1. Introduction
In 2020, there have been an estimated 2,261,419 new cases (11.7% of all cancer types), and 684,996 mortalities (6.9% of all cancer types) caused by breast cancer (BC), globally. BC, among other cancers, remains the leading cause of cancer-related mortality among women [
1]. BC is divided mainly into two types, invasive and non-invasive which is based on their spreading ability to the different organs. Invasive BC is more common and is further subdivided into invasive ductal carcinoma and invasive lobular carcinoma based on the origin of the tumor. BC is also divided into triple-negative breast cancer (TNBC), luminal (A and B), and human epidermal growth factor receptor (HER2) based on the status of the hormonal receptor. Among these, TNBC is highly aggressive with absence of estrogen receptor (ER), progesterone receptor (PR), and HER2 on the tumor [
2]. Depending upon the stage of cancer, treatment modalities, either alone or in combination, include surgical removal of the tumor, radiation therapy, chemotherapy, and hormonal therapy. Among these, chemotherapy is considered the first-line treatment choice [
3] even after having severe dose-dependent systemic toxicity and a lack of specificity [
4]. Tremendous efforts have been made to overcome these limitations either by adopting the alternative route of administration or site-specific delivery strategies for the existing chemo drugs.
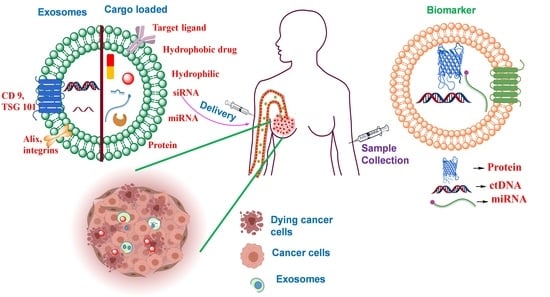
For the past decade, extensive research has been carried out in the area of nanotechnology and targeted therapy. Some of these candidates were approved for clinics based upon the risk over benefit ratio for clinically available formulations. However, high cost, poor specificity, and elimination by the reticuloendothelial system (RES) are some of the limitations that narrowed down the wide application of nanotechnology [
5]. To further improve the efficacy, and overcome the present limitations, various novel nano-carriers have emerged; extracellular vesicles (EVs) are one of them. EVs are nano-sized, bilayer membrane-bound vesicles that are naturally secreted by the cells into the extracellular environment. The EVs are further divided into three different types based on their molecular genesis and composition; (i) apoptotic bodies, (ii) exosomes, and (iii) microvesicles, which are naturally loaded with a variety of cargos and rich in disease-specific biomarkers [
6]].
The apoptotic bodies originate from the plasma membrane at the time of the cell death process, i.e., apoptosis. They are heterogeneous-shaped vesicles in the size range of 50–5000 nm. Apoptotic bodies contain DNA, RNA, histone, cell signaling molecules (messengers), nuclear fractions, and cell organelles. Apoptotic bodies display high expression of Annexin V (biomarker for apoptotic bodies) in their membrane [
7]. On the other hand, the microvesicles, which are also known as exovesicles, ectosomes, shedding vehicles, microparticles, and plasma membrane-originated vesicles, either originate from the outward budding of the plasma membrane or by the fusion of the plasma membrane. The size of microvesicles ranges from 50 to 1000 nm which may contain a variety of cargos,
viz., miRNA, mRNA, other non-coding RNA, receptor proteins, membrane protein, and cytoplasmic protein [
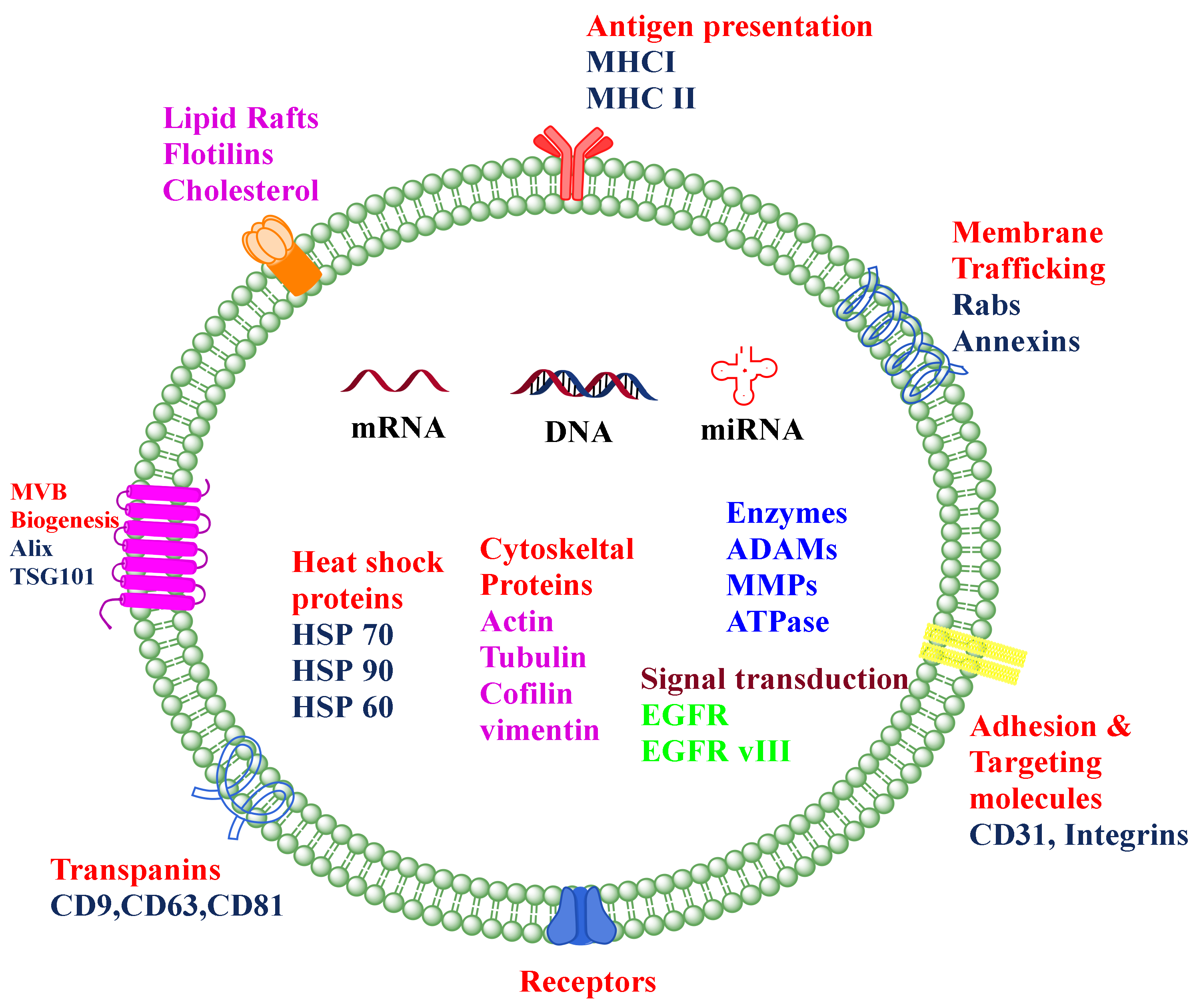
8]. Exosomes originate from multivesicular bodies (MVB) and are formed by the inward budding of late endosomes. The representative diagram of a typical exosome is depicted in
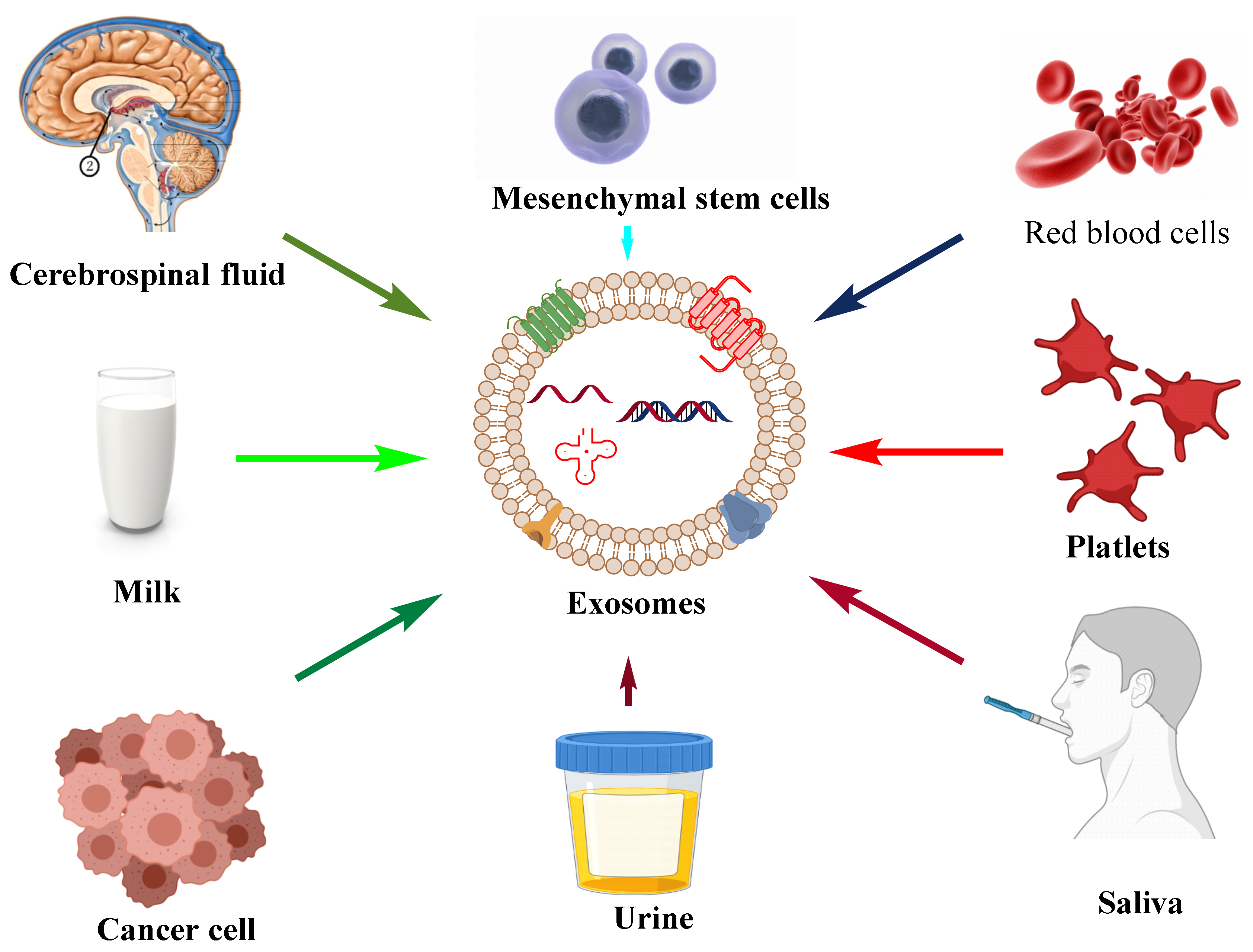
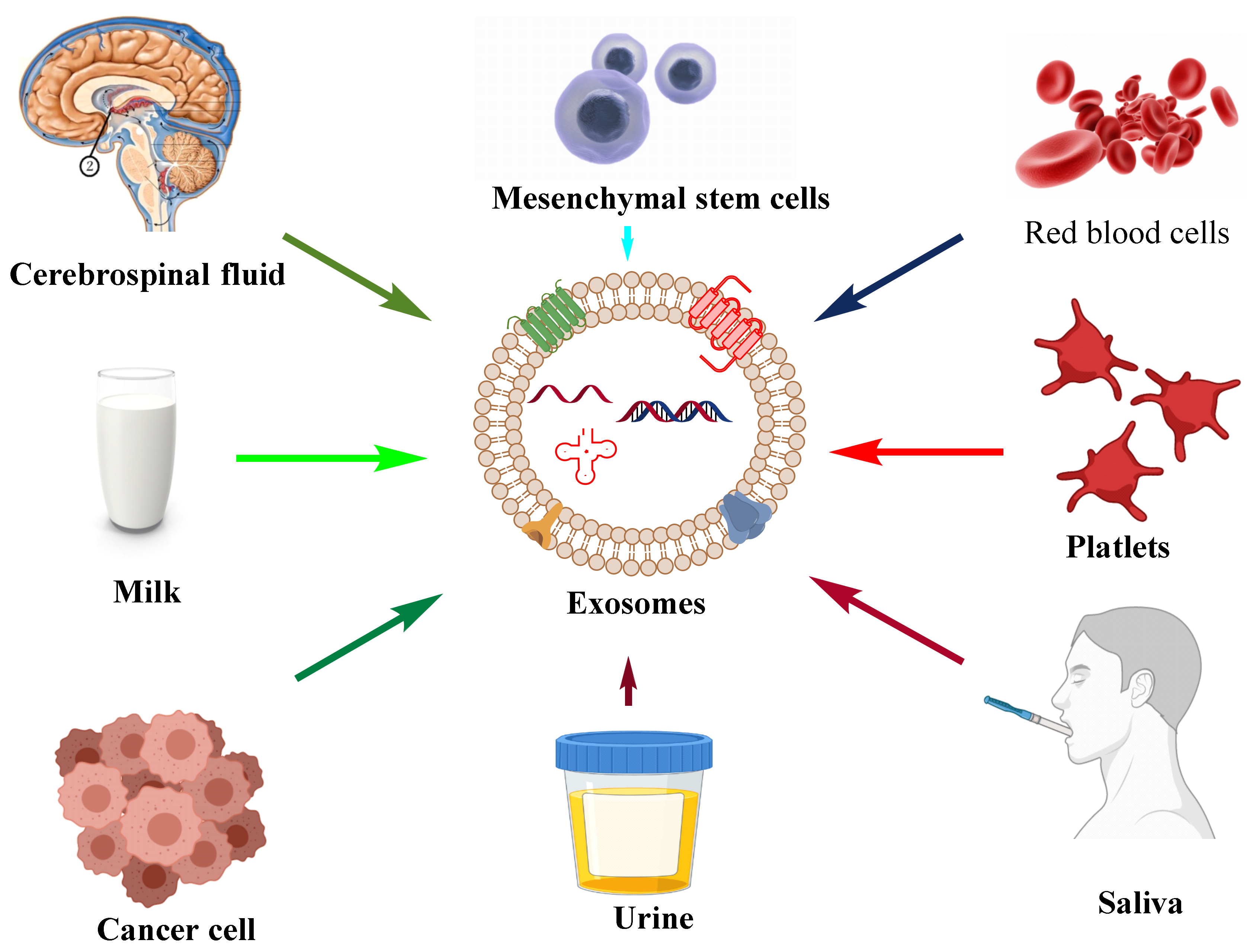
Figure 1. Their size ranges between 30 and 150 nm and is secreted by various biological fluids such as blood, tears, saliva, cerebrospinal fluid, and milk. They contain mRNA, miRNA, other non-coding RNA, immune protein (major histocompatibility complex (MHC)), cytoplasmic, receptor, and membrane proteins [
9,
10]. The comparison of different types of EVs and different sources of exosomes are represented in
Table 1 and
Figure 2, respectively.
The growing interest in EVs, especially on exosomes, is not only represented by the increasing number of scientific works on exosomes but also by the increasing number of scientific association portals and databases such as
- 1.
ISEV: The International Society for Extracellular Vesicles.
- 2.
EU ME-HaD: the European Network on Microvesicles and Exosomes in Health and Disease.
- 3.
Vesiclepedia.
- 4.
ExoCarta created in 2009 as an open access resource for compiling proteins and RNAs identified in exosomes (referenced in ‘Biochemistry of Exosomes’).
The present review, hence, explores the recent developments in the exosome technology more specifically to their use in breast cancer diagnosis and drug delivery. The review starts with the biogenesis of exosomes and further discusses the various exosomal approaches that have been recently investigated in this domain. Selection of different drug loading techniques either before or after the isolation of exosomes has been discussed in detail. The review also includes in-depth discussion about the concept of exosome surface functionalization by different targeting ligands and its application for obtaining effective diagnosis and treatment for breast cancer. Ongoing clinical trials, in the subject area, have also been included to gain a better understanding of the current status of the exosome technology. Overall, the prime target of the current manuscript is to provide the state-of-the-art involved in the field of exosomes and their advancements in the diagnosis and drug delivery in breast cancer.
2. Biogenesis of Exosomes
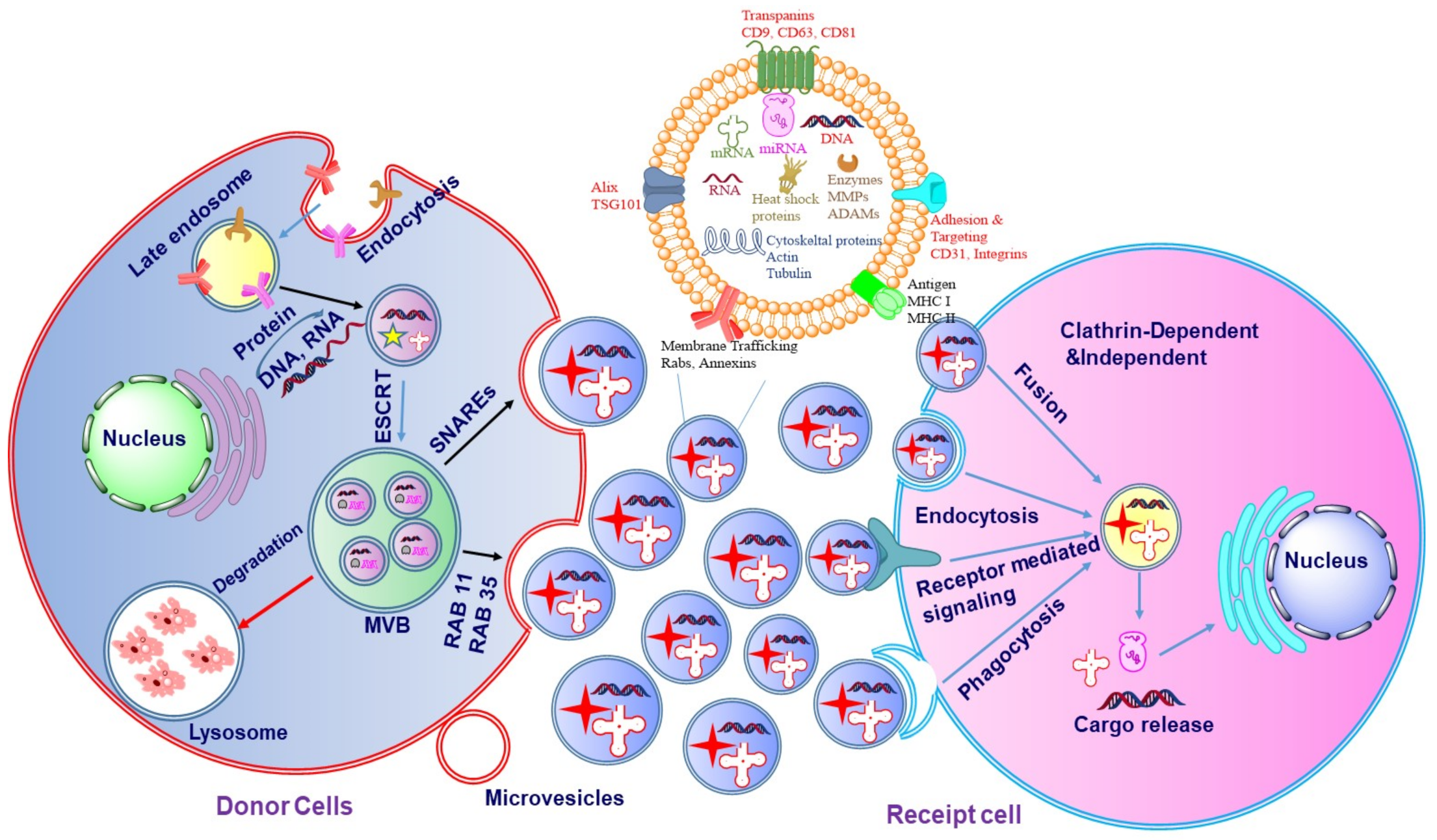
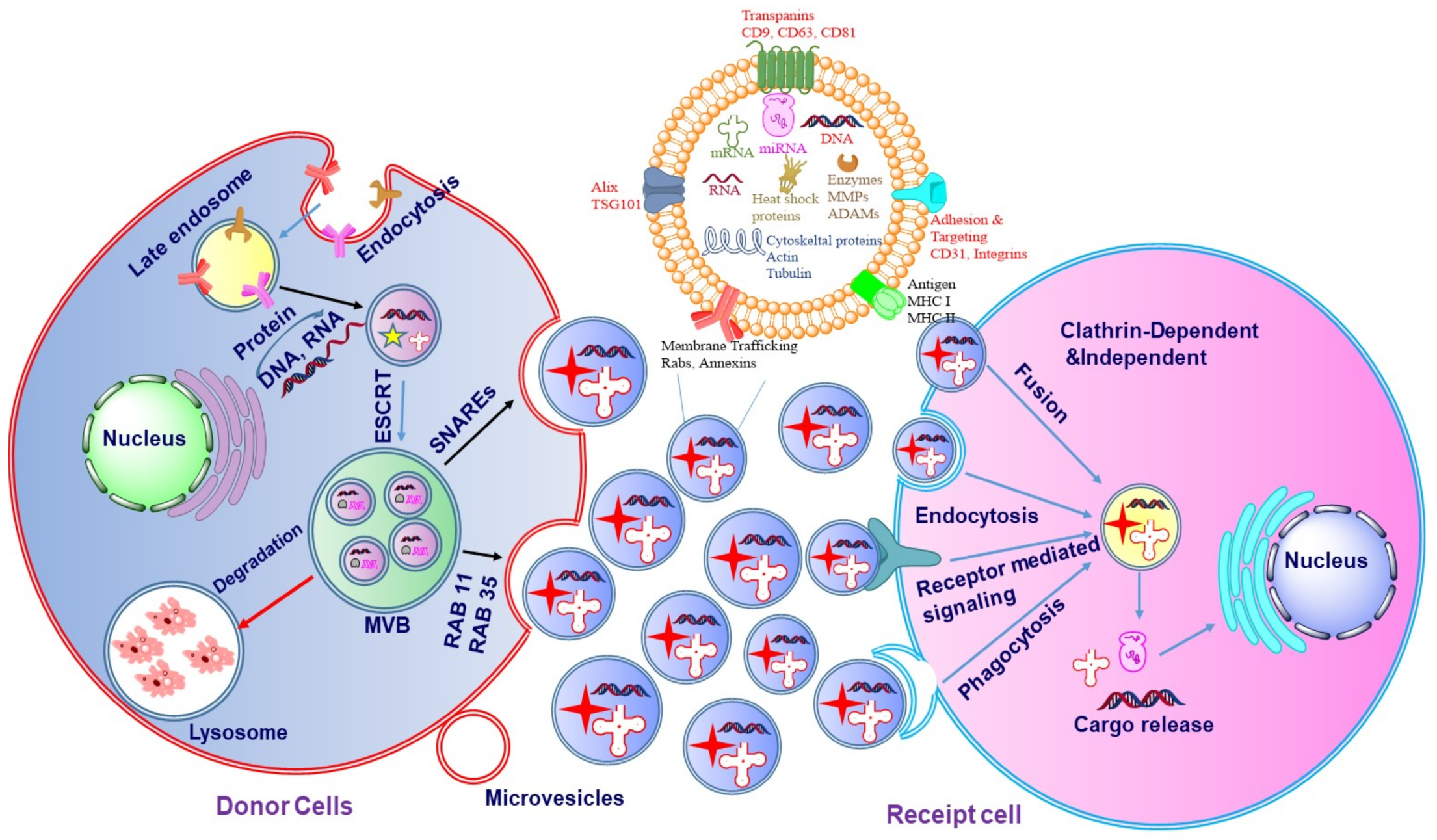
The biogenesis of exosomes may be divided into three major stages (
Figure 3): (i) formation of the endocytic vesicle, (ii) formation of multivesicular bodies (MVBs) which comprise intraluminal vesicles (ILVs), and (iii) the fusion of these MVBs with the plasma membrane [
11,
12,
13]. The exosomes can be secreted both in a healthy state and disease state by a wide array of cell types [
14]. During the process of biogenesis, the endocytic vesicle formed from the plasma membrane is first developed into an early endosome and then into late endosome. The limiting membranes of the late endosomes undergo invagination to produce ILVs within the lumen of organelles [
15]. The assembling of ILVs into late endosomes leads to the formation of MVBs. The formation of endocyclic vesicles is controlled by specific proteins that belong to the Rab family. The formation of endosomal vesicles occurs via two pathways namely, endosomal sorting complexes required for the transport (ESCRT) dependent pathway and ESCRT-independent pathway. The Rab family proteins include four ESCRT proteins namely ESCRT-0, ESCRT-I, ESCRT-II, ESCRT-III. Apart from the Rab proteins, the other two proteins associated with the transition of endosomes to exosomes include TSG101 and ALIX [
16,
17,
18]. Such a process needs ubiquitination of the cytosolic tail of the endocytosed receptors.
TSG101 belongs to the ESCRT-I protein and frames a complex with the ubiquitinated proteins that activate the ESCRT-II protein and promote the oligomerization and formation of the ESCRT-III complex. This complex then gets involved in the sequestration of MVB proteins and the enrollment of the de-ubiquitination catalyst to eliminate the ubiquitin from the load proteins before arranging them into ILVs. In the last stage of exosome formation, the ESCRT-III complex is degraded by vacuolar protein sorting associate protein 4 (VPS4) and ATPase [
19,
20]. The schematic diagram of the biogenesis of exosomes is represented in
Figure 3. In ESCRT-independent biogenesis of exosomes, ceramide plays an important role which is synthesized by sphingomylinase enzyme [
21]. Inhibition of this sphingomyelinase enzyme reduces the secretion of exosomes in Oli-neu cells [
12].
Some recent studies have shown that inhibition of these complex proteins (ESCRT, TSG101, and ALIX) resulted in a decrease in exosomal secretion. Four separate investigations have shown that inhibiting Hrs numbers of ESCRT-0 resulted in a decrease in exosomal secretion in different cell types which includes cervical cancer cells (HeLa)—a lineage of cancer human epithelial cells, human embryonic kidney cell line 293 (HEK293), mouse dendritic cells, squamous carcinoma cell line [
22,
23,
24]. The in vivo exosomal secretions are also decreased by the presence of certain drugs such as calpeptin, manumycin, imipramine, etc. Calpeptin is a calcium-dependent neutral cytosolic cysteine protease that inhibits calpains which plays a role in the production of multivesicular bodies [
25]. Manumycine is an anti-bacterial agent which inhibits the small GTPases Ras enzyme in F11, mouse neuroblastoma hybrid cells, and rat dorsal root ganglion cells [
26], and imipramine is a tri-cyclic anti-depressant which inhibits the acid sphingomyelinase (aSMase) both in vitro and in vivo [
27,
28]. Depletion of TSG101 also reduces the secretion of exosomes from immortalized retinal pigment epithelial cells 1 (RPE1) [
22].
3. Isolation of Exosomes
Several approaches have been reported in the literature for the isolation of EVs, viz., (1) ultra-centrifugation, (2) size-exclusion chromatography, (3) precipitation, (4) ultra-filtration, and (5) immunoaffinity. Every method has its benefits and drawbacks, however, based upon the exosome source and purpose, the method can be adopted for the isolation of exosomes.
3.1. Centrifugation
There are two types of centrifugation methods used for the isolation of exosomes: (1) differential ultra-centrifugation, separation based on the size of the different components and (2) density gradient ultra-centrifugation based on their densities.
3.1.1. Differential Ultra-Centrifugation
In differential ultracentrifugation, exosomes are isolated by serial centrifugation at different times and speeds. Johnstone et al., 1992 first used the differential ultracentrifugation method to separate exosomes from the tissue culture medium [
29], which was further upgraded by Thery et al., 2006 [
30]. According to Thery et al., cell culture supernatant underwent serial centrifugations at 300×
g and 2000×
g for each cycle of 10 min, and 10,000×
g for 30 min to remove the live cells, dead cells, and cell debris, respectively. The final supernatant obtained was ultra-centrifuged at 100,000×
g for 70 min to obtain the pellets containing exosomes and proteins which were washed with PBS to procure pure exosomes.
Although, various research groups have adopted ultracentrifugation-based exosomes isolation, however, isolation via differential centrifugation method does not yield a large number of pure exosomes which limits its utilization. Therefore, it is advised to either optimize the existing technique or we need to find an alternative method of isolation that will overcome the problems associated with the differential ultracentrifugation method. This may include the optimization of different process variables (number of steps with variable speeds) for obtaining the pure population of exosomes.
3.1.2. Density Gradient Ultra-Centrifugation
In the density gradient ultracentrifugation (DG centrifugation) method, similar steps are being followed as the differential ultracentrifugation method except for the last stage where the separation/purification is completed in high-density sugar solution or iodixanol which leads to the collection of pure exosomes at the middle layers of the test tube.
Yamada et al., 2012, isolated exosomes from bovine milk via ultra-centrifugation, the whey from 100 mL of milk was ultra-centrifuged at 100,000×
g for 60 min, after which the pellets were collected and re-suspended in 1 mL of fresh chilled PBS. The EVs in suspension were segmented on a linear sucrose density gradient (10–40%) solution (9 mL) and were again ultra-centrifuged at 200,000×
g for 18 h. Fractions (0.9 mL) were collected from the top of the centrifuge tube and categorized from 1 to 10, where the fraction numbered 7 was confirmed as exosomes by using the Western blot analysis. Fractions numbered 7–9 were diluted with chilled PBS and again ultra-centrifuged at 100,000×
g for 60 min to obtain pure exosomes which were further characterized by electron microscope [
31]. However, DG centrifugation is a more convenient method to isolate exosomes at high yield and purity, although it consumes more time and is a high equipment-dependent method which limits its usefulness.
3.2. Chromatography-Based Isolation Method
In this method, exosomes are separated based on size differences between the EVs in biological samples by using size-exclusion chromatography (SEC) which is also known as gel filtration or molecular sieve chromatography. The source material of the EVs is loaded on the column packed with Sepharose©, Sephacryl©, or BioGel P, as a stationary phase. Thus, the elution is based upon their size in the order of decreasing molecular weight [
32]. The commercially available columns which are employed to isolate the exosomes include qEV separation columns, EV Second purification columns, Exo Cap™, and Exo-spin. Compared to ultracentrifugation and ultrafiltration, the chromatography-based isolation method is quick and of relatively low cost. Additionally, the isolated fractions are uniform in size and their biological characteristics remain significantly unaffected by this method. However, there are some chances where exosomal fractions may get contaminated with other particles of the same size which may compromise the purity of exosomes. Nevertheless, owing to its time-consuming process, the SEC method limits its use in treatment and research.
A comparative study was performed by Gamez-Valero et al., 2016 [
33], where the exosomes were isolated from plasma by SEC using Sepharose
® CL-2B as a stationary column, followed by precipitation with PEG, and the PROSPR method. Exosomes from the SEC method may contain a lower amount of contaminated proteins with the highest quantity of exosomal biomarkers such as CD9, CD63, CD81, and CD5L [
33].
3.3. Ultrafiltration-Based Isolation
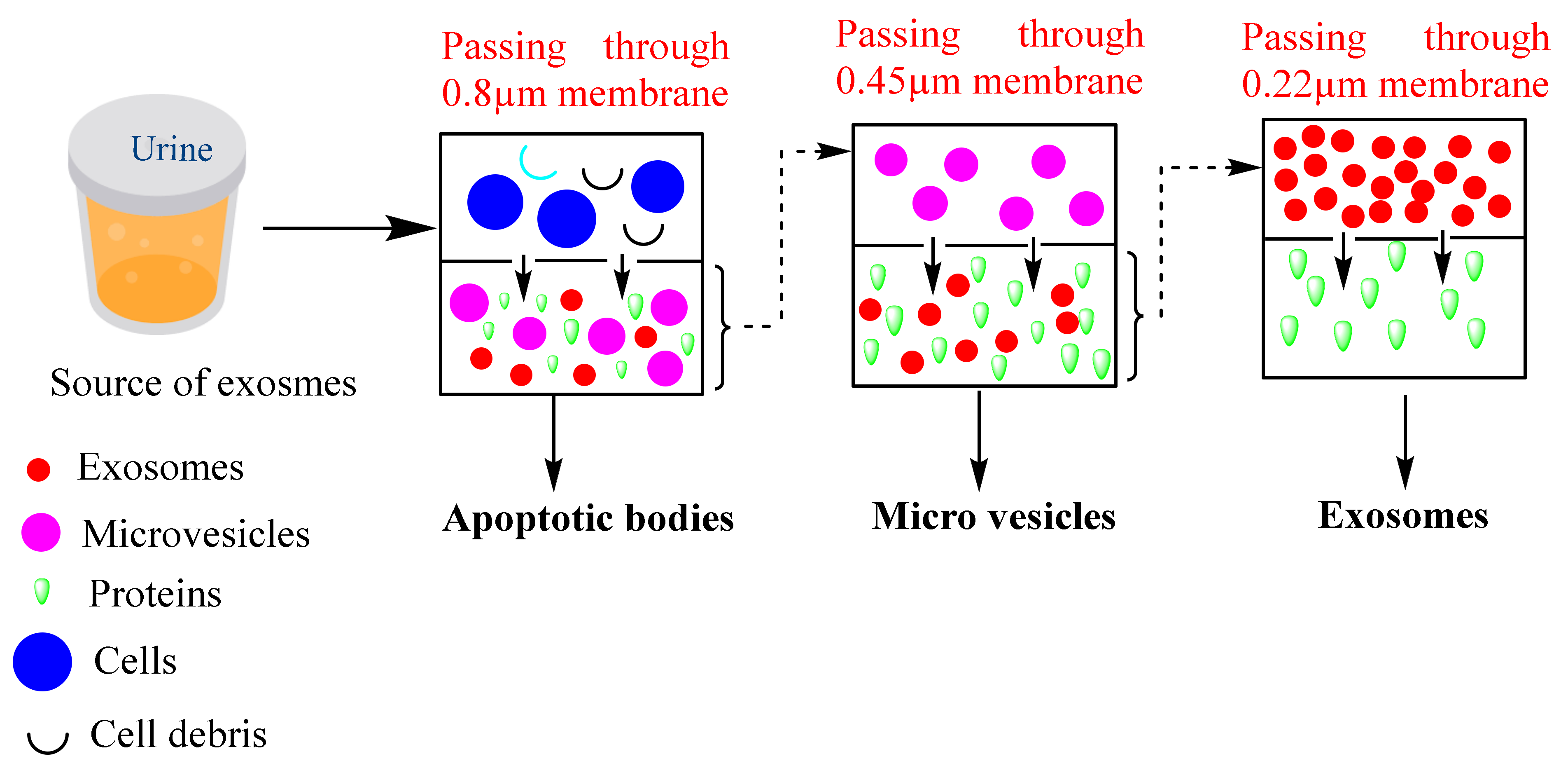
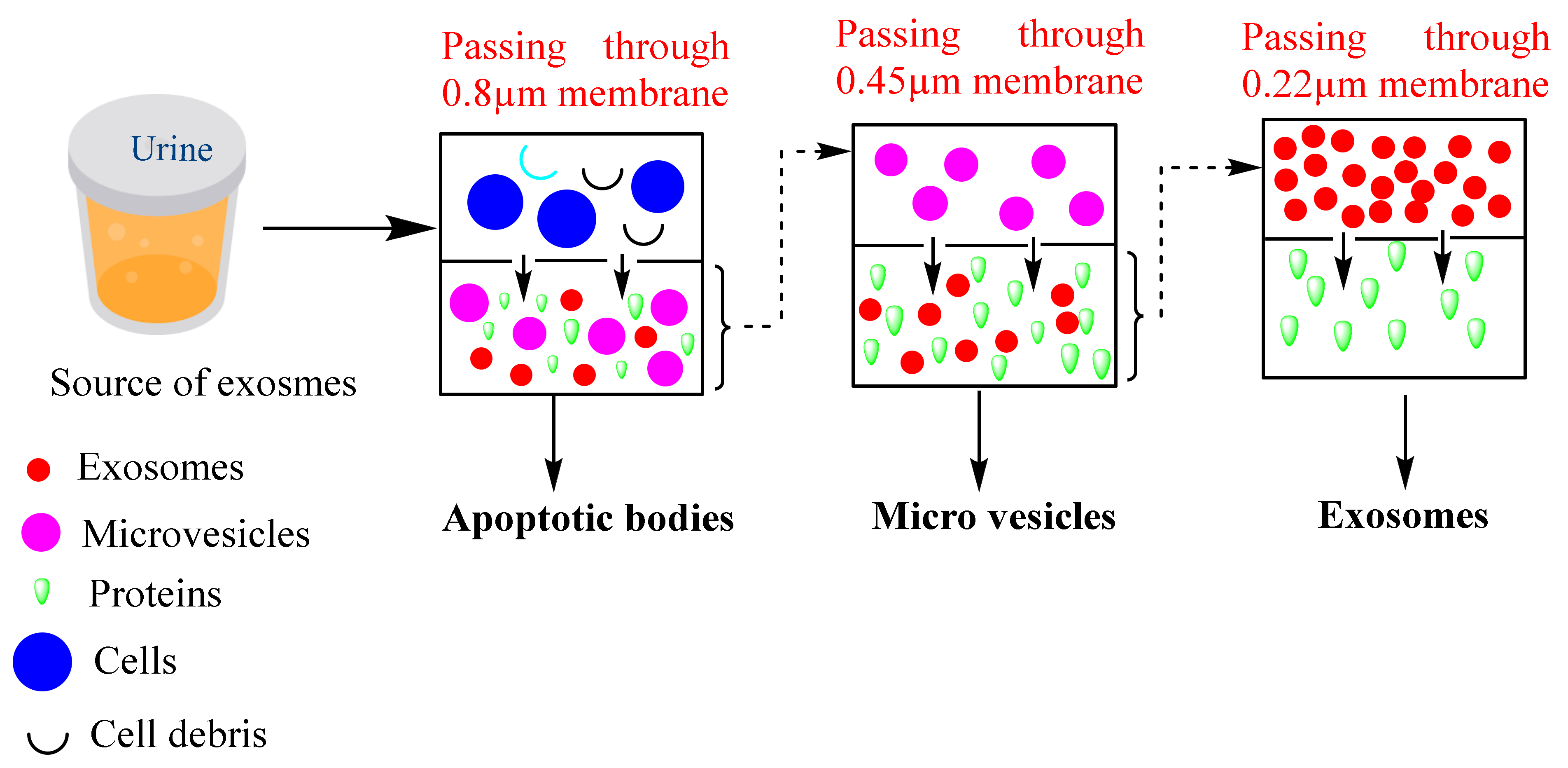
Ultrafiltration-based isolation of exosomes depends upon the separation of the different classes of EVs such as apoptotic bodies, microvesicles, exosomes, and other contamination proteins based on their different sizes. The filtration-based isolation method is classified into three types,
viz., ultrafiltration, hydrostatic dialysis, and gel filtration. In the ultrafiltration method, the ultrafiltration membranes are used to isolate exosomes that are available in different pore sizes such as 0.8, 0.45, 0.22, and 0.1 µm capable of retaining the particles with diameters of 800, 450, 220, and 100 nm, respectively. Larger particles such as apoptotic bodies are separated first followed by microvesicles and exosomes [
34]. A schematic diagram of the filtration-based method is represented in
Figure 4. Based on this method, Cheruvanky et al., 2007 have isolated exosomes from urine. They utilized ultrafiltration cells with nanomembranes which exhausted the protein with a molecular weight greater than 100 kDa, then centrifugation at 3000×
g. Such a method does not require any further ultracentrifugation [
35]. Recently a comparative study showed that the yield of the exosomes is greater in the ultrafiltration method compared to the ultracentrifugation method. Moreover, in the ultrafiltration method, the size of the exosomes is below 100 nm which indicates that the ultrafiltration method is most suitable for isolating exosomes from large sample sizes [
36].
3.4. Precipitation-Based Isolation
The precipitation-based isolation method is widely used for the isolation of exosomes. As per an earlier report, precipitation-based strategies are utilized particularly for biologics having a low initial volume. It was observed that 84% of scientists utilize the precipitation method for investigating the encapsulation of RNA in EVs [
37]. In the precipitation method, polymers such as polyethylene glycol (PEG) [
38], salt solutions such as sodium acetate [
39], organic salts, and a charge-based moiety such as protamine are used. The exosomes are first incubated with a precipitating agent for 1–24 h depending on the nature and concentration of the precipitating agent, followed by low-speed centrifugation [
38]. With the emergence of the requirement for exosomes’ isolation, several biotech companies are developing commercial kits (
Table 2) for the precipitation-based isolation of exosomes. In the protein organic solvent precipitation (PROSPR) method, solvents such as acetone, chloroform, glacial acetic acid, and trichloroacetic acid are used to precipitate the solubilizing proteins. Somiya et al., 2018 isolated exosomes from cow milk based on the PROSPR method [
40]. In the charge-based precipitation technique, protamine sulfate is used for separation. Protamine sulfate is a positively charged molecule that interacts with the negatively charged EVs and forms a precipitate that is separated by low-speed multiple centrifugations [
41]. The precipitation-based isolation method is the most promising isolation method for clinical use due to the requirement of lower sample quantity, less time consuming, and absence of exosomal damage. However, the precipitation method is facing the problems of contamination with other exosomal proteins such as albumin and PEG which could lead to the toxicity [
42]. Nevertheless, to minimize the contamination, pre-filtration through 0.22 um or post-precipitation–purification steps such as filtration and centrifugation were performed to reduce the contamination with impurities [
43].
3.5. Immunoaffinity-Based Isolation
Immunoaffinity is a separation and purification method based on certain interactions between antibodies and antigens to isolate the required component from a mixture. This is the most promising method for the isolation of specific exosomes. The commonly used antibodies in immunoaffinity-based isolation are monoclonal antibodies against specific biomarkers: CD9, CD63, CD80, and TSG101 for exosomes; integrins and CD40 for microvesicles, and annexin V for apoptotic bodies. These antibodies are fixed on different types of materials such as magnetic beads or microfluidic devices. Nakai et al., 2016 isolated exosomes from mouse peritoneal macrophages cells. For this, Tim4-FC protein was used to bind with phosphatidylserine present on the surface of the exosomes which were then captured by using magnetic beads [
44]. However, immunoaffinity-based isolation is not suitable for isolating a large number of exosomes. Moreover, the high cost and low yield of exosomes limit their usefulness.
Patel et al., 2019, and Tang et al., 2017, isolated the exosomes using a different method and observed that exosomes isolated via the UC method showed higher purity as determined by particle and protein ratio, and exosomal biomarker (CD9) as compared to SEC [
45,
46]. However, further studies are indispensable to determine the purity of exosomes from different isolation methods.
The advantages and disadvantages of different isolation methods are discussed in (
Table 3).
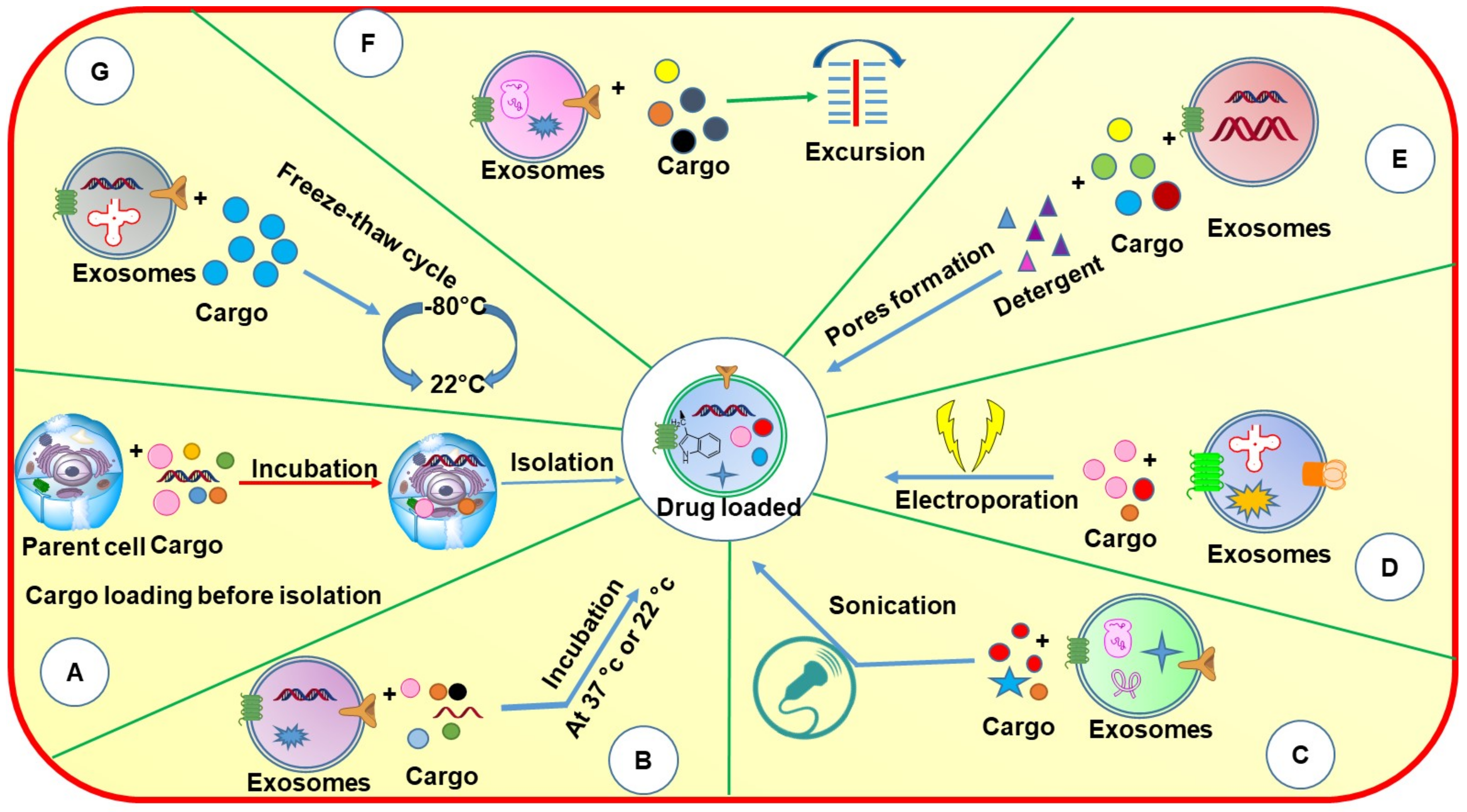
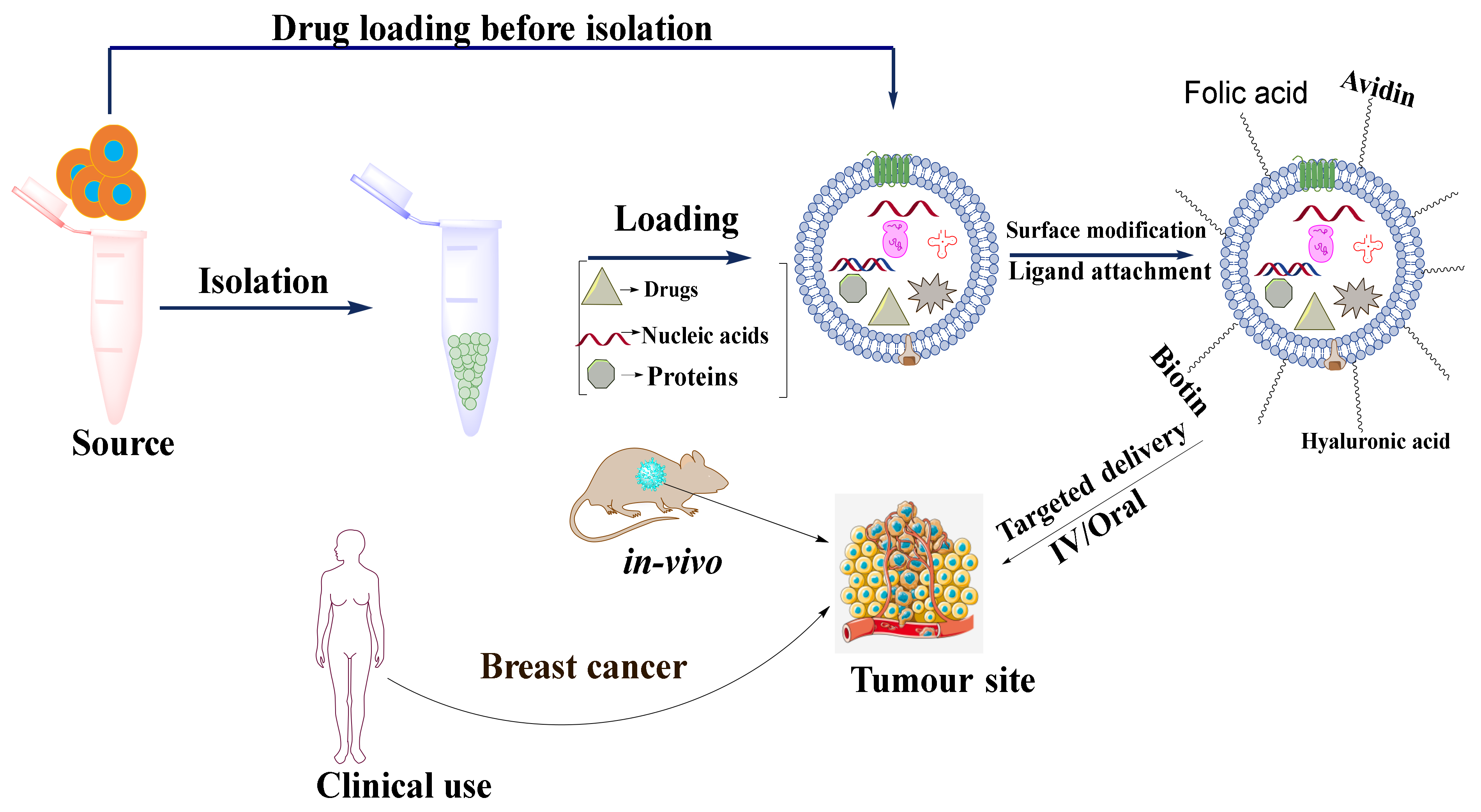
4. Drug Loading
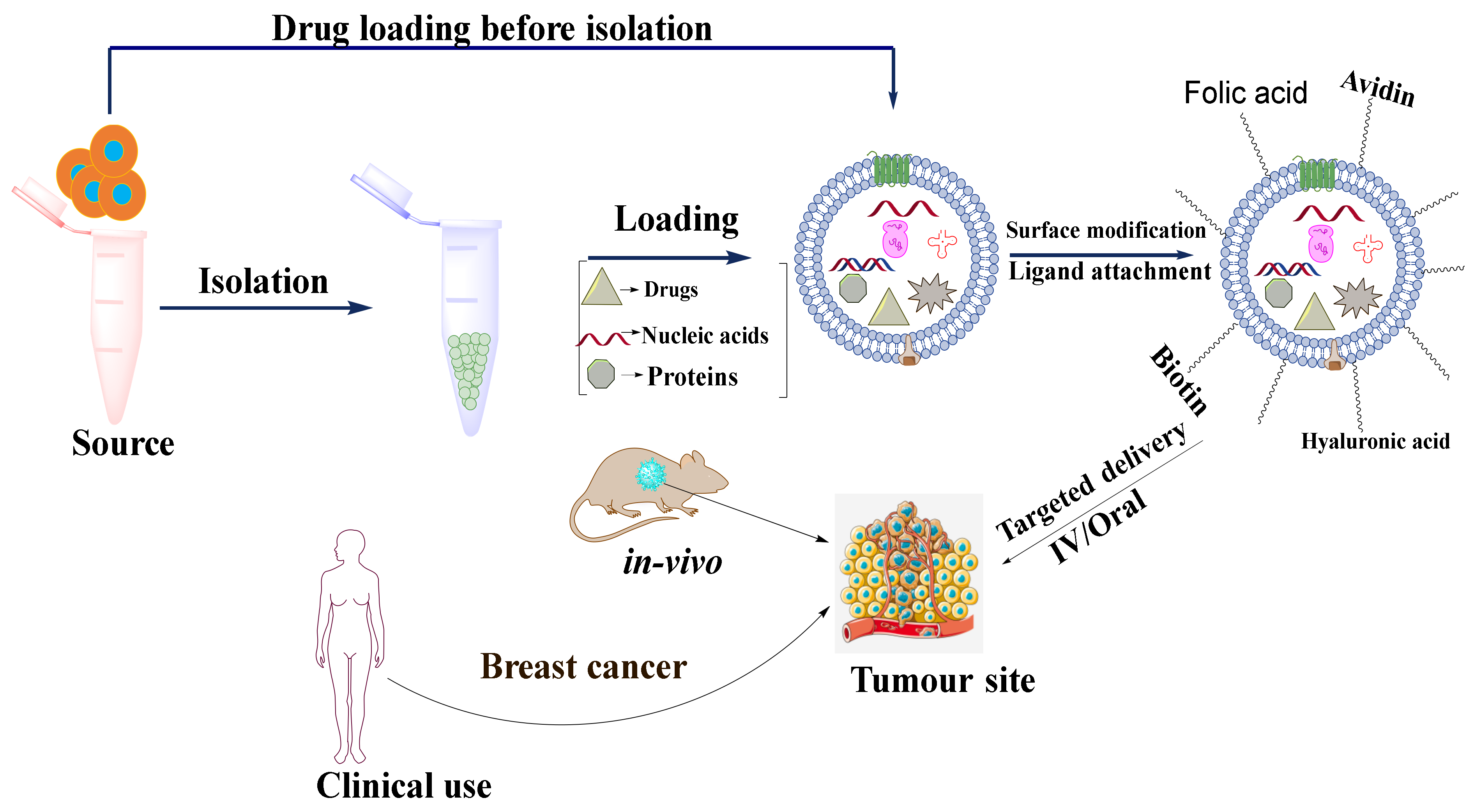
Exosomes can be used as nanocargos that are capable of loading different small and large molecules. They can load drugs either before or after isolation depending on the application (
Figure 5). Each of these loading methods has its pros and cons (
Table 4).
Drug loading before isolation of exosomes can be performed in two ways: (1) incubation of parent cells with a drug and (2) by gene editing. In the incubation method of drug loading, the compounds are directly mixed with the cell culture medium, where the drugs get internalized into cells and are loaded into EVs via an endogenous mechanism. Tang et al., 2012 incubated mouse hepatocarcinoma tumor cells (H22), Raw264.7 cells, human ovarian cancer cells (A2780, B16, HL60, and EL4) with four different drugs namely methotrexate (MTX), doxorubicin (DOX), cisplatin, and hydroxyl camptothecin for 12 h. MTX was incubated with H22 cell lines at different concentrations (1–10 μg mL
−1) and then ultracentrifuged to obtain microparticles which were again incubated with the same cell lines for 48–72 h at different concentrations. It was observed that both the high concentration and low concentration derived microparticles induced cell death, but the high dose derived microparticles resulted in microparticles packaged with a large amount of drugs, as compared to low dose derived microparticles [
47]. In another study, Pascucci and colleagues incubated paclitaxel (PTX) with SR4987 cells (bone marrow mesenchymal stromal cells) at low concentrations (100 µg mL
−1) for 24 h. After 48 h, exosomes were collected from the culture medium by differential centrifugation. The isolated PTX-exosomes showed significant anti-proliferative activity against CFPAC-1 cell lines (human pancreatic adenocarcinoma cells) [
48]. Interestingly, the exosomes devoid of paclitaxel also showed a minor anti-proliferative activity, due to the presence of some nucleic acids and proteins that may change the tumor environment.
4.1. Drug Loading after Isolation of Exosomes
Several methods have been used for drug loading after the isolation of exosomes. The drug loading can be achieved by incubation, sonication, detergent treatment, electroporation, extrusion, and freeze–thaw cycle to load the drugs following the isolation of exosomes from the respective biological source. All the loading techniques mentioned above are adopted from the field of liposomes by considering exosomes as a cousin of liposomes with higher stability and better biocompatibility. Except incubation, other loading techniques are based upon the assumption that the exosome lipid bilayer will rearrange in its original form similar to liposomes following the application and then removal of stress. Although this assumption may be correct, the effect on the biological activity of integral proteins and their rearrangement in the original position needs to be explored further. While every method has its advantages and disadvantages, incubation may be considered the simplest or safest method of drug loading which is not supposed to change the structures and the different components.
4.1.1. Incubation
Incubation is the simplest technique to load drugs into/onto EVs. In this method, the drug solution is simply mixed with isolated exosomes and incubated at different temperatures (body temperature and room temperature) and in the presence of a lower percentage of organic solvents which is supposed to allow the drug diffusion into the exosomes or hydrophobic interaction onto the exosomal surface protein. The level of drug diffusion is again dependent upon the physicochemical properties of the drug as the hydrophobic drugs can easily interact with the lipid layer of the exosome membrane and hence can permeate easily [
49]. Yang et al., 2015, compared the entrapment efficiency of two exosomal formulations loaded with paclitaxel and doxorubicin, respectively. Both the drugs were incubated with exosomes at body temperature for 2 h and it was found that doxorubicin showed slightly increased entrapment efficiency in comparison with paclitaxel [
50]. We reported the loading of milk-derived exosomes with chemotherapeutic drug PTX by incubation method and observed around 8% of practical loading. Interestingly, the exosomal drug exhibited excellent stability in the presence of simulated-gastrointestinal fluids, and during the low-temperature storage. The exosomal paclitaxel delivered orally showed significant inhibition of lung tumor xenografts and did not elicit any systemic and immunotoxicity compared to
i.v paclitaxel [
51]. Similarly, by using the incubation method, various drugs, and nucleotides,
viz., curcumin, celastrol, anthocyanidins, siRNA, and withaferin A were loaded into/onto milk-derived exosomes and delivered successfully into the animals to treat various types of cancers including breast, lung, ovarian, etc. [
52,
53,
54,
55,
56]. The incubation method is simple and inexpensive in comparison with other techniques, however, not efficient in terms of loading a large number of hydrophilic drugs, nucleic acids, and proteins.
4.1.2. Sonication
Sonication is a physical method in which mechanical shear force is applied by using a probe sonicator which decreases the exosome’s membrane integrity and allows the entry of drugs and other proteins into the exosomes. After which, the membrane is recovered by incubating the exosomes for 30–60 min at 37 °C. This method can be used to load a relatively large amount of drugs into exosomes.
Kim et al., 2016 formulated paclitaxel and doxorubicin-loaded exosomes using three different methods, including incubation at room temperature, electroporation, and sonication. It was found that the sonication method provided higher entrapment efficiency of PTX in the exosomes followed by electroporation and incubation at room temperature. Hence, it was indicated that the sonication method is the best method to load large amounts of drugs into exosomes [
57]. However, exosomes may lose integrity due to high mechanical shear. Although, the sonication method was observed to provide the maximum drug loading to exosomes and could be used for both small and large molecules, the effect of sonication on the exosomal surface biomarkers/proteins and their integrity need to be confirmed.
4.1.3. Electroporation
Electroporation is a well-reported method for the loading of hydrophilic drugs, siRNA, and miRNA into the exosomes as these macromolecules are larger and cannot be loaded to the exosomes by simple diffusion. In this method, exosomes are suspended in an electroporation buffer and the electric field is applied which results in the formation of small pores in the exosomal membrane leading to entry/loading of nucleotides into the exosomes. Although there are several successful reports about this method, RNA precipitation and aggregation of exosomes are also reported in some of the studies [
58]. Hood et al., 2014 proposed that the exosomal membrane aggregation and RNA precipitation can be reduced by using membrane stabilizers such as trehalose pulse media due to their increased colloidal activity [
59]. However, some scientists do not prefer the use of stabilizers as they might get attached to the membrane of exosomes.
The drug entrapment efficiency in the electroporation method depends on parameters,
viz., voltage, capacity, and pulse duration. Generally, in all electroporation methods, a voltage of 350–400 V has been used. Aqil et al., (2019), showed that electroporation could only load about 5% siRNA while chemical transfection resulted in about 30% loading [
56].
4.1.4. Extrusion, Detergent Treatment, and Freeze–Thaw Cycle
In the extrusion method, the exosomes are first mixed with drug solution and loaded into a syringe-based lipid extruder. The solution is then allowed to extrude through membranes of 100–400 nm pore size at controlled temperature. While passing through the membrane, the exosomal membrane gets disrupted which then allows their vigorous mixing with the drug solutions, resulting in effective drug loading into the exosomes. However, due to the usage of mechanical forces, the properties of the exosomal membrane such as zeta potential and protein structures can undergo certain changes. Fuhrmann et al., 2014 demonstrated that loading of porphyrin in MDA-MB-231 breast cancer cell line-derived exosomes by using extrusion method (sample was extruded 31 times) have shown some significant phototoxicity, whereas porphyrin loaded into liposomes did not show any significant phototoxic effect, due to the lack of encapsulation [
60].
In the detergent treatment method, some of the detergents such as saponin have been used, which form the complex with cholesterol in the cell membrane resulting in pore formation and increased membrane permeation [
61]. The size of the membrane pores is approximately 100 Å, which promotes the incorporation of therapeutic agents into exosomes.
In the freeze–thaw cycle method, the drugs are first incubated with exosomes at 37 °C for a specific time, after which the mixture quickly freezes at −80 or −160 °C (in presence of liquid nitrogen) followed by defrosting at room temperature. This cycle of freezing and unfreezing is repeated at least three times [
62]. Although this method may result in a good loading capacity, stress-induced aggregation of exosomes may pose a problem of increase in the size of drug-loaded exosomes. Haney et al., 2015 proposed that the loading of drugs into exosomes by freeze–thaw cycle is significantly lower than the sonication and incubation methods [
63]. Sato et al., 2015 used the freeze-cycle method to fuse the exosomes with liposomes to mimic the actions of liposomes [
64]. However, the use of saponin as a loading material is limited because of the hematolytic nature and possible toxicity in vivo.
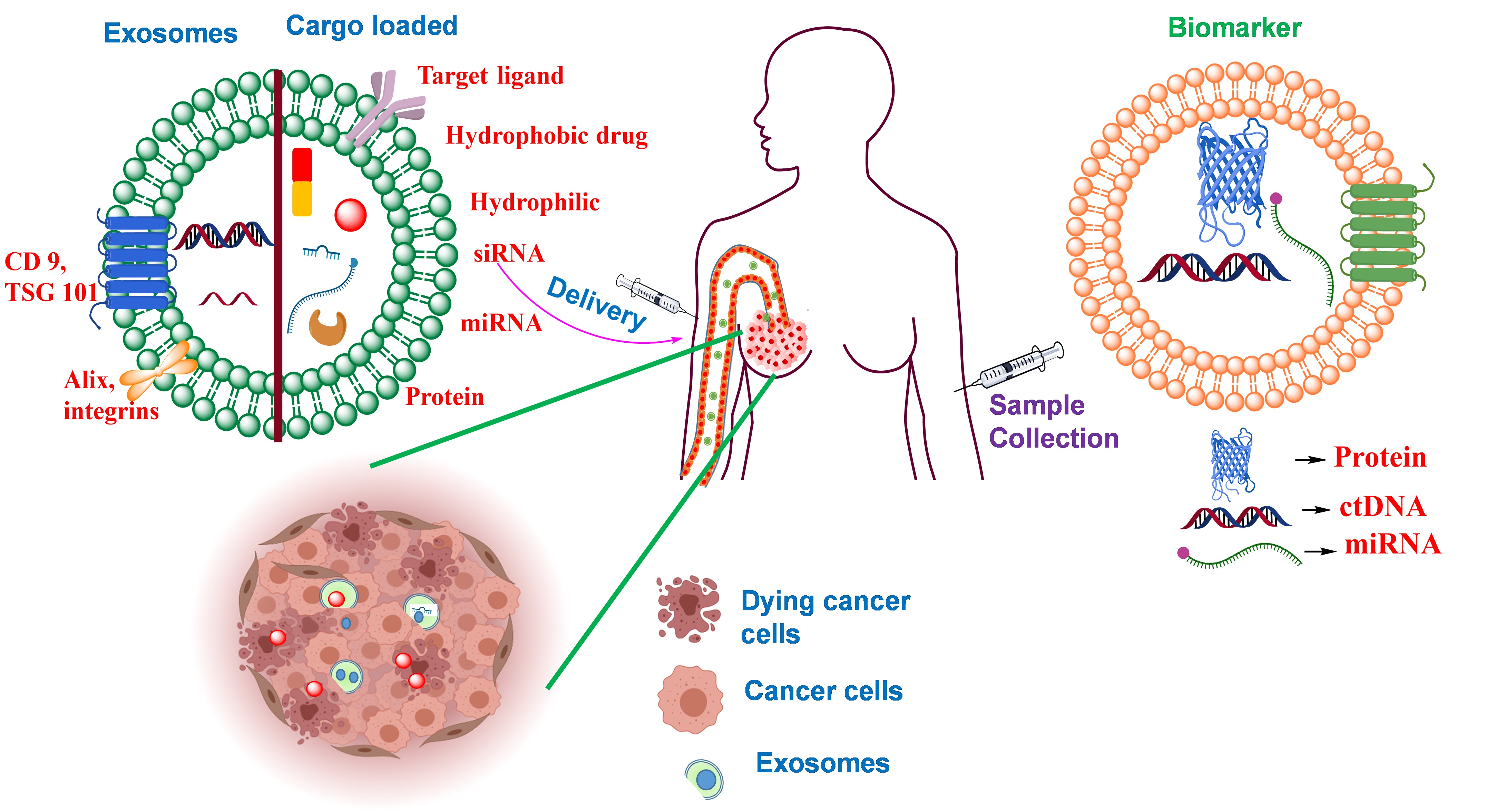
6. Challenges of Exosomes in Drug Delivery and Biomarker
The exosomes do possess certain challenges that need to be addressed. The first challenge faced by the exosomes is the lack of an appropriate standardization technique that should be employed for the isolation and purification of exosomes. The general method used for isolation is multi-step ultracentrifugation which is considered the “gold standard method” and at the same time, there arises a risk where the exosomes obtained might get contaminated with other types of EVs. Absence of a precise isolation and purification method leads to a risk of having both exosomal and non-exosomal EVs in the group of obtained exosomes. The existence of non-exosomal EVs may further compromise the therapeutic efficacy of the exosomal EVs. Secondly, for employing exosomes for cancer therapy, one should prevent sourcing exosomes from cancer cells because they might contain oncogenes that will facilitate cancer progression. Finally, the extraction of exosomes from cell cultures may display variability in their properties even though they are extracted from the same type of donor cells. It was observed that the cell culture and purification techniques limit the implementation of exosomes as therapeutic vesicles as well as hinder their mass production.
In addition, the exosomes are also used as biomarkers which are also presented with some challenges. Cells produce certain sets of proteins and miRNAs; it was observed that the exosomes also produce or express similar sets of proteins and miRNAs with few distinct cell-specific proteins. Further, because the exosomal populations expressed from single cells are heterogeneous, the content concentrations of the exosomes are expected to reside in a range rather than a set standard. Additionally, the exosomes are isolated from various types of cells, so unless they present some exceedingly specific cargoes, the determination of their tissue of origin would be challenging. To date, there remains a lack of compiled data based on exosomes required for the diagnosis of diseases. Additionally, there is no convenient method in terms of time, sample throughput, quality control, and accuracy or regulated technologies under the clinical level that could be used for detection and analysis of exosomes.
8. Conclusions
Exosomes are nanosized lipid-based small EVs that have been successfully used as a diagnostic tool and delivery vehicle for a variety of bioactive, viz., small molecules, proteins, DNAs, and RNAs such as mRNAs, miRNAs, etc., to the desired site and modulate the cellular communication within the tumor microenvironment. Surface functionalized exosomes further represent a novel approach to have their specific accumulation at the site of interest minimizing off-target effects and effective management of various cancers including breast cancer. Exosomes can be employed for therapeutic advantages by profiling their contents to assist with the diagnosis of breast cancer, evaluating their unique molecular characteristics to observe their distribution locally or distally, and transforming them into a targeted drug delivery system for cancer treatment. Recognition and modification of various cell-derived exosomal contents may provide a platform for novel diagnostic, preventive, and therapeutic strategies with minimally invasive techniques. Despite being a boon in targeted drug delivery systems against breast cancer, exosomes face certain challenges which include the need for standardization parameters for the classification of exosomes, finding the methods for determining the behavioral interaction of tumor microenvironment and exosomes as the tumor microenvironment is associated with the progression of different stages of breast cancer, and establishing their precise role in organ metastasis. Hence, further work in the field of exosomes is warranted to widen our understanding of the role of exosomes in breast cancer along with improved breast cancer treatment.


 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}