
Sting Is Commonly and Differentially Expressed in T- and Nk-Cell but Not B-Cell Non-Hodgkin Lymphomas


 , , , ,
, , , , 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Results
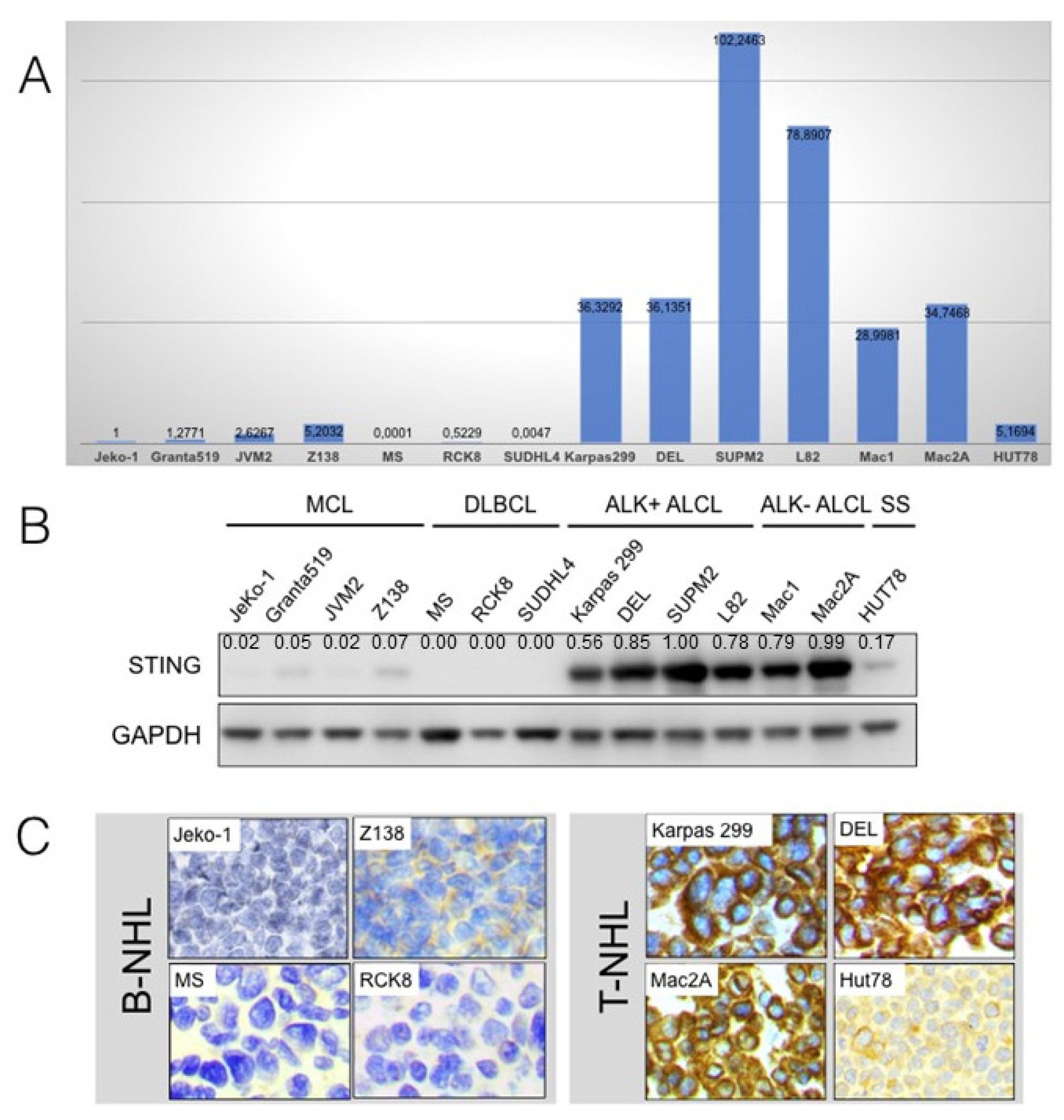
2.1. Expression of STING mRNA and Protein in NHL Cell Lines
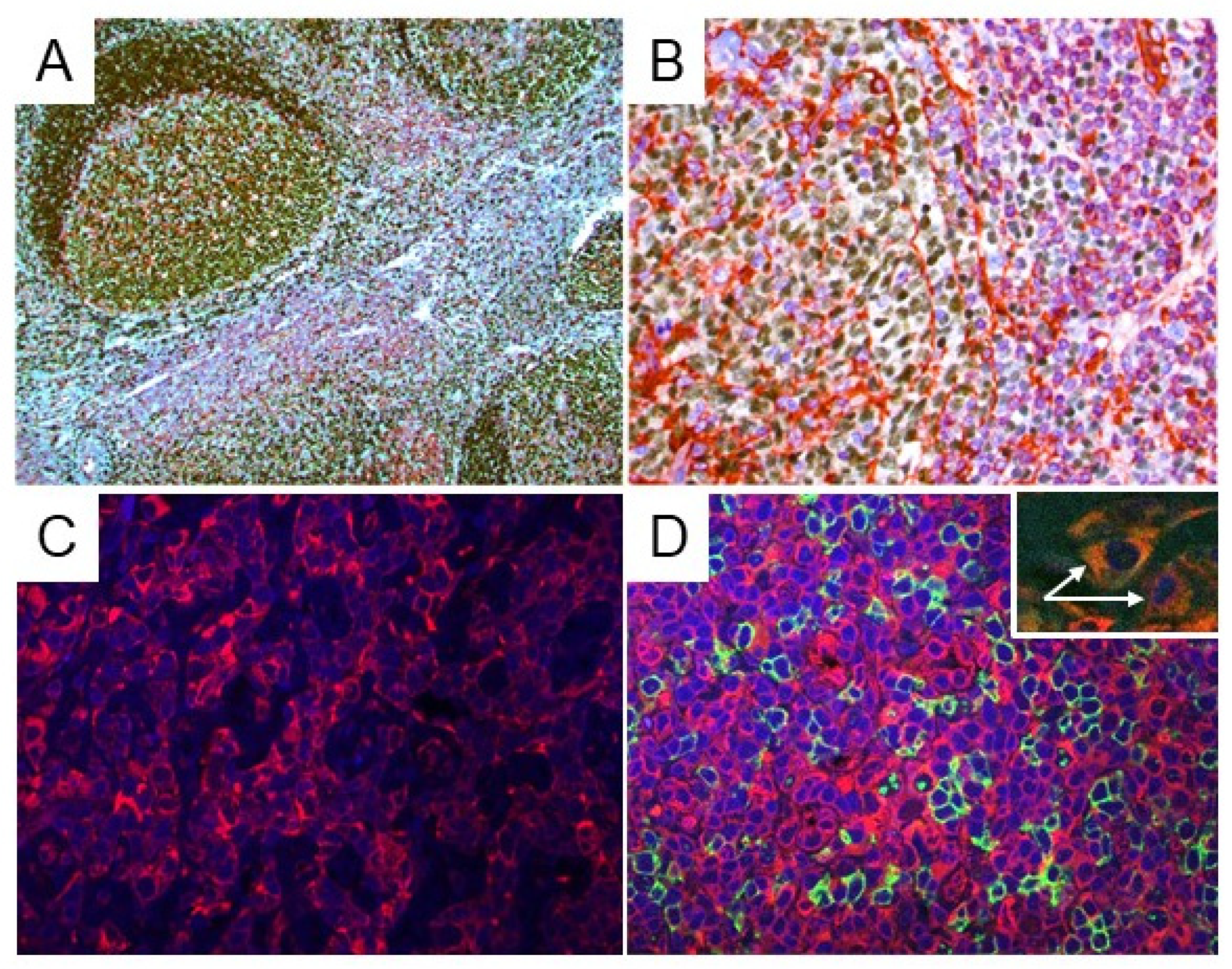
2.2. STING Expression in Reactive Lymph Nodes and Tonsils
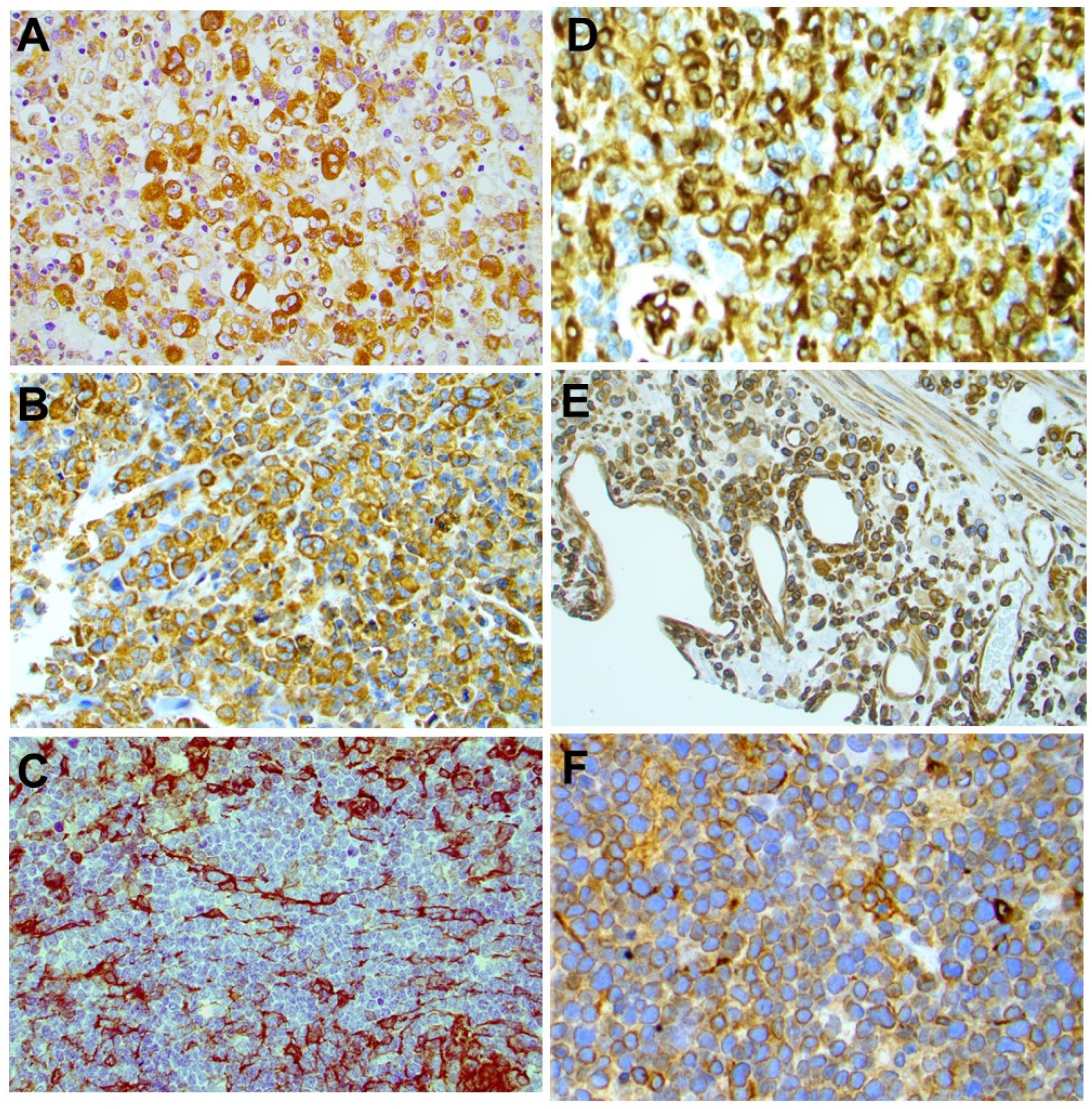
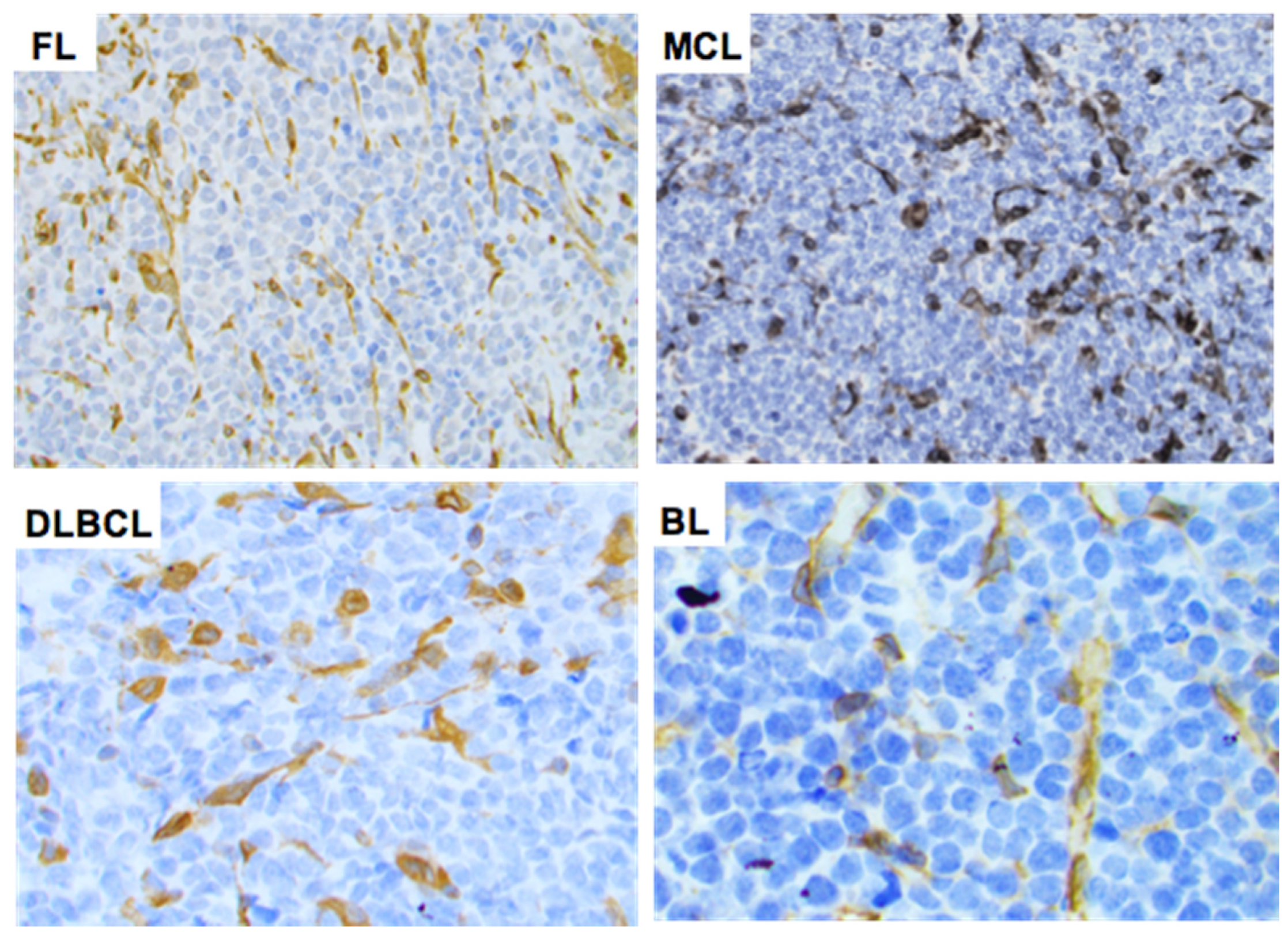
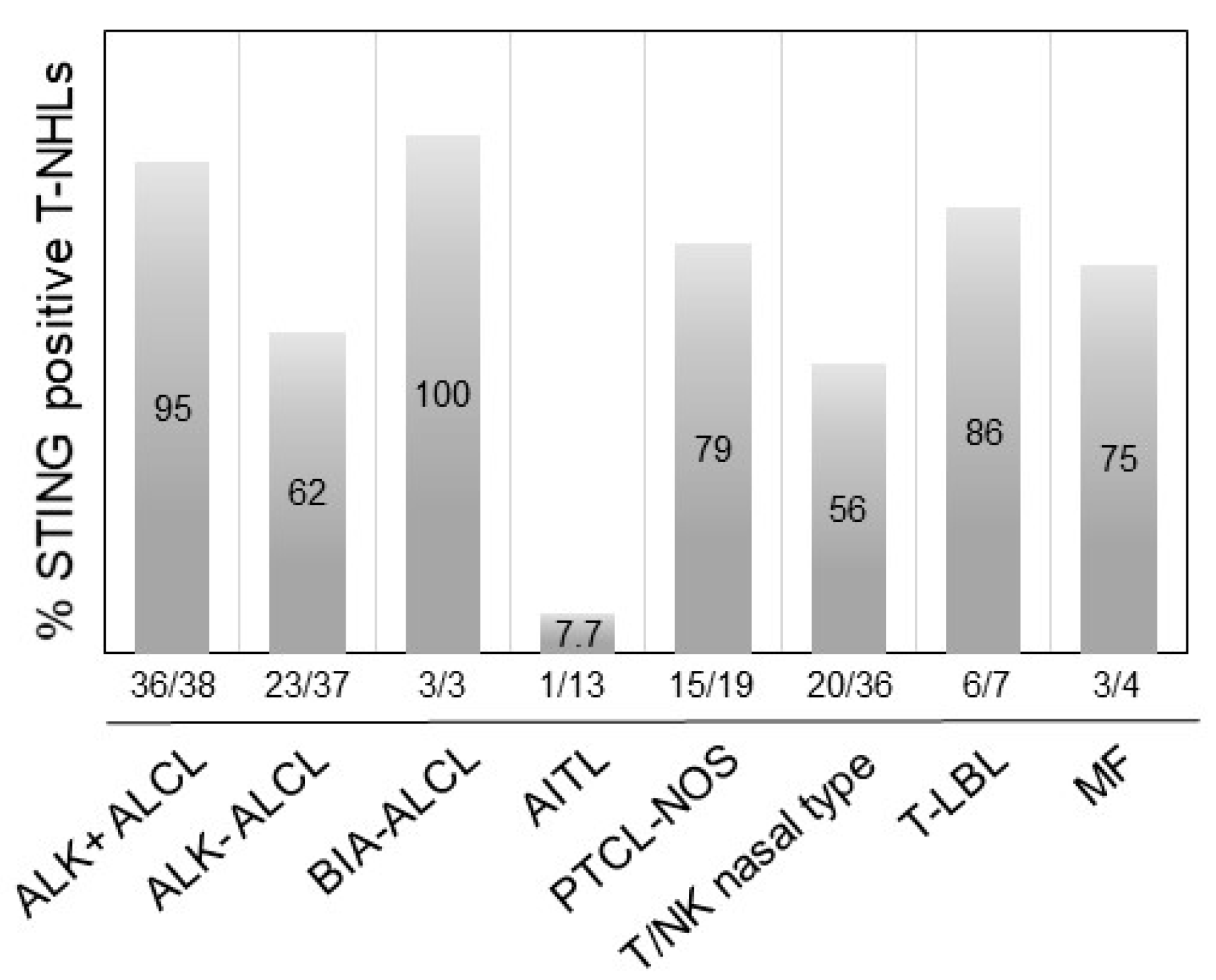
2.3. STING Expression in T-Cell and B-Cell NHLs
2.4. Survival Analysis
3. Discussion
4. Materials and Methods
4.1. Cell Lines
4.2. Gene Expression Analysis
4.3. Western Blot Analysis
4.4. Patient Groups
4.5. Therapy
4.6. Immunohistochemical Methods
4.7. Indirect Immunofluorescence
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kwon, J.; Bakhoum, S.F. The Cytosolic DNA-Sensing cGAS-STING Pathway in Cancer. Cancer Discov. 2020, 10, 26–39. [Google Scholar] [CrossRef] [PubMed]
- Swerdlow, S.H.; Campo, E.; Harris, N.L.; Jaffe, E.S.; Pileri, S.A.; Stein, H.; Thiele, J. World Health Organization Classification of Tumours. In Pathology and Genetics of Tumours of Haematopoietic and Lymphoid Tissues; IARC Press: Lyon, France, 2008. [Google Scholar]
- Mak, V.; Hamm, J.; Chhanabhai, M.; Shenkier, T.; Klasa, R.; Sehn, L.H.; Villa, D.; Gascoyne, R.D.; Connors, J.M.; Savage, K.J. Survival of patients with peripheral T-cell lymphoma after first relapse or progression: Spectrum of disease and rare long-term survivors. J. Clin. Oncol. 2013, 31, 1970–1976. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, J.; Wright, G.; Wang, C.; Rosenwald, A.; Gascoyne, R.D.; Weisenburger, D.D.; Greiner, T.C.; Smith, L.; Guo, S.; Wilcox, R.A.; et al. Gene expression signatures delineate biological and prognostic subgroups in peripheral T-cell lymphoma. Blood 2014, 123, 2915–2923. [Google Scholar] [CrossRef] [PubMed]
- Sakata-Yanagimoto, M.; Enami, T.; Yoshida, K.; Shiraishi, Y.; Ishii, R.; Miyake, Y.; Muto, H.; Tsuyama, N.; Sato-Otsubo, A.; Okuno, Y.; et al. Somatic RHOA mutation in angioimmunoblastic T cell lymphoma. Nat. Genet. 2014, 46, 171–175. [Google Scholar] [CrossRef]
- Palomero, T.; Couronné, L.; Khiabanian, H.; Kim, M.Y.; Ambesi-Impiombato, A.; Perez-Garcia, A.; Carpenter, Z.; Abate, F.; Allegretta, M.; Haydu, J.E.; et al. Recurrent mutations in epigenetic regulators, RHOA and FYN kinase in peripheral T cell lymphomas. Nat. Genet. 2014, 46, 166–170. [Google Scholar] [CrossRef]
- Crescenzo, R.; Abate, F.; Lasorsa, E.; Tabbo, F.; Gaudiano, M.; Chiesa, N.; Di Giacomo, F.; Spaccarotella, E.; Barbarossa, L.; Ercole, E.; et al. Convergent mutations and kinase fusions lead to oncogenic STAT3 activation in anaplastic large cell lymphoma. Cancer Cell 2015, 27, 516–532. [Google Scholar] [CrossRef] [Green Version]
- Nickelsen, M.; Ziepert, M.; Zeynalova, S.; Glass, B.; Metzner, B.; Leithaeuser, M.; Mueller-Hermelink, M.K.; Pfreundschuh, M.; Schmitz, N. High-dose CHOP plus etoposide (MegaCHOEP) in T-cell lymphoma: A comparative analysis of patients treated within trials of the German High-Grade Non-Hodgkin Lymphoma Study Group (DSHNHL). Ann. Oncol. 2009, 20, 1977–1984. [Google Scholar] [CrossRef]
- Ellin, F.; Landström, J.; Jerkeman, M.; Relander, T. Real-world data on prognostic factors and treatment in peripheral T-cell lymphomas: A study from the Swedish Lymphoma Registry. Blood 2014, 124, 1570–1577. [Google Scholar] [CrossRef] [Green Version]
- Gram, A.M.; Sun, C.; Landman, S.L.; Oosenbrug, T.; Koppejan, H.J.; Kwakkenbos, M.J.; Hoeben, R.C.; Paludan, S.R.; Ressing, M.E. Human B cells fail to secrete type I interferons upon cytoplasmic DNA exposure. Mol. Immunol. 2017, 91, 225–237. [Google Scholar] [CrossRef]
- Tang, C.-H.A.; Lee, A.C.; Chang, S.; Xu, Q.; Shao, A.; Lo, Y.; Spalek, W.T.; Pinilla-Ibarz, J.A.; Del Valle, J.R.; Hu, C.-C.A. STING regulates BCR signaling in normal and malignant B cells. Cell. Mol. Immunol. 2021, 18, 1016–1031. [Google Scholar] [CrossRef]
- Walker, M.M.; Crute, B.W.; Cambier, J.C.; Getahun, A. B Cell-Intrinsic STING Signaling Triggers Cell Activation, Synergizes with B Cell Receptor Signals, and Promotes Antibody Responses. J. Immunol. 2018, 201, 2641–2653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konno, H.; Yamauchi, S.; Berglund, A.; Putney, R.M.; Mulé, J.J.; Barber, G.N. Suppression of STING signaling through epigenetic silencing and missense mutation impedes DNA damage mediated cytokine production. Oncogene 2018, 37, 2037–2051. [Google Scholar] [CrossRef] [PubMed]
- Falahat, R.; Berglund, A.; Putney, R.M.; Perez-Villarroel, P.; Aoyama, S.; Pilon-Thomas, S.; Barber, G.N.; Mulé, J.J. Epigenetic reprogramming of tumor cell-intrinsic STING function sculpts antigenicity and T cell recognition of melanoma. Proc. Natl. Acad. Sci. USA 2021, 118, e2013598118. [Google Scholar] [CrossRef]
- Wang, Y.Y.; Jin, R.; Zhou, G.P.; Xu, H.G. Mechanisms of transcriptional activation of the stimulator of interferon genes by transcription factors CREB and c-Myc. Oncotarget 2016, 7, 85049–85057. [Google Scholar] [CrossRef] [Green Version]
- Ma, F.; Li, B.; Yu, Y.; Iyer, S.S.; Sun, M.; Cheng, G. Positive feedback regulation of type I interferon by the interferon-stimulated gene STING. EMBO Rep. 2015, 16, 202–212. [Google Scholar] [CrossRef]
- Wang, Y.; Luo, J.; Alu, A.; Han, X.; Wei, Y.; Wei, X. cGAS-STING pathway in cancer biotherapy. Mol. Cancer 2020, 19, 136. [Google Scholar] [CrossRef] [PubMed]
- Ohkuri, T.; Kosaka, A.; Ishibashi, K.; Kumai, T.; Hirata, Y.; Ohara, K.; Nagato, T.; Oikawa, K.; Aoki, N.; Harabuchi, Y.; et al. Intratumoral administration of cGAMP transiently accumulates potent macrophages for anti-tumor immunity at a mouse tumor site. Cancer Immunol. Immunother. 2017, 66, 705–716. [Google Scholar] [CrossRef]
- Demaria, O.; De Gassart, A.; Coso, S.; Gestermann, N.; Di Domizio, J.; Flatz, L.; Gaide, O.; Michielin, O.; Hwu, P.; Petrova, T.V.; et al. STING activation of tumor endothelial cells initiates spontaneous and therapeutic antitumor immunity. Proc. Natl. Acad. Sci. USA 2015, 112, 15408–15413. [Google Scholar] [CrossRef] [Green Version]
- Fu, J.; Kanne, D.B.; Leong, M.; Glickman, L.H.; McWhirter, S.M.; Lemmens, E.; Mechette, K.; Leong, J.J.; Lauer, P.; Liu, W.; et al. STING agonist formulated cancer vaccines can cure established tumors resistant to PD-1 blockade. Sci. Transl. Med. 2015, 7, 283ra52. [Google Scholar] [CrossRef] [Green Version]
- Sallets, A.; Robinson, S.; Kardosh, A.; Levy, R. Enhancing immunotherapy of STING agonist for lymphoma in preclinical models. Blood Adv. 2018, 2, 2230–2241. [Google Scholar] [CrossRef]
- Dahal, L.N.; Dou, L.; Hussain, K.; Liu, R.; Earley, A.; Cox, K.L.; Murinello, S.; Tracy, I.; Forconi, F.; Steele, A.J.; et al. STING Activation Reverses Lymphoma-Mediated Resistance to Antibody Immunotherapy. Cancer Res. 2017, 77, 3619–3631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atsaves, V.; Lekakis, L.; Drakos, E.; Leventaki, V.; Ghaderi, M.; Baltatzis, G.E.; Chioureas, D.; Jones, D.; Feretzaki, M.; Liakou, C.; et al. The oncogenic JUNB/CD30 axis contributes to cell cycle deregulation in ALK+ anaplastic large cell lymphoma. Br. J. Haematol. 2014, 167, 514–523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xagoraris, I.; Vassilakopoulos, T.P.; Drakos, E.; Angelopoulou, M.K.; Panitsas, F.; Herold, N.; Medeiros, L.J.; Giakoumis, X.; Pangalis, G.A.; Rassidakis, G.Z. Expression of the novel tumour suppressor sterile alpha motif and HD domain-containing protein 1 is an independent adverse prognostic factor in classical Hodgkin lymphoma. Br. J. Haematol. 2021, 193, 488–496. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lymphoma Type | Number of Patients | STING-Positive (n, %) |
|---|---|---|
| T-NHLs | 158 | 107 (68%) |
| ALK+ALCL | 38 | 36 (95%) |
| ALK-ALCL | 37 | 23 (62%) |
| BIA-ALCL | 3 | 3 (100%) |
| AITL | 13 | 1 (7.7%) |
| PTCL-NOS | 19 | 15 (79%) |
| MEITL | 1 | 0 (0%) |
| NK/T-nasal type | 36 | 20 (56%) |
| T-LBL | 7 | 6 (86%) |
| MF | 4 (2 typical, 2 transformed) | 3 (75%) |
| B-NHLs | 265 | 0 (0%) |
| FL | 84 | 0 (0%) |
| MCL | 72 (65 typical, 7 blastoid) | 0 (0%) |
| MZL | 19 | 0 (0%) |
| CLL/SLL | 19 | 0 (0%) |
| DLBCL | 61 | 0 (0%) |
| PMLBCL | 4 | 0 (0%) |
| BL | 3 | 0 (0%) |
| HG-NHL | 3 | 0 (0%) |
| TOTAL | 418 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xagoraris, I.; Farrajota Neves da Silva, P.; Kokaraki, G.; Stathopoulou, K.; Wahlin, B.; Österborg, A.; Herold, N.; Ng, S.-B.; Medeiros, L.J.; Drakos, E.; et al. Sting Is Commonly and Differentially Expressed in T- and Nk-Cell but Not B-Cell Non-Hodgkin Lymphomas. Cancers 2022, 14, 1186. https://doi.org/10.3390/cancers14051186
Xagoraris I, Farrajota Neves da Silva P, Kokaraki G, Stathopoulou K, Wahlin B, Österborg A, Herold N, Ng S-B, Medeiros LJ, Drakos E, et al. Sting Is Commonly and Differentially Expressed in T- and Nk-Cell but Not B-Cell Non-Hodgkin Lymphomas. Cancers. 2022; 14(5):1186. https://doi.org/10.3390/cancers14051186
Chicago/Turabian StyleXagoraris, Ioanna, Pedro Farrajota Neves da Silva, Georgia Kokaraki, Konstantina Stathopoulou, Björn Wahlin, Anders Österborg, Nikolas Herold, Siok-Bian Ng, L. Jeffrey Medeiros, Elias Drakos, and et al. 2022. "Sting Is Commonly and Differentially Expressed in T- and Nk-Cell but Not B-Cell Non-Hodgkin Lymphomas" Cancers 14, no. 5: 1186. https://doi.org/10.3390/cancers14051186
APA StyleXagoraris, I., Farrajota Neves da Silva, P., Kokaraki, G., Stathopoulou, K., Wahlin, B., Österborg, A., Herold, N., Ng, S.-B., Medeiros, L. J., Drakos, E., Sander, B., & Rassidakis, G. Z. (2022). Sting Is Commonly and Differentially Expressed in T- and Nk-Cell but Not B-Cell Non-Hodgkin Lymphomas. Cancers, 14(5), 1186. https://doi.org/10.3390/cancers14051186