
Sialyltransferase Inhibitor Ac53FaxNeu5Ac Reverts the Malignant Phenotype of Pancreatic Cancer Cells, and Reduces Tumor Volume and Favors T-Cell Infiltrates in Mice

 , , , , , and
, , , , , and 
Abstract
Simple Summary
Abstract
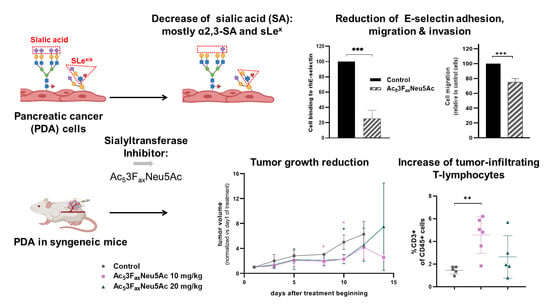
1. Introduction
2. Materials and Methods
2.1. Reagents and Cell Lines
2.2. Flow Cytometry
2.3. Protein Lysates and Western Blot (WB)
2.4. E-Selectin Binding Assay
2.5. Transwell Migration and Invasion Assays
2.6. Syngeneic Mice Tumor Generation
2.7. Immunohistochemistry
2.8. Statistical Analyses
3. Results
3.1. Ac53FaxNeu5Ac Reduced Cell Surface Sialylation in BxPC-3, Capan-1 and Panc-1 Cells
3.2. Ac53FaxNeu5Ac Impaired E-Selectin Binding in BxPC-3 and Capan-1 Cells
3.3. Ac53FaxNeu5Ac Reduced PDA Cells Migration and Invasion
3.4. Ac53FaxNeu5Ac Decreased SA Expression in KPC and KC Murine PDA Cell Lines, and Impaired KC Cells Migration and Invasion
3.5. Ac53FaxNeu5Ac Treatment Reduced the Growth of the Tumors Generated by Subcutaneous Injection of KC Cells in Syngeneic Mice
3.6. Ac53FaxNeu5Ac Treatment Reduced SA Expression on Tumor Cells and Altered the Tumor Immune Component
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer Statistics, 2022. CA Cancer J. Clin. 2022, 72, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Orth, M.; Metzger, P.; Gerum, S.; Mayerle, J.; Schneider, G.; Belka, C.; Schnurr, M.; Lauber, K. Pancreatic Ductal Adenocarcinoma: Biological Hallmarks, Current Status, and Future Perspectives of Combined Modality Treatment Approaches. Radiat. Oncol. 2019, 14, 141. [Google Scholar] [CrossRef] [PubMed]
- Pereira, S.P.; Oldfield, L.; Ney, A.; Hart, P.A.; Keane, M.G.; Pandol, S.J.; Li, D.; Greenhalf, W.; Jeon, C.Y.; Koay, E.J.; et al. Early Detection of Pancreatic Cancer. Lancet Gastroenterol. Hepatol. 2020, 5, 698–710. [Google Scholar] [CrossRef] [PubMed]
- Sahin, I.H.; Askan, G.; Hu, Z.I.; O’Reilly, E.M. Immunotherapy in Pancreatic Ductal Adenocarcinoma: An Emerging Entity? Ann. Oncol. 2017, 28, 2950–2961. [Google Scholar] [CrossRef] [PubMed]
- Huber, M.; Brehm, C.U.; Gress, T.M.; Buchholz, M.; Alhamwe, B.A.; von Strandmann, E.P.; Slater, E.P.; Bartsch, J.W.; Bauer, C.; Lauth, M. The Immune Microenvironment in Pancreatic Cancer. Int. J. Mol. Sci. 2020, 21, 7307. [Google Scholar] [CrossRef]
- Munkley, J. The Glycosylation Landscape of Pancreatic Cancer (Review). Oncol. Lett. 2019, 17, 2569–2575. [Google Scholar] [CrossRef]
- Lumibao, J.C.; Tremblay, J.R.; Hsu, J.; Engle, D.D. Altered Glycosylation in Pancreatic Cancer and Beyond. J. Exp. Med. 2022, 219, 1–13. [Google Scholar] [CrossRef]
- Rodrigues, J.G.; Balmaña, M.; Macedo, J.A.; Poças, J.; Fernandes, Â.; de-Freitas-Junior, J.C.M.; Pinho, S.S.; Gomes, J.; Magalhães, A.; Gomes, C.; et al. Glycosylation in Cancer: Selected Roles in Tumour Progression, Immune Modulation and Metastasis. Cell Immunol. 2018, 333, 46–57. [Google Scholar] [CrossRef]
- Heise, T.; Pijnenborg, J.F.A.; Büll, C.; van Hilten, N.; Kers-Rebel, E.D.; Balneger, N.; Elferink, H.; Adema, G.J.; Boltje, T.J. Potent Metabolic Sialylation Inhibitors Based on C-5-Modified Fluorinated Sialic Acids. J. Med. Chem. 2019, 62, 1014–1021. [Google Scholar] [CrossRef]
- Büll, C.; Boltje, T.J.; Wassink, M.; de Graaf, A.M.A.; van Delft, F.L.; den Brok, M.H.; Adema, G.J.; Bull, C.; Boltje, T.J.; Adema, G.J.; et al. Targeting Aberrant Sialylation in Cancer Cells Using a Fluorinated Sialic Acid Analog Impairs Adhesion, Migration, and In Vivo Tumor Growth. Mol. Cancer Ther. 2013, 12, 1935–1946. [Google Scholar] [CrossRef] [PubMed]
- Büll, C.; Boltje, T.J.; Balneger, N.; Weischer, S.M.; Wassink, M.; van Gemst, J.J.; Bloemendal, V.R.; Boon, L.; van der Vlag, J.; Heise, T.; et al. Sialic Acid Blockade Suppresses Tumor Growth by Enhancing T-Cell-Mediated Tumor Immunity. Cancer Res. 2018, 78, 3574–3588. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Garay, M.; Arteta, B.; Pagés, L.; de Llorens, R.; de Bolós, C.; Vidal-Vanaclocha, F.; Peracaula, R. A2,3-Sialyltransferase ST3Gal III Modulates Pancreatic Cancer Cell Motility and Adhesion in Vitro and Enhances Its Metastatic Potential in Vivo. PLoS ONE 2010, 5, e12524. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Garay, M.; Arteta, B.; Llop, E.; Cobler, L.; Pagès, L.; Ortiz, R.; Ferri, M.J.; de Bolós, C.; Figueras, J.; de Llorens, R.; et al. A2,3-Sialyltransferase ST3Gal IV Promotes Migration and Metastasis in Pancreatic Adenocarcinoma Cells and Tends to Be Highly Expressed in Pancreatic Adenocarcinoma Tissues. Int. J. Biochem. Cell Biol. 2013, 45, 1748–1757. [Google Scholar] [CrossRef] [PubMed]
- Guerrero, P.E.; Miró, L.; Wong, B.S.; Massaguer, A.; Martínez-Bosch, N.; de Llorens, R.; Navarro, P.; Konstantopoulos, K.; Llop, E.; Peracaula, R. Knockdown of A2,3-Sialyltransferases Impairs Pancreatic Cancer Cell Migration, Invasion and E-Selectin-Dependent Adhesion. Int. J. Mol. Sci. 2020, 21, 6239. [Google Scholar] [CrossRef] [PubMed]
- Clark, C.E.; Hingorani, S.R.; Mick, R.; Combs, C.; Tuveson, D.A.; Vonderheide, R.H. Dynamics of the Immune Reaction to Pancreatic Cancer from Inception to Invasion. Cancer Res. 2007, 67, 9518–9527. [Google Scholar] [CrossRef]
- Guerrero, P.E.; Duran, A.; Ortiz, M.R.; Castro, E.; Garcia-Velasco, A.; Llop, E.; Peracaula, R. Microfibril Associated Protein 4 (MFAP4) Is a Carrier of the Tumor Associated Carbohydrate Sialyl-Lewis x (SLex) in Pancreatic Adenocarcinoma. J. Proteom. 2021, 231, 104004. [Google Scholar] [CrossRef]
- Rodriguez, E.; Boelaars, K.; Brown, K.; Eveline Li, R.J.; Kruijssen, L.; Bruijns, S.C.M.; van Ee, T.; Schetters, S.T.T.; Crommentuijn, M.H.W.; van der Horst, J.C.; et al. Sialic Acids in Pancreatic Cancer Cells Drive Tumour-Associated Macrophage Differentiation via the Siglec Receptors Siglec-7 and Siglec-9. Nat. Commun. 2021, 12, 1270. [Google Scholar] [CrossRef]
- Rillahan, C.D.; Antonopoulos, A.; Lefort, C.T.; Sonon, R.; Ley, K.; Dell, A.; Haslam, S.M.; Paulson, J.C. Global Metabolic Inhibitors of Sialyl- and Fucosyltransferases. Nat. Chem. Biol. 2012, 8, 661–668. [Google Scholar] [CrossRef]
- Natoni, A.; Farrell, M.L.; Harris, S.; Falank, C.; Kirkham-McCarthy, L.; Macauley, M.S.; Reagan, M.R.; O’Dwyer, M. Sialyltransferase Inhibition Leads to Inhibition of Tumor Cell Interactions with E-Selectin, VCAM1, and MADCAM1, and Improves Sur Vival in a Human Multiple Myeloma Mouse Model. Haematologica 2020, 105, 457–467. [Google Scholar] [CrossRef]
- Hsu, C.C.; Lin, T.W.; Chang, W.W.; Wu, C.Y.; Lo, W.H.; Wang, P.H.; Tsai, Y.C. Soyasaponin-I-Modified Invasive Behavior of Cancer by Changing Cell Surface Sialic Acids. Gynecol. Oncol. 2005, 96, 415–422. [Google Scholar] [CrossRef] [PubMed]
- Zhan, L.; Chen, L.; Chen, Z. Knockdown of FUT3 Disrupts the Proliferation, Migration, Tumorigenesis and TGF-β Induced EMT in Pancreatic Cancer Cells. Oncol. Lett. 2018, 16, 924–930. [Google Scholar] [CrossRef] [PubMed]
- Pinho, S.; Marcos, N.T.; Ferreira, B.; Carvalho, A.S.; Oliveira, M.J.; Santos-Silva, F.; Harduin-Lepers, A.; Reis, C.A. Biological Significance of Cancer-Associated Sialyl-Tn Antigen: Modulation of Malignant Phenotype in Gastric Carcinoma Cells. Cancer Lett. 2007, 249, 157–170. [Google Scholar] [CrossRef] [PubMed]
- Gomes, C.; Osório, H.; Pinto, M.T.; Campos, D.; Oliveira, M.J.; Reis, C.A. Expression of ST3GAL4 Leads to SLex Expression and Induces C-Met Activation and an Invasive Phenotype in Gastric Carcinoma Cells. PLoS ONE 2013, 8, e66737. [Google Scholar] [CrossRef]
- Cui, H.X.; Wang, H.; Wang, Y.; Song, J.; Tian, H.U.A.; Xia, C.; Shen, Y. ST3Gal III Modulates Breast Cancer Cell Adhesion and Invasion by Altering the Expression of Invasion-Related Molecules. Oncol. Rep. 2016, 36, 3317–3324. [Google Scholar] [CrossRef]
- Shen, L.; Luo, Z.; Wu, J.; Qiu, L.; Luo, M.; Ke, Q.; Dong, X. Enhanced Expression of A2,3-Linked Sialic Acids Promotes Gastric Cancer Cell Metastasis and Correlates with Poor Prognosis. Int. J. Oncol. 2017, 50, 1201–1210. [Google Scholar] [CrossRef]
- Bassagañas, S.; Carvalho, S.; Dias, A.M.; Pérez-Garay, M.; Ortiz, M.R.; Figueras, J.; Reis, C.A.; Pinho, S.S.; Peracaula, R. Pancreatic Cancer Cell Glycosylation Regulates Cell Adhesion and Invasion through the Modulation of A2β1 Integrin and E-Cadherin Function. PLoS ONE 2014, 9, e98595. [Google Scholar] [CrossRef]
- Chiang, C.H.; Wang, C.H.; Chang, H.C.; More, S.V.; Li, W.S.; Hung, W.C. A Novel Sialyltransferase Inhibitor AL10 Suppresses Invasion and Metastasis of Lung Cancer Cells by Inhibiting Integrin-Mediated Signaling. J. Cell Physiol. 2010, 223, 492–499. [Google Scholar] [CrossRef]
- Sung, P.L.; Wen, K.C.; Horng, H.C.; Chang, C.M.; Chen, Y.J.; Lee, W.L.; Wang, P.H. The Role of A2,3-Linked Sialylation on Clear Cell Type Epithelial Ovarian Cancer. Taiwan J. Obstet. Gynecol. 2018, 57, 255–263. [Google Scholar] [CrossRef]
- Bassagañas, S.; Pérez-Garay, M.; Peracaula, R. Cell Surface Sialic Acid Modulates Extracellular Matrix Adhesion and Migration in Pancreatic Adenocarcinoma Cells. Pancreas 2014, 43, 109–117. [Google Scholar] [CrossRef]
- Macauley, M.S.; Arlian, B.M.; Rillahan, C.D.; Pang, P.-C.C.; Bortell, N.; Marcondes, M.C.G.; Haslam, S.M.; Dell, A.; Paulson, J.C.; Macauley, M.S.; et al. Systemic Blockade of Sialylation in Mice with a Global Inhibitor of Sialyltransferases. J. Biol. Chem. 2014, 289, 35149–35158. [Google Scholar] [CrossRef] [PubMed]
- Goulart, M.R.; Stasinos, K.; Fincham, R.E.A.; Delvecchio, F.R.; Kocher, H.M. T Cells in Pancreatic Cancer Stroma. World J. Gastroenterol. 2021, 27, 7956–7968. [Google Scholar] [CrossRef] [PubMed]
- Ino, Y.; Yamazaki-Itoh, R.; Shimada, K.; Iwasaki, M.; Kosuge, T.; Kanai, Y.; Hiraoka, N. Immune Cell Infiltration as an Indicator of the Immune Microenvironment of Pancreatic Cancer. Br. J. Cancer 2013, 108, 914–923. [Google Scholar] [CrossRef] [PubMed]
- Cortesi, M.; Zanoni, M.; Pirini, F.; Tumedei, M.M.; Ravaioli, S.; Rapposelli, I.G.; Frassineti, G.L.; Bravaccini, S. Pancreatic Cancer and Cellular Senescence: Tumor Microenvironment under the Spotlight. Int. J. Mol. Sci. 2022, 23, 254. [Google Scholar] [CrossRef] [PubMed]
- De Palma, M.; Lewis, C.E. Macrophage Regulation of Tumor Responses to Anticancer Therapies. Cancer Cell 2013, 23, 277–286. [Google Scholar] [CrossRef]
- Khaled, Y.S.; Ammori, B.J.; Elkord, E. Increased Levels of Granulocytic Myeloid-Derived Suppressor Cells in Peripheral Blood and Tumour Tissue of Pancreatic Cancer Patients. J. Immunol. Res. 2014, 2014, 879897. [Google Scholar] [CrossRef]
- Büll, C.; den Brok, M.H.; Adema, G.J. Sweet Escape: Sialic Acids in Tumor Immune Evasion. Biochim. Biophys. Acta Rev. Cancer 2014, 1846, 238–246. [Google Scholar] [CrossRef] [PubMed]
- Torphy, R.J.; Schulick, R.D.; Zhu, Y. Understanding the Immune Landscape and Tumor Microenvironment of Pancreatic Cancer to Improve Immunotherapy. Mol. Carcinog. 2020, 59, 775–782. [Google Scholar] [CrossRef] [PubMed]
- Van de Wall, S.; Santegoets, K.C.M.; van Houtum, E.J.H.; Büll, C.; Adema, G.J. Sialoglycans and Siglecs Can Shape the Tumor Immune Microenvironment. Trends Immunol. 2020, 41, 274–285. [Google Scholar] [CrossRef]
- Adams, O.J.; Stanczak, M.A.; von Gunten, S.; Läubli, H. Targeting Sialic Acid-Siglec Interactions to Reverse Immune Suppression in Cancer. Glycobiology 2018, 28, 640–647. [Google Scholar] [CrossRef]
- Meril, S.; Harush, O.; Reboh, Y.; Matikhina, T.; Barliya, T.; Cohen, C.J. Targeting Glycosylated Antigens on Cancer Cells Using Siglec-7/9-Based CAR T-Cells. Mol. Carcinog. 2020, 59, 713–723. [Google Scholar] [CrossRef] [PubMed]
- Perdicchio, M.; Cornelissen, L.A.M.; Streng-Ouwehand, I.; Engels, S.; Verstege, M.I.; Boon, L.; Geerts, D.; van Kooyk, Y.; Unger, W.W.J. Tumor Sialylation Impedes T Cell Mediated Anti-Tumor Responses While Promoting Tumor Associated-Regulatory T Cells. Oncotarget 2016, 7, 8771–8782. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody/Biotinylated Lectin /Conjugated Reagent | Clone or Host/Isotype | Dilution | Use | Supplier |
|---|---|---|---|---|
| anti-sLex | CSLEX | 1/10 (FC) | FC/WB | Cat# 563529, BD Biosciences (San Jose, CA, USA) |
| 1/70 (WB) | ||||
| anti-sLea | 121SLE | 1/500 (FC) | FC/WB | Cat# ab3982, Abcam (Cambridge, UK) |
| 1/1000 (WB) | ||||
| anti-Tubulin | B-7 | 1/500 | WB | Cat# sc-5286, Santa Cruz Biotechnology (Dallas, TX, USA) |
| Biotinylated SNA | 1/100 (FC) | FC/WB | Cat# B-1305, Vector Laboratories (Burlingame, CA, USA) | |
| 1/1000 (WB) | ||||
| Biotinylated MAA-II | 1/50 (FC) | FC/WB | Cat# B-1265, Vector Laboratories | |
| 1/500 (WB) | ||||
| Biotinylated PNA | 1/100 | FC | Cat# B-1075-5, Vector Laboratories | |
| Peroxidase-Conjugated goat anti-Mouse IgG + IgM | 1/4000 | WB | Cat#115-035-06, Jackson immune Research (West Grove, PA, USA) | |
| Peroxidase-Conjugate goat anti-Mouse IgG | 1/1000 | WB | Cat# 401215, Millipore (Darmstadt, Germany) | |
| Streptavidin-HRP Conjugate | 1/100,000 | WB | Cat# GERPN1231, GE Healthcare (Little Chalfont, UK) | |
| Anti-mouse IgG conjugated to Alexa Fluor 488 | 1/400 | FC | Cat# A-11029, Thermo Fisher Scientific (Waltham, MA, USA) | |
| Streptavidin conjugated to Alexa Fluor 488 | 1/1000 | FC | Cat# S32354, Invitrogen (Carlsbad, CA, USA) | |
| anti-phosphoHistone H3 | MC463 | 1/300 | IHC | Cat# 04-817, Millipore |
| anti-CD3 | 1/100 | IHC | Cat# ab5690, Abcam | |
| horse biotinylated antibody anti-IgGs | 1/100 | IHC | Cat# BA-1400, Vector Laboratories | |
| avidin-peroxidase complex | IHC | Cat# BA-1400, Vector Laboratories | ||
| Anti-mouse CD16 antibody | Rat IgG2a, λ | 1/100 | Spectral FC | Cat# 101302, Biolegend (San Diego, CA, USA) |
| Brilliant Violet 510™ anti-mouse CD8a Antibody | Rat IgG2a, κ | 1/1000 | Spectral FC | Cat# 100751, BioLegend |
| APC anti-mouse CD3 Antibody) | Rat IgG2b, κ | 1/500 | Spectral FC | Cat# 100235, BioLegend |
| PerCP-Cy™5.5 Rat Anti-Mouse CD45 | Rat LOU | 1/200 | Spectral FC | Cat# 567310, BD Biosciences |
| APC-Cyanine7 anti-mouse CD4 | Rat DA, also known as DA/HA IgG2a, κ | 1/400 | Spectral FC | Cat# 552051, BD Biosciences |
| PE/Cyanine7 anti-mouse Ly-6G Antibody | Rat IgG2a, κ | 1/1000 | Spectral FC | Cat# 127617, BioLegend |
| FITC anti-mouse/human CD11b Antibody | Rat IgG2b, κ | 1/4000 | Spectral FC | Cat# 101205, BioLegend |
| PE-Cy™5 Anti-Mouse CD45R/B220 | Rat IgG2b, κ | 1/200 | Spectral FC | Cat# 561879, BD Biosciences |
| Brilliant Violet 785™ anti-mouse F4/80 Antibody | Rat IgG2a, κ | 1/100 | Spectral FC | Cat# 123141, BioLegend |
| Alexa Fluor® 700 anti-mouse Ly-6C Antibody | Rat IgG2c, κ | 1/200 | Spectral FC | Cat# 128023, BioLegend |
| PE/Dazzle™ 594 anti-mouse CD49b (pan-NK1.1 cells) Antibody | Rat IgM, κ | 1/1000 | Spectral FC | Cat# 108923, BioLegend |
| Brilliant Violet 650™ anti-mouse/human CD45R/B220 Antibody | Rat IgG2a, κ | 1/100 | Spectral FC | Cat# 103241, BioLegend |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miró, L.; López, J.; Guerrero, P.E.; Martínez-Bosch, N.; Manero-Rupérez, N.; Moreno, M.; Ortiz, M.R.; Llop, E.; Navarro, P.; Peracaula, R. Sialyltransferase Inhibitor Ac53FaxNeu5Ac Reverts the Malignant Phenotype of Pancreatic Cancer Cells, and Reduces Tumor Volume and Favors T-Cell Infiltrates in Mice. Cancers 2022, 14, 6133. https://doi.org/10.3390/cancers14246133
Miró L, López J, Guerrero PE, Martínez-Bosch N, Manero-Rupérez N, Moreno M, Ortiz MR, Llop E, Navarro P, Peracaula R. Sialyltransferase Inhibitor Ac53FaxNeu5Ac Reverts the Malignant Phenotype of Pancreatic Cancer Cells, and Reduces Tumor Volume and Favors T-Cell Infiltrates in Mice. Cancers. 2022; 14(24):6133. https://doi.org/10.3390/cancers14246133
Chicago/Turabian StyleMiró, Laura, Júlia López, Pedro E. Guerrero, Neus Martínez-Bosch, Noemí Manero-Rupérez, Mireia Moreno, M. Rosa Ortiz, Esther Llop, Pilar Navarro, and Rosa Peracaula. 2022. "Sialyltransferase Inhibitor Ac53FaxNeu5Ac Reverts the Malignant Phenotype of Pancreatic Cancer Cells, and Reduces Tumor Volume and Favors T-Cell Infiltrates in Mice" Cancers 14, no. 24: 6133. https://doi.org/10.3390/cancers14246133
APA StyleMiró, L., López, J., Guerrero, P. E., Martínez-Bosch, N., Manero-Rupérez, N., Moreno, M., Ortiz, M. R., Llop, E., Navarro, P., & Peracaula, R. (2022). Sialyltransferase Inhibitor Ac53FaxNeu5Ac Reverts the Malignant Phenotype of Pancreatic Cancer Cells, and Reduces Tumor Volume and Favors T-Cell Infiltrates in Mice. Cancers, 14(24), 6133. https://doi.org/10.3390/cancers14246133