
ITIH5-Derived Polypeptides Covering the VIT Domain Suppress the Growth of Human Cancer Cells In Vitro

 , ,
, ,  , ,
, ,  , and
, and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Results
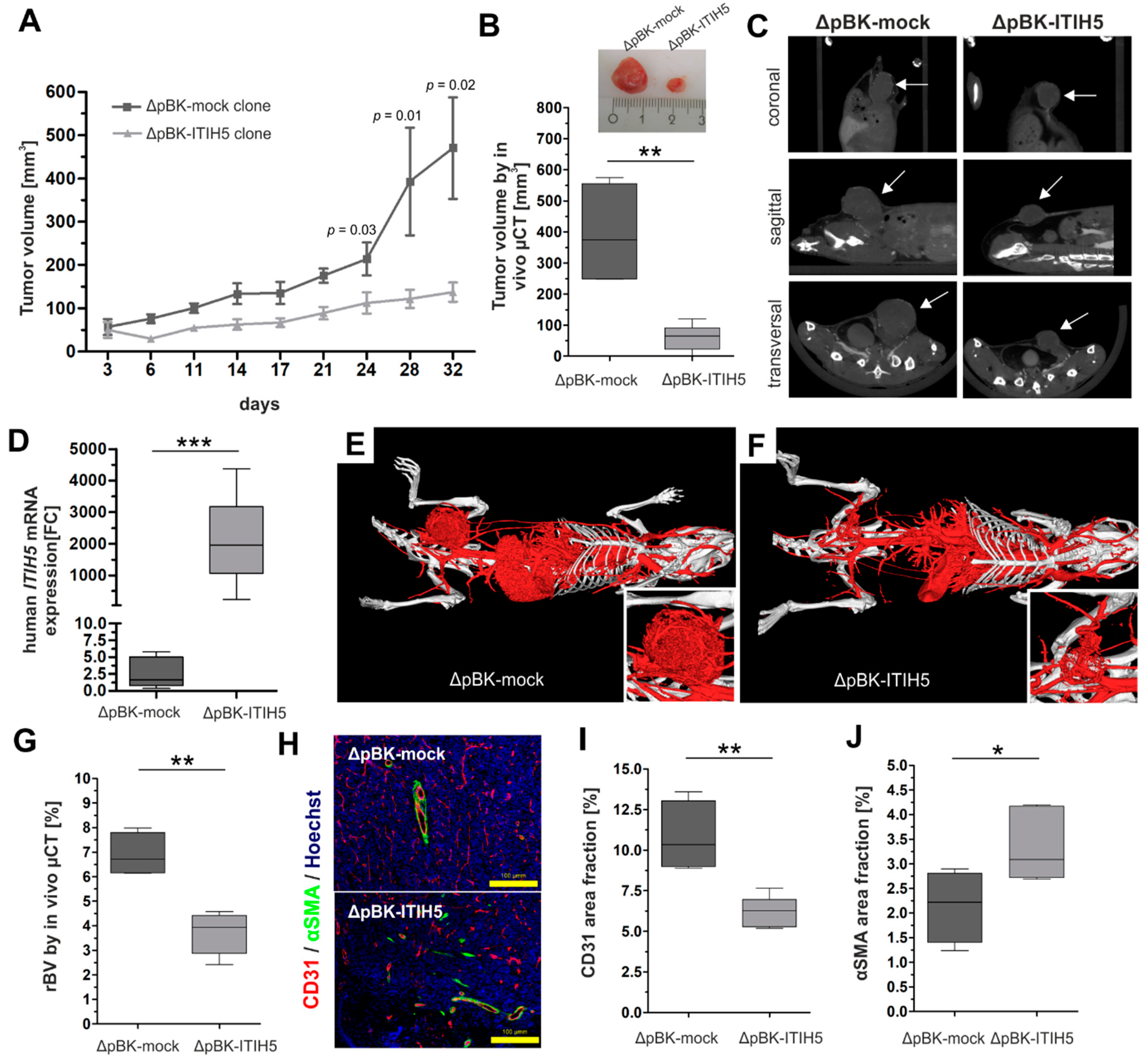
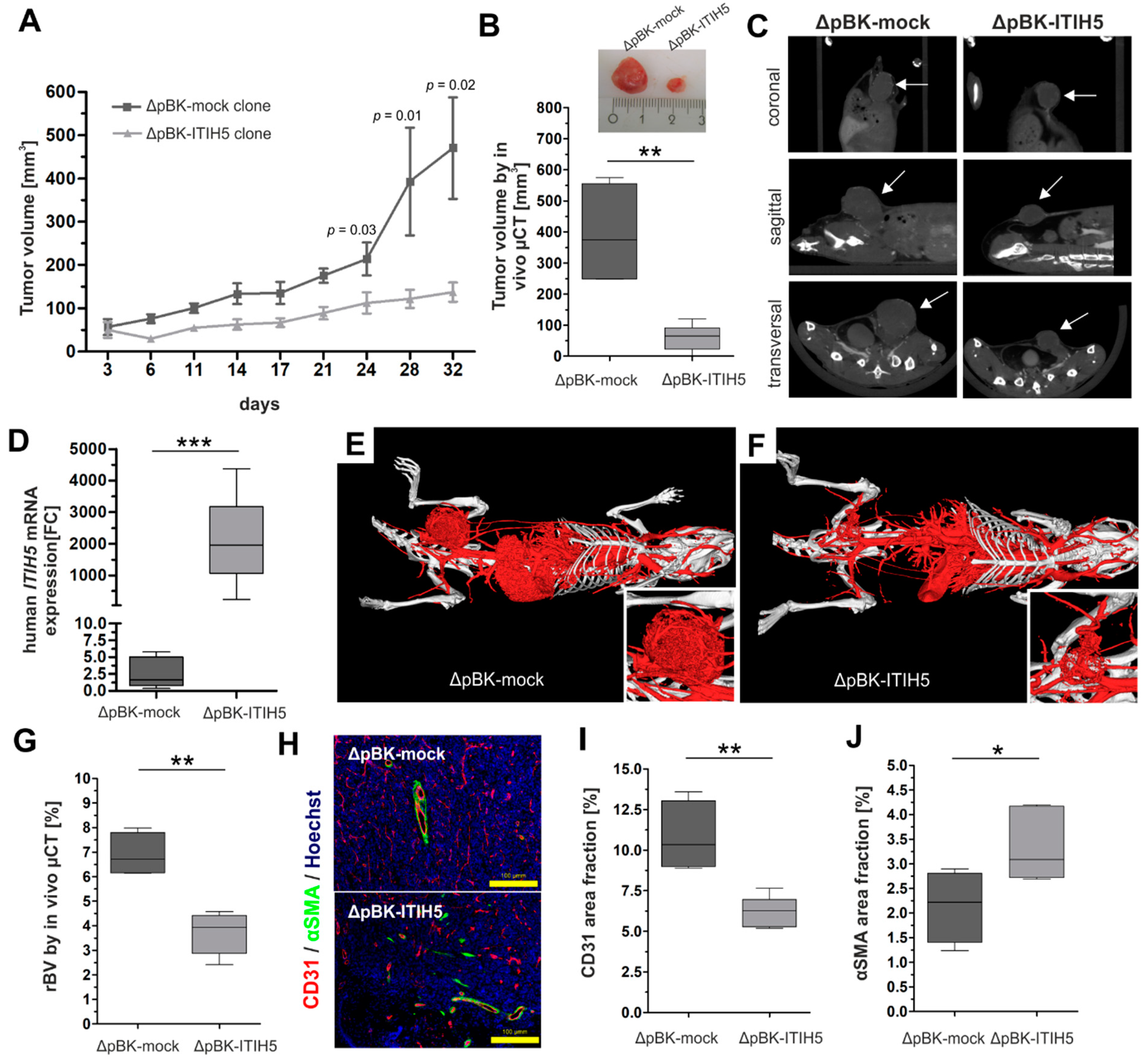
2.1. Full-Length ITIH5 Inhibited the Cell Growth of TNBC Breast Cancer Cells In Vivo
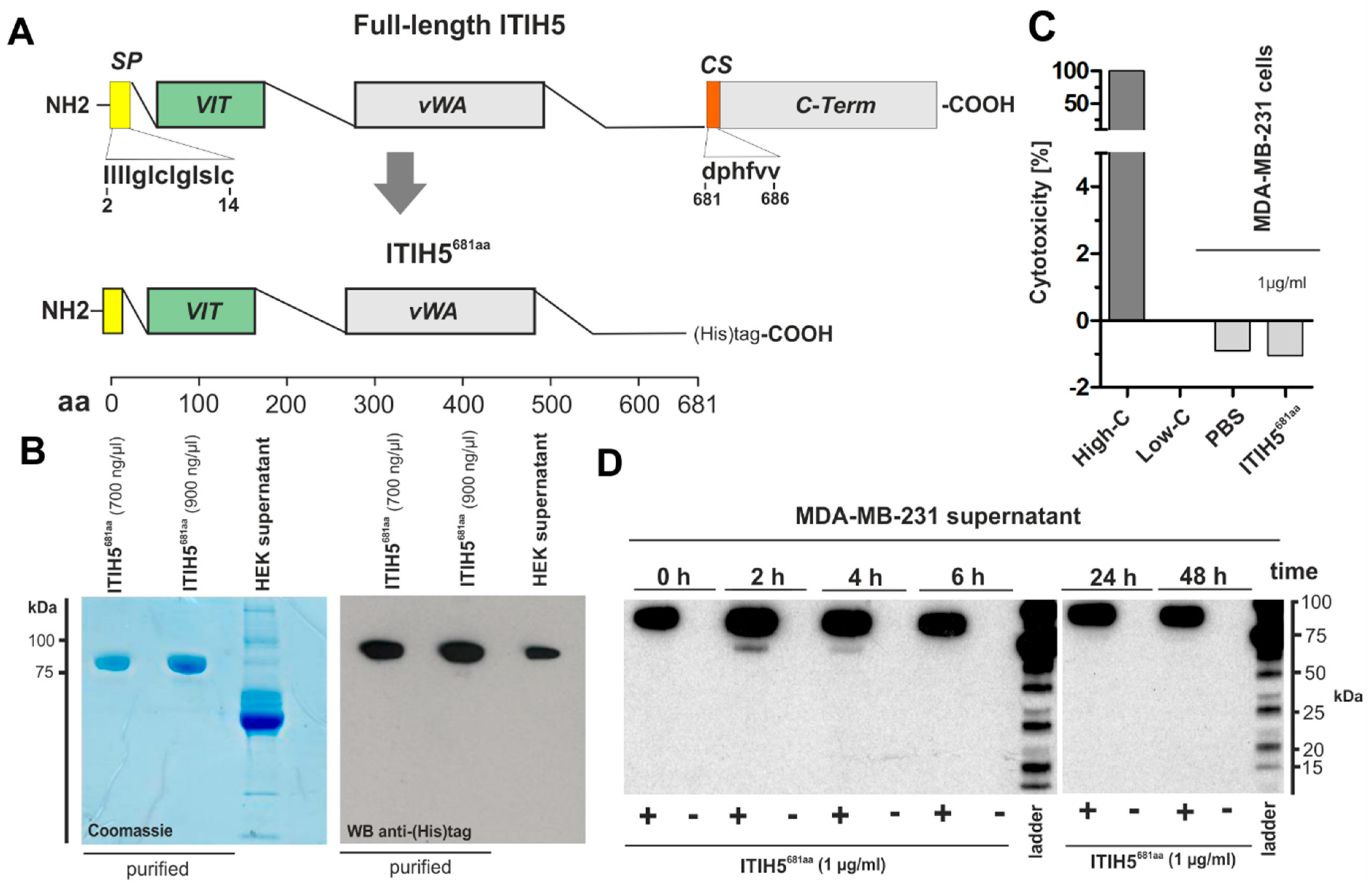
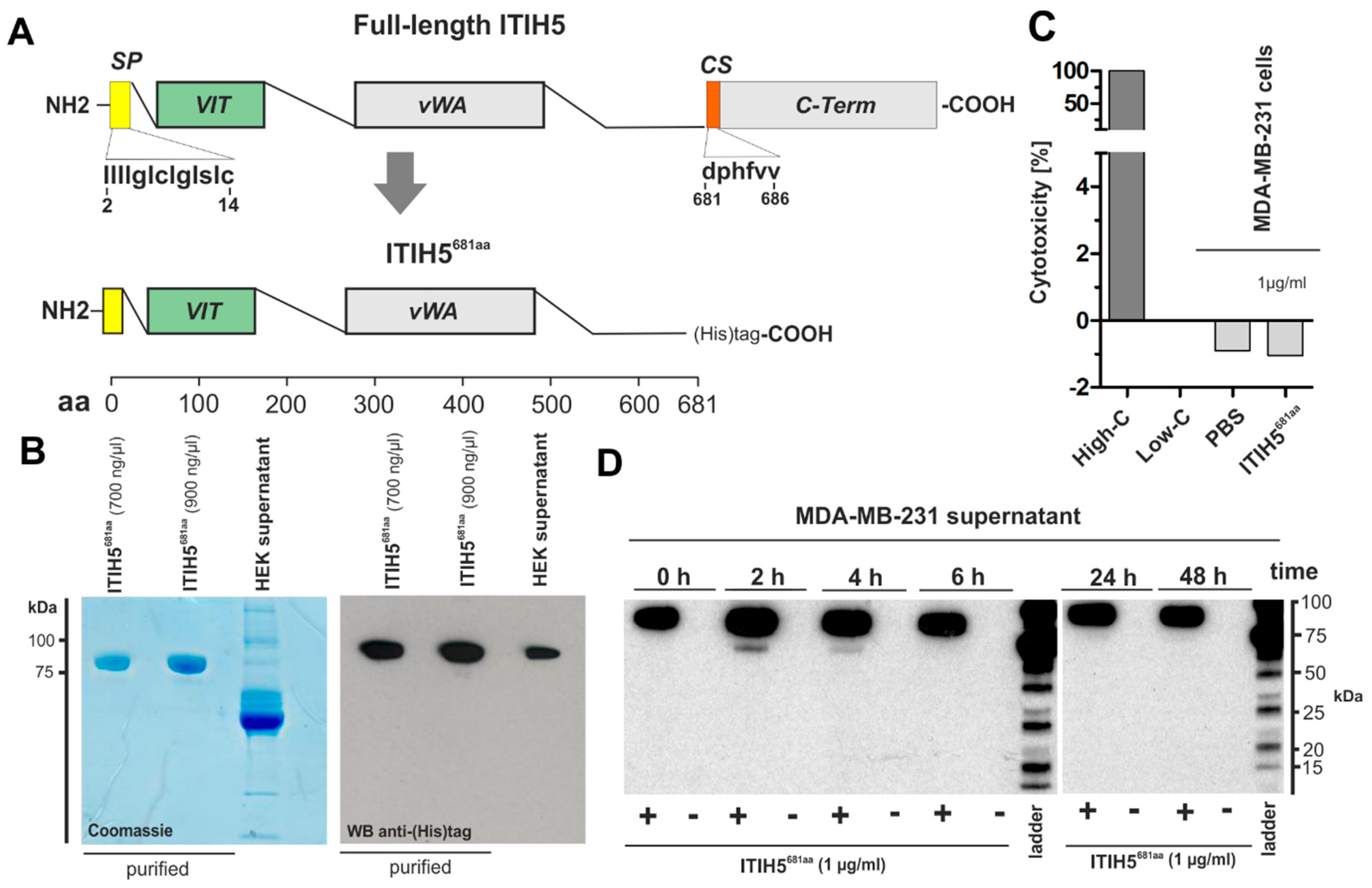
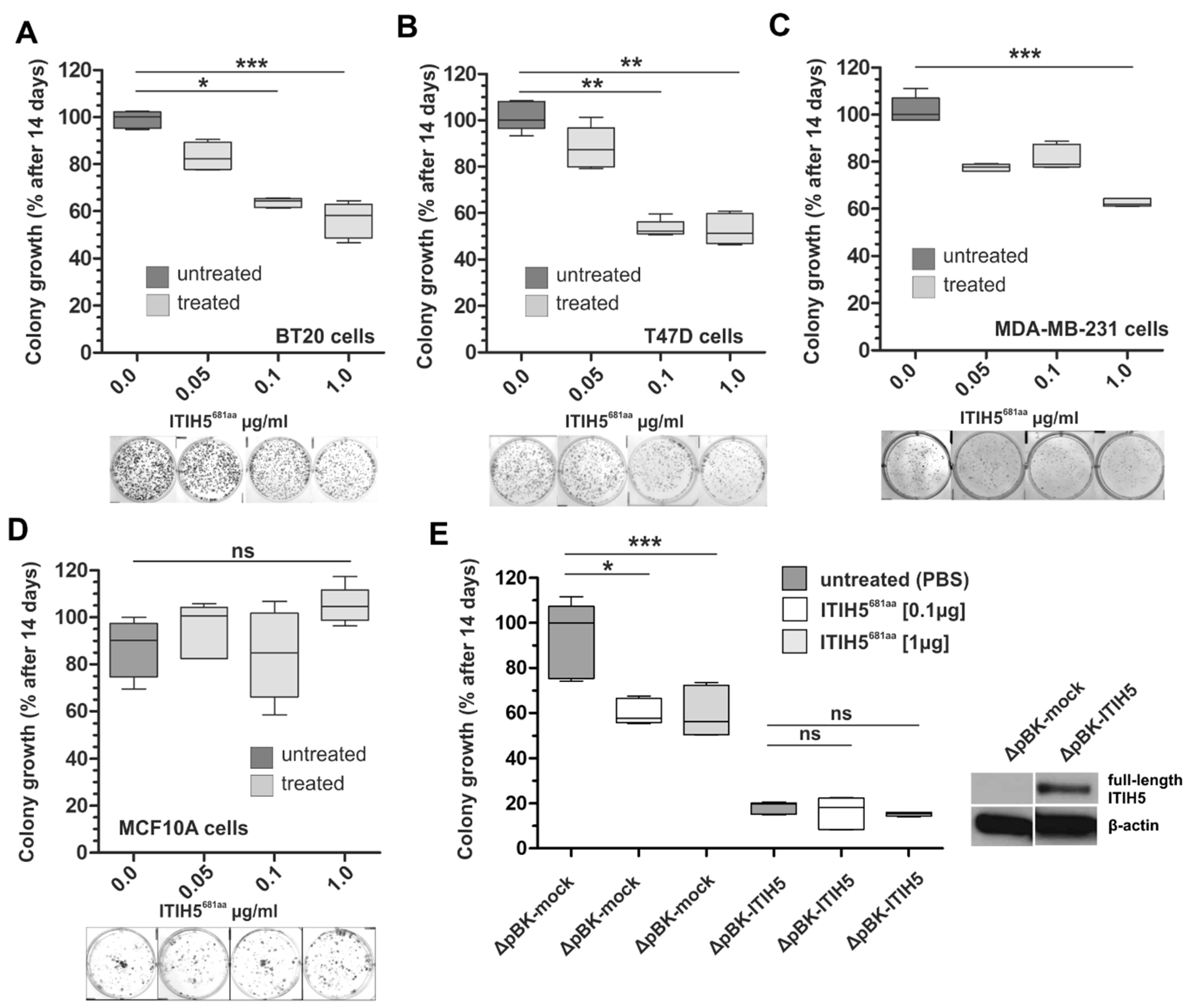
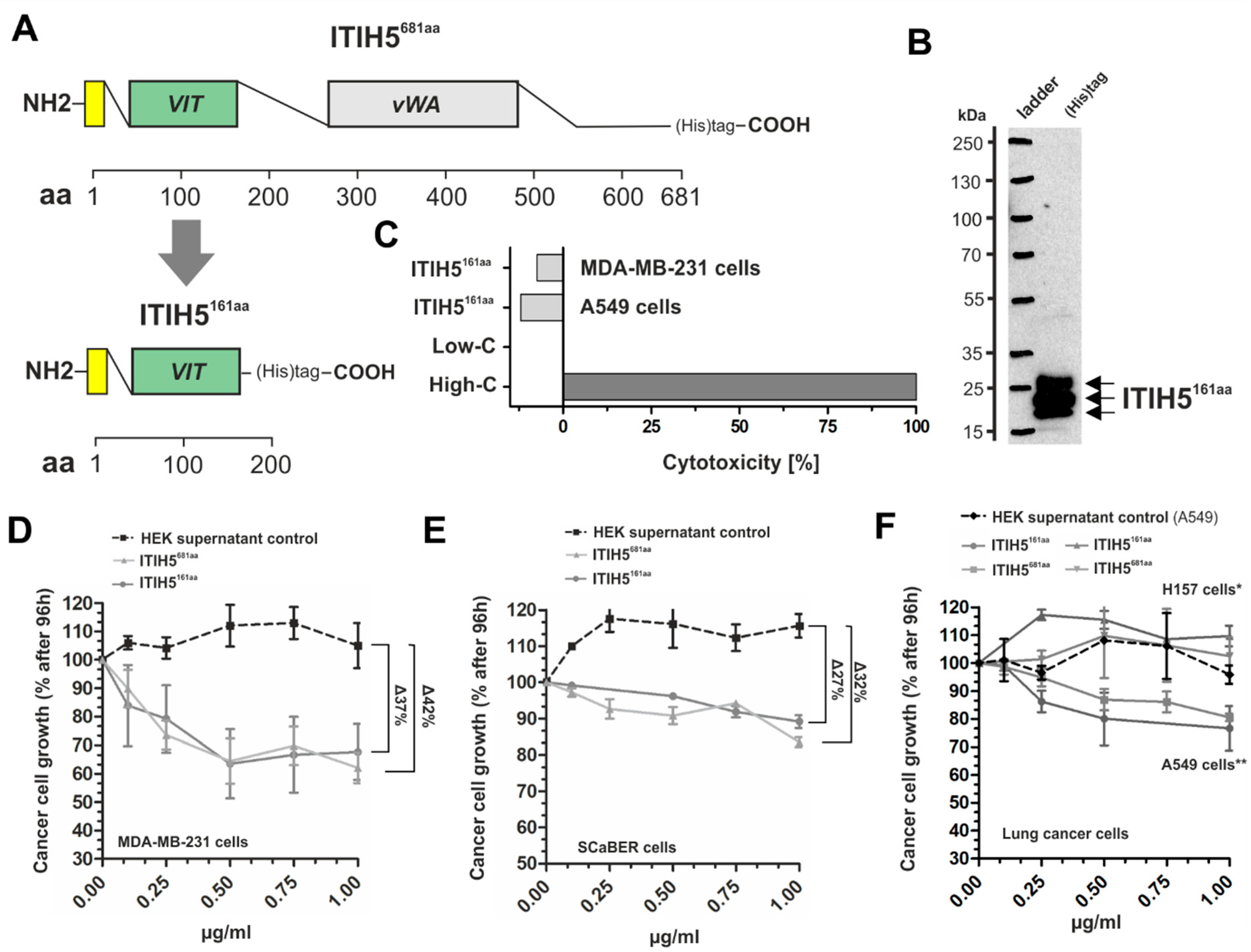
2.2. ITIH5 Protein Downsized to the Secreted N-Terminal Part without the HA-Binding Function Inhibited Cancer Cell and Colony Growth In Vitro
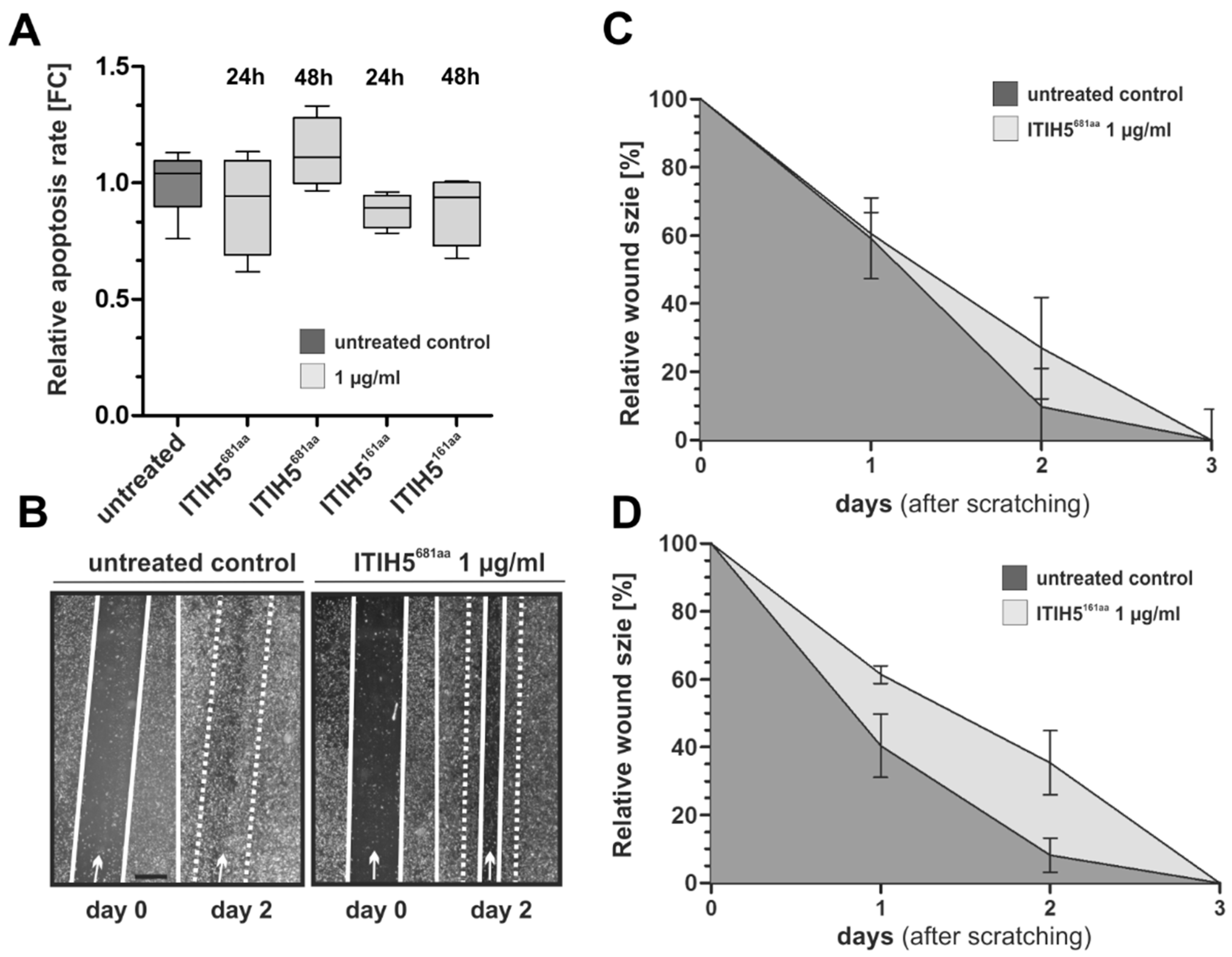
2.3. ITIH5 Downsized to the VIT Domain Was Sufficient and Responsible for Cancer Cell Growth Inhibition, Similar to the N-Terminal Protein
3. Discussion
3.1. Animal Model Showing ITIH5-Mediated Tumor Cell Growth Inhibition
3.2. ITIH5 VIT Domain Applied Extracellularly Is Sufficient to Mediate Tumor Cell Growth
4. Materials and Methods
4.1. Wild-Type and Genetically Modified Cell Lines
4.2. Animals
4.3. Construction and Generation of Truncated Recombinant ITIH5-Derived Proteins
4.4. Nucleic Acid Extraction and Reverse Transcription PCR
4.5. Real-Time qPCR
4.6. Two-Dimensional (2D) Gel Electrophoresis (2-DIGE)
4.7. Coomassie Blue Staining
4.8. Protein Verification by Western Blotting
4.9. Protein Identification by MALDI-TOF MS/MS
4.10. Cytotoxicity Assay
4.11. Short-Term Drug Response Assay
4.12. Clonogenic Survival Assay
4.13. Apoptosis Assay
4.14. Wound Healing Assay
4.15. In Vivo Mouse Model
4.16. Fluorescence Imaging
4.17. µCT Imaging
4.18. Statistical Data Acquisition
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Himmelfarb, M.; Klopocki, E.; Grube, S.; Staub, E.; Klaman, I.; Hinzmann, B.; Kristiansen, G.; Rosenthal, A.; Durst, M.; Dahl, E. ITIH5, a novel member of the inter-alpha-trypsin inhibitor heavy chain family is downregulated in breast cancer. Cancer Lett. 2004, 204, 69–77. [Google Scholar] [CrossRef]
- Veeck, J.; Chorovicer, M.; Naami, A.; Breuer, E.; Zafrakas, M.; Bektas, N.; Dürst, M.; Kristiansen, G.; Wild, P.J.; Hartmann, A.; et al. The extracellular matrix protein ITIH5 is a novel prognostic marker in invasive node-negative breast cancer and its aberrant expression is caused by promoter hypermethylation. Oncogene 2008, 27, 865–876. [Google Scholar] [CrossRef] [Green Version]
- Rose, M.; Gaisa, N.T.; Antony, P.; Fiedler, D.; Heidenreich, A.; Otto, W.; Denzinger, S.; Bertz, S.; Hartmann, A.; Karl, A.; et al. Epigenetic inactivation of ITIH5 promotes bladder cancer progression and predicts early relapse of pT1 high-grade urothelial tumours. Carcinogenesis 2014, 35, 727–736. [Google Scholar] [CrossRef] [Green Version]
- Kloten, V.; Rose, M.; Kaspar, S.; von Stillfried, S.; Knüchel, R.; Dahl, E. Epigenetic inactivation of the novel candidate tumor suppressor gene ITIH5 in colon cancer predicts unfavorable overall survival in the CpG island methylator phenotype. Epigenetics 2014, 9, 1290–1301. [Google Scholar] [CrossRef] [Green Version]
- Dötsch, M.M.; Kloten, V.; Schlensog, M.; Heide, T.; Braunschweig, T.; Veeck, J.; Petersen, I.; Knüchel, R.; Dahl, E. Low expression of ITIH5 in adenocarcinoma of the lung is associated with unfavorable patients’ outcome. Epigenetics 2015, 10, 903–912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dittmann, J.; Ziegfeld, A.; Jansen, L.; Gajda, M.; Kloten, V.; Dahl, E.; Runnebaum, I.B.; Dürst, M.; Backsch, C. Gene expression analysis combined with functional genomics approach identifies ITIH5 as tumor suppressor gene in cervical carcinogenesis. Mol. Carcinog. 2017, 56, 1578–1589. [Google Scholar] [CrossRef]
- Rose, M.; Kloten, V.; Noetzel, E.; Gola, L.; Ehling, J.; Heide, T.; Meurer, S.K.; Gaiko-Shcherbak, A.; Sechi, A.S.; Huth, S.; et al. ITIH5 mediates epigenetic reprogramming of breast cancer cells. Mol. Cancer 2017, 16, 44. [Google Scholar] [CrossRef] [Green Version]
- Rose, M.; Meurer, S.K.; Kloten, V.; Weiskirchen, R.; Denecke, B.; Antonopoulos, W.; Deckert, M.; Knüchel, R.; Dahl, E. ITIH5 induces a shift in TGF-β superfamily signaling involving Endoglin and reduces risk for breast cancer metastasis and tumor death. Mol. Carcinog. 2018, 57, 167–181. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Cao, F.; Li, X.; Zhang, L.; Liu, Z.; Li, X.; Lin, J.; Han, C. ITIH5, a p53-responsive gene, inhibits the growth and metastasis of melanoma cells by downregulating the transcriptional activity of KLF4. Cell Death Dis. 2021, 12, 438. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, K.; Kurahara, H.; Young, E.D.; Natsugoe, S.; Ijichi, A.; Iwakuma, T.; Welch, D.R. Genome-wide in vivo RNAi screen identifies ITIH5 as a metastasis suppressor in pancreatic cancer. Clin. Exp. Metastasis 2017, 34, 229–239. [Google Scholar] [CrossRef] [PubMed]
- Zou, L.; He, H.; Li, Z.; Chen, O.; Jia, X.; Zhang, H. Long noncoding RNA LINC00261 upregulates ITIH5 to impair tumorigenic ability of pancreatic cancer stem cells. Cell Death Discov. 2021, 7, 220. [Google Scholar] [CrossRef] [PubMed]
- Martin, J.; Midgley, A.; Meran, S.; Woods, E.; Bowen, T.; Phillips, A.O.; Steadman, R. Tumor Necrosis Factor-stimulated Gene 6 (TSG-6)-mediated Interactions with the Inter-α-inhibitor Heavy Chain 5 Facilitate Tumor Growth Factor β1 (TGFβ1)-dependent Fibroblast to Myofibroblast Differentiation. J. Biol. Chem. 2016, 291, 13789–13801. [Google Scholar] [PubMed] [Green Version]
- Huth, S.; Huth, L.; Marquardt, Y.; Fietkau, K.; Dahl, E.; Esser, P.R.; Martin, S.F.; Heise, R.; Merk, H.F.; Baron, J.M. Inter-α-Trypsin Inhibitor Heavy Chain 5 (ITIH5) Is a Natural Stabilizer of Hyaluronan That Modulates Biological Processes in the Skin. Ski. Pharm. Physiol. 2020, 33, 198–206. [Google Scholar] [CrossRef] [PubMed]
- Salier, J.P.; Rouet, P.; Raguenez, G.; Daveau, M. The inter-alpha-inhibitor family: From structure to regulation. Biochem. J. 1996, 315, 1–9. [Google Scholar] [CrossRef]
- Héron, A.; Bourguignon, J.; Callé, A.; Borghi, H.; Sesboüé, R.; Diarra-Mehrpour, M.; Martin, J.P. Post-translational processing of the inter-alpha-trypsin inhibitor in the human hepatoma HepG2 cell line. Biochem. J. 1994, 302, 573–580. [Google Scholar] [CrossRef] [PubMed]
- Bost, F.; Diarra-Mehrpour, M.; Martin, J.P. Inter-alpha-trypsin inhibitor proteoglycan family—A group of proteins binding and stabilizing the extracellular matrix. Eur. J. Biochem. 1998, 252, 339–346. [Google Scholar] [CrossRef]
- Zhuo, L.S.; Hascall, V.C.; Kimata, K. Inter-alpha-trypsin inhibitor, a covalent protein-glycosaminoglycan-protein complex. J. Biol. Chem. 2004, 279, 38079–38082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rose, M.; Noetzel, E.; Kistermann, J.; Eschenbruch, J.; Rushrush, S.; Gan, L.; Knüchel, R.; Gaisa, N.T.; Dahl, E. The ECM Modulator ITIH5 Affects Cell Adhesion, Motility and Chemotherapeutic Response of Basal/Squamous-Like (BASQ) Bladder Cancer Cells. Cells 2021, 10, 1038. [Google Scholar] [CrossRef]
- Zhuo, L.; Kimata, K. Structure and Function of Inter-alpha-Trypsin Inhibitor Heavy Chains. Connect. Tissue Res. 2008, 49, 311–320. [Google Scholar] [CrossRef]
- Weidle, U.H.; Birzele, F.; Tiefenthaler, G. Potential of Protein-based Anti-metastatic Therapy with Serpins and Inter α-Trypsin Inhibitors. Cancer Genom. Proteom. 2018, 15, 225–238. [Google Scholar] [CrossRef] [Green Version]
- Ehling, J.; Theek, B.; Gremse, F.; Baetke, S.; Möckel, D.; Maynard, J.; Ricketts, S.A.; Grüll, H.; Neeman, M.; Knuechel, R.; et al. Micro-CT imaging of tumor angiogenesis: Quantitative measures describing micromorphology and vascularization. Am. J. Pathol. 2014, 184, 431–441. [Google Scholar] [CrossRef]
- De Lisser, H.M.; Christofidou-Solomidou, M.; Strieter, R.M.; Burdick, M.D.; Robinson, C.S.; Wexler, R.S.; Kerr, J.S.; Garlanda, C.; Merwin, J.R.; Madri, J.A.; et al. Involvement of endothelial PECAM-1/CD31 in angiogenesis. Am. J. Pathol. 1997, 151, 671–677. [Google Scholar]
- Zhu, K.; Zhao, J.; Lubman, D.M.; Miller, F.R.; Barder, T.J. Protein pI shifts due to posttranslational modifications in the separation and characterization of proteins. Anal. Chem. 2005, 77, 2745–2755. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.F.; Wootton, J.C.; Gertz, E.M.; Agarwala, R.; Morgulis, A.; Schäffer, A.A.; Yu, Y.K. Protein database searches using compositionally adjusted substitution matrices. FEBS J. 2005, 72, 5101–5109. [Google Scholar] [CrossRef] [PubMed]
- Lumachi, F.; Brunello, A.; Maruzzo, M.; Basso, U.; Basso, S.M.M. Treatment of estrogen receptor-positive breast cancer. Curr. Med. Chem. 2013, 20, 596–604. [Google Scholar] [CrossRef]
- Turner, N.C.; Neven, P.; Loibl, S.; Andre, F. Advances in the treatment of advanced oestrogen-receptor-positive breast cancer. Lancet 2017, 389, 2403–2414. [Google Scholar] [CrossRef]
- Yin, L.; Duan, J.J.; Bian, X.W.; Yu, S.C. Triple-negative breast cancer molecular subtyping and treatment progress. Breast Cancer Res. 2020, 22, 61. [Google Scholar] [CrossRef] [PubMed]
- Stenehjem, D.D.; Tran, D.; Nkrumah, M.A.; Gupta, S. PD1/PDL1 inhibitors for the treatment of advanced urothelial bladder cancer. OncoTargets Ther. 2018, 11, 5973–5989. [Google Scholar] [CrossRef] [Green Version]
- Emens, L.A. Breast Cancer Immunotherapy: Facts and Hopes. Clin. Cancer Res. 2018, 24, 511–520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michel, L.L.; von Au, A.; Mavratzas, A.; Smetanay, K.; Schütz, F.; Schneeweiss, A. Immune Checkpoint Blockade in Patients with Triple-Negative Breast Cancer. Target. Oncol. 2020, 15, 415–428. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Beltran, A.; Cimadamore, A.; Blanca, A.; Massari, F.; Vau, N.; Scarpelli, M.; Cheng, L.; Montironi, R. Immune Checkpoint Inhibitors for the Treatment of Bladder Cancer. Cancers 2021, 13, 131. [Google Scholar] [CrossRef] [PubMed]
- Galsky, M.D.; Arija, J.A.A.; Bamias, A.; Davis, I.D.; de Santis, M.; Kikuchi, E.; Garcia-Del-Muro, X.; de Giorgi, U.; Mencinger, M.; Izumi, K.; et al. Atezolizumab with or without chemotherapy in metastatic urothelial cancer (IMvigor130): A multicentre, randomised, placebo-controlled phase 3 trial. Lancet 2020, 395, 1547–1557. [Google Scholar] [CrossRef]
- Lee, S.W.; Tomasetto, C.; Sager, R. Positive selection of candidate tumor-suppressor genes by subtractive hybridization. Proc Natl. Acad. Sci. USA 1991, 88, 2825–2829. [Google Scholar] [CrossRef] [Green Version]
- Young, E.D.; Manley, S.J.; Beadnell, T.C.; Shearin, A.E.; Sasaki, K.; Zimmerman, R.; Kauffman, E.; Vivian, C.J.; Parasuram, A.; Iwakuma, T.; et al. Suppression of pancreatic cancer liver metastasis by secretion-deficient ITIH5. Br. J. Cancer 2021, 124, 166–175. [Google Scholar] [CrossRef] [PubMed]
- Ruggiero, F.; Koch, M. Making recombinant extracellular matrix proteins. Methods 2008, 45, 75–85. [Google Scholar] [CrossRef]
- Wei, Z.; Yang, Y.; Zhang, P.; Andrianakos, R.; Hasegawa, K.; Lyu, J.; Chen, X.; Bai, G.; Liu, C.; Pera, M.; et al. Klf4 Interacts Directly with Oct4 and Sox2 to Promote Reprogramming. Stem Cells 2009, 27, 2969–2978. [Google Scholar] [CrossRef]
- Yu, F.; Li, J.; Chen, H.; Fu, J.; Ray, S.; Huang, S.; Zheng, H.; Ai, W. Kruppel-like factor 4 (KLF4) is required for maintenance of breast cancer stem cells and for cell migration and invasion. Oncogene 2011, 30, 2161–2172. [Google Scholar] [CrossRef] [Green Version]
- Hamm, A.; Veeck, J.; Bektas, N.; Wild, P.J.; Hartmann, A.; Heindrichs, U.; Kristiansen, G.; Werbowetski-Ogilvie, T.; Del Maestro, R.; Knuechel, R.; et al. Frequent expression loss of Inter-alpha-trypsin inhibitor heavy chain (ITIH) genes in multiple human solid tumors: A systematic expression analysis. BMC Cancer 2008, 8, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cerami, E.; Gao, J.; Dogrusoz, U.; Gross, B.E.; Sumer, S.O.; Aksoy, B.A.; Jacobsen, A.; Byrne, C.J.; Heuer, M.L.; Larsson, E.; et al. The cBio cancer genomics portal: An open platform for exploring multidimensional cancer genomics data. Cancer Discov. 2012, 2, 401–404. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.; Aksoy, B.A.; Dogrusoz, U.; Dresdner, G.; Gross, B.; Sumer, S.O.; Sun, Y.; Jacobsen, A.; Sinha, R.; Larsson, E.; et al. Integrative analysis of complex cancer genomics and clinical profiles using the cBioPortal. Sci. Signal 2013, 6, pl1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Labbus, K.; Henning, M.; Borkham-Kamphorst, E.; Geisler, C.; Berger, T.; Mak, T.W.; Knüchel, R.; Meyer, H.E.; Weiskirchen, R.; Henkel, C. Proteomic profiling in Lipocalin 2 deficient mice under normal and inflammatory conditions. J. Proteomics 2013, 78, 188–196. [Google Scholar] [CrossRef] [PubMed]
- Perkins, D.N.; Pappin, D.J.; Creasy, D.M.; Cottrell, J.S. Probability-based protein identification by searching sequence databases using mass spectrometry data. Electrophoresis 1999, 20, 3551–3567. [Google Scholar] [CrossRef]
- Tugues, S.; Honjo, S.; König, C.; Noguer, O.; Hedlund, M.; Botling, J.; Deschoemaeker, S.; Wenes, M.; Rolny, C.; Jahnen-Dechent, W.; et al. Genetic de_ciency in plasma protein HRG enhances tumor growth and metastasis by exacerbating immune escape and vessel abnormalization. Cancer Res. 2012, 72, 1953–1963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loening, A.M.; Gambhir, S.S. AMIDE: A free software tool for multimodality medical image analysis. Mol. Imaging. 2003, 2, 131–137. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Aligned Sequences | Expected | Method | Identities | Positives | Gaps |
|---|---|---|---|---|---|
| Matching ITIH5 VIT (56–161 aa)–ITIH1 VIT (58–166 aa) | 2 × 10−10 | Compositional matrix adjustment | 29/111 (26%) | 55/111 (49%) | 7/111 (6%) |
| Matching ITIH5 VIT (51–114 aa)–ITIH2 VIT (72–135 aa) | 7 × 10−14 | Compositional matrix adjustment | 23/64 (36%) | 41/64 (64%) | 0/64 (0%) |
| Matching ITIH5 VIT (56–161 aa)-ITIH3 VIT (50–158 aa) | 6 × 10−16 | Compositional matrix adjustment | 34/109 (31%) | 56/109 (51%) | 3/109 (2%) |
| Matching ITIH5 VIT (54–161 aa)–ITIH3 VIT (38–148 aa) | 1 × 10−17 | Compositional matrix adjustment | 41/112 (37%) | 59/112 (52%) | 5/112 (4%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rose, M.; Huth, S.; Wiesehöfer, M.; Ehling, J.; Henkel, C.; Steitz, J.; Lammers, T.; Kistermann, J.; Klaas, O.; Koch, M.; et al. ITIH5-Derived Polypeptides Covering the VIT Domain Suppress the Growth of Human Cancer Cells In Vitro. Cancers 2022, 14, 488. https://doi.org/10.3390/cancers14030488
Rose M, Huth S, Wiesehöfer M, Ehling J, Henkel C, Steitz J, Lammers T, Kistermann J, Klaas O, Koch M, et al. ITIH5-Derived Polypeptides Covering the VIT Domain Suppress the Growth of Human Cancer Cells In Vitro. Cancers. 2022; 14(3):488. https://doi.org/10.3390/cancers14030488
Chicago/Turabian StyleRose, Michael, Sebastian Huth, Marc Wiesehöfer, Josef Ehling, Corinna Henkel, Julia Steitz, Twan Lammers, Jennifer Kistermann, Oliver Klaas, Maximilian Koch, and et al. 2022. "ITIH5-Derived Polypeptides Covering the VIT Domain Suppress the Growth of Human Cancer Cells In Vitro" Cancers 14, no. 3: 488. https://doi.org/10.3390/cancers14030488
APA StyleRose, M., Huth, S., Wiesehöfer, M., Ehling, J., Henkel, C., Steitz, J., Lammers, T., Kistermann, J., Klaas, O., Koch, M., Rushrush, S., Knüchel, R., & Dahl, E. (2022). ITIH5-Derived Polypeptides Covering the VIT Domain Suppress the Growth of Human Cancer Cells In Vitro. Cancers, 14(3), 488. https://doi.org/10.3390/cancers14030488