
Comprehensive Analysis of DMRT3 as a Potential Biomarker Associated with the Immune Infiltration in a Pan-Cancer Analysis and Validation in Lung Adenocarcinoma
 ,
,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Analysis of the DMRT3 Expression in Pan-Cancer
2.2. Correlation Analysis of DMRT3 and Prognosis in Pan-Cancer
2.3. Correlation of the DMRT3 Expression with the Stage in Pan-Cancer
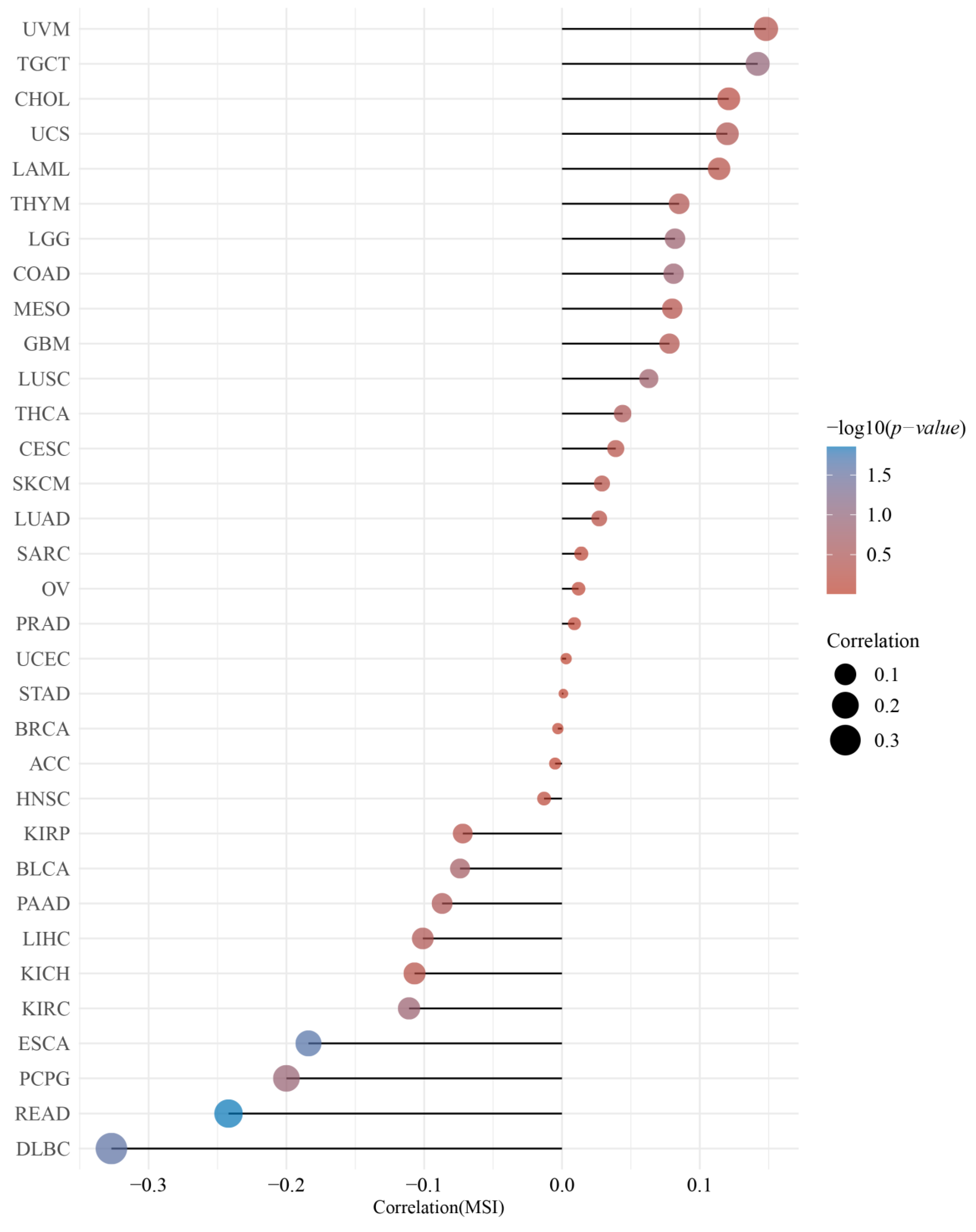
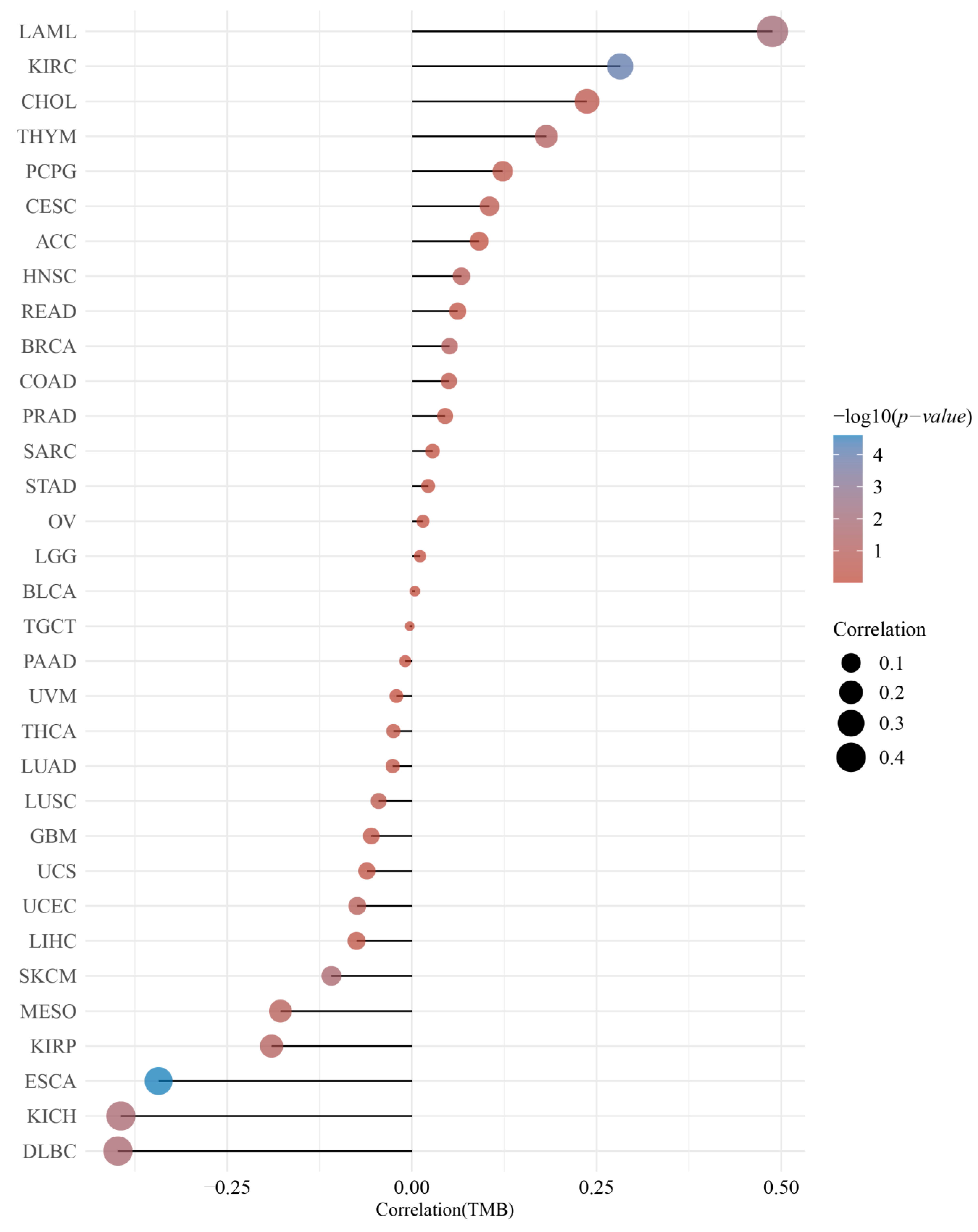
2.4. Correlation of the DMRT3 Expression with TMB and MSI in Pan-Cancer
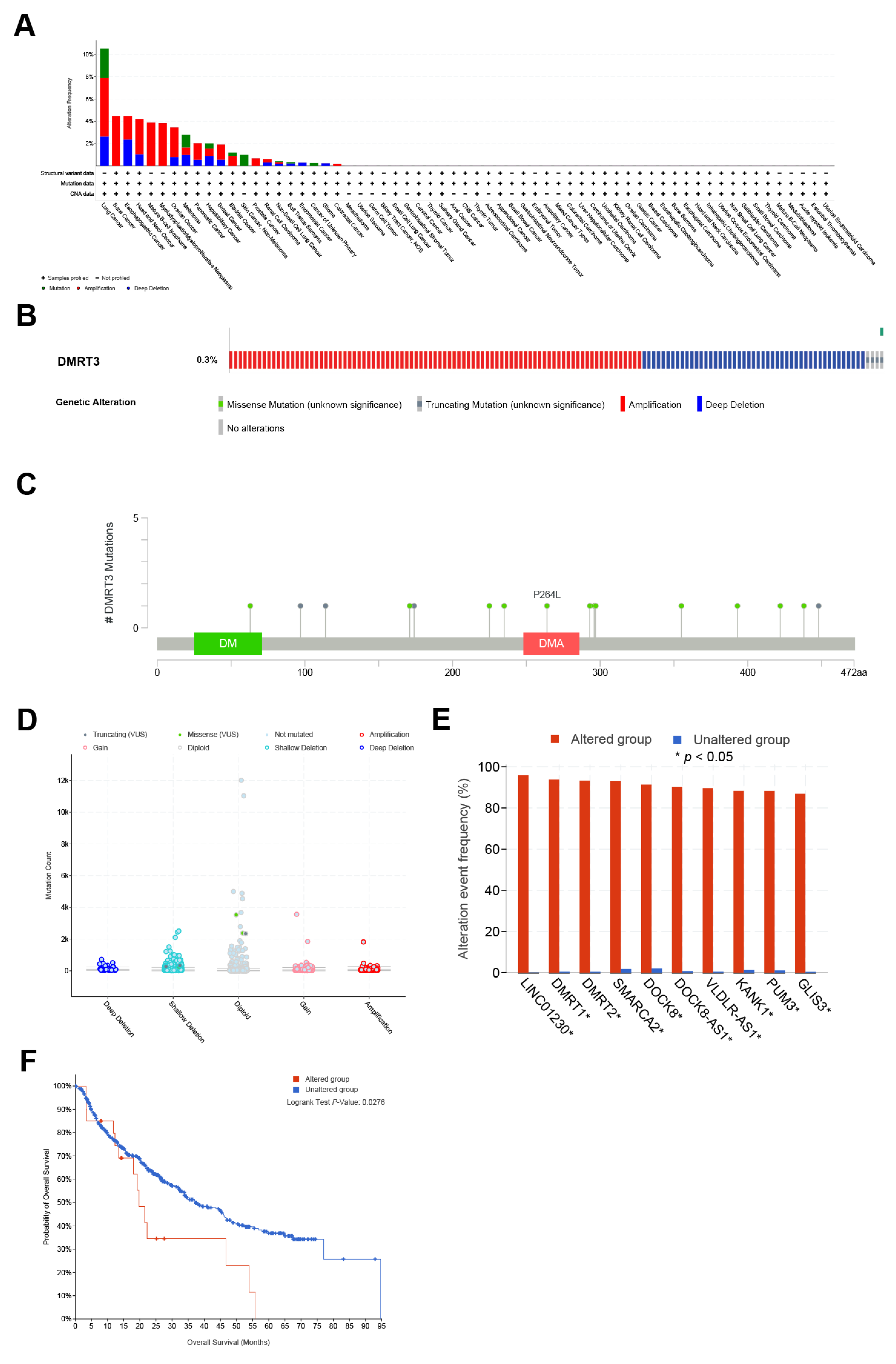
2.5. Genomic Alterations of DMRT3 in Pan-Cancer
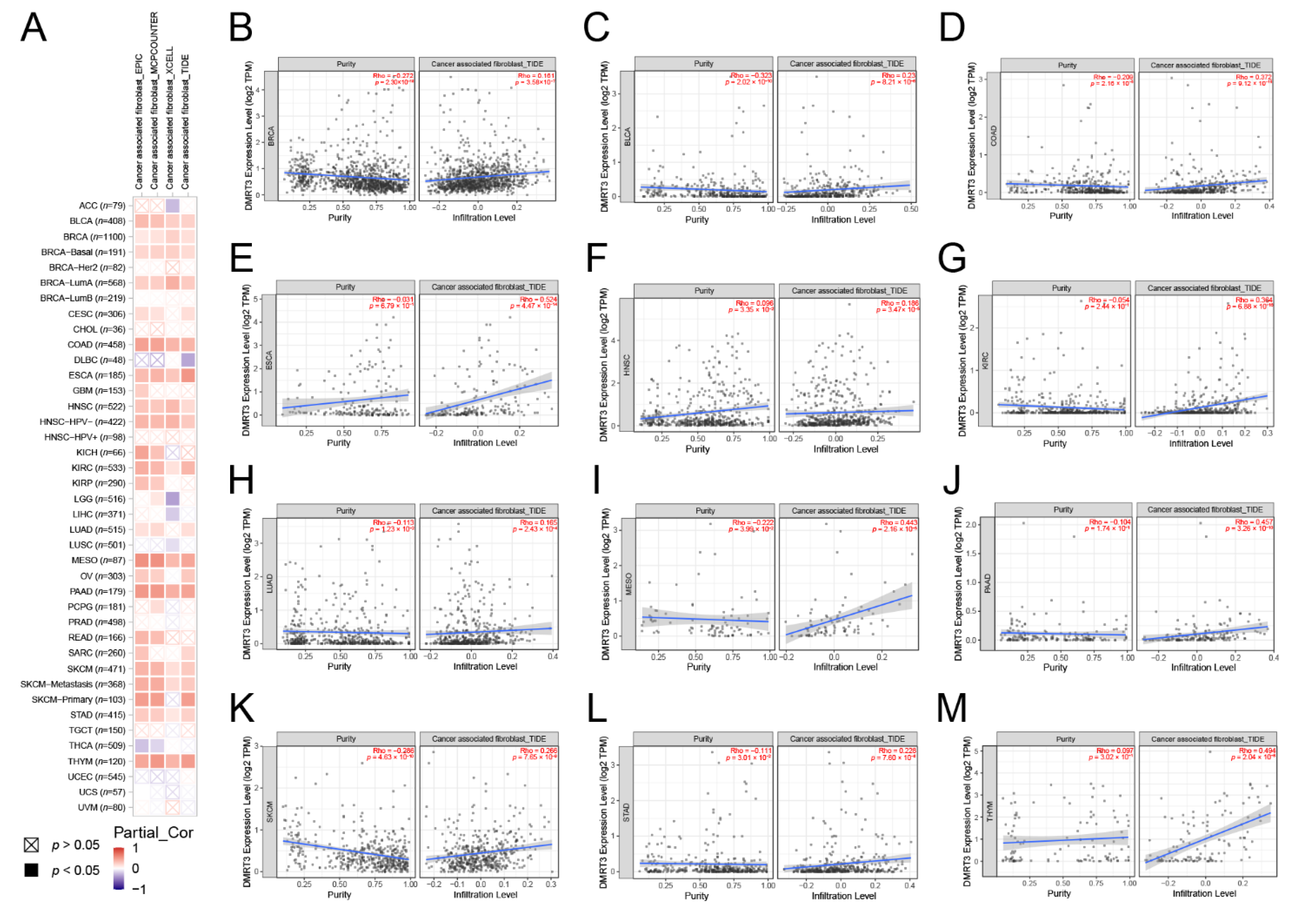
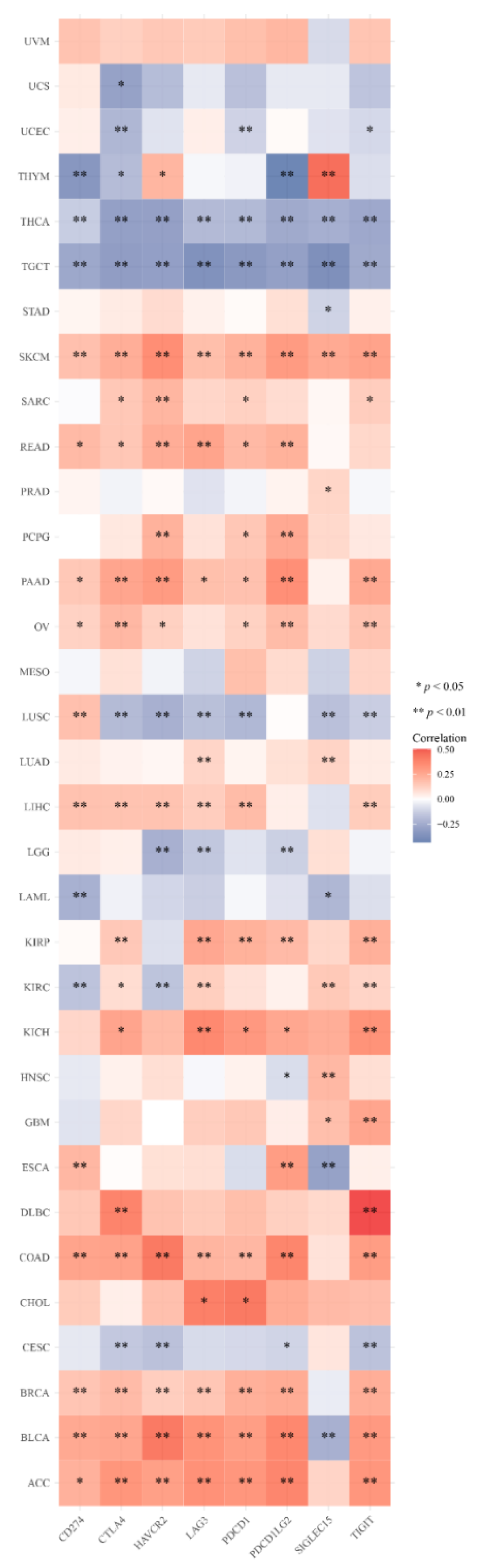
2.6. Analysis of the Association between the DMRT3 Expression and the Tumor Immune Microenvironment in Pan-Cancer
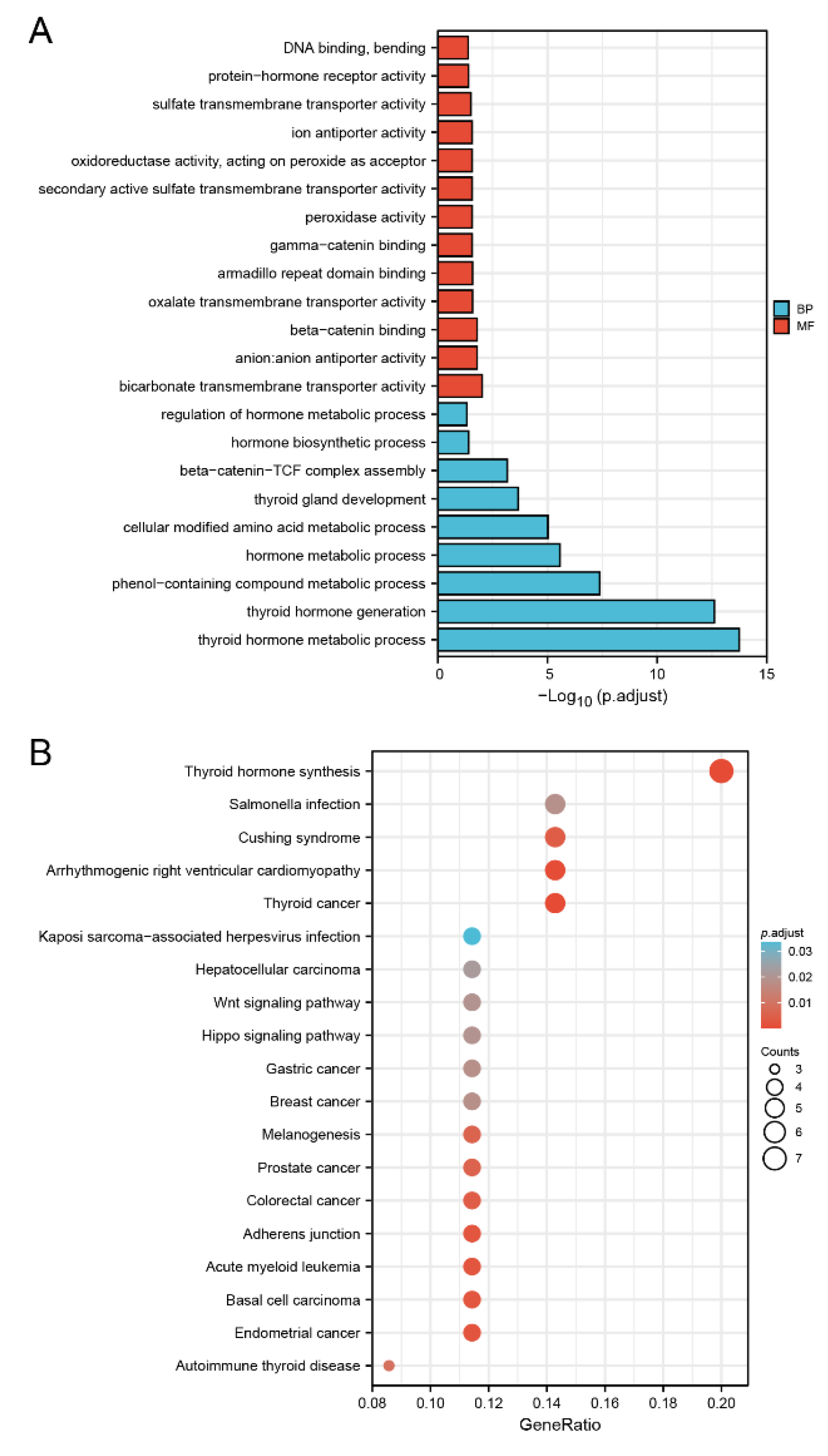
2.7. DMRT3-Related Functional Enrichment Analysis
2.8. Drug Sensitivity of DMRT3 in Pan-Cancer
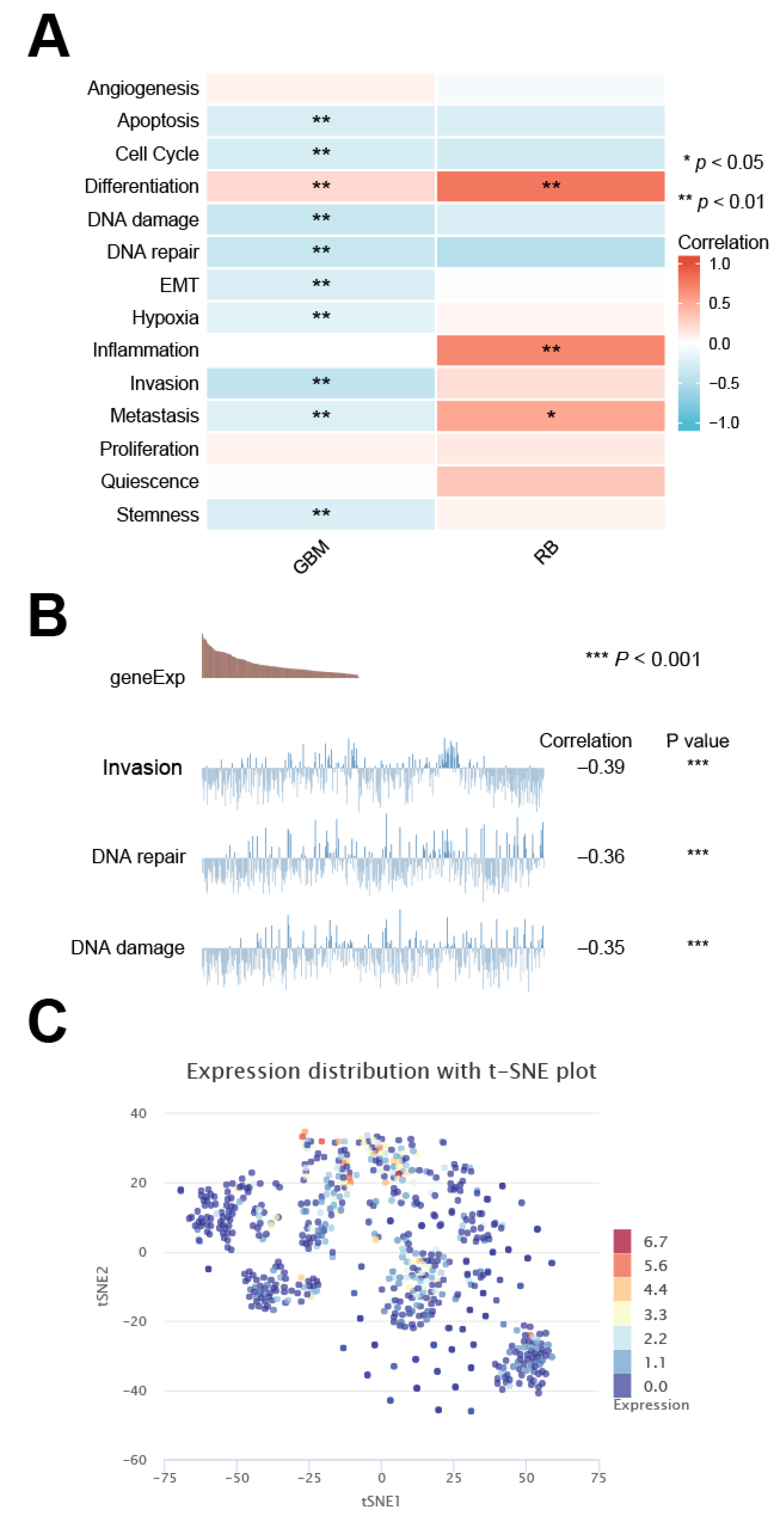
2.9. Single Cell Sequencing Data Analysis
2.10. Conservation Analysis of DMRT3
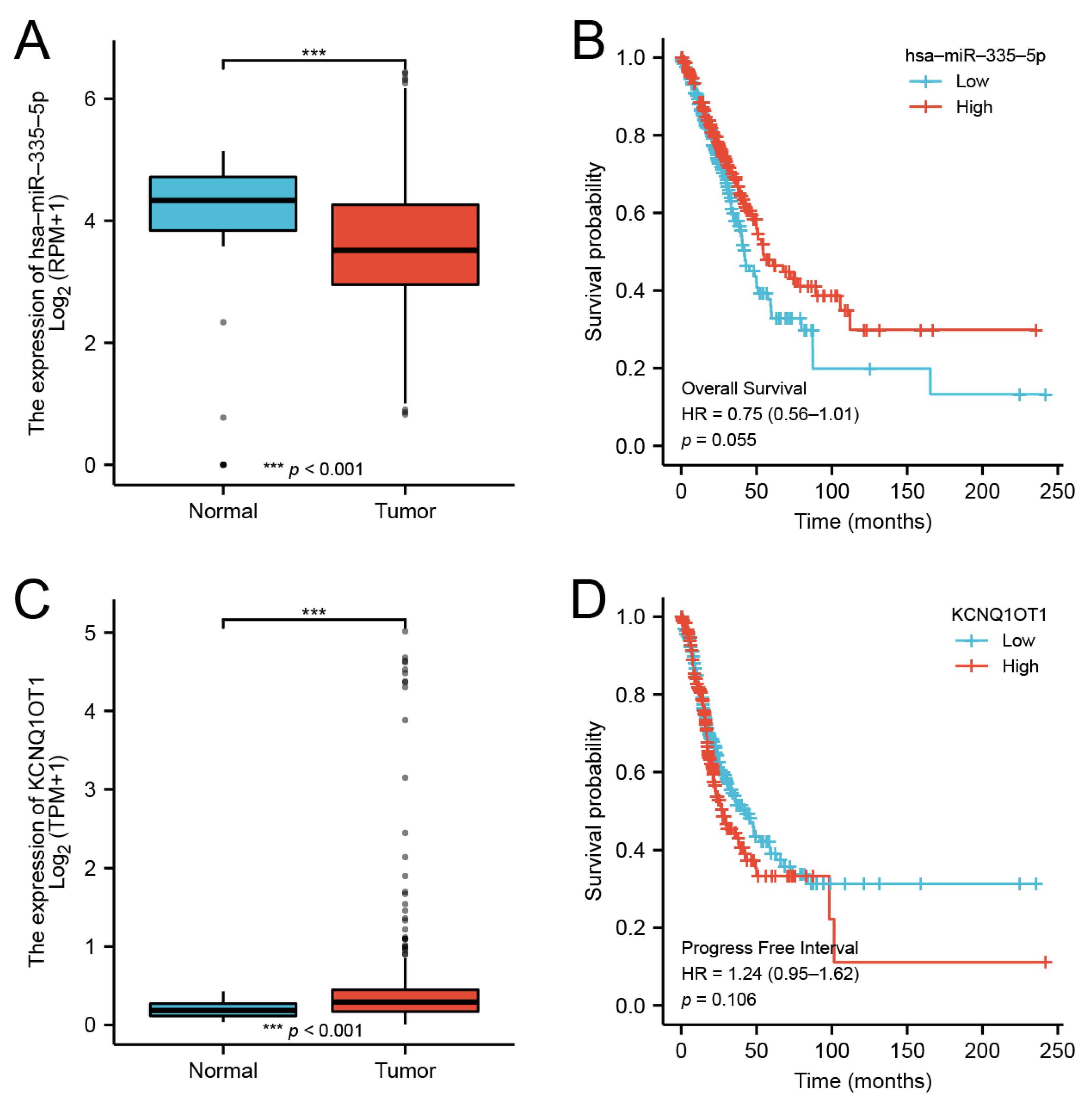
2.11. CeRNA Network Construction
2.12. QRT-PCR
2.13. Statistical Analysis
3. Results
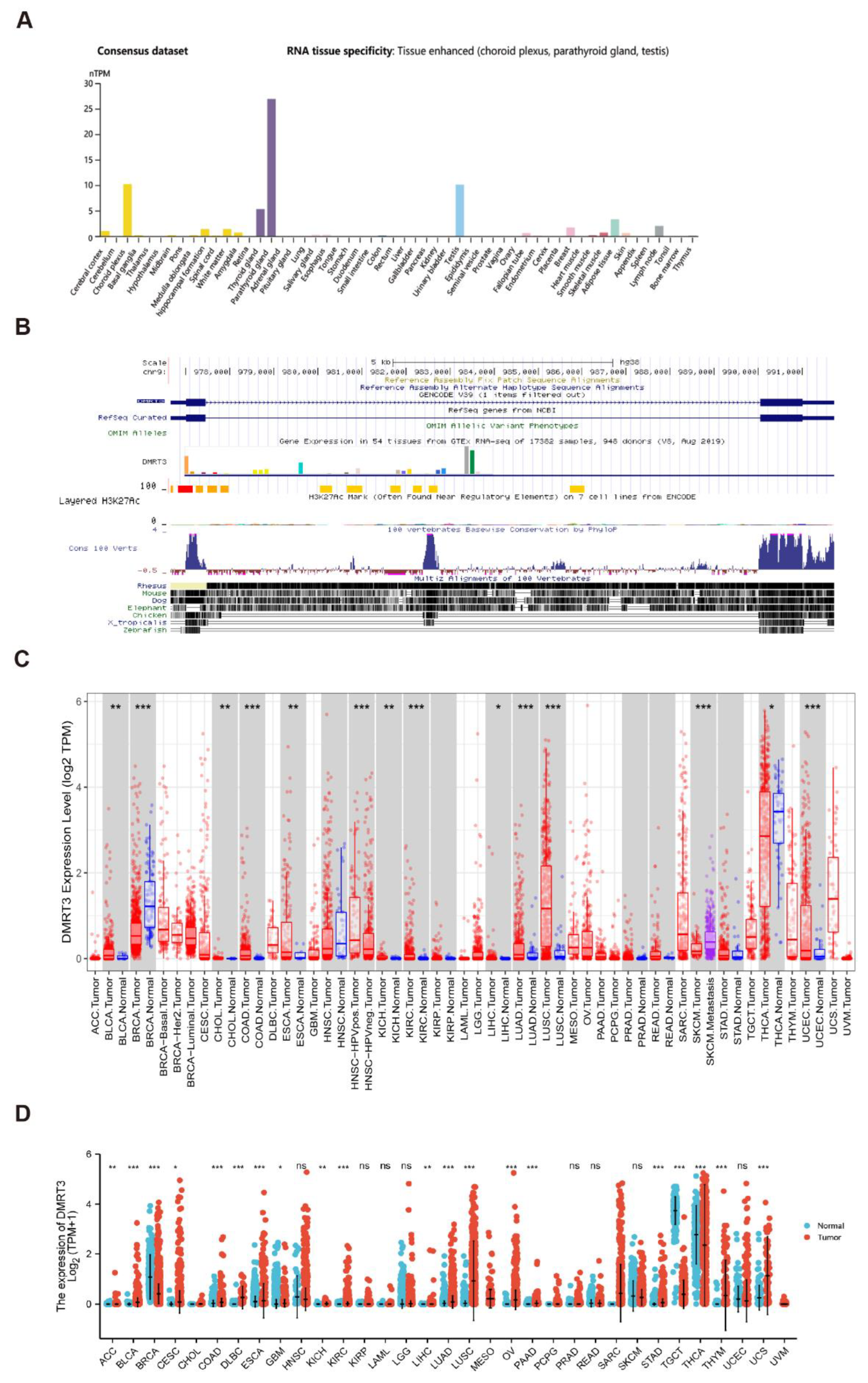
3.1. Gene Expression Analysis of DMRT3
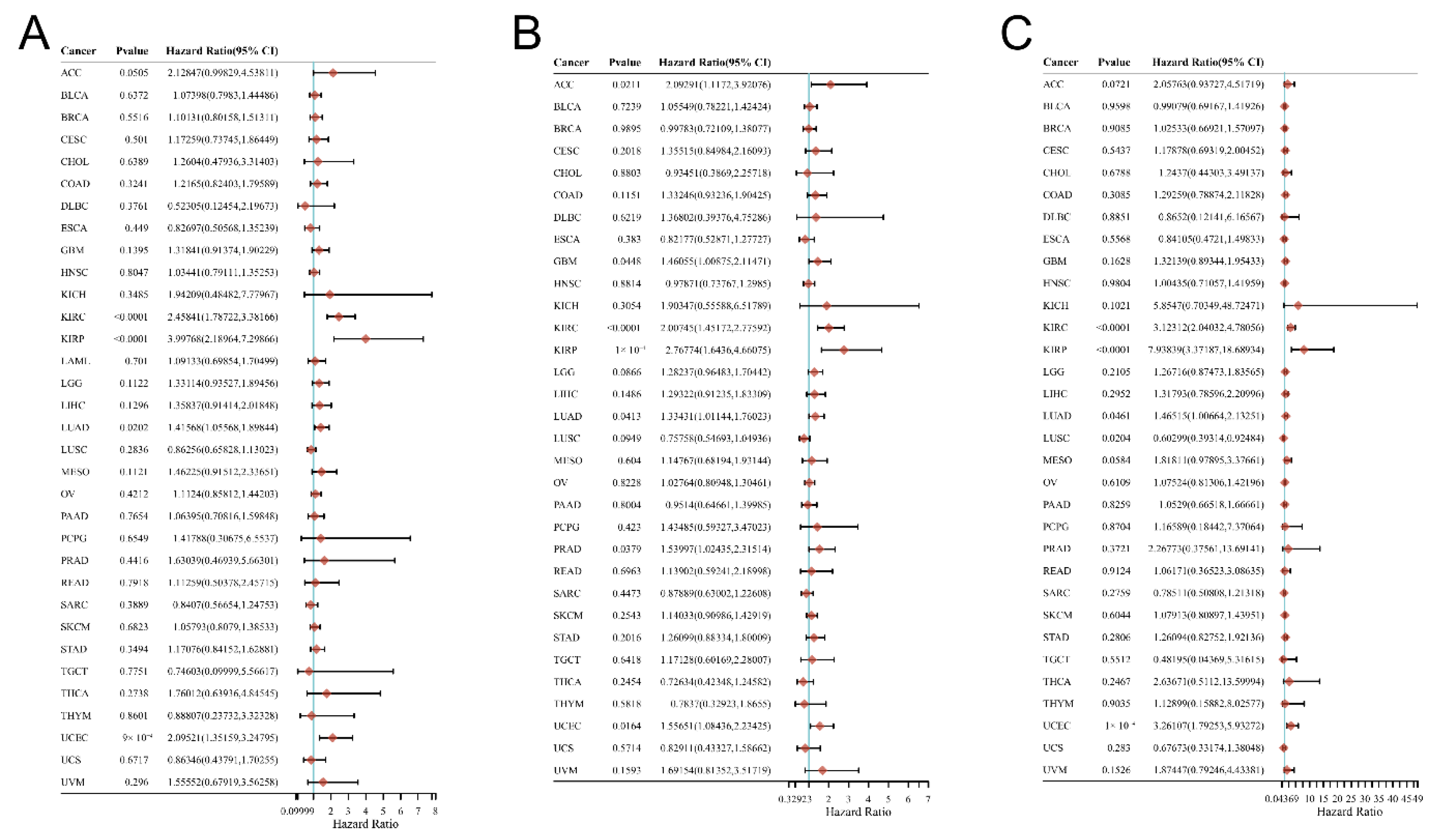
3.2. Relationship between the DMRT3 Expression and the Prognosis of Cancer Patients
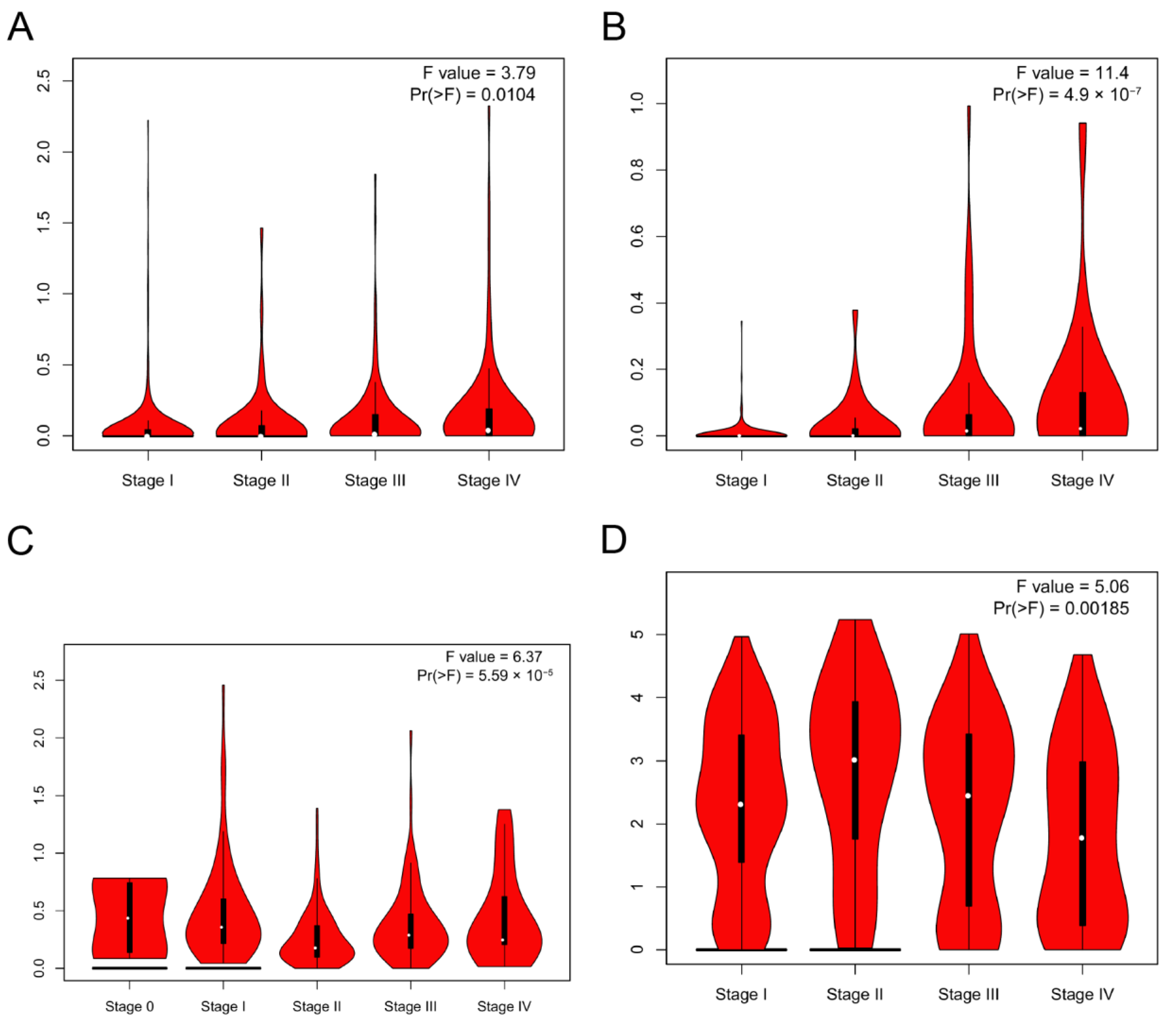
3.3. DMRT3 Expression between the Different Clinical Characteristics
3.4. The Landscape of the Genetic Alterations in DMRT3 in Different Tumors
3.5. DMRT3 Gene Expression in the Pan-Cancer Tumors Correlates with the Immune Infiltration
3.6. DMRT3 Gene Expression in Pan-Cancer Tumors Correlates with the Immune Checkpoint Genes
3.7. Biological Function of DMRT3 in Cancer
3.8. Drug Sensitivity Analysis of DMRT3
3.9. Cancer-Related Expression Pattern of DMRT3 in Single Cells
3.10. CeRNA Network of DMRT3
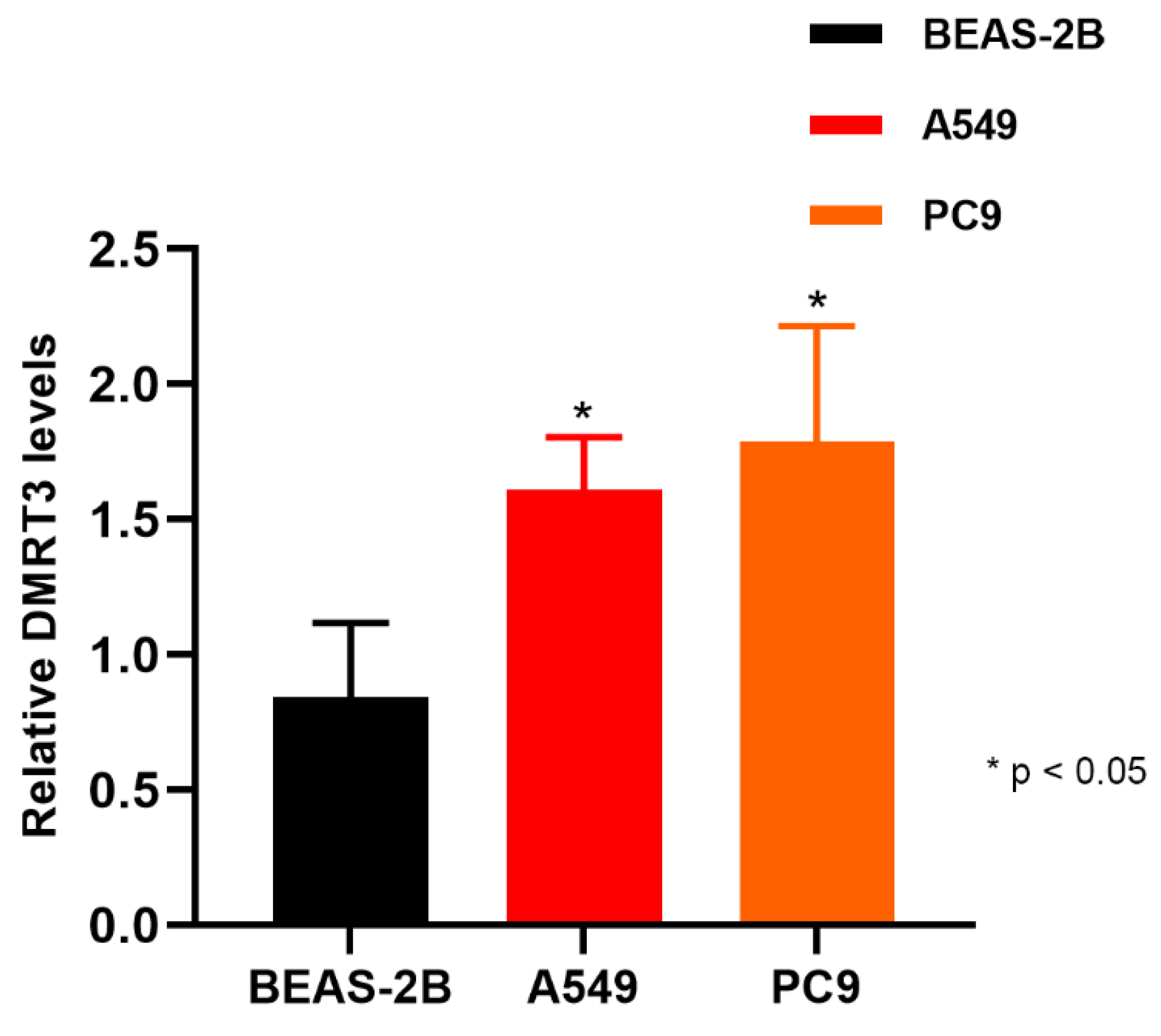
3.11. Validation of the DMRT3 Expression in the Cell Lines
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2020. CA Cancer J. Clin. 2020, 70, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer Statistics, 2021. CA Cancer J. Clin. 2021, 71, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Hua, X.; Zhao, W.; Pesatori, A.C.; Consonni, D.; Caporaso, N.E.; Zhang, T.; Zhu, B.; Wang, M.; Jones, K.; Hicks, B.; et al. Genetic and epigenetic intratumor heterogeneity impacts prognosis of lung adenocarcinoma. Nat. Commun. 2020, 11, 2459. [Google Scholar] [CrossRef] [PubMed]
- He, D.; Wang, D.; Lu, P.; Yang, N.; Xue, Z.; Zhu, X.; Zhang, P.; Fan, G. Single-cell RNA sequencing reveals heterogeneous tumor and immune cell populations in early-stage lung adenocarcinomas harboring EGFR mutations. Oncogene 2021, 40, 355–368. [Google Scholar] [CrossRef]
- Lei, X.; Lei, Y.; Li, J.K.; Du, W.X.; Li, R.G.; Yang, J.; Li, J.; Li, F.; Tan, H.B. Immune cells within the tumor microenvironment: Biological functions and roles in cancer immunotherapy. Cancer Lett. 2020, 470, 126–133. [Google Scholar] [CrossRef]
- Bellefroid, E.J.; Leclère, L.; Saulnier, A.; Keruzore, M.; Sirakov, M.; Vervoort, M.; De Clercq, S. Expanding roles for the evolutionarily conserved Dmrt sex transcriptional regulators during embryogenesis. Cell. Mol. Life Sci. 2013, 70, 3829–3845. [Google Scholar] [CrossRef]
- Kopp, A. Dmrt genes in the development and evolution of sexual dimorphism. Trends Genet. 2012, 28, 175–184. [Google Scholar] [CrossRef]
- Priyadharshini, V.S.; Jiménez-Chobillon, M.A.; de Graaf, J.; Porras Gutiérrez de Velasco, R.; Gratziou, C.; Ramírez-Jiménez, F.; Teran, L.M. Transcriptome Analysis Identifies Doublesex and Mab-3 Related Transcription Factor (DMRT3) in Nasal Polyp Epithelial Cells of Patients Suffering from Non-Steroidal Anti-Inflammatory Drug-Exacerbated Respiratory Disease (AERD). Biomolecules 2021, 11, 81092. [Google Scholar] [CrossRef]
- Lu, X.; Jing, L.; Liu, S.; Wang, H.; Chen, B. miR-149-3p Is a Potential Prognosis Biomarker and Correlated with Immune Infiltrates in Uterine Corpus Endometrial Carcinoma. Int. J. Endocrinol. 2022, 2022, 5006123. [Google Scholar] [CrossRef]
- Vivian, J.; Rao, A.A.; Nothaft, F.A.; Ketchum, C.; Armstrong, J.; Novak, A.; Pfeil, J.; Narkizian, J.; Deran, A.D.; Musselman-Brown, A.; et al. Toil enables reproducible, open source, big biomedical data analyses. Nat. Biotechnol. 2017, 35, 314–316. [Google Scholar] [CrossRef]
- Lin, Z.; Huang, W.; Yi, Y.; Li, D.; Xie, Z.; Li, Z.; Ye, M. LncRNA ADAMTS9-AS2 is a Prognostic Biomarker and Correlated with Immune Infiltrates in Lung Adenocarcinoma. Int. J. Gen. Med. 2021, 14, 8541–8555. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Sun, Y.; Ma, Y.; Zhao, X.; Sun, X.; Wang, Y.; Zhang, X. Comprehensive Pan-Cancer Analysis of IRAK Family Genes Identifies IRAK1 as a Novel Oncogene in Low-Grade Glioma. J. Oncol. 2022, 2022, 6497241. [Google Scholar] [CrossRef] [PubMed]
- Frost, F.G.; Cherukuri, P.F.; Milanovich, S.; Boerkoel, C.F. Pan-cancer RNA-seq data stratifies tumours by some hallmarks of cancer. J. Cell. Mol. Med. 2020, 24, 418–430. [Google Scholar] [CrossRef] [PubMed]
- Lao, Y.; Li, T.; Xie, X.; Chen, K.; Li, M.; Huang, L. MiR-195-3p is a Novel Prognostic Biomarker Associated with Immune Infiltrates of Lung Adenocarcinoma. Int. J. Gen. Med. 2022, 15, 191–203. [Google Scholar] [CrossRef]
- Tian, Y.; Bai, F.; Zhang, D. HIF1α: A Novel Biomarker with Potential Prognostic and Immunotherapy in Pan-cancer. Oxid. Med. Cell. Longev. 2022, 2022, 1246267. [Google Scholar] [CrossRef]
- Yuan, Z.; Wang, L.; Chen, C. Analysis of the prognostic, diagnostic and immunological role of HSP90α in malignant tumors. Front. Oncol. 2022, 12, 963719. [Google Scholar] [CrossRef]
- Gao, H.; Xu, C.; Liang, J.; Ge, S.; Zhang, F.; Tuo, Y.; Shi, H.; Han, A. Pan-cancer analysis of oncogenic role of Programmed Cell Death 2 Like (PDCD2L) and validation in colorectal cancer. Cancer Cell Int. 2022, 22, 100. [Google Scholar] [CrossRef]
- Thorsson, V.; Gibbs, D.L.; Brown, S.D.; Wolf, D.; Bortone, D.S.; Ou Yang, T.-H.; Porta-Pardo, E.; Gao, G.F.; Plaisier, C.L.; Eddy, J.A.; et al. The Immune Landscape of Cancer. Immunity 2018, 48, 812–830.e14. [Google Scholar] [CrossRef]
- Bonneville, R.; Krook, M.A.; Kautto, E.A.; Miya, J.; Wing, M.R.; Chen, H.Z.; Reeser, J.W.; Yu, L.; Roychowdhury, S. Landscape of Microsatellite Instability Across 39 Cancer Types. JCO Precis. Oncol. 2017, 2017. [Google Scholar] [CrossRef]
- Zhao, W.J.; Ou, G.Y.; Lin, W.W. Integrative Analysis of Neuregulin Family Members-Related Tumor Microenvironment for Predicting the Prognosis in Gliomas. Front. Immunol. 2021, 12, 682415. [Google Scholar] [CrossRef]
- Liu, Y.; Xue, J.; Zhong, M.; Wang, Z.; Li, J.; Zhu, Y. Prognostic Prediction, Immune Microenvironment, and Drug Resistance Value of Collagen Type I Alpha 1 Chain: From Gastrointestinal Cancers to Pan-Cancer Analysis. Front. Mol. Biosci. 2021, 8, 692120. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Tang, H.; Li, T.; Jiang, K.; Zhong, H.; Wu, Y.; He, J.; Li, D.; Li, M.; Cai, X. Comprehensive Analysis of the Expression, Prognosis, and Biological Significance of OVOLs in Breast Cancer. Int. J. Gen. Med. 2021, 14, 3951–3960. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Mai, H.; Chen, H.; Zhou, B.; Hou, J.; Jiang, D.-K. Pan-Cancer Analysis Identified C1ORF112 as a Potential Biomarker for Multiple Tumor Types. Front. Mol. Biosci. 2021, 8, 693651. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Shi, W.; Miao, Y.; Gan, J.; Guan, Q.; Ran, J. Evaluation of tumor microenvironmental immune regulation and prognostic in lung adenocarcinoma from the perspective of purinergic receptor P2Y13. Bioengineered 2021, 12, 6286–6304. [Google Scholar] [CrossRef]
- Li, M.; Wang, X.; Liu, J.; Mao, X.; Li, D.; Wang, Z.; Tang, Y.; Wu, S. Identification of Core Prognosis-Related Candidate Genes in Chinese Gastric Cancer Population Based on Integrated Bioinformatics. Biomed. Res. Int. 2020, 2020, 8859826. [Google Scholar] [CrossRef]
- Yuan, H.; Yan, M.; Zhang, G.; Liu, W.; Deng, C.; Liao, G.; Xu, L.; Luo, T.; Yan, H.; Long, Z.; et al. CancerSEA: A cancer single-cell state atlas. Nucleic Acids Res. 2019, 47, D900–D908. [Google Scholar] [CrossRef]
- Lu, X.; Li, G.; Liu, S.; Wang, H.; Chen, B. MiR-585-3p suppresses tumor proliferation and migration by directly targeting CAPN9 in high grade serous ovarian cancer. J. Ovarian Res. 2021, 14, 90. [Google Scholar] [CrossRef]
- Chen, B.; Gao, C.; Wang, H.; Sun, J.; Han, Z. Molecular Analysis of Prognosis and Immune Infiltration of Ovarian Cancer Based on Homeobox D Genes. Comput. Math. Methods Med. 2022, 2022, 3268386. [Google Scholar] [CrossRef]
- Liu, J.; Lichtenberg, T.; Hoadley, K.A.; Poisson, L.M.; Lazar, A.J.; Cherniack, A.D.; Kovatich, A.J.; Benz, C.C.; Levine, D.A.; Lee, A.V.; et al. An Integrated TCGA Pan-Cancer Clinical Data Resource to Drive High-Quality Survival Outcome Analytics. Cell 2018, 173, 400–416.e11. [Google Scholar] [CrossRef]
- Lánczky, A.; Győrffy, B. Web-Based Survival Analysis Tool Tailored for Medical Research (KMplot): Development and Implementation. J. Med. Internet Res. 2021, 23, e27633. [Google Scholar] [CrossRef]
- De la Chapelle, A.; Hampel, H. Clinical relevance of microsatellite instability in colorectal cancer. J. Clin. Oncol. 2010, 28, 3380–3387. [Google Scholar] [CrossRef] [PubMed]
- Musha, K.; Ge, X.; Ablikim, N.; Lu, B.; Chen, C.; Huang, J. Comprehensive Analysis of RELL2 as a Potential Biomarker Associated with Tumor Immune Infiltrating Cells in a Pan-Cancer Analysis. Dis. Markers 2022, 2022, 5009512. [Google Scholar] [CrossRef] [PubMed]
- Mhaidly, R.; Mechta-Grigoriou, F. Role of cancer-associated fibroblast subpopulations in immune infiltration, as a new means of treatment in cancer. Immunol. Rev. 2021, 302, 259–272. [Google Scholar] [CrossRef] [PubMed]
- Weinstein, J.N.; Collisson, E.A.; Mills, G.B.; Shaw, K.R.; Ozenberger, B.A.; Ellrott, K.; Shmulevich, I.; Sander, C.; Stuart, J.M. The Cancer Genome Atlas Pan-Cancer analysis project. Nat. Genet. 2013, 45, 1113–1120. [Google Scholar] [CrossRef]
- Schaub, F.X.; Dhankani, V.; Berger, A.C.; Trivedi, M.; Richardson, A.B.; Shaw, R.; Zhao, W.; Zhang, X.; Ventura, A.; Liu, Y.; et al. Pan-cancer Alterations of the MYC Oncogene and Its Proximal Network across the Cancer Genome Atlas. Cell Syst. 2018, 6, 282–300.e2. [Google Scholar] [CrossRef]
- Katsuta, E.; Huyser, M.; Yan, L.; Takabe, K. A prognostic score based on long-term survivor unique transcriptomic signatures predicts patient survival in pancreatic ductal adenocarcinoma. Am. J. Cancer Res. 2021, 11, 4294–4307. [Google Scholar]
- Meng, Y.; Li, S.; Gu, D.; Xu, K.; Du, M.; Zhu, L.; Chu, H.; Zhang, Z.; Wu, Y.; Fu, Z.; et al. Genetic variants in m6A modification genes are associated with colorectal cancer risk. Carcinogenesis 2020, 41, 8–17. [Google Scholar] [CrossRef]
- Zhang, S.; Li, M.; Ji, H.; Fang, Z. Landscape of transcriptional deregulation in lung cancer. BMC Genom. 2018, 19, 435. [Google Scholar] [CrossRef]
- Tsai, C.L.; Tsai, C.N.; Lee, Y.S.; Wang, H.S.; Lee, L.Y.; Lin, C.Y.; Yang, S.Y.; Chao, A. Genetic analysis of a Taiwanese family identifies a DMRT3-OAS3 interaction that is involved in human sexual differentiation through the regulation of ESR1 expression. Fertil. Steril. 2020, 114, 133–143. [Google Scholar] [CrossRef]
- Desmaris, E.; Keruzore, M.; Saulnier, A.; Ratié, L.; Assimacopoulos, S.; De Clercq, S.; Nan, X.; Roychoudhury, K.; Qin, S.; Kricha, S.; et al. DMRT5, DMRT3, and EMX2 Cooperatively Repress Gsx2 at the Pallium-Subpallium Boundary to Maintain Cortical Identity in Dorsal Telencephalic Progenitors. J. Neurosci. 2018, 38, 9105–9121. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | RNA Molecule | Source | Spearman.Stat | Spearman.Fdr | p Value |
|---|---|---|---|---|---|
| Tanespimycin | DMRT3 | GDSC | 0.194987316 | 8.98 × 10−9 | 1.33 × 10−9 |
| PD-0325901 | DMRT3 | CCLE | 0.164608073 | 0.044844396 | 0.00526032 |
| PLX4720 | DMRT3 | CCLE | 0.16154357 | 0.043031472 | 0.006181571 |
| Trametinib | DMRT3 | GDSC | 0.160737175 | 2.58 × 10−6 | 6.26794 × 10−7 |
| Refametinib | DMRT3 | GDSC | 0.154929636 | 6.85 × 10−6 | 1.58274 × 10−6 |
| GSK690693 | DMRT3 | GDSC | −0.15192674 | 1.09 × 10−5 | 2.52199 × 10−6 |
| PHA-793887 | DMRT3 | GDSC | −0.158247668 | 3.48 × 10−6 | 9.36157 × 10−7 |
| NPK76-II-72-1 | DMRT3 | GDSC | −0.159673169 | 2.33 × 10−6 | 7.44586 × 10−7 |
| TAK-715 | DMRT3 | GDSC | −0.159971009 | 2.25 × 10−6 | 7.09624 × 10−7 |
| PI-103 | DMRT3 | GDSC | −0.174534416 | 4.26 × 10−7 | 6.06437 × 10−8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, D.; Liu, M.; Jiang, J.; Luo, Y.; Wang, Y.; Chen, H.; Li, D.; Wang, D.; Yang, Z.; Chen, H. Comprehensive Analysis of DMRT3 as a Potential Biomarker Associated with the Immune Infiltration in a Pan-Cancer Analysis and Validation in Lung Adenocarcinoma. Cancers 2022, 14, 6220. https://doi.org/10.3390/cancers14246220
Yang D, Liu M, Jiang J, Luo Y, Wang Y, Chen H, Li D, Wang D, Yang Z, Chen H. Comprehensive Analysis of DMRT3 as a Potential Biomarker Associated with the Immune Infiltration in a Pan-Cancer Analysis and Validation in Lung Adenocarcinoma. Cancers. 2022; 14(24):6220. https://doi.org/10.3390/cancers14246220
Chicago/Turabian StyleYang, Donghong, Meilian Liu, Junhong Jiang, Yiping Luo, Yongcun Wang, Huoguang Chen, Dongbing Li, Dongliang Wang, Zhixiong Yang, and Hualin Chen. 2022. "Comprehensive Analysis of DMRT3 as a Potential Biomarker Associated with the Immune Infiltration in a Pan-Cancer Analysis and Validation in Lung Adenocarcinoma" Cancers 14, no. 24: 6220. https://doi.org/10.3390/cancers14246220
APA StyleYang, D., Liu, M., Jiang, J., Luo, Y., Wang, Y., Chen, H., Li, D., Wang, D., Yang, Z., & Chen, H. (2022). Comprehensive Analysis of DMRT3 as a Potential Biomarker Associated with the Immune Infiltration in a Pan-Cancer Analysis and Validation in Lung Adenocarcinoma. Cancers, 14(24), 6220. https://doi.org/10.3390/cancers14246220