kunze, P.; Kreiss, L.; Novosadová, V.; Roehe, A.V.; Steinmann, S.; Prochazka, J.; Geppert, C.I.; Hartmann, A.; Schürmann, S.; Friedrich, O.; et al. Multiphoton Microscopy Reveals DAPK1-Dependent Extracellular Matrix Remodeling in a Chorioallantoic Membrane (CAM) Model. Cancers 2022, 14, 2364. [Google Scholar] [CrossRef]
- Hagedorn, M.; Javerzat, S.; Gilges, D.; Meyre, A.; de Lafarge, B.; Eichmann, A.; Bikfalvi, A. Accessing key steps of human tumor progression in vivo by using an avian embryo model. Proc. Natl. Acad. Sci. USA 2005, 102, 1643–1648. [Google Scholar] [CrossRef] [PubMed]
- Dünker, N.; Jendrossek, V. Implementation of the Chick Chorioallantoic Membrane (CAM) Model in Radiation Biology and Experimental Radiation Oncology Research. Cancers 2019, 11, 1499. [Google Scholar] [CrossRef] [PubMed]
- Ndreshkjana, B.; Çapci, A.; Klein, V.; Chanvorachote, P.; Muenzner, J.K.; Huebner, K.; Steinmann, S.; Erlenbach-Wuensch, K.; Geppert, C.I.; Agaimy, A.; et al. Combination of 5-fluorouracil and thymoquinone targets stem cell gene signature in colorectal cancer cells. Cell Death Dis. 2019, 10, 379. [Google Scholar] [CrossRef]
- Sacks, D.; Baxter, B.; Campbell, B.C.V.; Carpenter, J.S.; Cognard, C.; Dippel, D.; Eesa, M.; Fischer, U.; Hausegger, K.; Hirsch, J.A.; et al. Multisociety Consensus Quality Improvement Revised Consensus Statement for Endovascular Therapy of Acute Ischemic Stroke. Int. J. Stroke 2018, 13, 612–632. [Google Scholar] [CrossRef]
- Senga, S.S.; Grose, R.P. Hallmarks of cancer-the new testament. Open Biol. 2021, 11, 200358. [Google Scholar] [CrossRef] [PubMed]
- Rich, J.N. Cancer stem cells: Understanding tumor hierarchy and heterogeneity. Medicine (Baltimore) 2016, 95, S2–S7. [Google Scholar] [CrossRef]
- De Angelis, M.L.; Francescangeli, F.; Zeuner, A. Breast Cancer Stem Cells as Drivers of Tumor Chemoresistance, Dormancy and Relapse: New Challenges and Therapeutic Opportunities. Cancers 2019, 11, 1569. [Google Scholar] [CrossRef]
- Muenzner, J.K.; Kunze, P.; Lindner, P.; Polaschek, S.; Menke, K.; Eckstein, M.; Geppert, C.I.; Chanvorachote, P.; Baeuerle, T.; Hartmann, A.; et al. Generation and characterization of hepatocellular carcinoma cell lines with enhanced cancer stem cell potential. J. Cell. Mol. Med. 2018, 22, 6238–6248. [Google Scholar] [CrossRef]
- Mélin, C.; Perraud, A.; Christou, N.; Bibes, R.; Cardot, P.; Jauberteau, M.O.; Battu, S.; Mathonnet, M. New ex-ovo colorectal-cancer models from different SdFFF-sorted tumor-initiating cells. Anal. Bioanal. Chem. 2015, 407, 8433–8443. [Google Scholar] [CrossRef]
- Dionísio, M.R.; Vieira, A.F.; Carvalho, R.; Conde, I.; Oliveira, M.; Gomes, M.; Pinto, M.T.; Pereira, P.; Pimentel, J.; Souza, C.; et al. BR-BCSC Signature: The Cancer Stem Cell Profile Enriched in Brain Metastases that Predicts a Worse Prognosis in Lymph Node-Positive Breast Cancer. Cells 2020, 9, 2442. [Google Scholar] [CrossRef]
- Pinto, M.T.; Ribeiro, A.S.; Conde, I.; Carvalho, R.; Paredes, J. The Chick Chorioallantoic Membrane Model: A New In Vivo Tool to Evaluate Breast Cancer Stem Cell Activity. Int. J. Mol. Sci. 2020, 22, 334. [Google Scholar] [CrossRef] [PubMed]
- Pizon, M.; Schott, D.; Pachmann, U.; Schobert, R.; Pizon, M.; Wozniak, M.; Bobinski, R.; Pachmann, K. Chick Chorioallantoic Membrane (CAM) Assays as a Model of Patient-Derived Xenografts from Circulating Cancer Stem Cells (cCSCs) in Breast Cancer Patients. Cancers 2022, 14, 1476. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. The hallmarks of cancer. Cell 2000, 100, 57–70. [Google Scholar] [CrossRef]
- Bragado, P.; Estrada, Y.; Parikh, F.; Krause, S.; Capobianco, C.; Farina, H.G.; Schewe, D.M.; Aguirre-Ghiso, J.A. TGF-β2 dictates disseminated tumour cell fate in target organs through TGF-β-RIII and p38α/β signalling. Nat. Cell Biol. 2013, 15, 1351–1361. [Google Scholar] [CrossRef] [PubMed]
- Fluegen, G.; Avivar-Valderas, A.; Wang, Y.; Padgen, M.R.; Williams, J.K.; Nobre, A.R.; Calvo, V.; Cheung, J.F.; Bravo-Cordero, J.J.; Entenberg, D.; et al. Phenotypic heterogeneity of disseminated tumour cells is preset by primary tumour hypoxic microenvironments. Nat. Cell Biol. 2017, 19, 120–132. [Google Scholar] [CrossRef] [PubMed]
- Nogami, H.; Urist, M.R. The classic: A morphogenetic matrix for differentiation of cartilage in tissue culture. Clin. Orthop. Relat. Res. 2009, 467, 3063–3067. [Google Scholar] [CrossRef][Green Version]
- Palaniappan, T.K.; Šlekienė, L.; Jonasson, A.K.; Gilthorpe, J.; Gunhaga, L. CAM-Delam: An in vivo approach to visualize and quantify the delamination and invasion capacity of human cancer cells. Sci. Rep. 2020, 10, 10472. [Google Scholar] [CrossRef]
- Gomes, C.; Osório, H.; Pinto, M.T.; Campos, D.; Oliveira, M.J.; Reis, C.A. Expression of ST3GAL4 leads to SLe(x) expression and induces c-Met activation and an invasive phenotype in gastric carcinoma cells. PLoS ONE 2013, 8, e66737. [Google Scholar] [CrossRef]
- Ferreira, L.B.; Tavares, C.; Pestana, A.; Pereira, C.L.; Eloy, C.; Pinto, M.T.; Castro, P.; Batista, R.; Rios, E.; Sobrinho-Simões, M.; et al. Osteopontin-a splice variant is overexpressed in papillary thyroid carcinoma and modulates invasive behavior. Oncotarget 2016, 7, 52003–52016. [Google Scholar] [CrossRef]
- Niemelä, E.; Desai, D.; Niemi, R.; Doroszko, M.; Özliseli, E.; Kemppainen, K.; Rahman, N.A.; Sahlgren, C.; Törnquist, K.; Eriksson, J.E.; et al. Nanoparticles carrying fingolimod and methotrexate enables targeted induction of apoptosis and immobilization of invasive thyroid cancer. Eur. J. Pharm.Biopharm. Off. J. Arb. Fur Pharm. Verfahr. e.V 2020, 148, 1–9. [Google Scholar] [CrossRef]
- Lokman, N.A.; Elder, A.S.; Ricciardelli, C.; Oehler, M.K. Chick chorioallantoic membrane (CAM) assay as an in vivo model to study the effect of newly identified molecules on ovarian cancer invasion and metastasis. Int. J. Mol. Sci. 2012, 13, 9959–9970. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Pathak, R.R.; Lopez-Rivera, E.; Friedman, S.L.; Aguirre-Ghiso, J.A.; Sikora, A.G. The In Ovo Chick Chorioallantoic Membrane (CAM) Assay as an Efficient Xenograft Model of Hepatocellular Carcinoma. J. Vis. Exp. 2015, 104, e52411. [Google Scholar] [CrossRef] [PubMed]
- Xiao, X.; Zhou, X.; Ming, H.; Zhang, J.; Huang, G.; Zhang, Z.; Li, P. Chick Chorioallantoic Membrane Assay: A 3D Animal Model for Study of Human Nasopharyngeal Carcinoma. PloS ONE 2015, 10, e0130935. [Google Scholar] [CrossRef] [PubMed]
- Martowicz, A.; Spizzo, G.; Gastl, G.; Untergasser, G. Phenotype-dependent effects of EpCAM expression on growth and invasion of human breast cancer cell lines. BMC Cancer 2012, 12, 501. [Google Scholar] [CrossRef]
- Steinmann, S.; Kunze, P.; Hampel, C.; Eckstein, M.; Bertram Bramsen, J.; Muenzner, J.K.; Carlé, B.; Ndreshkjana, B.; Kemenes, S.; Gasparini, P.; et al. DAPK1 loss triggers tumor invasion in colorectal tumor cells. Cell Death Dis. 2019, 10, 895. [Google Scholar] [CrossRef]
- Demir, R.; Naschberger, L.; Demir, I.; Melling, N.; Dimmler, A.; Papadopoulus, T.; Sturzl, M.; Klein, P.; Hohenberger, W. Hypoxia generates a more invasive phenotype of tumour cells: An in vivo experimental setup based on the chorioallantoic membrane. Pathol. Oncol. Res. POR 2009, 15, 417–422. [Google Scholar] [CrossRef]
- Sun, L.; Lin, P.; Qin, Z.; Liu, Y.; Deng, L.L.; Lu, C. Hypoxia promotes HO-8910PM ovarian cancer cell invasion via Snail-mediated MT1-MMP upregulation. Exp. Biol. Med. 2015, 240, 1434–1445. [Google Scholar] [CrossRef]
- Janser, F.A.; Ney, P.; Pinto, M.T.; Langer, R.; Tschan, M.P. The Chick Chorioallantoic Membrane (CAM) Assay as a Three-dimensional Model to Study Autophagy in Cancer Cells. Bio Protoc. 2019, 9, e3290. [Google Scholar] [CrossRef]
- Wang, Z.; Da Silva, T.G.; Jin, K.; Han, X.; Ranganathan, P.; Zhu, X.; Sanchez-Mejias, A.; Bai, F.; Li, B.; Fei, D.L.; et al. Notch signaling drives stemness and tumorigenicity of esophageal adenocarcinoma. Cancer Res. 2014, 74, 6364–6374. [Google Scholar] [CrossRef]
- Kain, K.H.; Miller, J.W.; Jones-Paris, C.R.; Thomason, R.T.; Lewis, J.D.; Bader, D.M.; Barnett, J.V.; Zijlstra, A. The chick embryo as an expanding experimental model for cancer and cardiovascular research. Dev. Dyn. Off. Publ. Am. Assoc. Anat. 2014, 243, 216–228. [Google Scholar] [CrossRef]
- Liu, M.; Scanlon, C.S.; Banerjee, R.; Russo, N.; Inglehart, R.C.; Willis, A.L.; Weiss, S.J.; D’Silva, N.J. The Histone Methyltransferase EZH2 Mediates Tumor Progression on the Chick Chorioallantoic Membrane Assay, a Novel Model of Head and Neck Squamous Cell Carcinoma. Transl. Oncol. 2013, 6, 273–281. [Google Scholar] [CrossRef] [PubMed]
- Zijlstra, A.; Mellor, R.; Panzarella, G.; Aimes, R.T.; Hooper, J.D.; Marchenko, N.D.; Quigley, J.P. A quantitative analysis of rate-limiting steps in the metastatic cascade using human-specific real-time polymerase chain reaction. Cancer Res. 2002, 62, 7083–7092. [Google Scholar] [PubMed]
- van der Horst, E.H.; Leupold, J.H.; Schubbert, R.; Ullrich, A.; Allgayer, H. TaqMan-based quantification of invasive cells in the chick embryo metastasis assay. Biotechniques 2004, 37, 940–942, 944, 946. [Google Scholar] [CrossRef] [PubMed]
- Ausprunk, D.H.; Folkman, J. Vascular injury in transplanted tissues. Fine structural changes in tumor, adult, and embryonic blood vessels. Virchows Arch. B Cell Pathol. 1976, 21, 31–44. [Google Scholar] [CrossRef]
- Ausprunk, D.H.; Knighton, D.R.; Folkman, J. Vascularization of normal and neoplastic tissues grafted to the chick chorioallantois. Role of host and preexisting graft blood vessels. Am. J. Pathol. 1975, 79, 597–618. [Google Scholar]
- Ribatti, D. The chick embryo chorioallantoic membrane as a model for tumor biology. Exp. Cell Res. 2014, 328, 314–324. [Google Scholar] [CrossRef]
- Hagedorn, M.; Balke, M.; Schmidt, A.; Bloch, W.; Kurz, H.; Javerzat, S.; Rousseau, B.; Wilting, J.; Bikfalvi, A. VEGF coordinates interaction of pericytes and endothelial cells during vasculogenesis and experimental angiogenesis. Dev.Dyn. Off. Publ. Am. Assoc. Anat. 2004, 230, 23–33. [Google Scholar] [CrossRef]
- Leong, H.S.; Steinmetz, N.F.; Ablack, A.; Destito, G.; Zijlstra, A.; Stuhlmann, H.; Manchester, M.; Lewis, J.D. Intravital imaging of embryonic and tumor neovasculature using viral nanoparticles. Nat. Protoc. 2010, 5, 1406–1417. [Google Scholar] [CrossRef]
- Cho, C.F.; Ablack, A.; Leong, H.S.; Zijlstra, A.; Lewis, J. Evaluation of nanoparticle uptake in tumors in real time using intravital imaging. J. Vis. Exp. 2011, 52, 2808. [Google Scholar] [CrossRef]
- Balke, M.; Neumann, A.; Kersting, C.; Agelopoulos, K.; Gebert, C.; Gosheger, G.; Buerger, H.; Hagedorn, M. Morphologic characterization of osteosarcoma growth on the chick chorioallantoic membrane. BMC Res. Notes 2010, 3, 58. [Google Scholar] [CrossRef]
- Jefferies, B.; Lenze, F.; Sathe, A.; Truong, N.; Anton, M.; von Eisenhart-Rothe, R.; Nawroth, R.; Mayer-Kuckuk, P. Non-invasive imaging of engineered human tumors in the living chicken embryo. Sci. Rep. 2017, 7, 4991. [Google Scholar] [CrossRef] [PubMed]
- Sys, G.M.; Lapeire, L.; Stevens, N.; Favoreel, H.; Forsyth, R.; Bracke, M.; De Wever, O. The in ovo CAM-assay as a xenograft model for sarcoma. J. Vis. Exp. 2013, 77, e50522. [Google Scholar] [CrossRef]
- Khan, S.; Pathak, P.; Vasudevan, S.; Nayak, D. Non-invasive photoacoustic screening of blood vasculature during anti-angiogenesis using CAM assay. OSA Continuum 2021, 4, 2821–2836. [Google Scholar] [CrossRef]
- Eckrich, J.; Kugler, P.; Buhr, C.R.; Ernst, B.P.; Mendler, S.; Baumgart, J.; Brieger, J.; Wiesmann, N. Monitoring of tumor growth and vascularization with repetitive ultrasonography in the chicken chorioallantoic-membrane-assay. Sci. Rep. 2020, 10, 18585. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.S.; Min, J.; Recknagel, A.K.; Riccio, M.; Butcher, J.T. Quantitative three-dimensional analysis of embryonic chick morphogenesis via microcomputed tomography. Anat. Rec. 2011, 294, 1–10. [Google Scholar] [CrossRef]
- Zuo, Z.; Syrovets, T.; Genze, F.; Abaei, A.; Ma, G.; Simmet, T.; Rasche, V. High-resolution MRI analysis of breast cancer xenograft on the chick chorioallantoic membrane. NMR Biomed. 2015, 28, 440–447. [Google Scholar] [CrossRef]
- Gebhardt, P.; Würbach, L.; Heidrich, A.; Heinrich, L.; Walther, M.; Opfermann, T.; Sørensen, B.; Saluz, H.P. Dynamic behaviour of selected PET tracers in embryonated chicken eggs. Rev. Esp. Med. Nucl. Imagen Mol. 2013, 32, 371–377. [Google Scholar] [CrossRef]
- Warnock, G.; Turtoi, A.; Blomme, A.; Bretin, F.; Bahri, M.A.; Lemaire, C.; Libert, L.C.; Seret, A.E.; Luxen, A.; Castronovo, V.; et al. In vivo PET/CT in a human glioblastoma chicken chorioallantoic membrane model: A new tool for oncology and radiotracer development. J. Nucl. Med. 2013, 54, 1782–1788. [Google Scholar] [CrossRef]
- Löffler, J.; Hamp, C.; Scheidhauer, E.; Di Carlo, D.; Solbach, C.; Abaei, A.; Hao, L.; Glatting, G.; Beer, A.J.; Rasche, V.; et al. Comparison of Quantification of Target-Specific Accumulation of [18F]F-siPSMA-14 in the HET-CAM Model and in Mice Using PET/MRI. Cancers 2021, 13, 4007. [Google Scholar] [CrossRef]
- Simons, M.; Alitalo, K.; Annex, B.H.; Augustin, H.G.; Beam, C.; Berk, B.C.; Byzova, T.; Carmeliet, P.; Chilian, W.; Cooke, J.P.; et al. State-of-the-Art Methods for Evaluation of Angiogenesis and Tissue Vascularization: A Scientific Statement From the American Heart Association. Circ. Res. 2015, 116, e99–e132. [Google Scholar] [CrossRef]
- Naik, M.; Brahma, P.; Dixit, M. A Cost-Effective and Efficient Chick Ex-Ovo CAM Assay Protocol to Assess Angiogenesis. Methods Protoc. 2018, 1, 19. [Google Scholar] [CrossRef] [PubMed]
- Borozdova, M.; Stiukhina, E.; Sdobnov, A.; Fedosov, I.; Postnov, D.; Tuchin, V. Quantitative measurement of blood flow dynamics in chorioallantoic membrane of chicken embryo using laser Doppler anemometry. In Proceedings of the Saratov Fall Meeting 2015: Third International Symposium on Optics and Biophotonics and Seventh Finnish-Russian Photonics and Laser Symposium (PALS), Saratov, Russian, 21 April 2016; Volume 9917, pp. 231–239. Available online: https://ui.adsabs.harvard.edu/abs/2016SPIE.9917E..0WB/abstract (accessed on 2 November 2022). [CrossRef]
- Mangir, N.; Raza, A.; Haycock, J.W.; Chapple, C.; Macneil, S. An Improved In Vivo Methodology to Visualise Tumour Induced Changes in Vasculature Using the Chick Chorionic Allantoic Membrane Assay. In Vivo 2018, 32, 461–472. [Google Scholar] [CrossRef] [PubMed]
- Shi, P.; Hong, J.; Huang, Y.; Zhang, Z.; Zhang, M.; Zhang, L. Automated computational framework of blood vessel quantification in chick chorioallantoic membrane angiogenesis. J. Biomed. Opt. 2014, 19, 106005. [Google Scholar] [CrossRef] [PubMed]
- Blacher, S.; Devy, L.; Hlushchuk, R.; Larger, E.; Lamande, N.; Burri, P.; Corvol, P.; Djonov, V.; Foidart, J.; Noel, A. Quantification of angiogenesis in the chicken chorioallantoic membrane (CAM). Image Anal. Stereol. 2005, 24. [Google Scholar] [CrossRef]
- Kuri, P.M.; Pion, E.; Mahl, L.; Kainz, P.; Schwarz, S.; Brochhausen, C.; Aung, T.; Haerteis, S. Deep Learning-Based Image Analysis for the Quantification of Tumor-Induced Angiogenesis in the 3D In Vivo Tumor Model—Establishment and Addition to Laser Speckle Contrast Imaging (LSCI). Cells 2022, 11, 2321. [Google Scholar]
- Power, E.A.; Fernandez-Torres, J.; Zhang, L.; Yaun, R.; Lucien, F.; Daniels, D.J. Chorioallantoic membrane (CAM) assay to study treatment effects in diffuse intrinsic pontine glioma. PloS ONE 2022, 17, e0263822. [Google Scholar] [CrossRef]
- Waschkies, C.F.; Pfiffner, F.K.; Heuberger, D.M.; Schneider, M.A.; Tian, Y.; Wolint, P.; Calcagni, M.; Giovanoli, P.; Buschmann, J. Tumor grafts grown on the chicken chorioallantoic membrane are distinctively characterized by MRI under functional gas challenge. Sci. Rep. 2020, 10, 7505. [Google Scholar] [CrossRef]
- Dupertuis, Y.M.; Delie, F.; Cohen, M.; Pichard, C. In ovo method for evaluating the effect of nutritional therapies on tumor development, growth and vascularization. Clin. Nutr. Exp. 2015, 2, 9–17. [Google Scholar] [CrossRef]
- Winter, G.; Koch, A.B.F.; Löffler, J.; Jelezko, F.; Lindén, M.; Li, H.; Abaei, A.; Zuo, Z.; Beer, A.J.; Rasche, V. In vivo PET/MRI Imaging of the Chorioallantoic Membrane. Front. Phys. 2020, 8, 151. [Google Scholar] [CrossRef]
- Ribatti, D.; Tamma, R.; Elieh Ali Komi, D. The morphological basis of the development of the chick embryo immune system. Exp. Cell Res. 2019, 381, 323–329. [Google Scholar] [CrossRef]
- Garcia, P.; Wang, Y.; Viallet, J.; Macek Jilkova, Z. The Chicken Embryo Model: A Novel and Relevant Model for Immune-Based Studies. Front. Immunol. 2021, 12, 791081. [Google Scholar] [CrossRef] [PubMed]
- Consortium, I.C.G.S. Sequence and comparative analysis of the chicken genome provide unique perspectives on vertebrate evolution. Nature 2004, 432, 695–716. [Google Scholar] [CrossRef]
- Alkie, T.N.; Yitbarek, A.; Hodgins, D.C.; Kulkarni, R.R.; Taha-Abdelaziz, K.; Sharif, S. Development of innate immunity in chicken embryos and newly hatched chicks: A disease control perspective. Avian Pathol. 2019, 48, 288–310. [Google Scholar] [CrossRef]
- Rashidi, H.; Sottile, V. The chick embryo: Hatching a model for contemporary biomedical research. BioEssays News Rev. Mol. Cell. Dev. Biol. 2009, 31, 459–465. [Google Scholar] [CrossRef]
- Rosenbruch, M.; Holst, A. The chick embryo yolk-sac blood vessel system as an experimental model for irritation and inflammation. Toxicol. In Vitro 1990, 4, 327–331. [Google Scholar] [CrossRef] [PubMed]
- Gil, M.; Ramil, F.; AgÍs, J.A. Hydroids (Cnidaria, Hydrozoa) from Mauritanian Coral Mounds. Zootaxa 2020, 4878, 412–466. [Google Scholar] [CrossRef]
- Greten, F.R.; Grivennikov, S.I. Inflammation and Cancer: Triggers, Mechanisms, and Consequences. Immunity 2019, 51, 27–41. [Google Scholar] [CrossRef]
- Giraldo, N.A.; Sanchez-Salas, R.; Peske, J.D.; Vano, Y.; Becht, E.; Petitprez, F.; Validire, P.; Ingels, A.; Cathelineau, X.; Fridman, W.H.; et al. The clinical role of the TME in solid cancer. Br. J. Cancer 2019, 120, 45–53. [Google Scholar] [CrossRef]
- Hibino, S.; Kawazoe, T.; Kasahara, H.; Itoh, S.; Ishimoto, T.; Sakata-Yanagimoto, M.; Taniguchi, K. Inflammation-Induced Tumorigenesis and Metastasis. Int. J. Mol. Sci. 2021, 22, 5421. [Google Scholar] [CrossRef]
- Kartikasari, A.E.R.; Huertas, C.S.; Mitchell, A.; Plebanski, M. Tumor-Induced Inflammatory Cytokines and the Emerging Diagnostic Devices for Cancer Detection and Prognosis. Front. Oncol. 2021, 11, 692142. [Google Scholar] [CrossRef]
- Benarafa, C. Tumor-induced inflammation alters neutrophil phenotype and disease progression. Breast Cancer Res. 2015, 17, 135. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.; Wan, G. Tumor-associated macrophages in immunotherapy. FEBS J. 2021, 288, 6174–6186. [Google Scholar] [CrossRef] [PubMed]
- Valdes, T.I.; Kreutzer, D.; Moussy, F. The chick chorioallantoic membrane as a novel in vivo model for the testing of biomaterials. J. Biomed. Mater. Res. 2002, 62, 273–282. [Google Scholar] [CrossRef] [PubMed]
- Kogut, M.H.; Lowry, V.K.; Moyes, R.B.; Bowden, L.L.; Bowden, R.; Genovese, K.; Deloach, J.R. Lymphokine-augmented activation of avian heterophils. Poult. Sci. 1998, 77, 964–971. [Google Scholar] [CrossRef]
- Kogut, M.H.; Holtzapple, C.; Lowry, V.K.; Genovese, K.; Stanker, L.H. Functional responses of neonatal chicken and turkey heterophils following stimulation by inflammatory agonists. Am. J. Vet. Res. 1998, 59, 1404–1408. [Google Scholar]
- Sharma, B.K.; Kakker, N.K.; Bhadouriya, S.; Chhabra, R. Effect of TLR agonist on infections bronchitis virus replication and cytokine expression in embryonated chicken eggs. Mol. Immunol. 2020, 120, 52–60. [Google Scholar] [CrossRef]
- Bar Shira, E.; Friedman, A. Innate immune functions of avian intestinal epithelial cells: Response to bacterial stimuli and localization of responding cells in the developing avian digestive tract. PloS ONE 2018, 13, e0200393. [Google Scholar] [CrossRef]
- Wang, Y.; Rousset, X.; Prunier, C.; Garcia, P.; Dosda, E.; Leplus, E.; Viallet, J. PD-1/PD-L1 Checkpoint Inhibitors Are Active in the Chicken Embryo Model and Show Antitumor Efficacy In Ovo. Cancers 2022, 14, 3095. [Google Scholar] [CrossRef]
- Rousset, X.; Dosda, E.; Viallet, J. Use of an Egg Grafted with Tumor Cells in Order to Study the Anti-Cancer Effectiveness of Immune Therapies in the Absence of Immune Effector Cells Other Than Those in the Grafted Egg. Patent FR3087793A1, 1 May 2020. [Google Scholar]
- Chen, L.; Wang, S.; Feng, Y.; Zhang, J.; Du, Y.; Zhang, J.; Ongeval, C.V.; Ni, Y.; Li, Y. Utilisation of Chick Embryo Chorioallantoic Membrane as a Model Platform for Imaging-Navigated Biomedical Research. Cells 2021, 10, 463. [Google Scholar] [CrossRef]
- Ribatti, D. The chick embryo chorioallantoic membrane (CAM). A multifaceted experimental model. Mech. Dev. 2016, 141, 70–77. [Google Scholar] [CrossRef]
- Ribatti, D.; Tamma, R. The chick embryo chorioallantoic membrane as an in vivo experimental model to study multiple myeloma. Enzymes 2019, 46, 23–35. [Google Scholar] [CrossRef] [PubMed]
- DeBord, L.C.; Pathak, R.R.; Villaneuva, M.; Liu, H.C.; Harrington, D.A.; Yu, W.; Lewis, M.T.; Sikora, A.G. The chick chorioallantoic membrane (CAM) as a versatile patient-derived xenograft (PDX) platform for precision medicine and preclinical research. Am. J. Cancer Res. 2018, 8, 1642–1660. [Google Scholar] [PubMed]
- Li, Q.X.; Feuer, G.; Ouyang, X.; An, X. Experimental animal modeling for immuno-oncology. Pharmacol. Ther. 2017, 173, 34–46. [Google Scholar] [CrossRef] [PubMed]
- Ochoa de Olza, M.; Oliva, M.; Hierro, C.; Matos, I.; Martin-Liberal, J.; Garralda, E. Early-drug development in the era of immuno-oncology: Are we ready to face the challenges? Ann.Oncol. Off. J. Eur. Soc. Med. Oncol. 2018, 29, 1727–1740. [Google Scholar] [CrossRef] [PubMed]
- Ohno, Y.; Kaneko, T.; Inoue, T.; Morikawa, Y.; Yoshida, T.; Fujii, A.; Masuda, M.; Ohno, T.; Hayashi, M.; Momma, J.; et al. Interlaboratory validation of the in vitro eye irritation tests for cosmetic ingredients. (1) Overview of the validation study and Draize scores for the evaluation of the tests. Toxicol. In Vitro 1999, 13, 73–98. [Google Scholar] [CrossRef]
- Wong, C.H.; Siah, K.W.; Lo, A.W. Estimation of clinical trial success rates and related parameters. Biostatistics 2018, 20, 273–286. [Google Scholar] [CrossRef]
- Victorelli, F.D.; Cardoso, V.M.O.; Ferreira, N.N.; Calixto, G.M.F.; Fontana, C.R.; Baltazar, F.; Gremião, M.P.D.; Chorilli, M. Chick embryo chorioallantoic membrane as a suitable in vivo model to evaluate drug delivery systems for cancer treatment: A review. Eur. J. Pharm.Biopharm. Off. J. Arb. Fur Pharm. Verfahr. e.V 2020, 153, 273–284. [Google Scholar] [CrossRef]
- Richardson, M.; Singh, G. Observations on the use of the avian chorioallantoic membrane (CAM) model in investigations into angiogenesis. Curr. Drug Targets Cardiovasc. Haematol. Disord. 2003, 3, 155–185. [Google Scholar] [CrossRef]
- Durupt, F.; Koppers-Lalic, D.; Balme, B.; Budel, L.; Terrier, O.; Lina, B.; Thomas, L.; Hoeben, R.C.; Rosa-Calatrava, M. The chicken chorioallantoic membrane tumor assay as model for qualitative testing of oncolytic adenoviruses. Cancer Gene Ther. 2012, 19, 58–68. [Google Scholar] [CrossRef]
- Ardelean, S.; Feflea, S.; Ionescu, D.; Năstase, V.; Dehelean, C.A. Toxicologic screening of some surfactants using modern in vivo bioassays. Rev. Med. Chir Soc. Med. Nat. Iasi 2011, 115, 251–258. [Google Scholar]
- Saw, C.L.; Heng, P.W.; Liew, C.V. Chick chorioallantoic membrane as an in situ biological membrane for pharmaceutical formulation development: A review. Drug Dev. Ind. Pharm. 2008, 34, 1168–1177. [Google Scholar] [CrossRef] [PubMed]
- Brun, S.; Bassissi, F.; Serdjebi, C.; Novello, M.; Tracz, J.; Autelitano, F.; Guillemot, M.; Fabre, P.; Courcambeck, J.; Ansaldi, C.; et al. GNS561, a new lysosomotropic small molecule, for the treatment of intrahepatic cholangiocarcinoma. Investig. New Drugs 2019, 37, 1135–1145. [Google Scholar] [CrossRef] [PubMed]
- Kroiss, A.; Vincent, S.; Decaussin-Petrucci, M.; Meugnier, E.; Viallet, J.; Ruffion, A.; Chalmel, F.; Samarut, J.; Allioli, N. Androgen-regulated microRNA-135a decreases prostate cancer cell migration and invasion through downregulating ROCK1 and ROCK2. Oncogene 2015, 34, 2846–2855. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.J.; Chiou, P.P.; Lin, P.; Lin, C.M.; Siri, S.; Peck, K.; Chen, T.T. Suppression of growth and cancer-induced angiogenesis of aggressive human breast cancer cells (MDA-MB-231) on the chorioallantoic membrane of developing chicken embryos by E-peptide of pro-IGF-I. J. Cell Biochem. 2007, 101, 1316–1327. [Google Scholar] [CrossRef] [PubMed]
- Colman, R.W.; Pixley, R.A.; Sainz, I.M.; Song, J.S.; Isordia-Salas, I.; Muhamed, S.N.; Powell, J.A., Jr.; Mousa, S.A. Inhibition of angiogenesis by antibody blocking the action of proangiogenic high-molecular-weight kininogen. J. Thromb. Haemost. 2003, 1, 164–170. [Google Scholar] [CrossRef]
- Marcion, G.; Hermetet, F.; Neiers, F.; Uyanik, B.; Dondaine, L.; Dias, A.M.M.; Da Costa, L.; Moreau, M.; Bellaye, P.S.; Collin, B.; et al. Nanofitins targeting heat shock protein 110: An innovative immunotherapeutic modality in cancer. Int. J. Cancer 2021, 148, 3019–3031. [Google Scholar] [CrossRef]
- Krutzke, L.; Allmendinger, E.; Hirt, K.; Kochanek, S. Chorioallantoic Membrane Tumor Model for Evaluating Oncolytic Viruses. Hum. Gene Ther. 2020, 31, 1100–1113. [Google Scholar] [CrossRef]
- Leclair, H.M.; Tardif, N.; Paris, A.; Galibert, M.D.; Corre, S. Role of Flavonoids in the Prevention of AhR-Dependent Resistance During Treatment with BRAF Inhibitors. Int. J. Mol. Sci. 2020, 21, 5025. [Google Scholar] [CrossRef]
- Bjørnstad, S.; Austdal, L.P.; Roald, B.; Glover, J.C.; Paulsen, R.E. Cracking the Egg: Potential of the Developing Chicken as a Model System for Nonclinical Safety Studies of Pharmaceuticals. J. Pharmacol. Exp. Ther. 2015, 355, 386–396. [Google Scholar] [CrossRef]
- Pawlikowska, P.; Tayoun, T.; Oulhen, M.; Faugeroux, V.; Rouffiac, V.; Aberlenc, A.; Pommier, A.L.; Honore, A.; Marty, V.; Bawa, O.; et al. Exploitation of the chick embryo chorioallantoic membrane (CAM) as a platform for anti-metastatic drug testing. Sci. Rep. 2020, 10, 16876. [Google Scholar] [CrossRef]
- Nascimento, B.F.O.; Laranjo, M.; Pereira, N.A.M.; Dias-Ferreira, J.; Piñeiro, M.; Botelho, M.F.; Pinho, E.M.T. Ring-Fused Diphenylchlorins as Potent Photosensitizers for Photodynamic Therapy Applications: In Vitro Tumor Cell Biology and in Vivo Chick Embryo Chorioallantoic Membrane Studies. ACS Omega 2019, 4, 17244–17250. [Google Scholar] [CrossRef] [PubMed]
- Klein, F.G.; Granier, C.; Zhao, Y.; Pan, Q.; Tong, Z.; Gschwend, J.E.; Holm, P.S.; Nawroth, R. Combination of Talazoparib and Palbociclib as a Potent Treatment Strategy in Bladder Cancer. J. Pers Med. 2021, 11, 340. [Google Scholar] [CrossRef] [PubMed]
- Dorrell, M.I.; Kast-Woelbern, H.R.; Botts, R.T.; Bravo, S.A.; Tremblay, J.R.; Giles, S.; Wada, J.F.; Alexander, M.; Garcia, E.; Villegas, G.; et al. A novel method of screening combinations of angiostatics identifies bevacizumab and temsirolimus as synergistic inhibitors of glioma-induced angiogenesis. PloS ONE 2021, 16, e0252233. [Google Scholar] [CrossRef] [PubMed]
- Kue, C.S.; Tan, K.Y.; Lam, M.L.; Lee, H.B. Chick embryo chorioallantoic membrane (CAM): An alternative predictive model in acute toxicological studies for anti-cancer drugs. Exp. Anim. 2015, 64, 129–138. [Google Scholar] [CrossRef] [PubMed]
- Šlekienė, L.; Stakišaitis, D.; Balnytė, I.; Valančiūtė, A. Sodium Valproate Inhibits Small Cell Lung Cancer Tumor Growth on the Chicken Embryo Chorioallantoic Membrane and Reduces the p53 and EZH2 Expression. Dose Response 2018, 16, 1559325818772486. [Google Scholar] [CrossRef]
- Tufan, A.C.; Satiroglu-Tufan, N.L. The chick embryo chorioallantoic membrane as a model system for the study of tumor angiogenesis, invasion and development of anti-angiogenic agents. Curr. Cancer Drug Targets 2005, 5, 249–266. [Google Scholar] [CrossRef]
- Ribatti, D. The chick embryo chorioallantoic membrane in the study of tumor angiogenesis. Rom. J. Morphol. Embryol. 2008, 49, 131–135. [Google Scholar]
- Achkar, I.W.; Kader, S.; Dib, S.S.; Junejo, K.; Al-Bader, S.B.; Hayat, S.; Bhagwat, A.M.; Rousset, X.; Wang, Y.; Viallet, J.; et al. Metabolic Signatures of Tumor Responses to Doxorubicin Elucidated by Metabolic Profiling in Ovo. Metabolites 2020, 10, 268. [Google Scholar] [CrossRef]
- Preis, E.; Schulze, J.; Gutberlet, B.; Pinnapireddy, S.R.; Jedelská, J.; Bakowsky, U. The chorioallantoic membrane as a bio-barrier model for the evaluation of nanoscale drug delivery systems for tumour therapy. Adv. Drug Deliv. Rev. 2021, 174, 317–336. [Google Scholar] [CrossRef]
- Vu, B.T.; Shahin, S.A.; Croissant, J.; Fatieiev, Y.; Matsumoto, K.; Le-Hoang Doan, T.; Yik, T.; Simargi, S.; Conteras, A.; Ratliff, L.; et al. Chick chorioallantoic membrane assay as an in vivo model to study the effect of nanoparticle-based anticancer drugs in ovarian cancer. Sci. Rep. 2018, 8, 8524. [Google Scholar] [CrossRef]
- Zuo, Z.; Syrovets, T.; Wu, Y.; Hafner, S.; Vernikouskaya, I.; Liu, W.; Ma, G.; Weil, T.; Simmet, T.; Rasche, V. The CAM cancer xenograft as a model for initial evaluation of MR labelled compounds. Sci. Rep. 2017, 7, 46690. [Google Scholar] [CrossRef] [PubMed]
- Lenkavska, L.; Blascakova, L.; Jurasekova, Z.; Macajova, M.; Bilcik, B.; Cavarga, I.; Miskovsky, P.; Huntosova, V. Benefits of hypericin transport and delivery by low- and high-density lipoproteins to cancer cells: From in vitro to ex ovo. Photodiagnosis Photodyn. Ther. 2019, 25, 214–224. [Google Scholar] [CrossRef] [PubMed]
- Löffler, J.; Herrmann, H.; Scheidhauer, E.; Wirth, M.; Wasserloos, A.; Solbach, C.; Glatting, G.; Beer, A.J.; Rasche, V.; Winter, G. Blocking Studies to Evaluate Receptor-Specific Radioligand Binding in the CAM Model by PET and MR Imaging. Cancers 2022, 14, 3870. [Google Scholar] [CrossRef] [PubMed]
- Kenakin, T.P. Pharmacokinetics. In A Pharmacology Primer—Techniques for More Effective and Strategic Drug Discovery, 5th ed.; Elsevier: Berlin/Heidelberg, Germany, 2018. [Google Scholar]
- Ribatti, D.; Nico, B.; Bertossi, M. The development of the blood-brain barrier in the chick. Studies with Evans blue and horseradish peroxidase. Ann. Anat.-Anat. Anz. 1993, 175, 85–88. [Google Scholar] [CrossRef]
- Roncali, L.; Nico, B.; Ribatti, D.; Bertossi, M.; Mancini, L. Microscopical and ultrastructural investigations on the development of the blood-brain barrier in the chick embryo optic tectum. Acta Neuropathol. 1986, 70, 193–201. [Google Scholar] [CrossRef]
- Chiba, Y.; Khandoker, A.H.; Nobuta, M.; Moriya, K.; Akiyama, R.; Tazawa, H. Development of respiratory rhythms in perinatal chick embryos. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2002, 131, 817–824. [Google Scholar] [CrossRef]
- Compton, S.K.; Goeringer, G.C. Lung development in the chick embryo. I. Phosphatidylcholine synthesis in the developing chick lung. Pediatr. Res. 1981, 15, 866–869. [Google Scholar] [CrossRef]
- Bolin, G.; Burggren, W.W. Metanephric kidney development in the chicken embryo: Glomerular numbers, characteristics and perfusion. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2013, 166, 343–350. [Google Scholar] [CrossRef]
- Wong, G.K.; Cavey, M.J. Development of the liver in the chicken embryo. I. Hepatic cords and sinusoids. Anat. Rec. 1992, 234, 555–567. [Google Scholar] [CrossRef]
- Wong, G.K.; Cavey, M.J. Development of the liver in the chicken embryo. II. Erythropoietic and granulopoietic cells. Anat. Rec. 1993, 235, 131–143. [Google Scholar] [CrossRef]
- Scanes, C.G. Avian Physiology: Are Birds Simply Feathered Mammals? Front. Physiol. 2020, 11, 542466. [Google Scholar] [CrossRef] [PubMed]
- Narbaitz, R.; Kacew, S. Ultrastructural and biochemical observations on the metanephros of normal and cultured chick embryos. Anat. Embryol. 1979, 155, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Wideman, R.F., Jr. Maturation of glomerular size distribution profiles in domestic fowl (Gallus gallus). J. Morphol. 1989, 201, 205–213. [Google Scholar] [CrossRef] [PubMed]
- Prince, N.; Oommen, V.; Bhaskar, A. Chicken intestine: An alternative to the mammalian intestine for physiology experimentation. Adv. Physiol. Educ. 2018, 42, 387–389. [Google Scholar] [CrossRef] [PubMed]
- Zaefarian, F.; Abdollahi, M.R.; Cowieson, A.; Ravindran, V. Avian Liver: The Forgotten Organ. Animals 2019, 9, 63. [Google Scholar] [CrossRef]
- Cook, M.E. Sturkie’s Avian Physiology. J. Wildl. Dis. 2017, 53, 703–705. [Google Scholar] [CrossRef]
- Hermier, D. Lipoprotein metabolism and fattening in poultry. J. Nutr. 1997, 127, 805s–808s. [Google Scholar] [CrossRef]
- Southwell, B.R. Staging of intestinal development in the chick embryo. Anat. Rec. A Discov. Mol. Cell Evol. Biol. 2006, 288, 909–920. [Google Scholar] [CrossRef]
- Nielsen, C.; Murtaugh, L.C.; Chyung, J.C.; Lassar, A.; Roberts, D.J. Gizzard formation and the role of Bapx1. Dev. Biol. 2001, 231, 164–174. [Google Scholar] [CrossRef]
- Bennett, T.; Cobb, J.L.S. Studies on the avian gizzard: The development of the gizzard and its innervation. Z. Zellforsch. Mikrosk. Anat. 1969, 98, 599–621. [Google Scholar] [CrossRef]
- Tokunaga, K.; Akimoto, S.; Aiba, M.; Nakagomi, M.; Suzuki, T.; Tatsumi, R.; Nakamura, M. Connexin 43 is Localized in Gizzard Smooth Muscle Cells during Chicken Development. J. Poult. Sci. 2022, 59, 338–347. [Google Scholar] [CrossRef]
- Stern, R.A.; Mozdziak, P.E. Differential ammonia metabolism and toxicity between avian and mammalian species, and effect of ammonia on skeletal muscle: A comparative review. J. Anim. Physiol. Anim. Nutr. 2019, 103, 774–785. [Google Scholar] [CrossRef]
- Fite, C.D. A Study of the Effects of Six Drugs on Chick Embryo Development. Proc. Okla. Acad. Sci. 1965, 45, 268–274. [Google Scholar]
- Whitsel, A.I.; Johnson, C.B.; Forehand, C.J. An in ovo chicken model to study the systemic and localized teratogenic effects of valproic acid. Teratology 2002, 66, 153–163. [Google Scholar] [CrossRef]
- Hsieh, C.L.; Chen, K.C.; Ding, C.Y.; Tsai, W.J.; Wu, J.F.; Peng, C.C. Valproic acid substantially downregulated genes folr1, IGF2R, RGS2, COL6A3, EDNRB, KLF6, and pax-3, N-acetylcysteine alleviated most of the induced gene alterations in chicken embryo model. Rom. J. Morphol. Embryol. 2013, 54, 993–1004. [Google Scholar]
- Turgut, U.; Kazan, S.; Cakin, H.; Ozak, A. Valproic acid effect on neural tube defects is not prevented by concomitant folic acid supplementation: Early chick embryo model pilot study. Int. J. Dev. Neurosci. 2019, 78, 45–48. [Google Scholar] [CrossRef]
- Zosen, D.; Austdal, L.P.E.; Bjørnstad, S.; Lumor, J.S.; Paulsen, R.E. Antiepileptic drugs lamotrigine and valproate differentially affect neuronal maturation in the developing chick embryo, yet with PAX6 as a potential common mediator. Neurotoxicology Teratol. 2022, 90, 107057. [Google Scholar] [CrossRef]
- Kimmel, C.A.; Young, J.F. Correlating pharmacokinetics and teratogenic endpoints. Fundam. Appl. Toxicol. 1983, 3, 250–255. [Google Scholar] [CrossRef]
- Nau, H. Species differences in pharmacokinetics and drug teratogenesis. Environ. Health Perspect. 1986, 70, 113–129. [Google Scholar] [CrossRef]
- Ghimire, S.; Zhang, X.; Zhang, J.; Wu, C. Use of Chicken Embryo Model in Toxicity Studies of Endocrine-Disrupting Chemicals and Nanoparticles. Chem. Res. Toxicol. 2022, 35, 550–568. [Google Scholar] [CrossRef]
- Kopańska, M.; Batoryna, M.; Banaś-Ząbczyk, A.; Błajda, J.; Lis, M.W. The Effect of α-Tocopherol on the Reduction of Inflammatory Processes and the Negative Effect of Acrylamide. Molecules 2022, 27, 965. [Google Scholar] [CrossRef] [PubMed]
- Samak, D.H.; El-Sayed, Y.S.; Shaheen, H.M.; El-Far, A.H.; Abd El-Hack, M.E.; Noreldin, A.E.; El-Naggar, K.; Abdelnour, S.A.; Saied, E.M.; El-Seedi, H.R.; et al. Developmental toxicity of carbon nanoparticles during embryogenesis in chicken. Environ. Sci. Pollut. Res. Int. 2020, 27, 19058–19072. [Google Scholar] [CrossRef] [PubMed]
- Ribatti, D. The chick embryo chorioallantoic membrane (CAM) assay. Reprod. Toxicol. 2017, 70, 97–101. [Google Scholar] [CrossRef]
- Zosen, D.; Hadera, M.G.; Lumor, J.S.; Andersen, J.M.; Paulsen, R.E. Chicken embryo as animal model to study drug distribution to the developing brain. J. Pharmacol. Toxicol. Methods 2021, 112, 107105. [Google Scholar] [CrossRef] [PubMed]
- Johnson, B.A.; Cheang, M.S.; Goldenberg, G.J. Comparison of Adriamycin Uptake in Chick Embryo Heart and Liver Cells and Murine L5178Y Lymphoblasts in Vitro: Role of Drug Uptake in Cardiotoxicity1. Cancer Res. 1986, 46, 218–223. [Google Scholar] [PubMed]
- Hornung, R.; Hammer-Wilson, M.J.; Kimel, S.; Liaw, L.-H.; Tadir, Y.; Berns, M.W. Systemic application of photosensitizers in the chick chorioallantoic membrane (CAM) model: Photodynamic response of CAM vessels and 5-aminolevulinic acid uptake kinetics by transplantable tumors. J. Photochem. Photobiol. B Biol. 1999, 49, 41–49. [Google Scholar] [CrossRef][Green Version]
- Fonseca, B.B.; da Silva, M.V.; de Morais Ribeiro, L.N. The chicken embryo as an in vivo experimental model for drug testing: Advantages and limitations. Lab. Anim. 2021, 50, 138–139. [Google Scholar] [CrossRef]
- Khosravi, A.; Sharifi, I.; Tavakkoli, H.; Derakhshanfar, A.; Keyhani, A.R.; Salari, Z.; Mosallanejad, S.S.; Bamorovat, M. Embryonic toxico-pathological effects of meglumine antimoniate using a chick embryo model. PloS ONE 2018, 13, e0196424. [Google Scholar] [CrossRef]
- Syrovets, T.; Gschwend, J.E.; Büchele, B.; Laumonnier, Y.; Zugmaier, W.; Genze, F.; Simmet, T. Inhibition of IkappaB kinase activity by acetyl-boswellic acids promotes apoptosis in androgen-independent PC-3 prostate cancer cells in vitro and in vivo. J. Biol. Chem. 2005, 280, 6170–6180. [Google Scholar] [CrossRef]
- Vargas, A.; Zeisser-Labouèbe, M.; Lange, N.; Gurny, R.; Delie, F. The chick embryo and its chorioallantoic membrane (CAM) for the in vivo evaluation of drug delivery systems. Adv. Drug Deliv. Rev. 2007, 59, 1162–1176. [Google Scholar] [CrossRef]
- Yadav, A.; Verhaegen, S.; Hadera, M.G.; Berntsen, H.F.; Berg, V.; Lyche, J.L.; Sabaredzovic, A.; Haug, L.S.; Myhre, O.; Zimmer, K.E.; et al. Peripherally administered persistent organic pollutants distribute to the brain of developing chicken embryo in concentrations relevant for human exposure. NeuroToxicology 2022, 88, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Rousset, X.; Maillet, D.; Grolleau, E.; Barthelemy, D.; Calattini, S.; Brevet, M.; Balandier, J.; Raffin, M.; Geiguer, F.; Garcia, J.; et al. Embryonated Chicken Tumor Xenografts Derived from Circulating Tumor Cells as a Relevant Model to Study Metastatic Dissemination: A Proof of Concept. Cancers 2022, 14, 4085. [Google Scholar] [CrossRef] [PubMed]
- Shoin, K.; Yamashita, J.; Enkaku, F.; Sasaki, T.; Tanaka, M.; Yoshio, Y. Chick Embryo Assay as Chemosensitivity Test for Malignant Glioma. Jpn. J. Cancer Res. 1991, 82, 1165–1170. [Google Scholar] [CrossRef] [PubMed]
- Lodhia, K.A.; Hadley, A.M.; Haluska, P.; Scott, C.L. Prioritizing therapeutic targets using patient-derived xenograft models. Biochim. Biophys. Acta 2015, 1855, 223–234. [Google Scholar] [CrossRef]
- American Veterinary Medical Association. AVMA Guidelines for the Euthanasia of Animals, 2020th ed.; American Veterinary Medical Association: Schaumburg, IL, USA, 2020. [Google Scholar]
- Lierz, M.; Korbel, R. Anesthesia and Analgesia in Birds. J. Exot. Pet. Med. 2012, 21, 44–58. [Google Scholar] [CrossRef]
- Rosenbruch, M. The sensitivity of chicken embryos in incubated eggs. Altex 1997, 14, 111–113. [Google Scholar]
- Ribatti, D. The Chick Embryo Chorioallantoic Membrane as an In Vivo Assay to Study Antiangiogenesis. Pharmaceuticals 2010, 3, 482–513. [Google Scholar] [CrossRef]
- FDA Wound Healing Clinical Focus Groupa. Guidance for industry: Chronic cutaneous ulcer and burn wounds-developing products for treatment. Wound Repair Regen. 2001, 9, 258–268. [Google Scholar] [CrossRef]
- Cimpean, A.M.; Ribatti, D.; Raica, M. The chick embryo chorioallantoic membrane as a model to study tumor metastasis. Angiogenesis 2008, 11, 311–319. [Google Scholar] [CrossRef]
- Deryugina, E.I.; Quigley, J.P. Chick embryo chorioallantoic membrane model systems to study and visualize human tumor cell metastasis. Histochem. Cell Biol. 2008, 130, 1119–1130. [Google Scholar] [CrossRef]
- Nigri, M.; Åhlgren, J.; Wolfer, D.P.; Voikar, V. Role of Environment and Experimenter in Reproducibility of Behavioral Studies With Laboratory Mice. Front. Behav. Neurosci. 2022, 16, 835444. [Google Scholar] [CrossRef] [PubMed]
- Voelkl, B.; Altman, N.S.; Forsman, A.; Forstmeier, W.; Gurevitch, J.; Jaric, I.; Karp, N.A.; Kas, M.J.; Schielzeth, H.; Van de Casteele, T.; et al. Author Correction: Reproducibility of animal research in light of biological variation. Nat. Rev. Neurosci. 2020, 21, 394. [Google Scholar] [CrossRef] [PubMed]
- Komatsu, A.; Higashi, Y.; Matsumoto, K. Various CAM tumor models. Enzymes 2019, 46, 37–57. [Google Scholar] [CrossRef] [PubMed]
- Inovotion. Available online: https://www.inovotion.com/ (accessed on 23 May 2022).




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fischer, D.; Fluegen, G.; Garcia, P.; Ghaffari-Tabrizi-Wizsy, N.; Gribaldo, L.; Huang, R.Y.-J.; Rasche, V.; Ribatti, D.; Rousset, X.; Pinto, M.T.; et al. The CAM Model—Q&A with Experts. Cancers 2023, 15, 191. https://doi.org/10.3390/cancers15010191
Fischer D, Fluegen G, Garcia P, Ghaffari-Tabrizi-Wizsy N, Gribaldo L, Huang RY-J, Rasche V, Ribatti D, Rousset X, Pinto MT, et al. The CAM Model—Q&A with Experts. Cancers. 2023; 15(1):191. https://doi.org/10.3390/cancers15010191
Chicago/Turabian StyleFischer, Dagmar, Georg Fluegen, Paul Garcia, Nassim Ghaffari-Tabrizi-Wizsy, Laura Gribaldo, Ruby Yun-Ju Huang, Volker Rasche, Domenico Ribatti, Xavier Rousset, Marta Texeira Pinto, and et al. 2023. "The CAM Model—Q&A with Experts" Cancers 15, no. 1: 191. https://doi.org/10.3390/cancers15010191
APA StyleFischer, D., Fluegen, G., Garcia, P., Ghaffari-Tabrizi-Wizsy, N., Gribaldo, L., Huang, R. Y.-J., Rasche, V., Ribatti, D., Rousset, X., Pinto, M. T., Viallet, J., Wang, Y., & Schneider-Stock, R. (2023). The CAM Model—Q&A with Experts. Cancers, 15(1), 191. https://doi.org/10.3390/cancers15010191