


Natural Killer Cells in Chronic Lymphocytic Leukemia: Functional Impairment and Therapeutic Potential


{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Background
1.1. Introduction to CLL and Its Immunosuppressive Effects
1.2. Natural Killer Cells
1.3. Evidence for NK Cell Importance in Leukemia
2. NK Dysfunction in CLL
2.1. Counts and Cytotoxicity
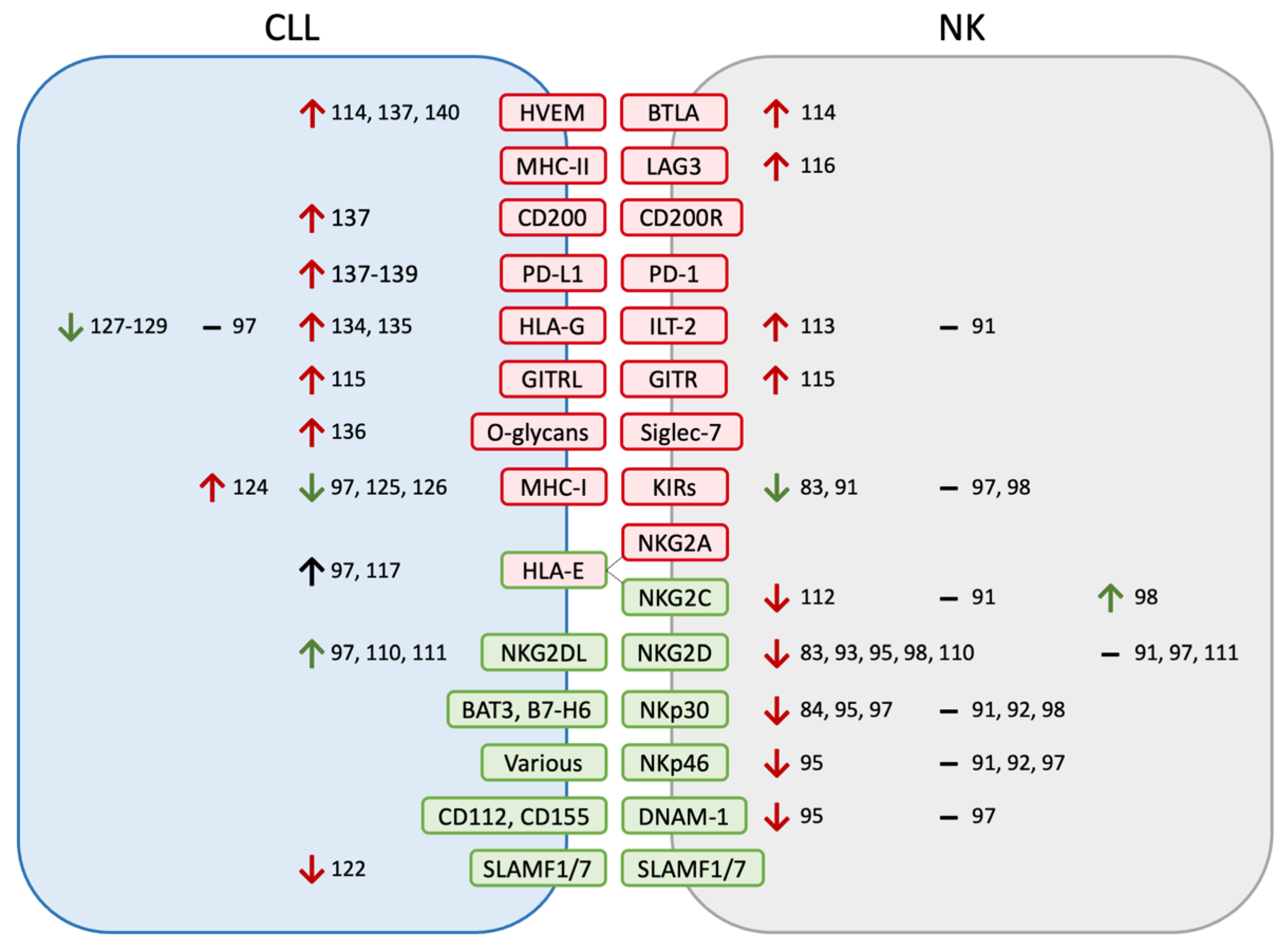
2.2. NK Phenotypes
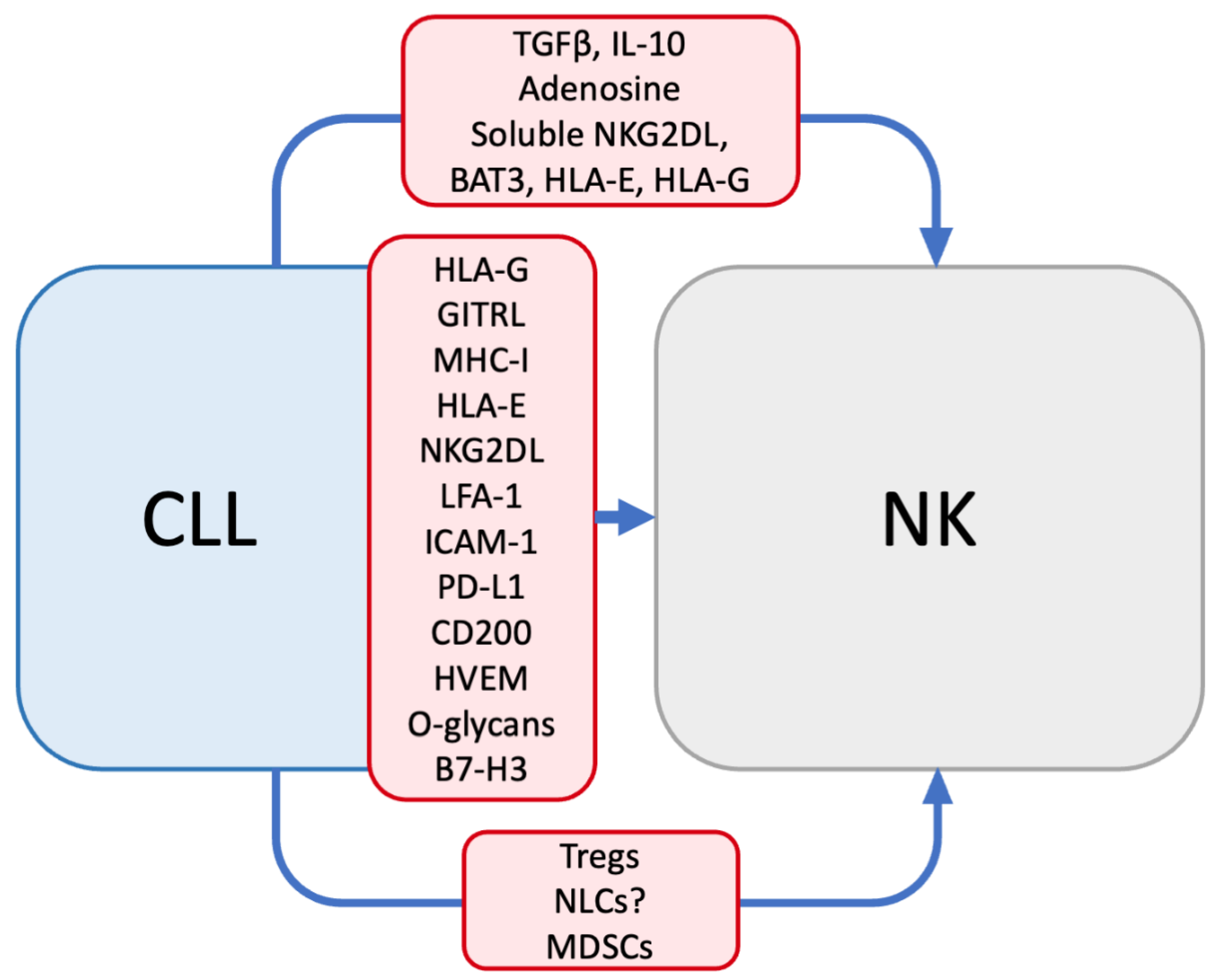
3. Mechanisms of NK Dysfunction in CLL
3.1. Receptor–Ligand Interactions
3.2. Soluble Factors
3.3. Altered Immune Populations
4. CLL Resistance to NK Cytotoxicity
5. Effects of CLL Therapeutics on NK Function
5.1. BTK Inhibitors
5.2. PI3K Inhibitors
5.3. Venetoclax
5.4. Antibodies
5.5. Cytotoxic Chemotherapy
5.6. Lenalidomide
6. Advantages of NK Therapy for CLL
7. Previous and Ongoing Attempts at NK Therapy for CLL
8. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Surveillance, Epidemiology, and End Results (SEER) Program. Cancer Stat Facts: Chronic Lymphocytic Leukemia (CLL). Available online: www.Seer.Cancer.Gov (accessed on 23 November 2022).
- Redaelli, A.; Laskin, B.L.; Stephens, J.M.; Botteman, M.F.; Pashos, C.L. The Clinical and Epidemiological Burden of Chronic Lymphocytic Leukaemia. Eur. J. Cancer Care 2004, 13, 279–287. [Google Scholar] [CrossRef] [PubMed]
- Hallek, M.; Cheson, B.D.; Catovsky, D.; Caligaris-Cappio, F.; Dighiero, G.; Döhner, H.; Hillmen, P.; Keating, M.; Montserrat, E.; Chiorazzi, N.; et al. IwCLL Guidelines for Diagnosis, Indications for Treatment, Response Assessment, and Supportive Management of CLL. Blood 2018, 131, 2745–2760. [Google Scholar] [CrossRef] [PubMed]
- Moreira, J.; Rabe, K.G.; Cerhan, J.R.; Kay, N.E.; Wilson, J.W.; Call, T.G.; Leis, J.F.; Jelinek, D.F.; Schwager, S.M.; Bowen, D.A.; et al. Infectious Complications among Individuals with Clinical Monoclonal B-Cell Lymphocytosis (MBL): A Cohort Study of Newly Diagnosed Cases Compared to Controls. Leukemia 2013, 27, 136–141. [Google Scholar] [CrossRef] [PubMed]
- Rossi, D.; De Paoli, L.; Rossi, F.M.; Cerri, M.; Deambrogi, C.; Rasi, S.; Zucchetto, A.; Capello, D.; Gattei, V.; Gaidano, G. Early Stage Chronic Lymphocytic Leukaemia Carrying Unmutated IGHV Genes Is at Risk of Recurrent Infections during Watch and Wait. Br. J. Haematol. 2008, 141, 734–736. [Google Scholar] [CrossRef] [PubMed]
- Williams, A.M.; Baran, A.M.; Meacham, P.J.; Feldman, M.M.; Valencia, H.E.; Newsom-Stewart, C.; Gupta, N.; Janelsins, M.C.; Barr, P.M.; Zent, C.S. Analysis of the Risk of Infection in Patients with Chronic Lymphocytic Leukemia in the Era of Novel Therapies. Leuk. Lymphoma 2017, 59, 625–632. [Google Scholar] [CrossRef] [PubMed]
- Hilal, T.; Gea-Banacloche, J.C.; Leis, J.F. Chronic Lymphocytic Leukemia and Infection Risk in the Era of Targeted Therapies: Linking Mechanisms with Infections. Blood Rev. 2018, 32, 387–399. [Google Scholar] [CrossRef]
- Dasanu, C.A.; Alexandrescu, D.T. Risk for Second Nonlymphoid Neoplasms in Chronic Lymphocytic Leukemia. MedGenMed 2007, 9, 35. [Google Scholar]
- Royle, J.A.; Baade, P.D.; Joske, D.; Girschik, J.; Fritschi, L. Second Cancer Incidence and Cancer Mortality among Chronic Lymphocytic Leukaemia Patients: A Population-Based Study. Br. J. Cancer 2011, 105, 1076–1081. [Google Scholar] [CrossRef]
- Solomon, B.M.; Rabe, K.G.; Slager, S.L.; Brewer, J.D.; Cerhan, J.R.; Shanafelt, T.D. Overall and Cancer-Specific Survival of Patients with Breast, Colon, Kidney, and Lung Cancers with and without Chronic Lymphocytic Leukemia: A SEER Population-Based Study. J. Clin. Oncol. 2013, 31, 930–937. [Google Scholar] [CrossRef]
- Brewer, J.D.; Shanafelt, T.D.; Otley, C.C.; Roenigk, R.K.; Cerhan, J.R.; Kay, N.E.; Weaver, A.L.; Call, T.G. Chronic Lymphocytic Leukemia Is Associated With Decreased Survival of Patients With Malignant Melanoma and Merkel Cell Carcinoma in a SEER Population-Based Study. J. Clin. Oncol. 2012, 30, 843–849. [Google Scholar] [CrossRef]
- Hodgson, K.; Ferrer, G.; Montserrat, E.; Moreno, C. Chronic Lymphocytic Leukemia and Autoimmunity: A Systematic Review. Haematologica 2011, 96, 752–761. [Google Scholar] [CrossRef] [PubMed]
- Rogers, K.A.; Woyach, J.A. Secondary Autoimmune Cytopenias in Chronic Lymphocytic Leukemia. Semin. Oncol. 2016, 43, 300–310. [Google Scholar] [CrossRef] [PubMed]
- Orsini, E.; Guarini, A.; Chiaretti, S.; Response, T.; Mauro, F.R.; Foa, R. The Circulating Dendritic Cell Compartment in Patients with Chronic Lymphocytic Leukemia Is Severely Defective and Unable to Stimulate an Effective T-Cell Response The Circulating Dendritic Cell Compartment in Patients with Chronic Lymphocytic Leukemia Is. Cancer Res. 2003, 63, 4497–4506. [Google Scholar]
- Zeya, H.I.; Keku, E.; Richards, F., 2nd; Spurr, C.L. Monocyte and Granulocyte Defect in Chronic Lymphocytic Leukemia. Am. J. Pathol. 1979, 95, 43–54. [Google Scholar] [PubMed]
- Maffei, R.; Bulgarelli, J.; Fiorcari, S.; Bertoncelli, L.; Martinelli, S.; Guarnotta, C.; Castelli, I.; Deaglio, S.; Debbia, G.; De Biasi, S.; et al. The Monocytic Population in Chronic Lymphocytic Leukemia Shows Altered Composition and Deregulation of Genes Involved in Phagocytosis and Inflammation. Haematologica 2013, 98, 1115–1123. [Google Scholar] [CrossRef] [PubMed]
- Jurado-Camino, T.; Córdoba, R.; Esteban-Burgos, L.; Hernández-Jiménez, E.; Toledano, V.; Hernandez-Rivas, J.-A.; Ruiz-Sainz, E.; Cobo, T.; Siliceo, M.; de Diego, R.; et al. Chronic Lymphocytic Leukemia: A Paradigm of Innate Immune Cross-Tolerance. J. Immunol. 2015, 194, 719–727. [Google Scholar] [CrossRef]
- Itälä, M.; Vainio, O.; Remes, K. Functional Abnormalities in Granulocytes Predict Susceptibility to Bacterial Infections in Chronic Lymphocytic Leukaemia. Eur. J. Haematol. 1996, 57, 46–53. [Google Scholar] [CrossRef]
- Kontoyiannis, D.P.; Georgiadou, S.P.; Wierda, W.G.; Wright, S.; Albert, N.D.; Ferrajoli, A.; Keating, M.; Lewis, R.E. Impaired Bactericidal but Not Fungicidal Activity of Polymorphonuclear Neutrophils in Patients with Chronic Lymphocytic Leukemia. Leuk. Lymphoma 2013, 54, 1730–1733. [Google Scholar] [CrossRef]
- Manukyan, G.; Papajik, T.; Gajdos, P.; Mikulkova, Z.; Urbanova, R.; Gabcova, G.; Kudelka, M.; Turcsányi, P.; Ryznerova, P.; Prochazka, V.; et al. Neutrophils in Chronic Lymphocytic Leukemia Are Permanently Activated and Have Functional Defects. Oncotarget 2017, 8, 84889–84901. [Google Scholar] [CrossRef]
- Yan, X.-J.; Dozmorov, I.; Li, W.; Yancopoulos, S.; Sison, C.; Centola, M.; Jain, P.; Allen, S.L.; Kolitz, J.E.; Rai, K.R.; et al. Identification of Outcome-Correlated Cytokine Clusters in Chronic Lymphocytic Leukemia. Blood 2011, 118, 5201–5210. [Google Scholar] [CrossRef]
- Heath, M.E.; Cheson, B.D. Defective Complement Activity in Chronic Lymphocytic Leukemia. Am. J. Hematol. 1985, 19, 63–73. [Google Scholar] [CrossRef]
- Füst, G.; Czink, E.; Minh, D.; Miszlay, Z.; Varga, L.; Hollán, S.R. Depressed Classical Complement Pathway Activities in Chronic Lymphocytic Leukaemia. Clin. Exp. Immunol. 1985, 60, 489–495. [Google Scholar]
- Füst, G.; Miszlay, Z.; Czink, E.; Varga, L.; Pálóczi, K.; Szegedi, G.; Hollán, S.R. C1 and C4 Abnormalities in Chronic Lymphocytic Leukaemia and Their Significance. Immunol. Lett. 1987, 14, 255–259. [Google Scholar] [CrossRef] [PubMed]
- Schlesinger, M.; Broman, I.; Lugassy, G. The Complement System Is Defective in Chronic Lymphatic Leukemia Patients and in Their Healthy Relatives. Leukemia 1996, 10, 1509–1513. [Google Scholar] [PubMed]
- Gustafson, M.P.; Abraham, R.S.; Lin, Y.; Wu, W.; Gastineau, D.A.; Zent, C.S.; Dietz, A.B. Association of an Increased Frequency of CD14+ HLA-DR Lo/Neg Monocytes with Decreased Time to Progression in Chronic Lymphocytic Leukaemia (CLL). Br. J. Haematol. 2012, 156, 674–676. [Google Scholar] [CrossRef] [PubMed]
- Jitschin, R.; Braun, M.; Büttner, M.; Dettmer-Wilde, K.; Bricks, J.; Berger, J.; Eckart, M.J.; Krause, S.W.; Oefner, P.J.; Le Blanc, K.; et al. CLL-Cells Induce IDOhi CD14+HLA-DRlo Myeloid-Derived Suppressor Cells That Inhibit T-Cell Responses and Promote TRegs. Blood 2014, 124, 750–760. [Google Scholar] [CrossRef]
- Ramsay, A.G.; Johnson, A.J.; Lee, A.M.; Gorgün, G.; Dieu, R.L.; Blum, W.; Byrd, J.C.; Gribben, J.G. Chronic Lymphocytic Leukemia T Cells Show Impaired Immunological Synapse Formation That Can Be Reversed with an Immunomodulating Drug. J. Clin. Investig. 2008, 118, 2427–2437. [Google Scholar] [CrossRef]
- Riches, J.C.; Davies, J.K.; McClanahan, F.; Fatah, R.; Iqbal, S.; Agrawal, S.; Ramsay, A.G.; Gribben, J.G. T Cells from CLL Patients Exhibit Features of T-Cell Exhaustion but Retain Capacity for Cytokine Production. Blood 2013, 121, 1612–1621. [Google Scholar] [CrossRef]
- Dazzi, F.; D’Andrea, E.; Biasi, G.; De Silvestro, G.; Gaidano, G.; Schena, M.; Tison, T.; Vianello, F.; Girolami, A.; Caligaris-Cappio, F. Failure of B Cells of Chronic Lymphocytic Leukemia in Presenting Soluble and Alloantigens. Clin. Immunol. Immunopathol. 1995, 75, 26–32. [Google Scholar] [CrossRef]
- Motta, M.; Chiarini, M.; Ghidini, C.; Zanotti, C.; Lamorgese, C.; Caimi, L.; Rossi, G.; Imberti, L. Quantification of Newly Produced B and T Lymphocytes in Untreated Chronic Lymphocytic Leukemia Patients. J. Transl. Med. 2010, 8, 111. [Google Scholar] [CrossRef]
- Sampalo, A.; Navas, G.; Medina, F.; Segundo, C.; Cámara, C.; Brieva, J.A. Chronic Lymphocytic Leukemia B Cells Inhibit Spontaneous Ig Production by Autologous Bone Marrow Cells: Role of CD95-CD95L Interaction. Blood 2000, 96, 3168–3174. [Google Scholar] [CrossRef] [PubMed]
- De Weerdt, I.; van Hoeven, V.; Munneke, J.M.; Endstra, S.; Hofland, T.; Hazenberg, M.D.; Kater, A.P. Innate Lymphoid Cells Are Expanded and Functionally Altered in Chronic Lymphocytic Leukemia. Haematologica 2016, 101, e461–e464. [Google Scholar] [CrossRef] [PubMed]
- Howell, K.M. The Failure of Antibody Formation in Leukemia. Arch. Intern. Med. 1920, 26, 706–714. [Google Scholar] [CrossRef][Green Version]
- Hudson, R.P.; Wilson, S.J. Hypogammaglobulinemia and Chronic Lymphatic Leukemia. Cancer 1960, 13, 200–204. [Google Scholar] [CrossRef] [PubMed]
- Fiddes, P.; Penny, R.; Wells, J.V.; Rozenberg, M.C. Clinical Correlations with Immunoglobulin Levels in Chronic Lymphatic Leukaemia§. Aust. N. Z. J. Med. 1972, 2, 346–350. [Google Scholar] [CrossRef]
- Freeman, J.A.; Crassini, K.R.; Best, O.G.; Forsyth, C.J.; Mackinlay, N.J.; Han, P.; Stevenson, W.; Mulligan, S.P. Immunoglobulin G Subclass Deficiency and Infection Risk in 150 Patients with Chronic Lymphocytic Leukemia. Leuk. Lymphoma 2013, 54, 99–104. [Google Scholar] [CrossRef]
- Burger, J.A.; Tsukada, N.; Burger, M.; Zvaifler, N.J.; Dell’Aquila, M.; Kipps, T.J. Blood-Derived Nurse-like Cells Protect Chronic Lymphocytic Leukemia B Cells from Spontaneous Apoptosis through Stromal Cell–Derived Factor-1. Blood 2000, 96, 2655–2663. [Google Scholar] [CrossRef]
- Herishanu, Y.; Pe, P.; Liu, D.; Pittaluga, S.; Vire, B.; Gibellini, F.; Njuguna, N.; Lee, E.; Stennett, L.; Raghavachari, N.; et al. The Lymph Node Microenvironment Promotes B-Cell Receptor Signaling, NF-KB Activation, and Tumor Proliferation in Chronic Lymphocytic Leukemia. Blood 2011, 117, 563–575. [Google Scholar] [CrossRef]
- Bagnara, D.; Kaufman, M.S.; Calissano, C.; Marsilio, S.; Patten, P.E.M.; Simone, R.; Chum, P.; Yan, X.-J.; Allen, S.L.; Kolitz, J.E.; et al. A Novel Adoptive Transfer Model of Chronic Lymphocytic Leukemia Suggests a Key Role for T Lymphocytes in the Disease. Blood 2011, 117, 5463–5472. [Google Scholar] [CrossRef]
- Riches, J.C.; Gribben, J.G. Understanding the Immunodeficiency in Chronic Lymphocytic Leukemia. Potential Clinical Implications. Hematol. Oncol. Clin. N. Am. 2013, 27, 207–235. [Google Scholar] [CrossRef]
- Hanna, B.S.; Öztürk, S.; Seiffert, M. Beyond Bystanders: Myeloid Cells in Chronic Lymphocytic Leukemia. Mol. Immunol. 2019, 110, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Arruga, F.; Gyau, B.B.; Iannello, A.; Vitale, N.; Vaisitti, T.; Deaglio, S. Immune Response Dysfunction in Chronic Lymphocytic Leukemia: Dissecting Molecular Mechanisms and Microenvironmental Conditions. Int. J. Mol. Sci. 2020, 21, 1825. [Google Scholar] [CrossRef] [PubMed]
- Forconi, F.; Moss, P. Perturbation of the Normal Immune System in Patients with CLL. Blood 2015, 126, 573–581. [Google Scholar] [CrossRef] [PubMed]
- Kay, N.E.; Perri, R.T. Evidence That Large Granular Lymphocytes from B-CLL Patients with Hypogammaglobulinemia down-Regulate B-Cell Immunoglobulin Synthesis. Blood 1989, 73, 1016–1019. [Google Scholar] [CrossRef]
- Vivier, E.; Tomasello, E.; Baratin, M.; Walzer, T.; Ugolini, S. Functions of Natural Killer Cells. Nat. Immunol. 2008, 9, 503–510. [Google Scholar] [CrossRef]
- Imai, K.; Matsuyama, S.; Miyake, S.; Suga, K.; Nakachi, K. Natural Cytotoxic Activity of Peripheral-Blood Lymphocytes and Cancer Incidence: An 11-Year Follow-up Study of a General Population. Lancet 2000, 356, 1795–1799. [Google Scholar] [CrossRef]
- Coca, S.; Perez-Piqueras, J.; Martinez, D.; Colmenarejo, A.; Saez, M.A.; Vallejo, C.; Martos, J.A.; Moreno, M. The Prognostic Significance of Intratumoral Natural Killer Cells in Patients with Colorectal Carcinoma. Cancer 1997, 79, 2320–2328. [Google Scholar] [CrossRef]
- Ishigami, S.; Natsugoe, S.; Tokuda, K.; Nakajo, A.; Che, X.; Iwashige, H.; Aridome, K.; Hokita, S.; Aikou, T. Prognostic Value of Intratumoral Natural Killer Cells in Gastric Carcinoma. Cancer 2000, 88, 577–583. [Google Scholar] [CrossRef]
- Takanami, I.; Takeuchi, K.; Giga, M. The Prognostic Value of Natural Killer Cell Infiltration in Resected Pulmonary Adenocarcinoma. J. Thorac. Cardiovasc. Surg. 2001, 121, 1058–1063. [Google Scholar] [CrossRef]
- Villegas, F.R.; Coca, S.; Villarrubia, V.G.; Jiménez, R.; Chillón, M.J.; Jareño, J.; Zuil, M.; Callol, L. Prognostic Significance of Tumor Infiltrating Natural Killer Cells Subset CD57 in Patients with Squamous Cell Lung Cancer. Lung Cancer 2002, 35, 23–28. [Google Scholar] [CrossRef]
- Schuster, I.S.; Coudert, J.D.; Andoniou, C.E.; Degli-Esposti, M.A. “Natural Regulators”: NK Cells as Modulators of T Cell Immunity. Front. Immunol. 2016, 7, 235. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Freud, A.G.; Caligiuri, M.A. Location and Cellular Stages of Natural Killer Cell Development. Trends Immunol. 2013, 34, 573–582. [Google Scholar] [CrossRef] [PubMed]
- Horowitz, A.; Strauss-Albee, D.M.; Leipold, M.; Kubo, J.; Nemat-Gorgani, N.; Dogan, O.C.; Dekker, C.L.; Mackey, S.; Maecker, H.; Swan, G.E.; et al. Genetic and Environmental Determinants of Human NK Cell Diversity Revealed by Mass Cytometry. Sci. Transl. Med. 2013, 5, 208ra145. [Google Scholar] [CrossRef] [PubMed]
- Björkström, N.K.; Riese, P.; Heuts, F.; Andersson, S.; Fauriat, C.; Ivarsson, M.A.; Björklund, A.T.; Flodström-Tullberg, M.; Michaëlsson, J.; Rottenberg, M.E.; et al. Expression Patterns of NKG2A, KIR, and CD57 Define a Process of CD56dim NK-Cell Differentiation Uncoupled from NK-Cell Education. Blood 2010, 116, 3853–3864. [Google Scholar] [CrossRef]
- Pfefferle, A.; Jacobs, B.; Netskar, H.; Ask, E.H.; Lorenz, S.; Clancy, T.; Goodridge, J.P.; Sohlberg, E.; Malmberg, K.-J. Intra-Lineage Plasticity and Functional Reprogramming Maintain Natural Killer Cell Repertoire Diversity. Cell Rep. 2019, 29, 2284–2294.e4. [Google Scholar] [CrossRef]
- Pahl, J.H.W.; Cerwenka, A.; Ni, J. Memory-Like NK Cells: Remembering a Previous Activation by Cytokines and NK Cell Receptors. Front. Immunol. 2018, 9, 2796. [Google Scholar] [CrossRef]
- Thielens, A.; Vivier, E.; Romagné, F. NK Cell MHC Class I Specific Receptors (KIR): From Biology to Clinical Intervention. Curr. Opin. Immunol. 2012, 24, 239–245. [Google Scholar] [CrossRef]
- Long, E.O.; Kim, H.S.; Liu, D.; Peterson, M.E.; Rajagopalan, S. Controlling Natural Killer Cell Responses: Integration of Signals for Activation and Inhibition. Annu. Rev. Immunol. 2013, 31, 227–258. [Google Scholar] [CrossRef]
- Lanier, L.L. Up on the Tightrope: Natural Killer Cell Activation and Inhibition. Nat. Immunol. 2008, 9, 495–502. [Google Scholar] [CrossRef]
- Chan, C.J.; Smyth, M.J.; Martinet, L. Molecular Mechanisms of Natural Killer Cell Activation in Response to Cellular Stress. Cell Death Differ. 2014, 21, 5–14. [Google Scholar] [CrossRef]
- Barrow, A.D.; Martin, C.J.; Colonna, M. The Natural Cytotoxicity Receptors in Health and Disease. Front. Immunol. 2019, 10, 909. [Google Scholar] [CrossRef] [PubMed]
- Prager, I.; Watzl, C. Mechanisms of Natural Killer Cell-Mediated Cellular Cytotoxicity. J. Leukoc. Biol. 2019, 105, 1319–1329. [Google Scholar] [CrossRef] [PubMed]
- Ruggeri, L.; Capanni, M.; Urbani, E.; Perruccio, K.; Shlomchik, W.D.; Tosti, A.; Posati, S.; Rogaia, D.; Frassoni, F.; Aversa, F.; et al. Effectiveness of Donor Natural Killer Cell Alloreactivity in Mismatched Hematopoietic Transplants. Science 2002, 295, 2097–2100. [Google Scholar] [CrossRef]
- Gao, F.; Ye, Y.; Gao, Y.; Huang, H.; Zhao, Y. Influence of KIR and NK Cell Reconstitution in the Outcomes of Hematopoietic Stem Cell Transplantation. Front. Immunol. 2020, 11, 2022. [Google Scholar] [CrossRef] [PubMed]
- McClanahan, F.; Gribben, J. Transplantation in Chronic Lymphocytic Leukemia: Does It Still Matter in the Era of Novel Targeted Therapies? Hematol. Oncol. Clin. N. Am. 2014, 28, 1055–1071. [Google Scholar] [CrossRef]
- Bachanova, V.; Weisdorf, D.J.; Wang, T.; Marsh, S.G.E.; Cereb, N.; Haagenson, M.D.; Spellman, S.R.; Lee, S.J.; Guethlein, L.A.; Parham, P.; et al. Donor Killer Cell Immunoglobulin-Like Receptor Genotype Does Not Improve Graft-versus-Leukemia Responses in Chronic Lymphocytic Leukemia after Unrelated Donor Transplant: A Center for International Blood and Marrow Transplant Research Analysis. Biol. Blood Marrow Transplant. 2019, 25, 949–954. [Google Scholar] [CrossRef] [PubMed]
- Karabon, L.; Jedynak, A.; Giebel, S.; Wołowiec, D.; Kielbinski, M.; Woszczyk, D.; Kapelko-Slowik, K.; Kuliczkowski, K.; Frydecka, I. KIR/HLA Gene Combinations Influence Susceptibility to B-Cell Chronic Lymphocytic Leukemia and the Clinical Course of Disease. Tissue Antigens 2011, 78, 129–138. [Google Scholar] [CrossRef] [PubMed]
- Fischer, L.; Penack, O.; Gentilini, C.; Nogai, A.; Muessig, A.; Thiel, E.; Uharek, L. The Anti-Lymphoma Effect of Antibody-Mediated Immunotherapy Is Based on an Increased Degranulation of Peripheral Blood Natural Killer (NK) Cells. Exp. Hematol. 2006, 34, 753–759. [Google Scholar] [CrossRef]
- Bologna, L.; Gotti, E.; Manganini, M.; Rambaldi, A.; Intermesoli, T.; Introna, M.; Golay, J. Mechanism of Action of Type II, Glycoengineered, Anti-CD20 Monoclonal Antibody GA101 in B-Chronic Lymphocytic Leukemia Whole Blood Assays in Comparison with Rituximab and Alemtuzumab. J. Immunol. 2011, 186, 3762–3769. [Google Scholar] [CrossRef]
- Wu, J.; Edberg, J.C.; Redecha, P.B.; Bansal, V.; Guyre, P.M.; Coleman, K.; Salmon, J.E.; Kimberly, R.P. A Novel Polymorphism of FcgammaRIIIa (CD16) Alters Receptor Function and Predisposes to Autoimmune Disease. J. Clin. Investig. 1997, 100, 1059–1070. [Google Scholar] [CrossRef]
- Koene, H.R.; Kleijer, M.; Algra, J.; Roos, D.; von dem Borne, A.E.; de Haas, M. Fc GammaRIIIa-158V/F Polymorphism Influences the Binding of IgG by Natural Killer Cell Fc GammaRIIIa, Independently of the Fc GammaRIIIa-48L/R/H Phenotype. Blood 1997, 90, 1109–1114. [Google Scholar] [CrossRef] [PubMed]
- Cartron, G.; Dacheux, L.; Salles, G.; Solal-Celigny, P.; Bardos, P.; Colombat, P.; Watier, H. Therapeutic Activity of Humanized Anti-CD20 Monoclonal Antibody and Polymorphism in IgG Fc Receptor FcγRIIIa Gene. Blood 2002, 99, 754–758. [Google Scholar] [CrossRef] [PubMed]
- Weng, W.-K.; Levy, R. Two Immunoglobulin G Fragment C Receptor Polymorphisms Independently Predict Response to Rituximab in Patients With Follicular Lymphoma. J. Clin. Oncol. 2003, 21, 3940–3947. [Google Scholar] [CrossRef] [PubMed]
- Treon, S.P.; Hansen, M.; Branagan, A.R.; Verselis, S.; Emmanouilides, C.; Kimby, E.; Frankel, S.R.; Touroutoglou, N.; Turnbull, B.; Anderson, K.C.; et al. Polymorphisms in FcγRIIIA (CD16) Receptor Expression Are Associated With Clinical Response to Rituximab in Waldenström’s Macroglobulinemia. J. Clin. Oncol. 2005, 23, 474–481. [Google Scholar] [CrossRef]
- Farag, S.S.; Flinn, I.W.; Modali, R.; Lehman, T.A.; Young, D.; Byrd, J.C. FcγRIIIa and FcγRIIa Polymorphisms Do Not Predict Response to Rituximab in B-Cell Chronic Lymphocytic Leukemia. Blood 2004, 103, 1472–1474. [Google Scholar] [CrossRef]
- Dornan, D.; Spleiss, O.; Yeh, R.-F.; Duchateau-Nguyen, G.; Dufour, A.; Zhi, J.; Robak, T.; Moiseev, S.I.; Dmoszynska, A.; Solal-Celigny, P.; et al. Effect of FCGR2A and FCGR3A Variants on CLL Outcome. Blood 2010, 116, 4212–4222. [Google Scholar] [CrossRef]
- Miller, J.S.; Soignier, Y.; Panoskaltsis-Mortari, A.; McNearney, S.A.; Yun, G.H.; Fautsch, S.K.; McKenna, D.; Le, C.; Defor, T.E.; Burns, L.J.; et al. Successful Adoptive Transfer and in Vivo Expansion of Human Haploidentical NK Cells in Patients with Cancer. Cancer 2005, 105, 3051–3057. [Google Scholar] [CrossRef]
- Choi, I.; Yoon, S.R.; Park, S.-Y.; Kim, H.; Jung, S.-J.; Jang, Y.J.; Kang, M.; Yeom, Y., II; Lee, J.-L.; Kim, D.-Y.; et al. Donor-Derived Natural Killer Cells Infused after Human Leukocyte Antigen–Haploidentical Hematopoietic Cell Transplantation: A Dose-Escalation Study. Biol. Blood Marrow Transplant. 2014, 20, 696–704. [Google Scholar] [CrossRef]
- Romee, R.; Rosario, M.; Berrien-Elliott, M.M.; Wagner, J.A.; Jewell, B.A.; Schappe, T.; Leong, J.W.; Abdel-Latif, S.; Schneider, S.E.; Willey, S.; et al. Cytokine-Induced Memory-like Natural Killer Cells Exhibit Enhanced Responses against Myeloid Leukemia. Sci. Transl. Med. 2016, 8, 357ra123. [Google Scholar] [CrossRef]
- Kimby, E.; Mellstedt, H.; Nilsson, B.; Bjorkholm, M.; Holm, G. Differences in Blood T and NK Cell Populations between Chronic Lymphocytic Leukemia of B Cell Type (B-CLL) and Monoclonal B-Lymphocytosis of Undetermined Significance (B-MLUS). Leukemia 1989, 3, 501–504. [Google Scholar]
- Villamor, N.; Reverter, J.C.; Montserrat, E.; Urbano-Ispízua, A.; Vives-Corrons, J.L.; Rozman, C. Recombinant Alpha-2b-Interferon May Restore Natural-Killer Activity in Patients with B-Chronic Lymphocytic Leukemia. Leukemia 1992, 6, 547–552. [Google Scholar] [PubMed]
- MacFarlane, A.W.; Jillab, M.; Smith, M.R.; Alpaugh, R.K.; Cole, M.E.; Litwin, S.; Millenson, M.M.; Al-Saleem, T.; Cohen, A.D.; Campbell, K.S. NK Cell Dysfunction in Chronic Lymphocytic Leukemia Is Associated with Loss of the Mature Cells Expressing Inhibitory Killer Cell Ig-like Receptors. Oncoimmunology 2017, 6, e1330235. [Google Scholar] [CrossRef] [PubMed]
- Hadadi, L.; Hafezi, M.; Amirzargar, A.A.; Sharifian, R.A.; Abediankenari, S.; Asgarian-Omran, H. Dysregulated Expression of Tim-3 and NKp30 Receptors on NK Cells of Patients with Chronic Lymphocytic Leukemia. Oncol. Res. Treat. 2019, 42, 197–203. [Google Scholar] [CrossRef] [PubMed]
- De Weerdt, I.; Hofland, T.; de Boer, R.; Dobber, J.A.; Dubois, J.; van Nieuwenhuize, D.; Mobasher, M.; de Boer, F.; Hoogendoorn, M.; Velders, G.A.; et al. Distinct Immune Composition in Lymph Node and Peripheral Blood of CLL Patients Is Reshaped during Venetoclax Treatment. Blood Adv. 2019, 3, 2642–2652. [Google Scholar] [CrossRef] [PubMed]
- Reyes, E.; Prieto, A.; Carrion, F.; Garcia-Suarez, J.; Esquivel, F.; Alvarez-Mon, M. Morphological Variants of Leukemic Cells in B Chronic Lymphocytic Leukemia Are Associated with Different T Cell and NK Cell Abnormalities. Am. J. Hematol. 1997, 55, 175–182. [Google Scholar] [CrossRef]
- Lundin, J.; Porwit-MacDonald, A.; Rossmann, E.D.; Karlsson, C.; Edman, P.; Rezvany, M.R.; Kimby, E.; Österborg, A.; Mellstedt, H. Cellular Immune Reconstitution after Subcutaneous Alemtuzumab (Anti-CD52 Monoclonal Antibody, CAMPATH-1H) Treatment as First-Line Therapy for B-Cell Chronic Lymphocytic Leukaemia. Leukemia 2004, 18, 484–490. [Google Scholar] [CrossRef]
- Palmer, S.; Hanson, C.A.; Zent, C.S.; Porrata, L.F.; LaPlant, B.; Geyer, S.M.; Markovic, S.N.; Call, T.G.; Bowen, D.A.; Jelinek, D.F.; et al. Prognostic Importance of T and NK-Cells in a Consecutive Series of Newly Diagnosed Patients with Chronic Lymphocytic Leukaemia. Br. J. Haematol. 2008, 141, 607–614. [Google Scholar] [CrossRef]
- Gonzalez-Rodriguez, A.P.; Contesti, J.; Huergo-Zapico, L.; Lopez-Soto, A.; Fernández-Guizán, A.; Acebes-Huerta, A.; Gonzalez-Huerta, A.J.; Gonzalez, E.; Fernandez-Alvarez, C.; Gonzalez, S. Prognostic Significance of CD8 and CD4 T Cells in Chronic Lymphocytic Leukemia. Leuk. Lymphoma 2010, 51, 1829–1836. [Google Scholar] [CrossRef]
- D’Arena, G.; Laurenti, L.; Minervini, M.M.; Deaglio, S.; Bonello, L.; De Martino, L.; De Padua, L.; Savino, L.; Tarnani, M.; De Feo, V.; et al. Regulatory T-Cell Number Is Increased in Chronic Lymphocytic Leukemia Patients and Correlates with Progressive Disease. Leuk. Res. 2011, 35, 363–368. [Google Scholar] [CrossRef]
- Le Garff-Tavernier, M.; Decocq, J.; de Romeuf, C.; Parizot, C.; Dutertre, C.A.; Chapiro, E.; Davi, F.; Debré, P.; Prost, J.F.; Teillaud, J.L.; et al. Analysis of CD16+CD56dim NK Cells from CLL Patients: Evidence Supporting a Therapeutic Strategy with Optimized Anti-CD20 Monoclonal Antibodies. Leukemia 2011, 25, 101–109. [Google Scholar] [CrossRef]
- Costello, R.T.; Knoblauch, B.; Sanchez, C.; Mercier, D.; Le Treut, T.; Sébahoun, G. Expression of Natural Killer Cell Activating Receptors in Patients with Chronic Lymphocytic Leukaemia. Immunology 2012, 135, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Huergo-Zapico, L.; Acebes-Huerta, A.; Gonzalez-Rodriguez, A.P.; Contesti, J.; Gonzalez-Garcia, E.; Payer, A.R.; Villa-Alvarez, M.; Fernández-Guizán, A.; López-Soto, A.; Gonzalez, S. Expansion of NK Cells and Reduction of NKG2D Expression in Chronic Lymphocytic Leukemia. Correlation with Progressive Disease. PLoS ONE 2014, 9, e108326. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.-T.; Zhu, H.-Y.; Wu, Y.-J.; Xia, Y.; Wu, J.-Z.; Wu, W.; Liang, J.-H.; Wang, L.; Fan, L.; Li, J.-Y.; et al. Elevated Absolute NK Cell Counts in Peripheral Blood Predict Good Prognosis in Chronic Lymphocytic Leukemia. J. Cancer Res. Clin. Oncol. 2018, 144, 449–457. [Google Scholar] [CrossRef]
- Parry, H.M.; Stevens, T.; Oldreive, C.; Zadran, B.; McSkeane, T.; Rudzki, Z.; Paneesha, S.; Chadwick, C.; Stankovic, T.; Pratt, G.; et al. NK Cell Function Is Markedly Impaired in Patients with Chronic Lymphocytic Leukaemia but Is Preserved in Patients with Small Lymphocytic Lymphoma. Oncotarget 2016, 7, 68513–68526. [Google Scholar] [CrossRef] [PubMed]
- Eskelund, C.W.; Nederby, L.; Thysen, A.H.; Skovbo, A.; Roug, A.S.; Hokland, M.E. Interleukin-21 and Rituximab Enhance NK Cell Functionality in Patients with B-Cell Chronic Lymphocytic Leukaemia. Leuk. Res. 2011, 35, 914–920. [Google Scholar] [CrossRef]
- Veuillen, C.; Aurran-Schleinitz, T.; Castellano, R.; Rey, J.; Mallet, F.; Orlanducci, F.; Pouyet, L.; Just-Landi, S.; Coso, D.; Ivanov, V.; et al. Primary B-CLL Resistance to NK Cell Cytotoxicity Can Be Overcome in Vitro and in Vivo by Priming NK Cells and Monoclonal Antibody Therapy. J. Clin. Immunol. 2012, 32, 632–646. [Google Scholar] [CrossRef]
- Hofland, T.; Endstra, S.; Gomes, C.K.P.; de Boer, R.; de Weerdt, I.; Bobkov, V.; Riedl, J.A.; Heukers, R.; Smit, M.J.; Eldering, E.; et al. Natural Killer Cell Hypo-Responsiveness in Chronic Lymphocytic Leukemia Can Be Circumvented In Vitro by Adequate Activating Signaling. HemaSphere 2019, 3, e308. [Google Scholar] [CrossRef]
- Lotzová, E.; Savary, C.A.; Herberman, R.B. Induction of NK Cell Activity against Fresh Human Leukemia in Culture with Interleukin 2. J. Immunol. 1987, 138, 2718–2727. [Google Scholar]
- Jewell, A.P.; Worman, C.P.; Giles, F.J.; Goldstone, A.H.; Lydyard, P.M. Resistance of Chronic Lymphocytic Leukaemia Cells to Interferon-α Generated Lymphokine Activated Killer Cells. Leuk. Lymphoma 1992, 7, 473–480. [Google Scholar] [CrossRef]
- Katrinakis, G.; Kyriakou, D.; Papadaki, H.; Kalokyri, I.; Markidou, F.; Eliopoulos, G.D. Defective Natural Killer Cell Activity in B-Cell Chronic Lymphocytic Leukaemia Is Associated with Impaired Release of Natural Killer Cytotoxic Factor(s) but Not of Tumour Necrosis Factor-α. Acta Haematol. 1996, 96, 16–23. [Google Scholar] [CrossRef]
- Weitzman, J.; Betancur, M.; Boissel, L.; Rabinowitz, A.P.; Klein, A.; Klingemann, H. Variable Contribution of Monoclonal Antibodies to ADCC in Patients with Chronic Lymphocytic Leukemia. Leuk. Lymphoma 2009, 50, 1361–1368. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, R.; Audrito, V.; Vacca, P.; Rossi, D.; Brusa, D.; Stignani, M.; Bortolotti, D.; D’Arena, G.; Coscia, M.; Laurenti, L.; et al. HLA-G Is a Component of the Chronic Lymphocytic Leukemia Escape Repertoire to Generate Immune Suppression: Impact of the HLA-G 14 Base Pair (Rs66554220) Polymorphism. Haematologica 2014, 99, 888–896. [Google Scholar] [CrossRef] [PubMed]
- Platsoucas, C.D.; Fernandes, G.; Gupta, S.L.; Kempin, S.; Clarkson, B.; Good, R.A.; Gupta, S. Defective Spontaneous and Antibody-Dependent Cytotoxicity Mediated by E-Rosette-Positive and E-Rosette-Negative Cells in Untreated Patients with Chronic Lymphocytic Leukemia: Augmentation by in Vitro Treatment with Interferon. J. Immunol. 1980, 125, 1216–1223. [Google Scholar]
- Ziegler, H.; Kay, N.E.; Zarling, J.M. Deficiency of Natural Killer Cell Activity in Patients with Chronic Lymphocytic Leukemia. Int. J. Cancer 1981, 27, 321–327. [Google Scholar] [CrossRef] [PubMed]
- Pattengale, P.K.; Gidlund, M.; Nilsson, K.; Sundstrom, C.; Sallstrom, J.; Simonsson, B.; Wigzell, H. Lysis of Fresh Human B-Lymphocyte-Derived Leukemia Cells by Interferon-Activated Natural Killer (NK) Cells. Int. J. Cancer 1982, 29, 1–7. [Google Scholar] [CrossRef]
- Kay, N.E.; Zarling, J.M. Impaired Natural Killer Activity in Patients With Chronic Lymphocytic Leukemia Is Associated With a Deficiency of Azurophilic Cytoplasmic Granules in Putative NK Cells. Blood 1984, 63, 305–309. [Google Scholar] [CrossRef]
- Kay, N.E.; Zarling, J. Restoration of Impaired Natural Killer Cell Activity of B-Chronic Lymphocytic Leukemia Patients by Recombinant Lnterleukin-2. Am. J. Hematol. 1987, 24, 161–167. [Google Scholar] [CrossRef]
- Alvarez-Mon, M.; Casas, J.; Laguna, R.; Jordá, J.; Durantez, A. Clinical Signification of Natural Killer Activity in B-Cell Chronic Lymphocytic Leukemia. Eur. J. Haematol. 1987, 38, 268–273. [Google Scholar] [CrossRef]
- Hilpert, J.; Grosse-Hovest, L.; Grunebach, F.; Buechele, C.; Nuebling, T.; Raum, T.; Steinle, A.; Salih, H.R. Comprehensive Analysis of NKG2D Ligand Expression and Release in Leukemia: Implications for NKG2D-Mediated NK Cell Responses. J. Immunol. 2012, 189, 1360–1371. [Google Scholar] [CrossRef]
- Nückel, H.; Switala, M.; Sellmann, L.; Horn, P.A.; Dürig, J.; Dührsen, U.; Küppers, R.; Grosse-Wilde, H.; Rebmann, V. The Prognostic Significance of Soluble NKG2D Ligands in B-Cell Chronic Lymphocytic Leukemia. Leukemia 2010, 24, 1152–1159. [Google Scholar] [CrossRef]
- Puiggros, A.; Blanco, G.; Muntasell, A.; Rodríguez-Rivera, M.; Nonell, L.; Altadill, M.; Puigdecanet, E.; Arnal, M.; Calvo, X.; Gimeno, E.; et al. Reduced Expansion of CD94/NKG2C+ NK Cells in Chronic Lymphocytic Leukemia and CLL-like Monoclonal B-Cell Lymphocytosis Is Not Related to Increased Human Cytomegalovirus Seronegativity or NKG2C Deletions. Int. J. Lab. Hematol. 2021, 43, 1032–1040. [Google Scholar] [CrossRef] [PubMed]
- Villa-Álvarez, M.; Sordo-Bahamonde, C.; Lorenzo-Herrero, S.; Gonzalez-Rodriguez, A.P.; Payer, A.R.; Gonzalez-Garcia, E.; Villa-Álvarez, M.C.; López-Soto, A.; Gonzalez, S. Ig-Like Transcript 2 (ILT2) Blockade and Lenalidomide Restore NK Cell Function in Chronic Lymphocytic Leukemia. Front. Immunol. 2018, 9, 2917. [Google Scholar] [CrossRef] [PubMed]
- Sordo-Bahamonde, C.; Lorenzo-Herrero, S.; Gonzalez-Rodriguez, A.P.; Payer, Á.R.; González-García, E.; López-Soto, A.; Gonzalez, S. BTLA/HVEM Axis Induces NK Cell Immunosuppression and Poor Outcome in Chronic Lymphocytic Leukemia. Cancers 2021, 13, 1766. [Google Scholar] [CrossRef]
- Buechele, C.; Baessler, T.; Wirths, S.; Schmohl, J.U.; Schmiedel, B.J.; Salih, H.R. Glucocorticoid-Induced TNFR-Related Protein (GITR) Ligand Modulates Cytokine Release and NK Cell Reactivity in Chronic Lymphocytic Leukemia (CLL). Leukemia 2012, 26, 991–1000. [Google Scholar] [CrossRef] [PubMed]
- Sordo-Bahamonde, C.; Lorenzo-Herrero, S.; González-Rodríguez, A.P.; Payer, Á.R.; González-García, E.; López-Soto, A.; Gonzalez, S. LAG-3 Blockade with Relatlimab (BMS-986016) Restores Anti-Leukemic Responses in Chronic Lymphocytic Leukemia. Cancers 2021, 13, 2112. [Google Scholar] [CrossRef]
- McWilliams, E.M.; Mele, J.M.; Cheney, C.; Timmerman, E.A.; Fiazuddin, F.; Strattan, E.J.; Mo, X.; Byrd, J.C.; Muthusamy, N.; Awan, F.T. Therapeutic CD94/NKG2A Blockade Improves Natural Killer Cell Dysfunction in Chronic Lymphocytic Leukemia. Oncoimmunology 2016, 5, e1226720. [Google Scholar] [CrossRef] [PubMed]
- Krzywinska, E.; Allende-Vega, N.; Cornillon, A.; Vo, D.-N.; Cayrefourcq, L.; Panabieres, C.; Vilches, C.; Déchanet-Merville, J.; Hicheri, Y.; Rossi, J.-F.; et al. Identification of Anti-Tumor Cells Carrying Natural Killer (NK) Cell Antigens in Patients With Hematological Cancers. EBioMedicine 2015, 2, 1364–1376. [Google Scholar] [CrossRef]
- Krzywinska, E.; Cornillon, A.; Allende-Vega, N.; Vo, D.-N.; Rene, C.; Lu, Z.-Y.; Pasero, C.; Olive, D.; Fegueux, N.; Ceballos, P.; et al. CD45 Isoform Profile Identifies Natural Killer (NK) Subsets with Differential Activity. PLoS ONE 2016, 11, e0150434. [Google Scholar] [CrossRef]
- Burton, J.D.; Weitz, C.H.; Kay, N.E. Malignant Chronic Lymphocytic Leukemia B Cells Elaborate Soluble Factors That Down-regulate T Cell and NK Function. Am. J. Hematol. 1989, 30, 61–67. [Google Scholar] [CrossRef]
- Reiners, K.; Topolar, D.; Henke, A.; Simhadri, V.; Sauer, M.; Bessler, M.; Hansen, H.; Tawadros, S.; Herling, M.; Kr, M. Soluble Ligands for NK Cell Receptors Promote Evasion of Chronic Lymphocytic Leukemia Cells from NK Cell Anti-Tumor Activity. Blood 2015, 121, 3658–3666. [Google Scholar] [CrossRef]
- Coma, M.; Tothova, E.; Guman, T.; Hajikova, M.; Giertlova, M.; Sarissky, M. Altered Expression Pattern of SLAM Family Receptors on Pathological B Cells of Patients with Chronic Lymphocytic Leukemia. Leuk. Lymphoma 2017, 58, 1726–1729. [Google Scholar] [CrossRef] [PubMed]
- Von Wenserski, L.; Schultheiß, C.; Bolz, S.; Schliffke, S.; Simnica, D.; Willscher, E.; Gerull, H.; Wolters-Eisfeld, G.; Riecken, K.; Fehse, B.; et al. SLAMF Receptors Negatively Regulate B Cell Receptor Signaling in Chronic Lymphocytic Leukemia via Recruitment of Prohibitin-2. Leukemia 2021, 35, 1073–1086. [Google Scholar] [CrossRef] [PubMed]
- Nouri, A.M.; Smith, S.; Oliver, T.R.; Newland, A.C.; Macey, M.G. Comparative Expression of Major Histocompatibility Complex (MHC) Antigens on CD5+ and CD5- B Cells in Patients with Chronic Lymphocytic Leukaemia (CLL). Eur. J. Cancer 1998, 34, 1618–1622. [Google Scholar] [CrossRef]
- Demanet, C.; Mulder, A.; Deneys, V.; Worsham, M.J.; Maes, P.; Claas, F.H.; Ferrone, S. Down-Regulation of HLA-A and HLA-Bw6, but Not HLA-Bw4, Allospecificities in Leukemic Cells: An Escape Mechanism from CTL and NK Attack? Blood 2004, 103, 3122–3130. [Google Scholar] [CrossRef]
- Verheyden, S.; Ferrone, S.; Mulder, A.; Claas, F.H.; Schots, R.; De Moerloose, B.; Benoit, Y.; Demanet, C. Role of the Inhibitory KIR Ligand HLA-Bw4 and HLA-C Expression Levels in the Recognition of Leukemic Cells by Natural Killer Cells. Cancer Immunol. Immunother. 2008, 58, 855. [Google Scholar] [CrossRef] [PubMed]
- Amiot, L.; Onno, M.; Drénou, B.; Monvoisin, C.; Fauchet, R. HLA-G Class I Gene Expression in Normal and Malignant Hematopoietic Cells. Hum. Immunol. 1998, 59, 524–528. [Google Scholar] [CrossRef] [PubMed]
- Amiot, L.; Friec, G.L.; Sebti, Y.; Drénou, B.; Pangault, C.; Guilloux, V.; Leleu, X.; Bernard, M.; Facon, T.; Fauchet, R. HLA-G and Lymphoproliferative Disorders. Semin. Cancer Biol. 2003, 13, 379–385. [Google Scholar] [CrossRef]
- Poláková, K.; Kŕčová, M.; Kuba, D.; Russ, G. Analysis of HLA-G Expression in Malignant Hematopoetic Cells from Leukemia Patients. Leuk. Res. 2003, 27, 643–648. [Google Scholar] [CrossRef]
- Nückel, H.; Rebmann, V.; Dürig, J.; Dührsen, U.; Grosse-Wilde, H. HLA-G Expression Is Associated with an Unfavorable Outcome and Immunodeficiency in Chronic Lymphocytic Leukemia. Blood 2005, 105, 1694–1698. [Google Scholar] [CrossRef]
- Maki, G.; Hayes, G.M.; Naji, A.; Tyler, T.; Carosella, E.D.; Rouas-Freiss, N.; Gregory, S.A. NK Resistance of Tumor Cells from Multiple Myeloma and Chronic Lymphocytic Leukemia Patients: Implication of HLA-G. Leukemia 2008, 22, 998–1006. [Google Scholar] [CrossRef]
- Giannopoulos, K.; Dmoszyńska, A.; Bojarska-Junak, A.; Schmitt, M.; Roliński, J. Expression of HLA-G in Patients with B-Cell Chronic Lymphocytic Leukemia (B-CLL). Folia Histochem. Cytobiol. 2008, 46, 457–460. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Erikci, A.A.; Karagoz, B.; Ozyurt, M.; Ozturk, A.; Kilic, S.; Bilgi, O. HLA-G Expression in B Chronic Lymphocytic Leukemia: A New Prognostic Marker? Hematology 2009, 14, 101–105. [Google Scholar] [CrossRef] [PubMed]
- Perez-Chacon, G.; Rosado, S.; Rebolleda, N.; Losada-Fernandez, I.; Vargas, J.A.; Morado, M.; Jorda, J.; Perez-Aciego, P. Prognostic Irrelevance of HLA-G in B-Cell Chronic Lymphocytic Leukemia. Int. J. Lab. Hematol. 2009, 31, 327–337. [Google Scholar] [CrossRef] [PubMed]
- Własiuk, P.; Tomczak, W.; Zajac, M.; Dmoszyńska, A.; Giannopoulos, K. Total Expression of HLA-G and TLR-9 in Chronic Lymphocytic Leukemia Patients. Hum. Immunol. 2013, 74, 1592–1597. [Google Scholar] [CrossRef] [PubMed]
- Chang, L.-Y.; Liang, S.-Y.; Lu, S.-C.; Tseng, H.C.; Tsai, H.-Y.; Tang, C.-J.; Sugata, M.; Chen, Y.-J.; Chen, Y.-J.; Wu, S.-J.; et al. Molecular Basis and Role of Siglec-7 Ligand Expression on Chronic Lymphocytic Leukemia B Cells. Front. Immunol. 2022, 13, 840388. [Google Scholar] [CrossRef]
- Ramsay, A.G.; Clear, A.J.; Fatah, R.; Gribben, J.G. Multiple Inhibitory Ligands Induce Impaired T-Cell Immunologic Synapse Function in Chronic Lymphocytic Leukemia That Can Be Blocked with Lenalidomide: Establishing a Reversible Immune Evasion Mechanism in Human Cancer. Blood 2012, 120, 1412–1421. [Google Scholar] [CrossRef]
- Grzywnowicz, M.; Zaleska, J.; Mertens, D.; Tomczak, W.; Wlasiuk, P.; Kosior, K.; Piechnik, A.; Bojarska-Junak, A.; Dmoszynska, A.; Giannopoulos, K. Programmed Death-1 and Its Ligand Are Novel Immunotolerant Molecules Expressed on Leukemic B Cells in Chronic Lymphocytic Leukemia. PLoS ONE 2012, 7, e35178. [Google Scholar] [CrossRef]
- Brusa, D.; Serra, S.; Coscia, M.; Rossi, D.; D’Arena, G.; Laurenti, L.; Jaksic, O.; Fedele, G.; Inghirami, G.; Gaidano, G.; et al. The PD-1/PD-L1 Axis Contributes to T-Cell Dysfunction in Chronic Lymphocytic Leukemia. Haematologica 2013, 98, 953–963. [Google Scholar] [CrossRef]
- M’Hidi, H.; Thibult, M.L.; Chetaille, B.; Rey, F.; Bouadallah, R.; Nicollas, R.; Olive, D.; Xerri, L. High Expression of the Inhibitory Receptor BTLA in T-Follicular Helper Cells and in B-Cell Small Lymphocytic Lymphoma/Chronic Lymphocytic Leukemia. Am. J. Clin. Pathol. 2009, 132, 589–596. [Google Scholar] [CrossRef]
- Huergo-Zapico, L.; Gonzalez-Rodriguez, A.P.; Contesti, J.; Gonzalez, E.; López-Soto, A.; Fernandez-Guizan, A.; Acebes-Huerta, A.; de los Toyos, J.R.; Lopez-Larrea, C.; Groh, V.; et al. Expression of ERp5 and GRP78 on the Membrane of Chronic Lymphocytic Leukemia Cells: Association with Soluble MICA Shedding. Cancer Immunol. Immunother. 2012, 61, 1201–1210. [Google Scholar] [CrossRef]
- Groh, V.; Wu, J.; Yee, C.; Spies, T. Tumour-Derived Soluble MIC Ligands Impair Expression of NKG2D and T-Cell Activation. Nature 2002, 419, 734–738. [Google Scholar] [CrossRef] [PubMed]
- Wagner, B.; da Silva Nardi, F.; Schramm, S.; Kraemer, T.; Celik, A.A.; Dürig, J.; Horn, P.A.; Dührsen, U.; Nückel, H.; Rebmann, V. HLA-E Allelic Genotype Correlates with HLA-E Plasma Levels and Predicts Early Progression in Chronic Lymphocytic Leukemia. Cancer 2017, 123, 814–823. [Google Scholar] [CrossRef] [PubMed]
- Sebti, Y.; Le Friec, G.; Pangault, C.; Gros, F.; Drénou, B.; Guilloux, V.; Bernard, M.; Lamy, T.; Fauchet, R.; Amiot, L. Soluble HLA-G Molecules Are Increased in Lymphoproliferative Disorders. Hum. Immunol. 2003, 64, 1093–1101. [Google Scholar] [CrossRef] [PubMed]
- Giannopoulos, K.; Schmitt, M.; Kowal, M.; Własiuk, P.; Bojarska-Junak, A.; Roliński, J.; Dmoszyńska, A. The Significance of Soluble HLA-G Plasma Levels as Well as Messenger HLA-G for B-Cell Chronic Lymphocytic Leukemia (B-CLL). Leuk. Res. 2008, 32, 1815–1819. [Google Scholar] [CrossRef] [PubMed]
- Wdowiak, K.; Gallego-Colon, E.; Francuz, T.; Czajka-Francuz, P.; Ruiz-Agamez, N.; Kubeczko, M.; Grochoła, I.; Wybraniec, T.M.; Chudek, J.; Wojnar, J. Increased Serum Levels of Galectin-9 in Patients with Chronic Lymphocytic Leukemia. Oncol. Lett. 2019, 17, 1019–1029. [Google Scholar] [CrossRef]
- Kremer, J.-P.; Reisbach, G.; Nerl, C.; Döurmer, P. B-Cell Chronic Lymphocytic Leukaemia Cells Express and Release Transforming Growth Factor-β. Br. J. Haematol. 1992, 80, 480–487. [Google Scholar] [CrossRef]
- Lotz, M.; Ranheim, E.; Kipps, T.J. Transforming Growth Factor Beta as Endogenous Growth Inhibitor of Chronic Lymphocytic Leukemia B Cells. J. Exp. Med. 1994, 179, 999–1004. [Google Scholar] [CrossRef]
- Castriconi, R.; Cantoni, C.; Della Chiesa, M.; Vitale, M.; Marcenaro, E.; Conte, R.; Biassoni, R.; Bottino, C.; Moretta, L.; Moretta, A. Transforming Growth Factor Beta 1 Inhibits Expression of NKp30 and NKG2D Receptors: Consequences for the NK-Mediated Killing of Dendritic Cells. Proc. Natl. Acad. Sci. USA 2003, 100, 4120–4125. [Google Scholar] [CrossRef]
- Xu, Y.; Sun, J.; Sheard, M.A.; Tran, H.C.; Wan, Z.; Liu, W.Y.; Asgharzadeh, S.; Sposto, R.; Wu, H.W.; Seeger, R.C. Lenalidomide Overcomes Suppression of Human Natural Killer Cell Anti-Tumor Functions by Neuroblastoma Microenvironment-Associated IL-6 and TGFβ1. Cancer Immunol. Immunother. 2013, 62, 1637–1648. [Google Scholar] [CrossRef]
- Zaiatz-Bittencourt, V.; Finlay, D.K.; Gardiner, C.M. Canonical TGF-β Signaling Pathway Represses Human NK Cell Metabolism. J. Immunol. 2018, 200, 3934–3941. [Google Scholar] [CrossRef]
- DiLillo, D.J.; Weinberg, J.B.; Yoshizaki, A.; Horikawa, M.; Bryant, J.M.; Iwata, Y.; Matsushita, T.; Matta, K.M.; Chen, Y.; Venturi, G.M.; et al. Chronic Lymphocytic Leukemia and Regulatory B Cells Share IL-10 Competence and Immunosuppressive Function. Leukemia 2013, 27, 170–182. [Google Scholar] [CrossRef] [PubMed]
- Littwitz-Salomon, E.; Malyshkina, A.; Schimmer, S.; Dittmer, U. The Cytotoxic Activity of Natural Killer Cells Is Suppressed by IL-10+ Regulatory T Cells During Acute Retroviral Infection. Front. Immunol. 2018, 9, 1947. [Google Scholar] [CrossRef] [PubMed]
- Serra, S.; Horenstein, A.L.; Vaisitti, T.; Brusa, D.; Rossi, D.; Laurenti, L.; D’Arena, G.; Coscia, M.; Tripodo, C.; Inghirami, G.; et al. CD73-Generated Extracellular Adenosine in Chronic Lymphocytic Leukemia Creates Local Conditions Counteracting Drug-Induced Cell Death. Blood 2011, 118, 6141–6152. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Matosevic, S. Adenosinergic Signaling as a Target for Natural Killer Cell Immunotherapy. J. Mol. Med. 2018, 96, 903–913. [Google Scholar] [CrossRef]
- Ghiringhelli, F.; Ménard, C.; Terme, M.; Flament, C.; Taieb, J.; Chaput, N.; Puig, P.E.; Novault, S.; Escudier, B.; Vivier, E.; et al. CD4+CD25+ Regulatory T Cells Inhibit Natural Killer Cell Functions in a Transforming Growth Factor–β–Dependent Manner. J. Exp. Med. 2005, 202, 1075–1085. [Google Scholar] [CrossRef]
- Smyth, M.J.; Teng, M.W.L.; Swann, J.; Kyparissoudis, K.; Godfrey, D.I.; Hayakawa, Y. CD4+CD25+ T Regulatory Cells Suppress NK Cell-Mediated Immunotherapy of Cancer. J. Immunol. 2006, 176, 1582–1587. [Google Scholar] [CrossRef]
- Bachanova, V.; Cooley, S.; Defor, T.E.; Verneris, M.R.; Zhang, B.; McKenna, D.H.; Curtsinger, J.; Panoskaltsis-Mortari, A.; Lewis, D.; Hippen, K.; et al. Clearance of Acute Myeloid Leukemia by Haploidentical Natural Killer Cells Is Improved Using IL-2 Diphtheria Toxin Fusion Protein. Blood 2014, 123, 3855–3863. [Google Scholar] [CrossRef]
- Lad, D.; Hoeppli, R.; Huang, Q.; Garcia, R.; Xu, L.; Toze, C.; Broady, R.; Levings, M. Regulatory T-Cells Drive Immune Dysfunction in CLL. Leuk. Lymphoma 2018, 59, 486–489. [Google Scholar] [CrossRef]
- Solman, I.G.; Blum, L.K.; Burger, J.A.; Kipps, T.J.; Dean, J.P.; James, D.F.; Mongan, A. Impact of Long-Term Ibrutinib Treatment on Circulating Immune Cells in Previously Untreated Chronic Lymphocytic Leukemia. Leuk. Res. 2021, 102, 106520. [Google Scholar] [CrossRef]
- Bruno, A.; Mortara, L.; Baci, D.; Noonan, D.M.; Albini, A. Myeloid Derived Suppressor Cells Interactions With Natural Killer Cells and Pro-Angiogenic Activities: Roles in Tumor Progression. Front. Immunol. 2019, 10, 771. [Google Scholar] [CrossRef]
- Kowalska, W.; Bojarska-Junak, A. Monocytic MDSC as a Source of Immunosuppressive Cytokines in Chronic Lymphocytic Leukemia (CLL) Microenvironment. Folia Histochem. Cytobiol. 2020, 58, 25–36. [Google Scholar] [CrossRef] [PubMed]
- Beyer, M.; Kochanek, M.; Darabi, K.; Popov, A.; Jensen, M.; Endl, E.; Knolle, P.A.; Thomas, R.K.; Von Bergwelt-Baildon, M.; Debey, S.; et al. Reduced Frequencies and Suppressive Function of CD4+ CD25 Hi Regulatory T Cells in Patients with Chronic Lymphocytic Leukemia after Therapy with Fludarabine. Blood 2005, 106, 2018–2025. [Google Scholar] [CrossRef] [PubMed]
- Motta, M.; Rassenti, L.; Shelvin, B.J.; Lerner, S.; Kipps, T.J.; Keating, M.J.; Wierda, W.G. Increased Expression of CD152 (CTLA-4) by Normal T Lymphocytes in Untreated Patients with B-Cell Chronic Lymphocytic Leukemia. Leukemia 2005, 19, 1788–1793. [Google Scholar] [CrossRef] [PubMed]
- Jak, M.; Mous, R.; Remmerswaal, E.B.M.; Spijker, R.; Jaspers, A.; Yagüe, A.; Eldering, E.; Van Lier, R.A.W.; Van Oers, M.H.J. Enhanced Formation and Survival of CD4+ CD25hi Foxp3+ T-Cells in Chronic Lymphocytic Leukemia. Leuk. Lymphoma 2009, 50, 788–801. [Google Scholar] [CrossRef]
- Mpakou, V.E.; Ioannidou, H.-D.; Konsta, E.; Vikentiou, M.; Spathis, A.; Kontsioti, F.; Kontos, C.K.; Velentzas, A.D.; Papageorgiou, S.; Vasilatou, D.; et al. Quantitative and Qualitative Analysis of Regulatory T Cells in B Cell Chronic Lymphocytic Leukemia. Leuk. Res. 2017, 60, 74–81. [Google Scholar] [CrossRef]
- Palma, M.; Gentilcore, G.; Heimersson, K.; Mozaffari, F.; Näsman-Glaser, B.; Young, E.; Rosenquist, R.; Hansson, L.; Österborg, A.; Mellstedt, H. T Cells in Chronic Lymphocytic Leukemia Display Dysregulated Expression of Immune Checkpoints and Activation Markers. Haematologica 2017, 102, 562–572. [Google Scholar] [CrossRef]
- Nuñez, S.Y.; Ziblat, A.; Secchiari, F.; Torres, N.I.; Sierra, J.M.; Raffo Iraolagoitia, X.L.; Araya, R.E.; Domaica, C.I.; Fuertes, M.B.; Zwirner, N.W. Human M2 Macrophages Limit NK Cell Effector Functions through Secretion of TGF-β and Engagement of CD85j. J. Immunol. 2018, 200, 1008–1015. [Google Scholar] [CrossRef]
- Spitz, D.L.; Zucker-Franklin, D.; Nabi, Z.F. Unmasking of Cryptic Natural Killer (NK) Cell Recognition Sites on Chronic Lymphocytic Leukemia Lymphocytes. Am. J. Hematol. 1988, 28, 155–161. [Google Scholar] [CrossRef]
- Sánchez-Martínez, D.; Lanuza, P.M.; Gómez, N.; Muntasell, A.; Cisneros, E.; Moraru, M.; Azaceta, G.; Anel, A.; Martínez-Lostao, L.; Villalba, M.; et al. Activated Allogeneic NK Cells Preferentially Kill Poor Prognosis B-Cell Chronic Lymphocytic Leukemia Cells. Front. Immunol. 2016, 7, 454. [Google Scholar] [CrossRef]
- Wurzer, H.; Filali, L.; Hoffmann, C.; Krecke, M.; Biolato, A.M.; Mastio, J.; De Wilde, S.; François, J.H.; Largeot, A.; Berchem, G.; et al. Intrinsic Resistance of Chronic Lymphocytic Leukemia Cells to NK Cell-Mediated Lysis Can Be Overcome In Vitro by Pharmacological Inhibition of Cdc42-Induced Actin Cytoskeleton Remodeling. Front. Immunol. 2021, 12, 1696. [Google Scholar] [CrossRef]
- Wild, J.; Schmiedel, B.J.; Maurer, A.; Raab, S.; Prokop, L.; Stevanović, S.; Dörfel, D.; Schneider, P.; Salih, H.R. Neutralization of (NK-Cell-Derived) B-Cell Activating Factor by Belimumab Restores Sensitivity of Chronic Lymphoid Leukemia Cells to Direct and Rituximab-Induced NK Lysis. Leukemia 2015, 29, 1676–1683. [Google Scholar] [CrossRef] [PubMed]
- Buschle, M.; Campana, D.; Carding, S.R.; Richard, C.; Hoffbrand, A. V Interferon Gamma Inhibits Apopototic Cell Death in B Cell Chronic Lymphocytic Leukemia. J. Exp. Med. 1993, 177, 213–218. [Google Scholar] [CrossRef] [PubMed]
- Buechele, C.; Baessler, T.; Schmiedel, B.J.; Schumacher, C.E.; Grosse-Hovest, L.; Rittig, K.; Salih, H.R. 4-1BB Ligand Modulates Direct and Rituximab-Induced NK-Cell Reactivity in Chronic Lymphocytic Leukemia. Eur. J. Immunol. 2012, 42, 737–748. [Google Scholar] [CrossRef]
- Honigberg, L.A.; Smith, A.M.; Sirisawad, M.; Verner, E.; Loury, D.; Chang, B.; Li, S.; Pan, Z.; Thamm, D.H.; Miller, R.A.; et al. The Bruton Tyrosine Kinase Inhibitor PCI-32765 Blocks B-Cell Activation and Is Efficacious in Models of Autoimmune Disease and B-Cell Malignancy. Proc. Natl. Acad. Sci. USA 2010, 107, 13075–13080. [Google Scholar] [CrossRef]
- Herman, S.E.M.; Gordon, A.L.; Hertlein, E.; Ramanunni, A.; Zhang, X.; Flynn, J.; Jones, J.; Blum, K.a.; Buggy, J.J.; Hamdy, A.; et al. Bruton Tyrosine Kinase Represents a Promising Therapeutic Target for Treatment of Chronic Lymphocytic Leukemia and Is Effectively Targeted by PCI-32765. Blood 2011, 117, 6287–6296. [Google Scholar] [CrossRef] [PubMed]
- Ponader, S.; Chen, S.; Buggy, J.J.; Balakrishnan, K.; Gandhi, V.; Wierda, W.G.; Keating, M.J.; Brien, S.O.; Chiorazzi, N.; Burger, J.A. The Bruton Tyrosine Kinase Inhibitor PCI-32765 Thwarts Chronic Lymphocytic Leukemia Cell Survival and Tissue Homing in Vitro and in Vivo. Blood 2012, 119, 1182–1189. [Google Scholar] [CrossRef]
- De Rooij, M.F.M.; Kuil, A.; Geest, C.R.; Eldering, E.; Chang, B.Y.; Buggy, J.J.; Pals, S.T.; Spaargaren, M. The Clinically Active BTK Inhibitor PCI-32765 Targets B-Cell Receptor– and Chemokine-Controlled Adhesion and Migration in Chronic Lymphocytic Leukemia. Blood 2012, 119, 2590–2594. [Google Scholar] [CrossRef]
- Pleyer, C.; Sun, C.; Desai, S.; Ahn, I.E.; Tian, X.; Nierman, P.; Soto, S.; Superata, J.; Valdez, J.; Lotter, J.; et al. Reconstitution of Humoral Immunity and Decreased Risk of Infections in Patients with Chronic Lymphocytic Leukemia Treated with Bruton Tyrosine Kinase Inhibitors. Leuk. Lymphoma 2020, 61, 2375–2382. [Google Scholar] [CrossRef]
- Rogers, K.A.; Mousa, L.; Zhao, Q.; Bhat, S.A.; Byrd, J.C.; El Boghdadly, Z.; Guerrero, T.; Levine, L.B.; Lucas, F.; Shindiapina, P.; et al. Incidence of Opportunistic Infections during Ibrutinib Treatment for B-Cell Malignancies. Leukemia 2019, 33, 2527–2530. [Google Scholar] [CrossRef]
- Miklos, D.; Cutler, C.S.; Arora, M.; Waller, E.K.; Jagasia, M.; Pusic, I.; Flowers, M.E.; Logan, A.C.; Nakamura, R.; Blazar, B.R.; et al. Ibrutinib for Chronic Graft-versus-Host Disease after Failure of Prior Therapy. Blood 2017, 130, 2243–2250. [Google Scholar] [CrossRef]
- Khurana, D.; Arneson, L.N.; Schoon, R.A.; Dick, C.J.; Leibson, P.J. Differential Regulation of Human NK Cell-Mediated Cytotoxicity by the Tyrosine Kinase Itk. J. Immunol. 2007, 178, 3575–3582. [Google Scholar] [CrossRef]
- Kohrt, H.E.; Sagiv-Barfi, I.; Rafiq, S.; Herman, S.E.M.; Butchar, J.P.; Cheney, C.; Zhang, X.; Buggy, J.J.; Muthusamy, N.; Levy, R.; et al. Ibrutinib Antagonizes Rituximab-Dependent NK Cell–Mediated Cytotoxicity. Blood 2014, 123, 1957–1960. [Google Scholar] [CrossRef] [PubMed]
- Bao, Y.; Zheng, J.; Han, C.; Jin, J.; Han, H.; Liu, Y.; Lau, Y.-L.; Tu, W.; Cao, X. Tyrosine Kinase Btk Is Required for NK Cell Activation. J. Biol. Chem. 2012, 287, 23769–23778. [Google Scholar] [CrossRef] [PubMed]
- Bojarczuk, K.; Siernicka, M.; Dwojak, M.; Bobrowicz, M.; Pyrzynska, B.; Gaj, P.; Karp, M.; Giannopoulos, K.; Efremov, D.G.; Fauriat, C.; et al. B-Cell Receptor Pathway Inhibitors Affect CD20 Levels and Impair Antitumor Activity of Anti-CD20 Monoclonal Antibodies. Leukemia 2014, 28, 1163–1167. [Google Scholar] [CrossRef] [PubMed]
- Da Roit, F.; Engelberts, P.J.; Taylor, R.P.; Breij, E.C.W.; Gritti, G.; Rambaldi, A.; Introna, M.; Parren, P.W.H.I.; Beurskens, F.J.; Golay, J. Ibrutinib Interferes with the Cell-Mediated Anti-Tumor Activities of Therapeutic CD20 Antibodies: Implications for Combination Therapy. Haematologica 2015, 100, 77–86. [Google Scholar] [CrossRef]
- Duong, M.N.; Matera, E.L.; Mathé, D.; Evesque, A.; Valsesia-Wittmann, S.; Clémenceau, B.; Dumontet, C. Effect of Kinase Inhibitors on the Therapeutic Properties of Monoclonal Antibodies. MAbs 2015, 7, 192–198. [Google Scholar] [CrossRef][Green Version]
- Golay, J.; Ubiali, G.; Introna, M. The Specific Bruton Tyrosine Kinase Inhibitor Acalabrutinib (ACP-196) Shows Favorable in Vitro Activity against Chronic Lymphocytic Leukemia B Cells with CD20 Antibodies. Haematologica 2017, 102, e400–e403. [Google Scholar] [CrossRef]
- Hassenrück, F.; Knödgen, E.; Göckeritz, E.; Midda, S.H.; Vondey, V.; Neumann, L.; Herter, S.; Klein, C.; Hallek, M.; Krause, G. Sensitive Detection of the Natural Killer Cell-Mediated Cytotoxicity of Anti-CD20 Antibodies and Its Impairment by B-Cell Receptor Pathway Inhibitors. Biomed Res. Int. 2018, 2018, 1023490. [Google Scholar] [CrossRef]
- Pavlasova, G.; Borsky, M.; Seda, V.; Cerna, K.; Osickova, J.; Doubek, M.; Mayer, J.; Calogero, R.; Trbusek, M.; Pospisilova, S.; et al. Ibrutinib Inhibits CD20 Upregulation on CLL B Cells Mediated by the CXCR4/SDF-1 Axis. Blood 2016, 128, 1609–1613. [Google Scholar] [CrossRef]
- Skarzynski, M.; Niemann, C.U.; Lee, Y.S.; Martyr, S.; Maric, I.; Salem, D.; Stetler-Stevenson, M.; Marti, G.E.; Calvo, K.R.; Yuan, C.; et al. Interactions between Ibrutinib and Anti-CD20 Antibodies: Competing Effects on the Outcome of Combination Therapy. Clin. Cancer Res. 2016, 22, 86–95. [Google Scholar] [CrossRef]
- Patz, M.; Isaeva, P.; Forcob, N.; Müller, B.; Frenzel, L.P.; Wendtner, C.-M.; Klein, C.; Umana, P.; Hallek, M.; Krause, G. Comparison of the in Vitro Effects of the Anti-CD20 Antibodies Rituximab and GA101 on Chronic Lymphocytic Leukaemia Cells. Br. J. Haematol. 2011, 152, 295–306. [Google Scholar] [CrossRef] [PubMed]
- Long, M.; Beckwith, K.; Do, P.; Mundy, B.L.; Gordon, A.; Lehman, A.M.; Maddocks, K.J.; Cheney, C.; Jones, J.A.; Flynn, J.M.; et al. Ibrutinib Treatment Improves T Cell Number and Function in CLL Patients. J. Clin. Investig. 2017, 127, 3052–3064. [Google Scholar] [CrossRef] [PubMed]
- Hofland, T.; de Weerdt, I.; ter Burg, H.; de Boer, R.; Tannheimer, S.; Tonino, S.H.; Kater, A.P.; Eldering, E. Dissection of the Effects of JAK and BTK Inhibitors on the Functionality of Healthy and Malignant Lymphocytes. J. Immunol. 2019, 203, 2100–2109. [Google Scholar] [CrossRef] [PubMed]
- Kondo, K.; Shaim, H.; Thompson, P.A.; Burger, J.A.; Keating, M.; Estrov, Z.; Harris, D.; Kim, E.; Ferrajoli, A.; Daher, M.; et al. Ibrutinib Modulates the Immunosuppressive CLL Microenvironment through STAT3-Mediated Suppression of Regulatory B-Cell Function and Inhibition of the PD-1/PD-L1 Pathway. Leukemia 2018, 32, 960–970. [Google Scholar] [CrossRef]
- Podhorecka, M.; Goracy, A.; Szymczyk, A.; Kowal, M.; Ibanez, B.; Jankowska-Lecka, O.; Macheta, A.; Nowaczynska, A.; Drab-Urbanek, E.; Chocholska, S.; et al. Changes in T-Cell Subpopulations and Cytokine Network during Early Period of Ibrutinib Therapy in Chronic Lymphocytic Leukemia Patients—The Significant Decrease in T Regulatory Cells Number. Oncotarget 2017, 8, 34661. [Google Scholar] [CrossRef] [PubMed]
- Dubovsky, J.A.; Beckwith, K.A.; Natarajan, G.; Woyach, J.A.; Jaglowski, S.; Zhong, Y.; Hessler, J.D.; Liu, T.-M.; Chang, B.Y.; Larkin, K.M.; et al. Ibrutinib Is an Irreversible Molecular Inhibitor of ITK Driving a Th1-Selective Pressure in T Lymphocytes. Blood 2013, 122, 2539–2549. [Google Scholar] [CrossRef]
- Niemann, C.U.; Herman, S.E.M.; Maric, I.; Gomez-Rodriguez, J.; Biancotto, A.; Chang, B.Y.; Martyr, S.; Stetler-Stevenson, M.; Yuan, C.M.; Calvo, K.R.; et al. Disruption of in Vivo Chronic Lymphocytic Leukemia Tumor-Microenvironment Interactions by Ibrutinib—Findings from an Investigator-Initiated Phase II Study. Clin. Cancer Res. 2016, 22, 1572–1582. [Google Scholar] [CrossRef]
- Yin, Q.; Sivina, M.; Robins, H.; Yusko, E.; Vignali, M.; O’Brien, S.; Keating, M.J.; Ferrajoli, A.; Estrov, Z.; Jain, N.; et al. Ibrutinib Therapy Increases T Cell Repertoire Diversity in Patients with Chronic Lymphocytic Leukemia. J. Immunol. 2017, 198, 1740–1747. [Google Scholar] [CrossRef]
- Fiorcari, S.; Maffei, R.; Audrito, V.; Martinelli, S.; Hacken, E.T.; Zucchini, P.; Grisendi, G.; Potenza, L.; Luppi, M.; Burger, J.A.; et al. Ibrutinib Modifies the Function of Monocyte/Macrophage Population in Chronic Lymphocytic Leukemia. Oncotarget 2016, 7, 65968. [Google Scholar] [CrossRef]
- Borge, M.; Almejún, M.B.; Podaza, E.; Colado, A.; Grecco, H.F.; Cabrejo, M.; Bezares, R.F.; Giordano, M.; Gamberale, R. Ibrutinib Impairs the Phagocytosis of Rituximab-Coated Leukemic Cells from Chronic Lymphocytic Leukemia Patients by Human Macrophages. Haematologica 2015, 100, e140–e142. [Google Scholar] [CrossRef]
- Ren, L.; Campbell, A.; Fang, H.; Gautam, S.; Elavazhagan, S.; Fatehchand, K.; Mehta, P.; Stiff, A.; Reader, B.F.; Mo, X.; et al. Analysis of the Effects of the Bruton’s Tyrosine Kinase (Btk) Inhibitor Ibrutinib on Monocyte Fcγ Receptor (FcγR) Function. J. Biol. Chem. 2016, 291, 3043–3052. [Google Scholar] [CrossRef] [PubMed]
- Woyach, J.A.; Ruppert, A.S.; Heerema, N.A.; Zhao, W.; Booth, A.M.; Ding, W.; Bartlett, N.L.; Brander, D.M.; Barr, P.M.; Rogers, K.A.; et al. Ibrutinib Regimens versus Chemoimmunotherapy in Older Patients with Untreated CLL. N. Engl. J. Med. 2018, 379, 2517–2528. [Google Scholar] [CrossRef] [PubMed]
- Burger, J.A.; Sivina, M.; Jain, N.; Kim, E.; Kadia, T.; Estrov, Z.; Nogueras-Gonzalez, G.M.; Huang, X.; Jorgensen, J.; Li, J.; et al. Randomized Trial of Ibrutinib vs. Ibrutinib plus Rituximab in Patients with Chronic Lymphocytic Leukemia. Blood 2019, 133, 1011–1019. [Google Scholar] [CrossRef] [PubMed]
- Byrd, J.C.; Harrington, B.; O’Brien, S.; Jones, J.A.; Schuh, A.; Devereux, S.; Chaves, J.; Wierda, W.G.; Awan, F.T.; Brown, J.R.; et al. Acalabrutinib (ACP-196) in Relapsed Chronic Lymphocytic Leukemia. N. Engl. J. Med. 2016, 374, 323–332. [Google Scholar] [CrossRef]
- Sharman, J.P.; Egyed, M.; Jurczak, W.; Skarbnik, A.; Pagel, J.M.; Flinn, I.W.; Kamdar, M.; Munir, T.; Walewska, R.; Corbett, G.; et al. Acalabrutinib with or without Obinutuzumab versus Chlorambucil and Obinutuzmab for Treatment-Naive Chronic Lymphocytic Leukaemia (ELEVATE TN): A Randomised, Controlled, Phase 3 Trial. Lancet 2020, 395, 1278–1291. [Google Scholar] [CrossRef]
- Kumar, S. Natural Killer Cell Cytotoxicity and Its Regulation by Inhibitory Receptors. Immunology 2018, 154, 383–393. [Google Scholar] [CrossRef]
- Palazzo, A.; Herter, S.; Grosmaire, L.; Jones, R.; Frey, C.R.; Limani, F.; Bacac, M.; Umana, P.; Oldham, R.J.; Marshall, M.J.E.; et al. The PI3Kδ-Selective Inhibitor Idelalisib Minimally Interferes with Immune Effector Function Mediated by Rituximab or Obinutuzumab and Significantly Augments B Cell Depletion In Vivo. J. Immunol. 2018, 200, 2304–2312. [Google Scholar] [CrossRef]
- Akhiani, A.A.; Hallner, A.; Kiffin, R.; Aydin, E.; Werlenius, O.; Aurelius, J.; Martner, A.; Thorén, F.B.; Hellstrand, K. Idelalisib Rescues Natural Killer Cells from Monocyte-Induced Immunosuppression by Inhibiting NOX2-Derived Reactive Oxygen Species. Cancer Immunol. Res. 2020, 8, 1532–1541. [Google Scholar] [CrossRef]
- Rohrbacher, L.; Brauchle, B.; Ogrinc Wagner, A.; von Bergwelt-Baildon, M.; Bücklein, V.L.; Subklewe, M. The PI3K∂-Selective Inhibitor Idelalisib Induces T- and NK-Cell Dysfunction Independently of B-Cell Malignancy-Associated Immunosuppression. Front. Immunol. 2021, 12, 557. [Google Scholar] [CrossRef]
- Brown, J.R. Phosphatidylinositol 3 Kinase δ Inhibitors: Present and Future. Cancer J. 2019, 25, 394–400. [Google Scholar] [CrossRef]
- Dong, S.; Harrington, B.K.; Hu, E.Y.; Greene, J.T.; Lehman, A.M.; Tran, M.; Wasmuth, R.L.; Long, M.; Muthusamy, N.; Brown, J.R.; et al. PI3K P110δ Inactivation Antagonizes Chronic Lymphocytic Leukemia and Reverses T Cell Immune Suppression. J. Clin. Investig. 2019, 129, 122–136. [Google Scholar] [CrossRef] [PubMed]
- Lampson, B.L.; Kasar, S.N.; Matos, T.R.; Morgan, E.A.; Rassenti, L.; Davids, M.S.; Fisher, D.C.; Freedman, A.S.; Jacobson, C.A.; Armand, P.; et al. Idelalisib given Front-Line for Treatment of Chronic Lymphocytic Leukemia Causes Frequent Immune-Mediated Hepatotoxicity. Blood 2016, 128, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Chellappa, S.; Kushekhar, K.; Munthe, L.A.; Tjønnfjord, G.E.; Aandahl, E.M.; Okkenhaug, K.; Taskén, K. The PI3K P110δ Isoform Inhibitor Idelalisib Preferentially Inhibits Human Regulatory T Cell Function. J. Immunol. 2019, 202, 1397–1405. [Google Scholar] [CrossRef]
- Gentile, M.; Petrungaro, A.; Uccello, G.; Vigna, E.; Recchia, A.G.; Caruso, N.; Bossio, S.; De Stefano, L.; Palummo, A.; Storino, F.; et al. Venetoclax for the Treatment of Chronic Lymphocytic Leukemia. Expert Opin. Investig. Drugs 2017, 26, 1307–1316. [Google Scholar] [CrossRef] [PubMed]
- Viant, C.; Guia, S.; Hennessy, R.J.; Rautela, J.; Pham, K.; Bernat, C.; Goh, W.; Jiao, Y.; Delconte, R.; Roger, M.; et al. Cell Cycle Progression Dictates the Requirement for BCL2 in Natural Killer Cell Survival. J. Exp. Med. 2017, 214, 491–510. [Google Scholar] [CrossRef] [PubMed]
- Elías, E.E.; Almejún, M.B.; Colado, A.; Cordini, G.; Vergara-Rubio, M.; Podaza, E.; Risnik, D.; Cabrejo, M.; Fernández-Grecco, H.; Bezares, R.F.; et al. Autologous T-Cell Activation Fosters ABT-199 Resistance in Chronic Lymphocytic Leukemia: Rationale for a Combined Therapy with SYK Inhibitors and Anti-CD20 Monoclonal Antibodies. Haematologica 2018, 103, e458–e461. [Google Scholar] [CrossRef] [PubMed]
- Armant, M.; Delespesse, G.; Sarfati, M. IL-2 and IL-7 but Not IL-12 Protect Natural Killer Cells from Death by Apoptosis and up-Regulate Bcl-2 Expression. Immunology 1995, 85, 331–337. [Google Scholar]
- Jiang, S.; Munker, R.; Andreeff, M. Bcl-2 Is Expressed in Human Natural Killer Cells and Is Regulated by Interleukin-2. Nat. Immunol. 1996, 15, 312–317. [Google Scholar]
- Huntington, N.D.; Puthalakath, H.; Gunn, P.; Naik, E.; Michalak, E.M.; Smyth, M.J.; Tabarias, H.; Degli-Esposti, M.A.; Dewson, G.; Willis, S.N.; et al. Interleukin 15–Mediated Survival of Natural Killer Cells Is Determined by Interactions among Bim, Noxa and Mcl-1. Nat. Immunol. 2007, 8, 856–863. [Google Scholar] [CrossRef]
- Zheng, X.; Wang, Y.; Wei, H.; Ling, B.; Sun, R.; Tian, Z. Bcl-XL Is Associated with the Anti-Apoptotic Effect of IL-15 on the Survival of CD56dim Natural Killer Cells. Mol. Immunol. 2008, 45, 2559–2569. [Google Scholar] [CrossRef]
- Hodge, D.L.; Yang, J.; Buschman, M.D.; Schaughency, P.M.; Dang, H.; Bere, W.; Yang, Y.; Savan, R.; Subleski, J.J.; Yin, X.-M.; et al. Interleukin-15 Enhances Proteasomal Degradation of Bid in Normal Lymphocytes: Implications for Large Granular Lymphocyte Leukemias. Cancer Res. 2009, 69, 3986–3994. [Google Scholar] [CrossRef] [PubMed]
- Sathe, P.; Delconte, R.B.; Souza-Fonseca-Guimaraes, F.; Seillet, C.; Chopin, M.; Vandenberg, C.J.; Rankin, L.C.; Mielke, L.A.; Vikstrom, I.; Kolesnik, T.B.; et al. Innate Immunodeficiency Following Genetic Ablation of Mcl1 in Natural Killer Cells. Nat. Commun. 2014, 5, 4539. [Google Scholar] [CrossRef]
- Capuano, C.; Romanelli, M.; Pighi, C.; Cimino, G.; Rago, A.; Molfetta, R.; Paolini, R.; Santoni, A.; Galandrini, R. Anti-CD20 Therapy Acts via FcγRIIIA to Diminish Responsiveness of Human Natural Killer Cells. Cancer Res. 2015, 75, 4097–4108. [Google Scholar] [CrossRef] [PubMed]
- Teeling, J.L.; French, R.R.; Cragg, M.S.; van den Brakel, J.; Pluyter, M.; Huang, H.; Chan, C.; Parren, P.W.H.I.; Hack, C.E.; Dechant, M.; et al. Characterization of New Human CD20 Monoclonal Antibodies with Potent Cytolytic Activity against Non-Hodgkin Lymphomas. Blood 2004, 104, 1793–1800. [Google Scholar] [CrossRef] [PubMed]
- Werlenius, O.; Aurelius, J.; Hallner, A.; Akhiani, A.A.; Simpanen, M.; Martner, A.; Andersson, P.-O.; Hellstrand, K.; Thorén, F.B. Reactive Oxygen Species Induced by Therapeutic CD20 Antibodies Inhibit Natural Killer Cell-Mediated Antibody-Dependent Cellular Cytotoxicity against Primary CLL Cells. Oncotarget 2016, 7, 32046. [Google Scholar] [CrossRef]
- Werlenius, O.; Riise, R.E.; Simpanen, M.; Aurelius, J.; Thorén, F.B. CD20 Antibodies Induce Production and Release of Reactive Oxygen Species by Neutrophils. Blood 2014, 123, 4001–4002. [Google Scholar] [CrossRef]
- Mössner, E.; Brünker, P.; Moser, S.; Püntener, U.; Schmidt, C.; Herter, S.; Grau, R.; Gerdes, C.; Nopora, A.; van Puijenbroek, E.; et al. Increasing the Efficacy of CD20 Antibody Therapy through the Engineering of a New Type II Anti-CD20 Antibody with Enhanced Direct and Immune Effector Cell–Mediated B-Cell Cytotoxicity. Blood 2010, 115, 4393–4402. [Google Scholar] [CrossRef]
- Terszowski, G.; Klein, C.; Stern, M. KIR/HLA Interactions Negatively Affect Rituximab- but Not GA101 (Obinutuzumab)-Induced Antibody-Dependent Cellular Cytotoxicity. J. Immunol. 2014, 192, 5618–5624. [Google Scholar] [CrossRef]
- Capuano, C.; Pighi, C.; Molfetta, R.; Paolini, R.; Battella, S.; Palmieri, G.; Giannini, G.; Belardinilli, F.; Santoni, A.; Galandrini, R. Obinutuzumab-Mediated High-Affinity Ligation of FcγRIIIA/CD16 Primes NK Cells for IFNγ Production. Oncoimmunology 2017, 6, e1290037. [Google Scholar] [CrossRef]
- Capuano, C.; Pighi, C.; Maggio, R.; Battella, S.; Morrone, S.; Palmieri, G.; Santoni, A.; Klein, C.; Galandrini, R. CD16 Pre-Ligation by Defucosylated Tumor-Targeting MAb Sensitizes Human NK Cells to Γc Cytokine Stimulation via PI3K/MTOR Axis. Cancer Immunol. Immunother. 2020, 69, 501–512. [Google Scholar] [CrossRef]
- Capuano, C.; Battella, S.; Pighi, C.; Franchitti, L.; Turriziani, O.; Morrone, S.; Santoni, A.; Galandrini, R.; Palmieri, G. Tumor-Targeting Anti-CD20 Antibodies Mediate In Vitro Expansion of Memory Natural Killer Cells: Impact of CD16 Affinity Ligation Conditions and In Vivo Priming. Front. Immunol. 2018, 9, 1031. [Google Scholar] [CrossRef] [PubMed]
- Freeman, C.L.; Morschhauser, F.; Sehn, L.; Dixon, M.; Houghton, R.; Lamy, T.; Fingerle-Rowson, G.; Wassner-Fritsch, E.; Gribben, J.G.; Hallek, M.; et al. Cytokine Release in Patients with CLL Treated with Obinutuzumab and Possible Relationship with Infusion-Related Reactions. Blood 2015, 126, 2646–2649. [Google Scholar] [CrossRef] [PubMed]
- García-Muñoz, R.; Aguinaga, L.; Feliu, J.; Anton-Remirez, J.; Jorge-Del-Val, L.; Casajús-Navasal, A.; Nebot-Villacampa, M.J.; Daroca-Fernandez, I.; Domínguez-Garrido, E.; Rabasa, P.; et al. Obinutuzumab Induces Depletion of NK Cells in Patients with Chronic Lymphocytic Leukemia. Immunotherapy 2018, 10, 491–499. [Google Scholar] [CrossRef] [PubMed]
- García-Muñoz, R.; Nájera, M.J.; Feliu, J.; Antón-Remírez, J.; Ramalle-Gómara, E.; Marín-Gorricho, R.; Peralta, R.; Gutiérrez-Gamarra, E.; Nuñez-Rodriguez, J.; Zafra-Morales, R.; et al. Battle of Thermopylae: 300 Spartans (Natural Killer Cells plus Obinutuzumab) versus the Immortal Warriors (Chronic Lymphocytic Leukemia Cells) of Xerxes’ Army. Future Sci. 2019, 5, FSO425. [Google Scholar] [CrossRef]
- Enqvist, M.; Jacobs, B.; Junlén, H.R.; Schaffer, M.; Melén, C.M.; Friberg, D.; Wahlin, B.E.; Malmberg, K.-J. Systemic and Intra-Nodal Activation of NK Cells After Rituximab Monotherapy for Follicular Lymphoma. Front. Immunol. 2019, 10, 2085. [Google Scholar] [CrossRef]
- Lepretre, S.; Aurran, T.; Mahé, B.; Cazin, B.; Tournilhac, O.; Maisonneuve, H.; Casasnovas, O.; Delmer, A.; Leblond, V.; Royer, B.; et al. Excess Mortality after Treatment with Fludarabine and Cyclophosphamide in Combination with Alemtuzumab in Previously Untreated Patients with Chronic Lymphocytic Leukemia in a Randomized Phase 3 Trial. Blood 2012, 119, 5104–5110. [Google Scholar] [CrossRef]
- Ysebaert, L.; Gross, E.; Kühlein, E.; Blanc, A.; Corre, J.; Fournié, J.J.; Laurent, G.; Quillet-Mary, A. Immune Recovery after Fludarabine–Cyclophosphamide–Rituximab Treatment in B-Chronic Lymphocytic Leukemia: Implication for Maintenance Immunotherapy. Leukemia 2010, 24, 1310–1316. [Google Scholar] [CrossRef] [PubMed]
- García Muñoz, R.; Izquierdo-Gil, A.; Muñoz, A.; Roldan-Galiacho, V.; Rabasa, P.; Panizo, C. Lymphocyte Recovery Is Impaired in Patients with Chronic Lymphocytic Leukemia and Indolent Non-Hodgkin Lymphomas Treated with Bendamustine plus Rituximab. Ann. Hematol. 2014, 93, 1879–1887. [Google Scholar] [CrossRef]
- Martínez-Calle, N.; Hartley, S.; Ahearne, M.; Kasenda, B.; Beech, A.; Knight, H.; Balotis, C.; Kennedy, B.; Wagner, S.; Dyer, M.J.S.; et al. Kinetics of T-Cell Subset Reconstitution Following Treatment with Bendamustine and Rituximab for Low-Grade Lymphoproliferative Disease: A Population-Based Analysis. Br. J. Haematol. 2019, 184, 957–968. [Google Scholar] [CrossRef]
- Seymour, J.F.; Kipps, T.J.; Eichhorst, B.; Hillmen, P.; D’Rozario, J.; Assouline, S.; Owen, C.; Gerecitano, J.; Robak, T.; De la Serna, J.; et al. Venetoclax–Rituximab in Relapsed or Refractory Chronic Lymphocytic Leukemia. N. Engl. J. Med. 2018, 378, 1107–1120. [Google Scholar] [CrossRef]
- Itchaki, G.; Brown, J.R. Lenalidomide in the Treatment of Chronic Lymphocytic Leukemia. Expert Opin. Investig. Drugs 2017, 26, 633–650. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, T.; Hideshima, T.; Akiyama, M.; Podar, K.; Yasui, H.; Raje, N.; Kumar, S.; Chauhan, D.; Treon, S.P.; Richardson, P.; et al. Molecular Mechanisms Whereby Immunomodulatory Drugs Activate Natural Killer Cells: Clinical Application. Br. J. Haematol. 2005, 128, 192–203. [Google Scholar] [CrossRef] [PubMed]
- Reddy, N.; Hernandez-Ilizaliturri, F.J.; Deeb, G.; Roth, M.; Vaughn, M.; Knight, J.; Wallace, P.; Czuczman, M.S. Immunomodulatory Drugs Stimulate Natural Killer-Cell Function, Alter Cytokine Production by Dendritic Cells, and Inhibit Angiogenesis Enhancing the Anti-Tumour Activity of Rituximab in Vivo. Br. J. Haematol. 2008, 140, 36–45. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Adams, M.; Carter, T.; Chen, R.; Muller, G.; Stirling, D.; Schafer, P.; Bartlett, J.B. Lenalidomide Enhances Natural Killer Cell and Monocyte-Mediated Antibody-Dependent Cellular Cytotoxicity of Rituximab-Treated CD20+ Tumor Cells. Clin. Cancer Res. 2008, 14, 4650–4657. [Google Scholar] [CrossRef] [PubMed]
- Acebes-Huerta, A.; Huergo-Zapico, L.; Gonzalez-Rodriguez, A.P.; Fernandez-Guizan, A.; Payer, A.R.; López-Soto, A.; Gonzalez, S. Lenalidomide Induces Immunomodulation in Chronic Lymphocytic Leukemia and Enhances Antitumor Immune Responses Mediated by NK and CD4 T Cells. Biomed Res. Int. 2014, 2014, 265840. [Google Scholar] [CrossRef]
- Lagrue, K.; Carisey, A.; Morgan, D.J.; Chopra, R.; Davis, D.M. Lenalidomide Augments Actin Remodeling and Lowers NK-Cell Activation Thresholds. Blood 2015, 126, 50–60. [Google Scholar] [CrossRef]
- Vitale, C.; Strati, P. CAR T-Cell Therapy for B-Cell Non-Hodgkin Lymphoma and Chronic Lymphocytic Leukemia: Clinical Trials and Real-World Experiences. Front. Oncol. 2020, 10, 849. [Google Scholar] [CrossRef]
- Xu, X.; Sun, Q.; Liang, X.; Chen, Z.; Zhang, X.; Zhou, X.; Li, M.; Tu, H.; Liu, Y.; Tu, S.; et al. Mechanisms of Relapse After CD19 CAR T-Cell Therapy for Acute Lymphoblastic Leukemia and Its Prevention and Treatment Strategies. Front. Immunol. 2019, 10, 2664. [Google Scholar] [CrossRef]
- Sabry, M.; Lowdell, M.W. Killers at the Crossroads: The Use of Innate Immune Cells in Adoptive Cellular Therapy of Cancer. Stem Cells Transl. Med. 2020, 9, 974–984. [Google Scholar] [CrossRef]
- Hu, W.; Wang, G.; Huang, D.; Sui, M.; Xu, Y. Cancer Immunotherapy Based on Natural Killer Cells: Current Progress and New Opportunities. Front. Immunol. 2019, 10, 1205. [Google Scholar] [CrossRef]
- Liu, E.; Marin, D.; Banerjee, P.; Macapinlac, H.A.; Thompson, P.; Basar, R.; Nassif Kerbauy, L.; Overman, B.; Thall, P.; Kaplan, M.; et al. Use of CAR-Transduced Natural Killer Cells in CD19-Positive Lymphoid Tumors. N. Engl. J. Med. 2020, 382, 545–553. [Google Scholar] [CrossRef] [PubMed]
- Buitrago, J.; Adkins, S.; Hawkins, M.; Iyamu, K.; Oort, T. Adult Survivorship: Considerations Following CAR T-Cell Therapy. Clin. J. Oncol. Nurs. 2019, 23, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Van der Harst, D.; Brand, A.; van Luxemburg-Heys, S.; Kooy-Winkelaar, E.; van Rood, J. Lymphokine-Activated Killer Cell Functions in Patients with Leukemic B-Lymphoproliferative Diseases. Blood 1989, 74, 2464–2470. [Google Scholar] [CrossRef]
- Foa, B.R.; Fierro, M.T.; Raspadori, D.; Bonferroni, M.; Cardona, S.; Guarini, A.; Gillio Tos, A.; Francia di Celle, P.; Cesano, A.; Matera, L.; et al. Lymphokine-Activated Killer (LAK) Cell Activity in B and T Chronic Lymphoid Leukemia: Defective LAK Generation and Reduced Susceptibility of the Leukemic Cells to Allogeneic and Autologous LAK Effectors. Blood 1990, 76, 1349–1354. [Google Scholar] [CrossRef] [PubMed]
- Santiago-Schwarz, F.; Panagiotopoulos, C.; Sawitsky, A.; Rai, K.R. Distinct Characteristics of Lymphokine-Activated Killer (LAK) Cells Derived from Patients with B-Cell Chronic Lymphocytic Leukemia (B-CLL). A Factor in B-CLL Serum Promotes Natural Killer Cell-like LAK Cell Growth. Blood 1990, 76, 1355–1360. [Google Scholar] [CrossRef]
- Guven, H.; Gilljam, M.; Chambers, B.J.; Ljunggren, H.G.; Christensson, B.; Kimby, E.; Dilber, M.S. Expansion of Natural Killer (NK) and Natural Killer-like T (NKT)-Cell Populations Derived from Patients with B-Chronic Lymphocytic Leukemia (B-CLL): A Potential Source for Cellular Immunotherapy. Leukemia 2003, 17, 1973–1980. [Google Scholar] [CrossRef]
- Laprevotte, E.; Voisin, G.; Ysebaert, L.; Klein, C.; Daugrois, C.; Laurent, G.; Fournie, J.-J.; Quillet-Mary, A. Recombinant Human IL-15 Trans-Presentation by B Leukemic Cells from Chronic Lymphocytic Leukemia Induces Autologous NK Cell Proliferation Leading to Improved Anti-CD20 Immunotherapy. J. Immunol. 2013, 191, 3634–3640. [Google Scholar] [CrossRef]
- Sánchez-Martínez, D.; Azaceta, G.; Muntasell, A.; Aguiló, N.; Núñez, D.; Gálvez, E.M.; Naval, J.; Anel, A.; Palomera, L.; Vilches, C.; et al. Human NK Cells Activated by EBV(+) Lymphoblastoid Cells Overcome Anti-Apoptotic Mechanisms of Drug Resistance in Haematological Cancer Cells. Oncoimmunology 2015, 4, e991613. [Google Scholar] [CrossRef]
- Sanchez-Martinez, D.; Allende-Vega, N.; Orecchioni, S.; Talarico, G.; Cornillon, A.; Vo, D.-N.; Rene, C.; Lu, Z.-Y.; Krzywinska, E.; Anel, A.; et al. Expansion of Allogeneic NK Cells with Efficient Antibody-Dependent Cell Cytotoxicity against Multiple Tumors. Theranostics 2018, 8, 3856–3869. [Google Scholar] [CrossRef]
- Calvo, T.; Reina-Ortiz, C.; Giraldos, D.; Gascón, M.; Woods, D.; Asenjo, J.; Marco-Brualla, J.; Azaceta, G.; Izquierdo, I.; Palomera, L.; et al. Expanded and Activated Allogeneic NK Cells Are Cytotoxic against B-Chronic Lymphocytic Leukemia (B-CLL) Cells with Sporadic Cases of Resistance. Sci. Rep. 2020, 10, 19398. [Google Scholar] [CrossRef]
- Müller, T.; Uherek, C.; Maki, G.; Chow, K.U.; Schimpf, A.; Klingemann, H.-G.; Tonn, T.; Wels, W.S. Expression of a CD20-Specific Chimeric Antigen Receptor Enhances Cytotoxic Activity of NK Cells and Overcomes NK-Resistance of Lymphoma and Leukemia Cells. Cancer Immunol. Immunother. 2008, 57, 411–423. [Google Scholar] [CrossRef]
- Boissel, L.; Betancur, M.; Wels, W.S.; Tuncer, H.; Klingemann, H. Transfection with MRNA for CD19 Specific Chimeric Antigen Receptor Restores NK Cell Mediated Killing of CLL Cells. Leuk. Res. 2009, 33, 1255–1259. [Google Scholar] [CrossRef] [PubMed]
- Liu, E.; Tong, Y.; Dotti, G.; Shaim, H.; Savoldo, B.; Mukherjee, M.; Orange, J.; Wan, X.; Lu, X.; Reynolds, A.; et al. Cord Blood NK Cells Engineered to Express IL-15 and a CD19-Targeted CAR Show Long-Term Persistence and Potent Antitumor Activity. Leukemia 2018, 32, 520–531. [Google Scholar] [CrossRef] [PubMed]
- Cichocki, F.; Goodridge, J.; Bjordahl, R.; Mahmood, S.; Davis, Z.B.; Gaidarova, S.; Abujarour, R.; Groff, B.; Witty, A.D.; Wang, H.; et al. Dual-Antigen Targeted off-the-Shelf NK Cells Show Durable Response and Prevent Antigen Escape in Lymphoma and Leukemia. Blood 2022. [Google Scholar] [CrossRef] [PubMed]
- Yano, M.; Sharpe, C.; Lance, J.R.; Ravikrishnan, J.; Zapolnik, K.; Mo, X.M.; Woyach, J.A.; Sampath, D.; Kittai, A.S.; Vasu, S.; et al. Evaluation of Allogeneic and Autologous Membrane-Bound IL-21-Expanded NK Cells for Chronic Lymphocytic Leukemia Therapy. Blood Adv. 2022, 6, 5641–5654. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yano, M.; Byrd, J.C.; Muthusamy, N. Natural Killer Cells in Chronic Lymphocytic Leukemia: Functional Impairment and Therapeutic Potential. Cancers 2022, 14, 5787. https://doi.org/10.3390/cancers14235787
Yano M, Byrd JC, Muthusamy N. Natural Killer Cells in Chronic Lymphocytic Leukemia: Functional Impairment and Therapeutic Potential. Cancers. 2022; 14(23):5787. https://doi.org/10.3390/cancers14235787
Chicago/Turabian StyleYano, Max, John C. Byrd, and Natarajan Muthusamy. 2022. "Natural Killer Cells in Chronic Lymphocytic Leukemia: Functional Impairment and Therapeutic Potential" Cancers 14, no. 23: 5787. https://doi.org/10.3390/cancers14235787
APA StyleYano, M., Byrd, J. C., & Muthusamy, N. (2022). Natural Killer Cells in Chronic Lymphocytic Leukemia: Functional Impairment and Therapeutic Potential. Cancers, 14(23), 5787. https://doi.org/10.3390/cancers14235787