
AKR1B1 as a Prognostic Biomarker of High-Grade Serous Ovarian Cancer

 , and
, and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Groups
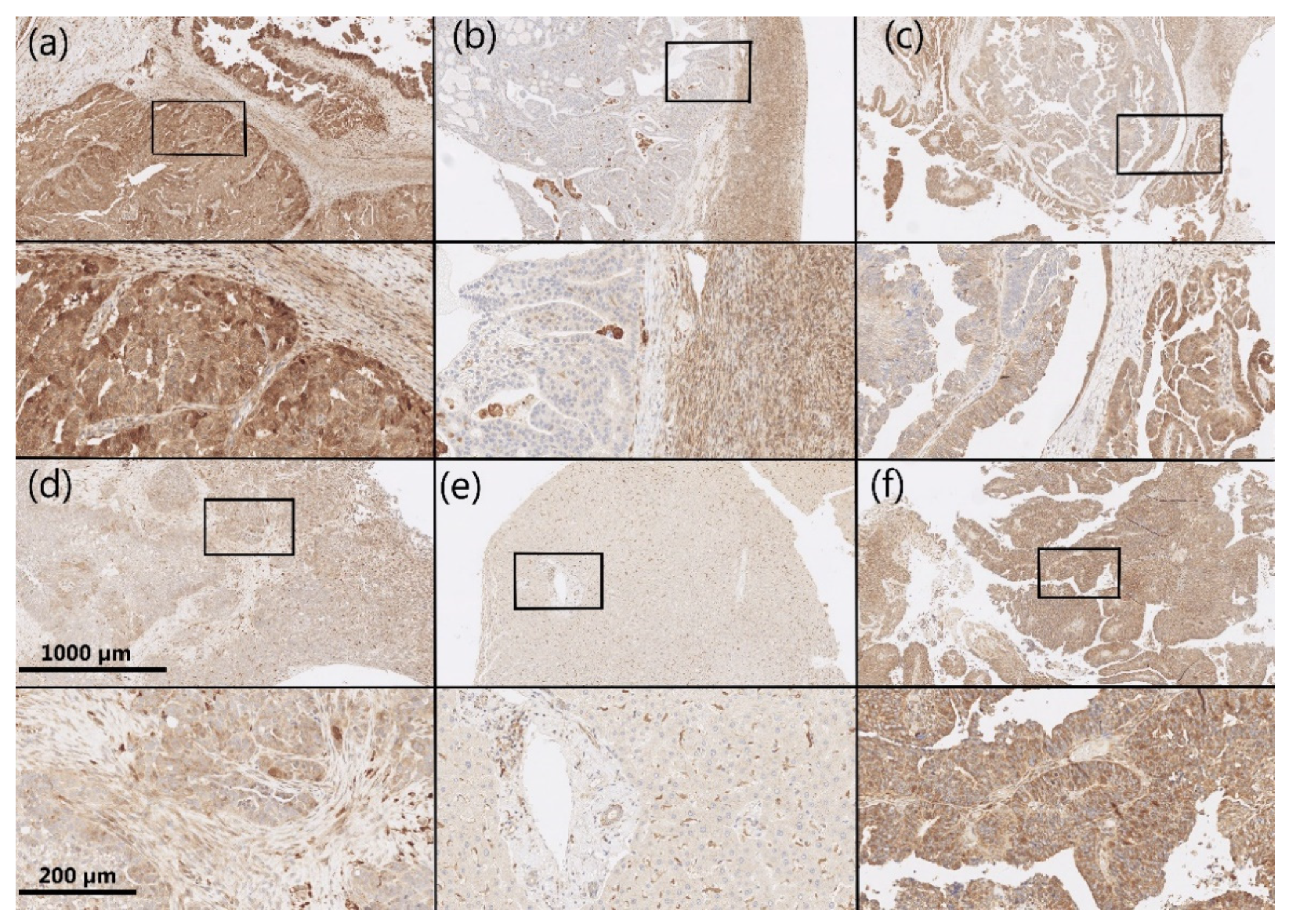
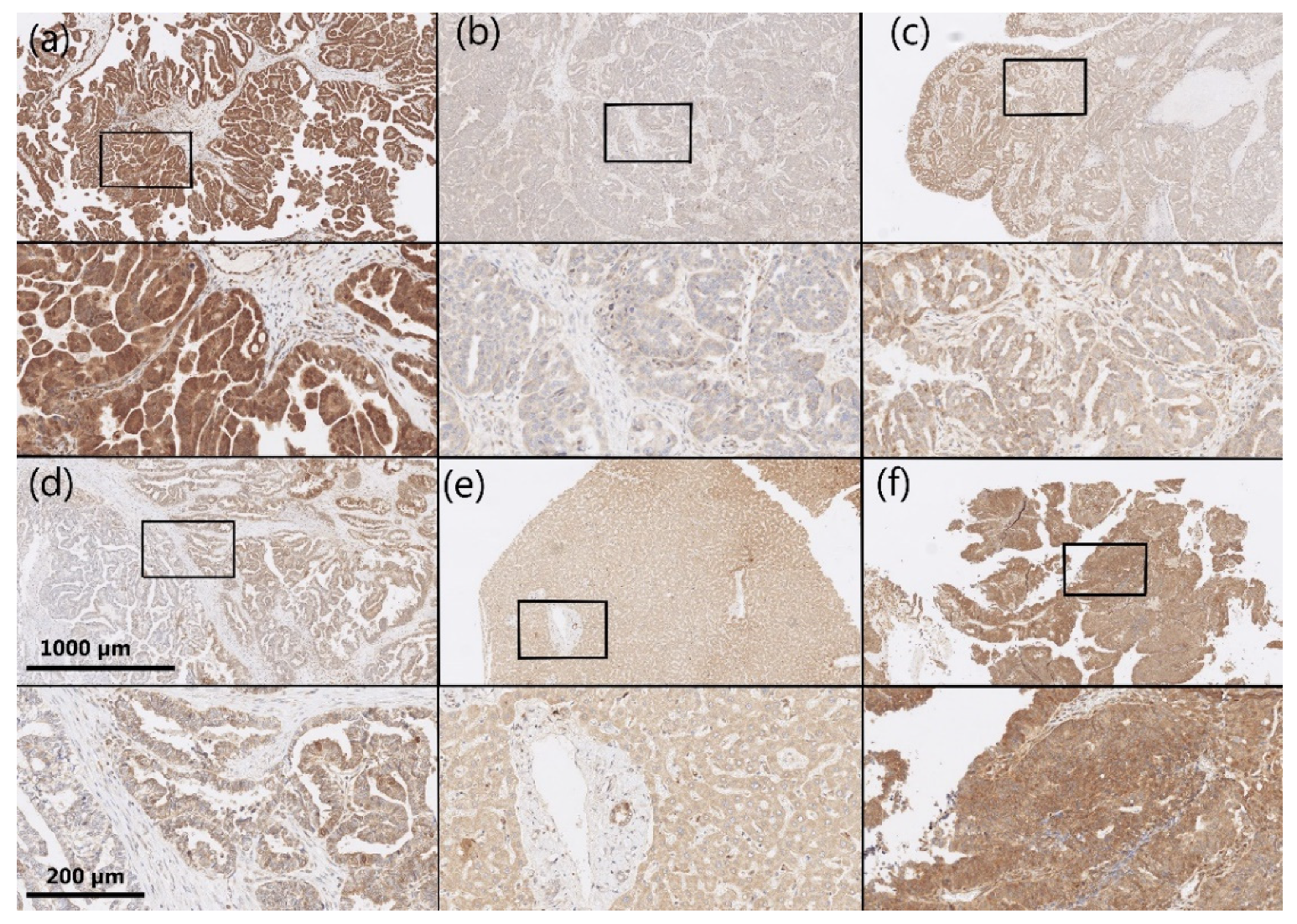
2.2. Immunohistochemistry
2.3. Statistics
2.4. Ethical Issues
3. Results
3.1. Demographic and Histopathological Characteristics of the Patients
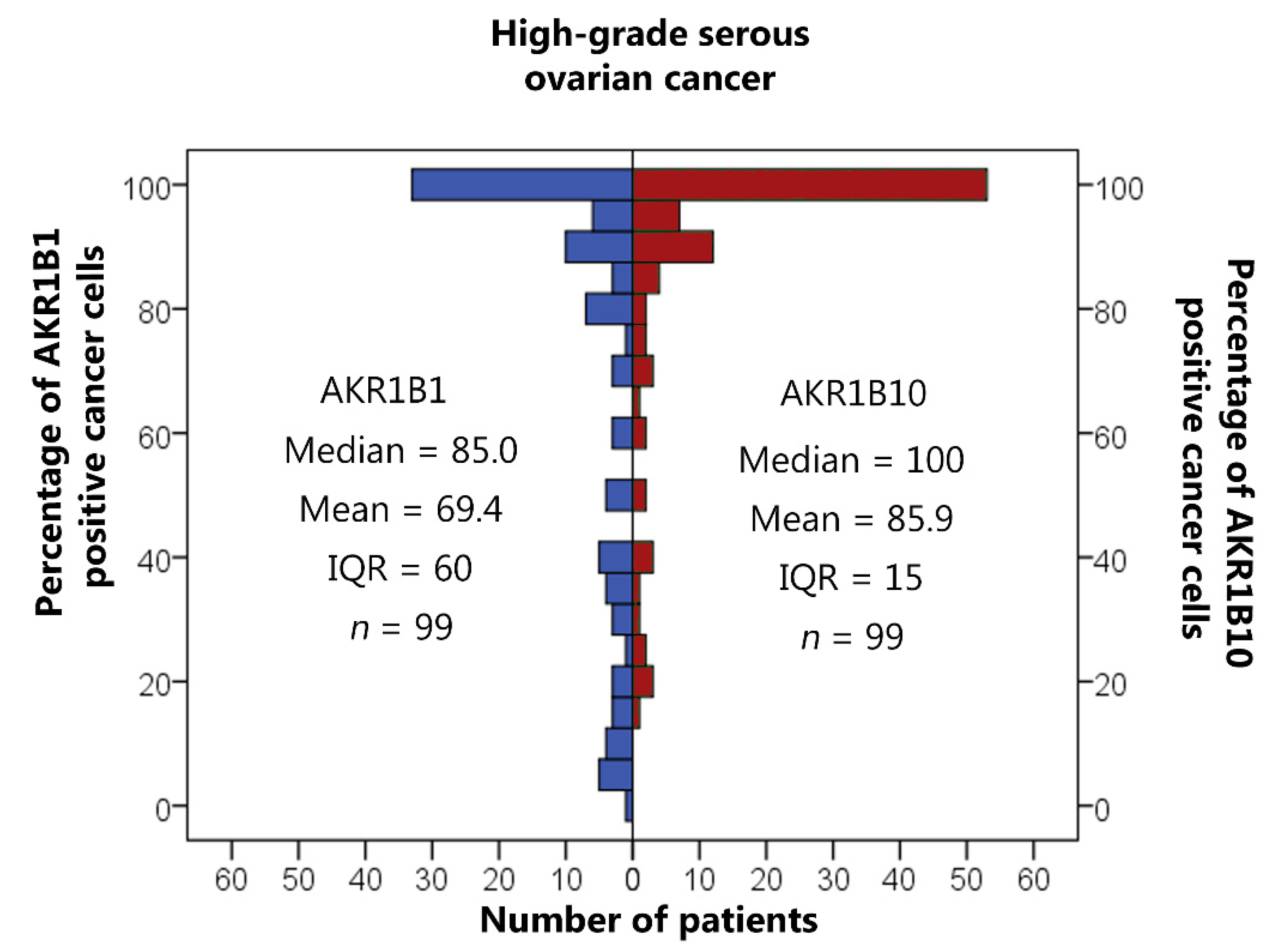
3.2. AKR1B1 and AKR1B10 Expression Levels in HGSC
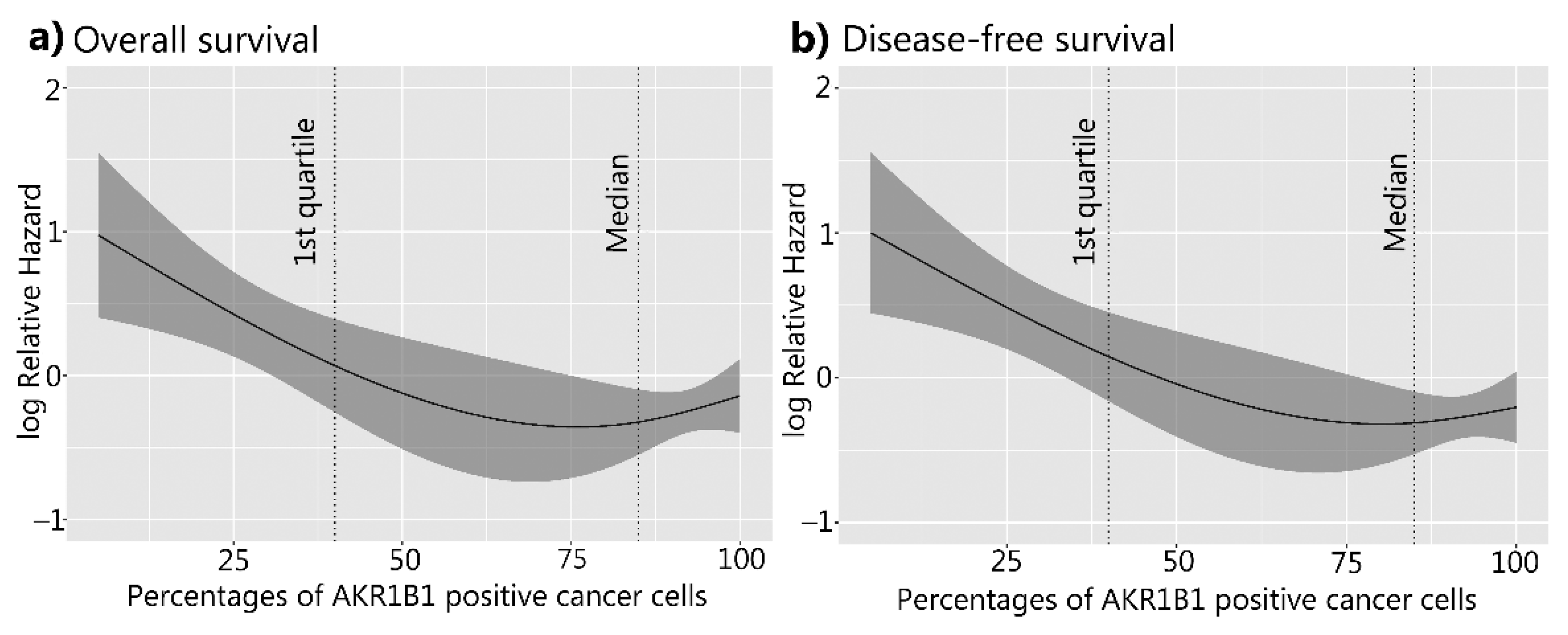
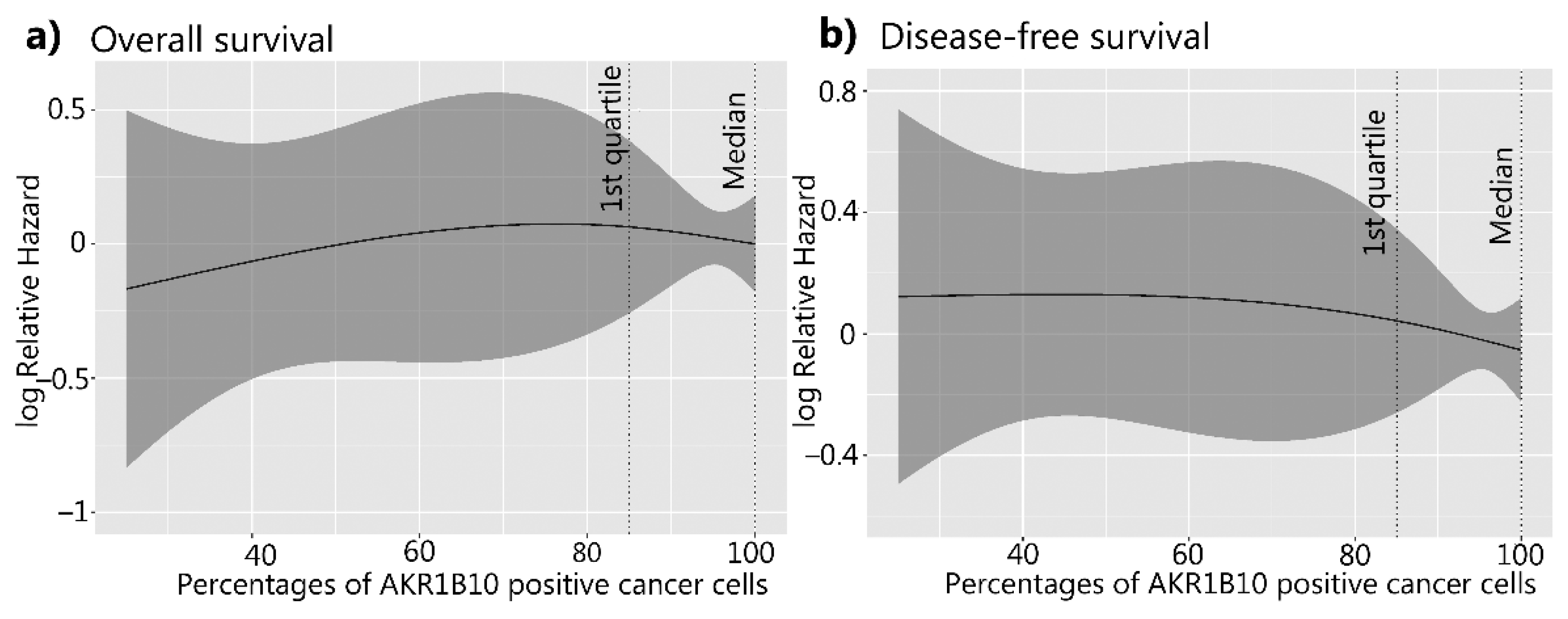
3.3. The Correlation between AKR1B1 and AKR1B10 Expression Levels and Survival
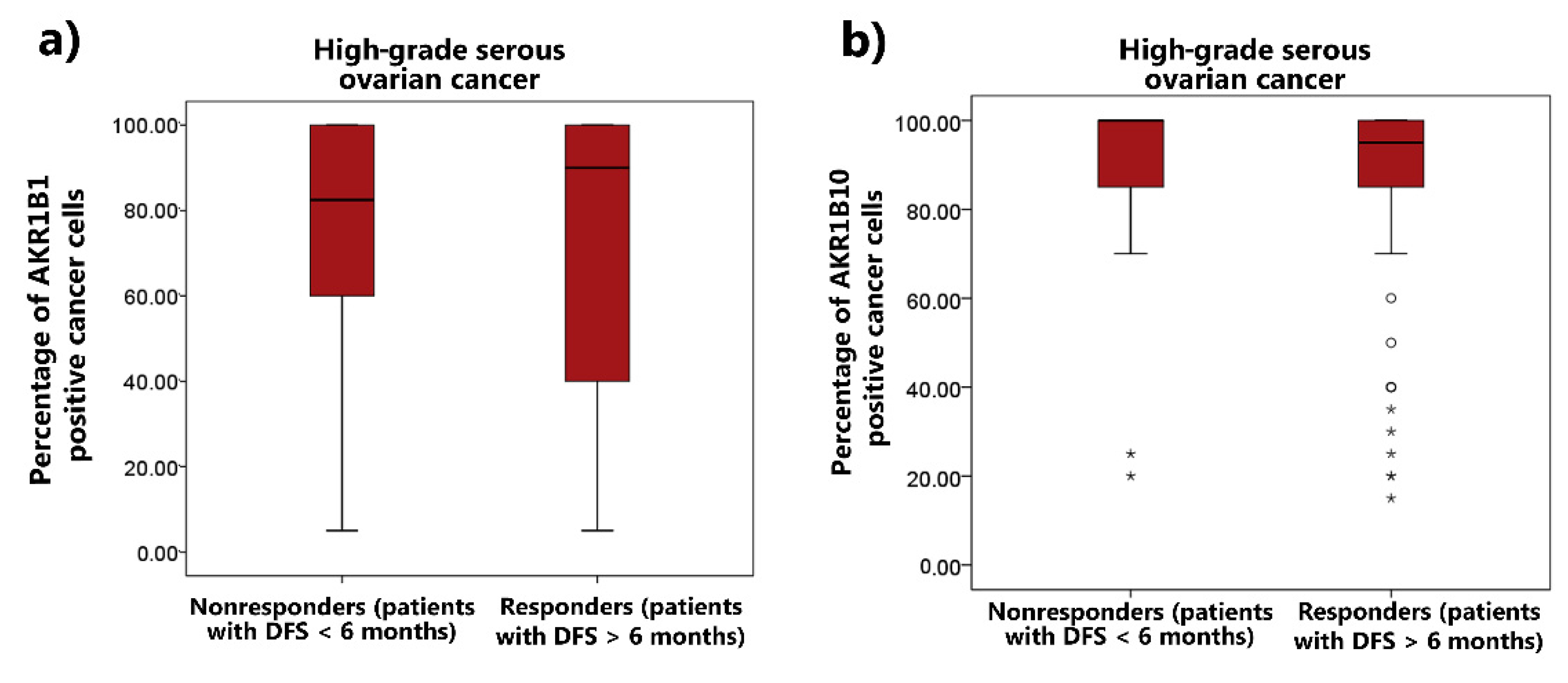
3.4. The Correlation between AKR1B1 and AKR1B10 Expression Levels and Chemoresistance
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Herrington, C.S.; WHO Classification of Tumours Editorial Board. WHO Classification of Tumours Female Genital Tumours; International Agency for Research on Cancer: Lyon, France, 2020. [Google Scholar]
- Kohn, E.C.; Ivy, S.P. Whence High-Grade Serous Ovarian Cancer. Am. Soc. Clin. Oncol. Educ. Book 2017, 37, 443–448. [Google Scholar] [CrossRef]
- Lisio, M.A.; Fu, L.; Goyeneche, A.; Gao, Z.H.; Telleria, C. High-Grade Serous Ovarian Cancer: Basic Sciences, Clinical and Therapeutic Standpoints. Int. J. Mol. Sci. 2019, 20, 952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- The Cancer Genome Atlas Research Network. Integrated genomic analyses of ovarian carcinoma. Nature 2011, 474, 609–615. [Google Scholar] [CrossRef]
- Gadducci, A.; Guarneri, V.; Peccatori, F.A.; Ronzino, G.; Scandurra, G.; Zamagni, C.; Zola, P.; Salutari, V. Current strategies for the targeted treatment of high-grade serous epithelial ovarian cancer and relevance of BRCA mutational status. J. Ovarian Res. 2019, 12, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matulonis, U.A.; Sood, A.K.; Fallowfield, L.; Howitt, B.E.; Sehouli, J.; Karlan, B.Y. Ovarian cancer. Nat. Rev. Dis. Primers 2016, 2, 16061. [Google Scholar] [CrossRef] [PubMed]
- Ushijima, K. Treatment for recurrent ovarian cancer-at first relapse. J. Oncol. 2010, 2010, 497429. [Google Scholar] [CrossRef] [PubMed]
- Kurnit, K.C.; Avila, M.; Hinchcliff, E.M.; Coleman, R.L.; Westin, S.N. PARP inhibition in the ovarian cancer patient: Current approvals and future directions. Pharmacol. Ther. 2020, 213, 107588. [Google Scholar] [CrossRef]
- Tew, W.P.; Lacchetti, C.; Ellis, A.; Maxian, K.; Banerjee, S.; Bookman, M.; Jones, M.B.; Lee, J.M.; Lheureux, S.; Liu, J.F.; et al. PARP Inhibitors in the Management of Ovarian Cancer: ASCO Guideline. J. Clin. Oncol. 2020, 38, 3468–3493. [Google Scholar] [CrossRef]
- da Cunha Colombo Bonadio, R.R.; Fogace, R.N.; Miranda, V.C.; Diz, M.D.P.E. Homologous recombination deficiency in ovarian cancer: A review of its epidemiology and management. Clinics 2018, 73, e450s. [Google Scholar] [CrossRef]
- Su, R.; Liu, Y.; Wu, X.; Xiang, J.; Xi, X. Dynamically Accumulating Homologous Recombination Deficiency Score Served as an Important Prognosis Factor in High-Grade Serous Ovarian Cancer. Front. Mol. Biosci. 2021, 8, 762741. [Google Scholar] [CrossRef]
- Mansoori, B.; Mohammadi, A.; Davudian, S.; Shirjang, S.; Baradaran, B. The Different Mechanisms of Cancer Drug Resistance: A Brief Review. Adv. Pharm. Bull. 2017, 7, 339–348. [Google Scholar] [CrossRef] [PubMed]
- Bell, C.C.; Gilan, O. Principles and mechanisms of non-genetic resistance in cancer. Br. J. Cancer 2019, 122, 465–472. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, Q.; Zhou, L.; Xie, N.; Nice, E.C.; Zhang, H.; Huang, C.; Lei, Y. Cancer drug resistance: Redox resetting renders a way. Oncotarget 2016, 7, 42740–42761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashworth, A. Drug resistance caused by reversion mutation. Cancer Res. 2008, 68, 10021–10023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rižner, T.L.; Penning, T.M. Role of aldo-keto reductase family 1 (AKR1) enzymes in human steroid metabolism. Steroids 2014, 79, 49–63. [Google Scholar] [CrossRef] [Green Version]
- Schwab, A.; Siddiqui, A.; Vazakidou, M.E.; Napoli, F.; Böttcher, M.; Menchicchi, B.; Raza, U.; Saatci, Ö.; Krebs, A.M.; Ferrazzi, F.; et al. Polyol Pathway Links Glucose Metabolism to the Aggressiveness of Cancer Cells. Cancer Res. 2018, 78, 1604–1618. [Google Scholar] [CrossRef] [Green Version]
- Khayami, R.; Hashemi, S.R.; Kerachian, M.A. Role of aldo-keto reductase family 1 member B1 (AKR1B1) in the cancer process and its therapeutic potential. J. Cell Mol. Med. 2020, 24, 8890–8902. [Google Scholar] [CrossRef]
- Alzamil, H.A.; Pawade, J.; Fortier, M.A.; Bernal, A.L. Expression of the prostaglandin F synthase AKR1B1 and the prostaglandin transporter SLCO2A1 in human fetal membranes in relation to spontaneous term and preterm labor. Front. Physiol. 2014, 5, 272. [Google Scholar] [CrossRef] [Green Version]
- Kang, E.S.; Kim, G.H.; Kim, H.J.; Woo, I.S.; Ham, S.A.; Jin, H.; Kim, M.Y.; Lee, J.H.; Chang, K.C.; Seo, H.G.; et al. Nrf2 regulates curcumin-induced aldose reductase expression indirectly via nuclear factor-kappaB. Pharmacol. Res. 2008, 58, 15–21. [Google Scholar] [CrossRef]
- Matsunaga, T.; Wada, Y.; Endo, S.; Soda, M.; El-Kabbani, O.; Hara, A. Aldo-Keto Reductase 1B10 and Its Role in Proliferation Capacity of Drug-Resistant Cancers. Front. Pharmacol. 2012, 3, 5. [Google Scholar] [CrossRef] [Green Version]
- Gallego, O.; Ruiz, F.X.; Ardèvol, A.; Domínguez, M.; Alvarez, R.; de Lera, A.R.; Rovira, C.; Farrés, J.; Fita, I.; Parés, X. Structural basis for the high all-trans-retinaldehyde reductase activity of the tumor marker AKR1B10. Proc. Natl. Acad. Sci. USA 2007, 104, 20764–20769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, J.; Yan, R.; Zu, X.; Cheng, J.M.; Rao, K.; Liao, D.F.; Cao, D. Aldo-keto reductase family 1 B10 affects fatty acid synthesis by regulating the stability of acetyl-CoA carboxylase-alpha in breast cancer cells. J. Biol. Chem. 2008, 283, 3418–3423. [Google Scholar] [CrossRef] [Green Version]
- Mounier, C.; Bouraoui, L.; Rassart, E. Lipogenesis in cancer progression (review). Int. J. Oncol. 2014, 45, 485–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, L.; Shen, H.; Huang, C.; Jing, H.; Cao, D. AKR1B10 induces cell resistance to daunorubicin and idarubicin by reducing C13 ketonic group. Toxicol. Appl. Pharmacol. 2011, 255, 40–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsunaga, T.; Suzuki, A.; Kezuka, C.; Okumura, N.; Iguchi, K.; Inoue, I.; Soda, M.; Endo, S.; El-Kabbani, O.; Hara, A.; et al. Aldo-keto reductase 1B10 promotes development of cisplatin resistance in gastrointestinal cancer cells through down-regulating peroxisome proliferator-activated receptor-γ-dependent mechanism. Chem. Biol. Interact. 2016, 256, 142–153. [Google Scholar] [CrossRef]
- Martin, H.J.; Breyer-Pfaff, U.; Wsol, V.; Venz, S.; Block, S.; Maser, E. Purification and characterization of akr1b10 from human liver: Role in carbonyl reduction of xenobiotics. Drug Metab. Dispos. 2006, 34, 464–470. [Google Scholar] [CrossRef]
- Hevir, N.; Sinkovec, J.; Lanišnik Rižner, T. Decreased levels of AKR1B1 and AKR1B10 in cancerous endometrium compared to adjacent non-cancerous tissue. Chem. Biol. Interact. 2013, 202, 226–233. [Google Scholar] [CrossRef]
- The R Development Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing: Vienna, Austria. 2021. Available online: https://www.R-project.org/ (accessed on 10 October 2021).
- Harrell, F.E., Jr. Regression Modeling Strategies. R Package Version 6.2-0. CRAN. Vienna, Austria. 2021. Available online: https://CRAN.R-project.org/package=rms (accessed on 10 October 2021).
- Cerami, E.; Gao, J.; Dogrusoz, U.; Gross, B.E.; Sumer, S.O.; Aksoy, B.A.; Jacobsen, A.; Byrne, C.J.; Heuer, M.L.; Larsson, E.; et al. The cBio cancer genomics portal: An open platform for exploring multidimensional cancer genomics data. Cancer Discov. 2012, 2, 401–404. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.; Aksoy, B.A.; Dogrusoz, U.; Dresdner, G.; Gross, B.; Sumer, S.O.; Sun, Y.; Jacobsen, A.; Sinha, R.; Larsson, E.; et al. Integrative analysis of complex cancer genomics and clinical profiles using the cBioPortal. Sci. Signal. 2013, 6, pl1. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Liu, T.; Zhang, Z.; Payne, S.H.; Zhang, B.; McDermott, J.E.; Zhou, J.Y.; Petyuk, V.A.; Chen, L.; Ray, D.; et al. Integrated Proteogenomic Characterization of Human High-Grade Serous Ovarian Cancer. Cell 2016, 166, 755–765. [Google Scholar] [CrossRef] [Green Version]
- Penning, T.M. The aldo-keto reductases (AKRs): Overview. Chem. Biol. Interact. 2015, 234, 236–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rižner, T.L. Enzymes of the AKR1B and AKR1C Subfamilies and Uterine Diseases. Front. Pharmacol. 2012, 3, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hojnik, M.; Kenda Šuster, N.; Smrkolj, Š.; Frković Grazio, S.; Verdenik, I.; Rižner, T.L. AKR1C3 Is Associated with Better Survival of Patients with Endometrial Carcinomas. J. Clin. Med. 2020, 9, 4105. [Google Scholar] [CrossRef] [PubMed]
- Hojnik, M.; Frković Grazio, S.; Verdenik, I.; Rižner, T.L. AKR1B1 and AKR1B10 as Prognostic Biomarkers of Endometrioid Endometrial Carcinomas. Cancers 2021, 13, 3398. [Google Scholar] [CrossRef]
- Reddy, K.A.; Kumar, P.U.; Srinivasulu, M.; Triveni, B.; Sharada, K.; Ismail, A.; Reddy, G.B. Overexpression and enhanced specific activity of aldoketo reductases (AKR1B1 & AKR1B10) in human breast cancers. Breast 2017, 31, 137–143. [Google Scholar] [CrossRef]
- Fang, C.Y.; Lin, Y.H.; Chen, C.L. Overexpression of AKR1B10 predicts tumor recurrence and short survival in oral squamous cell carcinoma patients. J. Oral Pathol. Med. 2019, 48, 712–719. [Google Scholar] [CrossRef]
- Taskoparan, B.; Seza, E.G.; Demirkol, S.; Tuncer, S.; Stefek, M.; Gure, A.O.; Banerjee, S. Opposing roles of the aldo-keto reductases AKR1B1 and AKR1B10 in colorectal cancer. Cell Oncol. 2017, 40, 563–578. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, S.M.U.; Jiang, Z.N.; Zheng, Z.H.; Li, Y.; Wang, X.J.; Tang, X. AKR1B10 expression predicts response of gastric cancer to neoadjuvant chemotherapy. Oncol. Lett. 2019, 17, 773–780. [Google Scholar] [CrossRef] [Green Version]
- DiStefano, J.K.; Davis, B. Diagnostic and Prognostic Potential of AKR1B10 in Human Hepatocellular Carcinoma. Cancers 2019, 11, 486. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Zhong, X.; Li, J.; Liu, B.; Guo, S.; Chen, J.; Tan, Q.; Wang, Q.; Ma, W.; Wu, Z.; et al. Screening and identification of lung cancer metastasis-related genes by suppression subtractive hybridization. Thorac. Cancer 2012, 3, 207–216. [Google Scholar] [CrossRef]
- Saraswat, M.; Mrudula, T.; Kumar, P.U.; Suneetha, A.; Rao Rao, T.S.; Srinivasulu, M.; Reddy, B. Overexpression of aldose reductase in human cancer tissues. Med. Sci. Monit. 2006, 12, CR525–CR529. [Google Scholar]
- Torres-Mena, J.E.; Salazar-Villegas, K.N.; Sánchez-Rodríguez, R.; López-Gabiño, B.; Del Pozo-Yauner, L.; Arellanes-Robledo, J.; Villa-Treviño, S.; Gutiérrez-Nava, M.A.; Pérez-Carreón, J.I. Aldo-Keto Reductases as Early Biomarkers of Hepatocellular Carcinoma: A Comparison Between Animal Models and Human HCC. Dig. Dis. Sci. 2018, 63, 934–944. [Google Scholar] [CrossRef] [PubMed]
- Demirkol Canlı, S.; Seza, E.G.; Sheraj, I.; Gömçeli, I.; Turhan, N.; Carberry, S.; Prehn, J.H.M.; Güre, A.O.; Banerjee, S. Evaluation of an aldo-keto reductase gene signature with prognostic significance in colon cancer via activation of epithelial to mesenchymal transition and the p70S6K pathway. Carcinogenesis 2020, 41, 1219–1228. [Google Scholar] [CrossRef]
- Uzozie, A.C.; Selevsek, N.; Wahlander, A.; Nanni, P.; Grossmann, J.; Weber, A.; Buffoli, F.; Marra, G. Targeted Proteomics for Multiplexed Verification of Markers of Colorectal Tumorigenesis. Mol. Cell Proteom. 2017, 16, 407–427. [Google Scholar] [CrossRef] [Green Version]
- Uzozie, A.; Nanni, P.; Staiano, T.; Grossmann, J.; Barkow-Oesterreicher, S.; Shay, J.W.; Tiwari, A.; Buffoli, F.; Laczko, E.; Marra, G. Sorbitol dehydrogenase overexpression and other aspects of dysregulated protein expression in human precancerous colorectal neoplasms: A quantitative proteomics study. Mol. Cell Proteom. 2014, 13, 1198–1218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kropotova, E.S.; Tychko, R.A.; Zinov’eva, O.L.; Zyrianova, A.F.; Khankin, S.L.; Cherkes, V.L.; Aliev, V.A.; Beresten, S.F.; Oparina, N.I.U.; Mashkova, T.D. Downregulation of AKR1B10 gene expression in colorectal cancer. Mol. Biol. 2010, 44, 243–250. [Google Scholar] [CrossRef]
- Hung, J.J.; Yeh, Y.C.; Hsu, W.H. Prognostic significance of AKR1B10 in patients with resected lung adenocarcinoma. Thorac. Cancer 2018, 9, 1492–1499. [Google Scholar] [CrossRef]
- Ko, H.H.; Cheng, S.L.; Lee, J.J.; Chen, H.M.; Kuo, M.Y.; Cheng, S.J. Expression of AKR1B10 as an independent marker for poor prognosis in human oral squamous cell carcinoma. Head Neck 2017, 39, 1327–1332. [Google Scholar] [CrossRef]
- Yao, Y.; Wang, X.; Zhou, D.; Li, H.; Qian, H.; Zhang, J.; Jiang, L.; Wang, B.; Lin, Q.; Zhu, X. Loss of AKR1B10 promotes colorectal cancer cells proliferation and migration via regulating FGF1-dependent pathway. Aging 2020, 12, 13059–13075. [Google Scholar] [CrossRef]
- Deng, H.B.; Parekh, H.K.; Chow, K.C.; Simpkkins, H. Increased expression of dihydrodiol dehydrogenase induces resistance to cisplatin in human ovarian carcinoma cells. J. Biol. Chem. 2002, 277, 15035–15043. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.Y.; Hong, S.S.; Zhang, M.; Cai, Q.Q.; Zhang, M.X.; Xu, C.J. Proteomic alterations of fibroblasts induced by ovarian cancer cells reveal potential cancer targets. Neoplasma 2018, 65, 104–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexiou, P.; Pegklidou, K.; Chatzopoulou, M.; Nicolaou, I.; Demopoulos, V.J. Aldose reductase enzyme and its implication to major health problems of the 21st century. Curr. Med. Chem. 2009, 16, 734–752. [Google Scholar] [CrossRef]
- Penning, T.M.; Jonnalagadda, S.; Trippier, P.C.; Rižner, T.L. Aldo-Keto Reductases and Cancer Drug Resistance. Pharmacol. Rev. 2021, 73, 1150–1171. [Google Scholar] [CrossRef]
- Sinreih, M.; Štupar, S.; Čemažar, L.; Verdenik, I.; Frković Grazio, S.; Smrkolj, Š.; Rižner, T.L. STAR and AKR1B10 are down-regulated in high-grade endometrial cancer. J. Steroid. Biochem. Mol. Biol. 2017, 171, 43–53. [Google Scholar] [CrossRef] [PubMed]
- Leone, A.; Roca, M.S.; Ciardiello, C.; Costantini, S.; Budillon, A. Oxidative Stress Gene Expression Profile Correlates with Cancer Patient Poor Prognosis: Identification of Crucial Pathways Might Select Novel Therapeutic Approaches. Oxid. Med. Cell Longev. 2017, 2017, 2597581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, M.; Kapoor, A.; Bhatnagar, A. Oxidative and reductive metabolism of lipid-peroxidation derived carbonyls. Chem. Biol. Interact. 2015, 234, 261–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, K.A.; Choi, B.H.; Nam, C.W.; Song, M.; Kim, S.T.; Lee, J.Y.; Kwak, M.K. Identification of aldo-keto reductases as NRF2-target marker genes in human cells. Toxicol. Lett. 2013, 218, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Penning, T.M. Aldo-Keto Reductase Regulation by the Nrf2 System: Implications for Stress Response, Chemotherapy Drug Resistance, and Carcinogenesis. Chem. Res. Toxicol. 2017, 30, 162–176. [Google Scholar] [CrossRef]
- Asangani, I.; Blair, I.A.; Van Duyne, G.; Hilser, V.J.; Moiseenkova-Bell, V.; Plymate, S.; Sprenger, C.; Wand, A.J.; Penning, T.M. Using Biochemistry & Biophysics to Extinguish Androgen Receptor Signaling in Prostate Cancer. J. Biol. Chem. 2021, 296, 10024. [Google Scholar] [CrossRef]
- Hofman, J.; Malcekova, B.; Skarka, A.; Novotna, E.; Wsol, V. Anthracycline resistance mediated by reductive metabolism in cancer cells: The role of aldo-keto reductase 1C3. Toxicol. Appl. Pharmacol. 2014, 278, 238–248. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | Detail | Datum |
|---|---|---|
| Age (y) | Mean ± SD | 61.3 ± 11.3 |
| Ascites (n (%)) | 54 (54.5) | |
| Chemotherapy with reported follow-up (n = 71) (n (%)) | Responders (at least 6 months of DFS) | 53 (74.6) |
| Non-responders (6 months of DFS was not achieved) | 18 (25.4) | |
| Primary chemotherapy (n) | Carboplatin | 13 |
| Docetaxel and carboplatin | 4 | |
| Doxorubicin | 1 | |
| Gemcitabine | 1 | |
| Gemcitabine and carboplatin | 1 | |
| Paclitaxel and carboplatin | 68 | |
| Paclitaxel and carboplatin and gemcitabine | 1 | |
| None | 6 | |
| NA | 4 | |
| Residual disease after primary chemotherapy (n) | Macroscopic | 51 |
| Microscopic | 46 | |
| No cytoreductive surgery | 2 | |
| FIGO stage (n = 99) | I–II | 19 |
| III–IV | 80 | |
| Grade (n = 99) | High-grade | 99 |
| Follow-up (y) | Range | 0.25–12.6 |
| Median | 3.2 | |
| Five-year survival rate a (n (%)) | 29 (29.9) |
| Antibody Information | |||||
|---|---|---|---|---|---|
| Antibody | Manufacturer, Catalogue Number, Lot Number | Peptide/ Protein Target | Antigen Sequence | Species Raised, Monoclonal, Polyclonal | Dilution |
| Anti-AKR1B1 | Abcam, Cambridge, UK, ab62795, GR64780-2 | Aldo-keto reductase family 1 member B1 | Aa 300 to the C-terminus (conjugated to keyhole limpet hemocyanin) | Polyclonal rabbit antibody | 1:200 |
| Anti-AKR1B10 | Abcam, Cambridge, UK, ab96417, GR13314-31 | Aldo-keto reductase family 1 member B10 | Fragment corresponding to aa 1–286 | Polyclonal rabbit antibody | 1:200 |
| Antibody Validation | |||||
| Published validation by our research team [28]. | |||||
| Current Validation Positive controls for AKR1B1: Kupffer cells, lymphocytes, endometrioid endometrial cancer cells. Positive controls for AKR1B10: hepatocytes, ductal liver epithelium, lymphocytes, endometrioid endometrial cancer cells. Negative controls for AKR1B1: hepatocytes, fibrous tissue. Negative controls for AKR1B10: fibrous tissue. | |||||
| Overall Survival | Significance | Hazard Ratio | Confidence Interval |
|---|---|---|---|
| FIGO (I–II vs. III–IV) | p = 0.01 | 2.41 | 1.23–4.72 |
| Ascites | p = 0.80 | 0.94 | 0.59–1.50 |
| AKR1B1 expression (continuous variable) | p = 0.010 | 0.991 | 0.984–0.998 |
| AKR1B10 expression (continuous variable) | p = 0.621 | 1.002 | 0.993–1.012 |
| Disease-Free Survival | Significance | Hazard Ratio | Confidence Interval |
|---|---|---|---|
| FIGO (I–II vs. III–IV) | p = 0.005 | 2.43 | 1.31–4.51 |
| Ascites | p = 0.761 | 0.93 | 0.60–1.46 |
| AKR1B1 expression (continuous variable) | p = 0.005 | 0.990 | 0.984–0.997 |
| AKR1B10 expression (continuous variable) | p = 0.715 | 0.998 | 0.989–1.008 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hojnik, M.; Šuster, N.K.; Smrkolj, Š.; Sisinger, D.; Grazio, S.F.; Verdenik, I.; Rižner, T.L. AKR1B1 as a Prognostic Biomarker of High-Grade Serous Ovarian Cancer. Cancers 2022, 14, 809. https://doi.org/10.3390/cancers14030809
Hojnik M, Šuster NK, Smrkolj Š, Sisinger D, Grazio SF, Verdenik I, Rižner TL. AKR1B1 as a Prognostic Biomarker of High-Grade Serous Ovarian Cancer. Cancers. 2022; 14(3):809. https://doi.org/10.3390/cancers14030809
Chicago/Turabian StyleHojnik, Marko, Nataša Kenda Šuster, Špela Smrkolj, Damjan Sisinger, Snježana Frković Grazio, Ivan Verdenik, and Tea Lanišnik Rižner. 2022. "AKR1B1 as a Prognostic Biomarker of High-Grade Serous Ovarian Cancer" Cancers 14, no. 3: 809. https://doi.org/10.3390/cancers14030809
APA StyleHojnik, M., Šuster, N. K., Smrkolj, Š., Sisinger, D., Grazio, S. F., Verdenik, I., & Rižner, T. L. (2022). AKR1B1 as a Prognostic Biomarker of High-Grade Serous Ovarian Cancer. Cancers, 14(3), 809. https://doi.org/10.3390/cancers14030809