

Insights on the Role of PGRMC1 in Mitotic and Meiotic Cell Division
 , , and
, , and
Abstract
Simple Summary
Abstract
1. Introduction
2. Cell Division
2.1. The Coordinated Role of the Cytoskeleton
2.2. The Spindle Assembly Checkpoint (SAC) and the Chromosomal Passenger Complex (CPC)
2.3. The Role of the Membranous Compartment
3. PGRMC1 and Cell Division
3.1. Experimental Evidence That PGRMC1 Participates in the Control of Cell Proliferation and Cell Division
3.1.1. Control of Mitotic Cell Division
3.1.2. Control of Meiotic Cell Division
3.1.3. Putative PGRMC1 Mechanisms of Action Controlling the Progression of Mitotic and Meiotic Cell Division
4. Other PGRMC1 Functions Found in Interphasic Cells Providing Insights into How PGRMC1 Might Participate in the Progression of Cell Division
4.1. Interaction with the Actin Cytoskeleton and Function Mediating Cell Shape and Migration

4.2. Control of Membrane Trafficking
5. Conclusions and Future Directions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cahill, M.A.; Neubauer, H. PGRMC Proteins Are Coming of Age: A Special Issue on the Role of PGRMC1 and PGRMC2 in Metabolism and Cancer Biology. Cancers 2021, 13, 512. [Google Scholar] [CrossRef] [PubMed]
- Cahill, M.A. Progesterone Receptor Membrane Component 1: An Integrative Review. J. Steroid Biochem. Mol. Biol. 2007, 105, 13–36. [Google Scholar] [CrossRef] [PubMed]
- Pru, J.K. Pleiotropic Actions of Pgrmc Proteins in Cancer. Endocrinology 2022, 163, 512. [Google Scholar] [CrossRef] [PubMed]
- Peluso, J.J. Progesterone Signaling and Mammalian Ovarian Follicle Growth Mediated by Progesterone Receptor Membrane Component Family Members. Cells 2022, 11, 1632. [Google Scholar] [CrossRef] [PubMed]
- Peluso, J.J.; Pru, J.K. Progesterone Receptor Membrane Component (PGRMC)1 and PGRMC2 and Their Roles in Ovarian and Endometrial Cancer. Cancers 2021, 13, 5953. [Google Scholar] [CrossRef] [PubMed]
- Markov, A.V.; Kaznacheev, I.S. Evolutionary consequences of polyploidy in prokaryotes and the origin of mitosis and meiosis. Biol. Direct. 2016, 11, 28. [Google Scholar] [CrossRef] [PubMed]
- Scholey, J.M.; Brust-Mascher, I.; Mogilner, A. Cell division. Nature 2003, 422, 746–752. [Google Scholar] [CrossRef] [PubMed]
- Wittmann, T.; Hyman, A.; Desai, A. The spindle: A dynamic assembly of microtubules and motors. Nat. Cell Biol. 2001, 3, E28–E34. [Google Scholar] [CrossRef] [PubMed]
- Vogt, E.; Kirsch-Volders, M.; Parry, J.; Eichenlaub-Ritter, U. Spindle formation, chromosome segregation and the spindle checkpoint in mammalian oocytes and susceptibility to meiotic error. Mutat. Res. 2008, 651, 14–29. [Google Scholar] [CrossRef] [PubMed]
- Nunes, V.; Ferreira, J.G. From the cytoskeleton to the nucleus: An integrated view on early spindle assembly. In Seminars in Cell & Developmental Biology; Academic Press: Cambridge, MA, USA, 2021; Volume 117, pp. 42–51. [Google Scholar]
- Davidson, P.M.; Cadot, B. Actin on and around the Nucleus. Trends Cell Biol. 2021, 31, 211–223. [Google Scholar] [CrossRef] [PubMed]
- Maddox, A.S.; Azoury, J.; Dumont, J. Polar body cytokinesis. Cytoskeleton 2012, 69, 855–868. [Google Scholar] [CrossRef]
- Normand, G.; King, R.W. Understanding cytokinesis failure. Adv. Exp. Med. Biol. 2010, 676, 27–55. [Google Scholar] [PubMed]
- Sagona, A.P.; Stenmark, H. Cytokinesis and cancer. FEBS Lett. 2010, 584, 2652–2661. [Google Scholar] [CrossRef] [PubMed]
- Straight, A.F.; Cheung, A.; Limouze, J.; Chen, I.; Westwood, N.J.; Sellers, J.R.; Mitchison, T.J. Dissecting temporal and spatial control of cytokinesis with a myosin II Inhibitor. Science 2003, 299, 1743–1747. [Google Scholar] [CrossRef] [PubMed]
- Musacchio, A.; Salmon, E.D. The spindle-assembly checkpoint in space and time. Nat. Rev. Mol. Cell Biol. 2007, 8, 379–393. [Google Scholar] [CrossRef]
- Jones, K.T.; Lane, S.I. Molecular causes of aneuploidy in mammalian eggs. Development 2013, 140, 3719–3730. [Google Scholar] [CrossRef]
- Akhshi, T.K.; Wernike, D.; Piekny, A. Microtubules and actin crosstalk in cell migration and division. Cytoskeleton 2014, 71, 1–23. [Google Scholar] [CrossRef]
- Ajduk, A.; Zernicka-Goetz, M. Polarity and cell division orientation in the cleavage embryo: From worm to human. Mol. Hum. Reprod. 2016, 22, 691–703. [Google Scholar] [CrossRef]
- Evans, J.P.; Robinson, D.N. The spatial and mechanical challenges of female meiosis. Mol. Reprod. Dev. 2011, 78, 769–777. [Google Scholar] [CrossRef]
- Edwards, R.G.; Beard, H.K. Oocyte polarity and cell determination in early mammalian embryos. Mol. Hum. Reprod. 1997, 3, 863–905. [Google Scholar] [CrossRef]
- Leung, C.Y.; Zhu, M.; Zernicka-Goetz, M. Polarity in Cell-Fate Acquisition in the Early Mouse Embryo. Curr. Top. Dev. Biol. 2016, 120, 203–234. [Google Scholar] [PubMed]
- Maitre, J.L.; Turlier, H.; Illukkumbura, R.; Eismann, B.; Niwayama, R.; Nedelec, F.; Hiiragi, T. Asymmetric division of contractile domains couples cell positioning and fate specification. Nature 2016, 536, 344–348. [Google Scholar] [CrossRef] [PubMed]
- Morin, X.; Bellaiche, Y. Mitotic spindle orientation in asymmetric and symmetric cell divisions during animal development. Dev. Cell 2011, 21, 102–119. [Google Scholar] [CrossRef]
- Noatynska, A.; Gotta, M. Cell polarity and asymmetric cell division: The C. elegans early embryo. Essays Biochem. 2012, 53, 1–14. [Google Scholar] [PubMed]
- Hyttel, P. Gametogenesis. In Essentials of Domestic Animal Embriology; Hyttel, P., Sinowatz, F., Vejlsted, M., Eds.; Saunders Elsevier: Amsterdam, The Netherlands, 2010; pp. 32–56. [Google Scholar]
- Sato, M.; Kakui, Y.; Toya, M. Tell the Difference Between Mitosis and Meiosis: Interplay Between Chromosomes, Cytoskeleton, and Cell Cycle Regulation. Front. Cell Dev. Biol. 2021, 9, 660322. [Google Scholar] [CrossRef]
- Hohmann, T.; Dehghani, F. The Cytoskeleton-A Complex Interacting Meshwork. Cells 2019, 8, 362. [Google Scholar] [CrossRef]
- Burute, M.; Kapitein, L.C. Cellular Logistics: Unraveling the Interplay Between Microtubule Organization and Intracellular Transport. Annu. Rev. Cell Dev. Biol. 2019, 35, 29–54. [Google Scholar] [CrossRef]
- Kitagawa, M.; Lee, S.H. The chromosomal passenger complex (CPC) as a key orchestrator of orderly mitotic exit and cytokinesis. Front. Cell Dev. Biol. 2015, 3, 14. [Google Scholar] [CrossRef]
- Mierzwa, B.; Gerlich, D.W. Cytokinetic abscission: Molecular mechanisms and temporal control. Dev. Cell 2014, 31, 525–538. [Google Scholar] [CrossRef]
- Green, R.A.; Paluch, E.; Oegema, K. Cytokinesis in animal cells. Annu. Rev. Cell Dev. Biol. 2012, 28, 29–58. [Google Scholar] [CrossRef]
- Ruchaud, S.; Carmena, M.; Earnshaw, W.C. Chromosomal passengers: Conducting cell division. Nat. Rev. Mol. Cell Biol. 2007, 8, 798–812. [Google Scholar] [CrossRef] [PubMed]
- Champion, L.; Linder, M.I.; Kutay, U. Cellular Reorganization during Mitotic Entry. Trends Cell Biol. 2017, 27, 26–41. [Google Scholar] [CrossRef] [PubMed]
- Dao, V.T.; Dupuy, A.G.; Gavet, O.; Caron, E.; de Gunzburg, J. Dynamic changes in Rap1 activity are required for cell retraction and spreading during mitosis. J. Cell Sci. 2009, 122, 2996–3004. [Google Scholar] [CrossRef] [PubMed]
- Dix, C.L.; Matthews, H.K.; Uroz, M.; McLaren, S.; Wolf, L.; Heatley, N.; Win, Z.; Almada, P.; Henriques, R.; Boutros, M.; et al. The Role of Mitotic Cell-Substrate Adhesion Re-modeling in Animal Cell Division. Dev. Cell 2018, 45, 132–145.e133. [Google Scholar] [CrossRef] [PubMed]
- Cramer, L.P.; Mitchison, T.J. Investigation of the mechanism of retraction of the cell margin and rearward flow of nodules during mitotic cell rounding. Mol. Biol. Cell 1997, 8, 109–119. [Google Scholar] [CrossRef][Green Version]
- Whitehead, C.M.; Winkfein, R.J.; Rattner, J.B. The relationship of HsEg5 and the actin cytoskeleton to centrosome separation. Cell Motil Cytoskelet. 1996, 35, 298–308. [Google Scholar] [CrossRef]
- Raaijmakers, J.A.; van Heesbeen, R.G.; Meaders, J.L.; Geers, E.F.; Fernandez-Garcia, B.; Medema, R.H.; Tanenbaum, M.E. Nuclear envelope-associated dynein drives prophase centrosome separation and enables Eg5-independent bipolar spindle formation. EMBO J. 2012, 31, 4179–4190. [Google Scholar] [CrossRef]
- De Simone, A.; Nedelec, F.; Gonczy, P. Dynein Transmits Polarized Actomyosin Cortical Flows to Promote Centrosome Separation. Cell Rep. 2016, 14, 2250–2262. [Google Scholar] [CrossRef]
- Uzbekov, R.; Kireyev, I.; Prigent, C. Centrosome separation: Respective role of microtubules and actin filaments. Biol. Cell 2002, 94, 275–288. [Google Scholar] [CrossRef]
- Chugh, P.; Paluch, E.K. The actin cortex at a glance. J. Cell Sci. 2018, 131, 186254. [Google Scholar] [CrossRef]
- Ramkumar, N.; Baum, B. Coupling changes in cell shape to chromosome segregation. Nat. Rev. Mol. Cell Biol. 2016, 17, 511–521. [Google Scholar] [CrossRef] [PubMed]
- Ungricht, R.; Kutay, U. Mechanisms and functions of nuclear envelope remodelling. Nat. Rev. Mol. Cell Biol. 2017, 18, 229–245. [Google Scholar] [CrossRef] [PubMed]
- Gavet, O.; Pines, J. Progressive activation of CyclinB1-Cdk1 coordinates entry to mitosis. Dev. Cell 2010, 18, 533–543. [Google Scholar] [CrossRef] [PubMed]
- Conduit, P.T.; Wainman, A.; Raff, J.W. Centrosome function and assembly in animal cells. Nat. Rev. Mol. Cell Biol. 2015, 16, 611–624. [Google Scholar] [CrossRef]
- Meraldi, P. Centrosomes in spindle organization and chromosome segregation: A mechanistic view. Chromosome Res. 2016, 24, 19–34. [Google Scholar] [CrossRef]
- Maiato, H.; Logarinho, E. Mitotic spindle multipolarity without centrosome amplification. Nat. Cell Biol. 2014, 16, 386–394. [Google Scholar] [CrossRef]
- Karsenti, E.; Vernos, I. The mitotic spindle: A self-made machine. Science 2001, 294, 543–547. [Google Scholar] [CrossRef]
- Schuh, M.; Ellenberg, J. Self-organization of MTOCs replaces centrosome function during acentrosomal spindle assembly in live mouse oocytes. Cell 2007, 130, 484–498. [Google Scholar] [CrossRef]
- Dumont, J.; Desai, A. Acentrosomal spindle assembly and chromosome segregation during oocyte meiosis. Trends Cell Biol. 2012, 22, 241–249. [Google Scholar] [CrossRef]
- So, C.; Menelaou, K.; Uraji, J.; Harasimov, K.; Steyer, A.M.; Seres, K.B.; Bucevicius, J.; Lukinavicius, G.; Mobius, W.; Sibold, C.; et al. Mechanism of spindle pole organization and instability in human oocytes. Science 2022, 375, eabj3944. [Google Scholar] [CrossRef]
- Holubcova, Z.; Blayney, M.; Elder, K.; Schuh, M. Human oocytes. Error-prone chromosome-mediated spindle assembly favors chromosome segregation defects in human oocytes. Science 2015, 348, 1143–1147. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.H.; Lee, I.W.; Jo, Y.J.; Kim, N.H.; Namgoong, S. Acentriolar microtubule organization centers and Ran-mediated microtubule formation pathways are both required in porcine oocytes. Mol. Reprod. Dev. 2019, 86, 972–983. [Google Scholar] [CrossRef] [PubMed]
- Kamijo, K.; Ohara, N.; Abe, M.; Uchimura, T.; Hosoya, H.; Lee, J.S.; Miki, T. Dissecting the role of Rho-mediated signaling in contractile ring formation. Mol. Biol. Cell 2006, 17, 43–55. [Google Scholar] [CrossRef] [PubMed]
- Duro, J.; Nilsson, J. SAC during early cell divisions: Sacrificing fidelity over timely division, regulated differently across organisms: Chromosome alignment and segregation are left unsupervised from the onset of development until checkpoint activity is acquired, varying from species to species. Bioessays 2021, 43, e2000174. [Google Scholar] [PubMed]
- Sun, S.C.; Kim, N.H. Spindle assembly checkpoint and its regulators in meiosis. Hum. Reprod. Update 2012, 18, 60–72. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Bian, M.; Jiang, Q.; Zhang, C. Roles of Aurora kinases in mitosis and tumorigenesis. Mol. Cancer Res. 2007, 5, 1–10. [Google Scholar] [CrossRef]
- Sharif, B.; Na, J.; Lykke-Hartmann, K.; McLaughlin, S.H.; Laue, E.; Glover, D.M.; Zernicka-Goetz, M. The chromosome passenger complex is required for fidelity of chromosome transmission and cytokinesis in meiosis of mouse oocytes. J. Cell Sci. 2010, 123, 4292–4300. [Google Scholar] [CrossRef]
- Yang, K.T.; Li, S.K.; Chang, C.C.; Tang, C.J.; Lin, Y.N.; Lee, S.C.; Tang, T.K. Aurora-C kinase deficiency causes cytokinesis failure in meiosis I and production of large polyploid oocytes in mice. Mol. Biol. Cell 2010, 21, 2371–2383. [Google Scholar] [CrossRef]
- Afonso, O.; Figueiredo, A.C.; Maiato, H. Late mitotic functions of Aurora kinases. Chromosoma 2016, 126, 93–103. [Google Scholar] [CrossRef]
- Smith, C.M.; Chircop, M. Clathrin-mediated endocytic proteins are involved in regulating mitotic progression and completion. Traffic 2012, 13, 1628–1641. [Google Scholar] [CrossRef]
- Holubcova, Z.; Howard, G.; Schuh, M. Vesicles modulate an actin network for asymmetric spindle positioning. Nat. Cell Biol. 2013, 15, 937–947. [Google Scholar] [CrossRef] [PubMed]
- Neto, H.; Collins, L.L.; Gould, G.W. Vesicle trafficking and membrane remodelling in cytokinesis. Biochem. J. 2011, 437, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Anderson, D.J.; Hetzer, M.W. Reshaping of the endoplasmic reticulum limits the rate for nuclear envelope formation. J. Cell Biol. 2008, 182, 911–924. [Google Scholar] [CrossRef]
- Anderson, D.J.; Hetzer, M.W. Shaping the endoplasmic reticulum into the nuclear envelope. J. Cell Sci. 2008, 121, 137–142. [Google Scholar] [CrossRef] [PubMed]
- Colanzi, A.; Corda, D. Mitosis controls the Golgi and the Golgi controls mitosis. Curr. Opin. Cell Biol. 2007, 19, 386–393. [Google Scholar] [CrossRef] [PubMed]
- Valente, C.; Colanzi, A. Mechanisms and Regulation of the Mitotic Inheritance of the Golgi Complex. Front. Cell Dev. Biol. 2015, 3, 79. [Google Scholar] [CrossRef]
- Shibata, Y.; Voeltz, G.K.; Rapoport, T.A. Rough sheets and smooth tubules. Cell 2006, 126, 435–439. [Google Scholar] [CrossRef]
- Lu, L.; Ladinsky, M.S.; Kirchhausen, T. Cisternal organization of the endoplasmic reticulum during mitosis. Mol. Biol. Cell 2009, 20, 3471–3480. [Google Scholar] [CrossRef]
- Bobinnec, Y.; Marcaillou, C.; Morin, X.; Debec, A. Dynamics of the endoplasmic reticulum during early development of Drosophila melanogaster. Cell Motil Cytoskelet. 2003, 54, 217–225. [Google Scholar] [CrossRef]
- Bergman, Z.J.; McLaurin, J.D.; Eritano, A.S.; Johnson, B.M.; Sims, A.Q.; Riggs, B. Spatial reorganization of the endoplasmic reticulum during mitosis relies on mitotic kinase cyclin A in the early Drosophila embryo. PLoS ONE 2015, 10, e0117859. [Google Scholar] [CrossRef]
- Zheng, Y. A membranous spindle matrix orchestrates cell division. Nat. Rev. Mol. Cell Biol. 2010, 11, 529–535. [Google Scholar] [CrossRef] [PubMed]
- Wakeham, D.E.; Abi-Rached, L.; Towler, M.C.; Wilbur, J.D.; Parham, P.; Brodsky, F.M. Clathrin heavy and light chain isoforms originated by independent mechanisms of gene duplication during chordate evolution. Proc. Natl. Acad. Sci. USA 2005, 102, 7209–7214. [Google Scholar] [CrossRef] [PubMed]
- Puhka, M.; Vihinen, H.; Joensuu, M.; Jokitalo, E. Endoplasmic reticulum remains continuous and undergoes sheet-to-tubule transformation during cell division in mammalian cells. J. Cell Biol. 2007, 179, 895–909. [Google Scholar] [CrossRef] [PubMed]
- Tooze, J.; Hollinshead, M. Evidence that globular Golgi clusters in mitotic HeLa cells are clustered tubular endosomes. Eur. J. Cell Biol. 1992, 58, 228–242. [Google Scholar] [PubMed]
- Lucocq, J.M.; Berger, E.G.; Warren, G. Mitotic Golgi fragments in HeLa cells and their role in the reassembly pathway. J. Cell Biol. 1989, 109, 463–474. [Google Scholar] [CrossRef]
- Xie, S.; Reinecke, J.B.; Farmer, T.; Bahl, K.; Yeow, I.; Nichols, B.J.; McLamarrah, T.A.; Naslavsky, N.; Rogers, G.C.; Caplan, S. Vesicular trafficking plays a role in centriole disengagement and duplication. Mol. Biol. Cell 2018, 29, 2622–2631. [Google Scholar] [CrossRef]
- Naslavsky, N.; Caplan, S. Endocytic membrane trafficking in the control of centrosome function. Curr. Opin. Cell Biol. 2020, 65, 150–155. [Google Scholar] [CrossRef]
- Wang, L.; Dynlacht, B.D. The regulation of cilium assembly and disassembly in development and disease. Development 2018, 145, 151407. [Google Scholar] [CrossRef]
- Nachury, M.V.; Mick, D.U. Establishing and regulating the composition of cilia for signal transduction. Nat. Rev. Mol. Cell Biol. 2019, 20, 389–405. [Google Scholar]
- Miserey-Lenkei, S.; Colombo, M.I. Small RAB GTPases Regulate Multiple Steps of Mitosis. Front. Cell Dev. Biol. 2016, 4, 2. [Google Scholar] [CrossRef]
- Stenmark, H. Rab GTPases as coordinators of vesicle traffic. Nat. Rev. Mol. Cell Biol. 2009, 10, 513–525. [Google Scholar] [CrossRef] [PubMed]
- Dunster, K.; Toh, B.H.; Sentry, J.W. Early endosomes, late endosomes, and lysosomes display distinct partitioning strategies of inheritance with similarities to Golgi-derived membranes. Eur. J. Cell Biol. 2002, 81, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Boucrot, E.; Kirchhausen, T. Endosomal recycling controls plasma membrane area during mitosis. Proc. Natl. Acad. Sci. USA 2007, 104, 7939–7944. [Google Scholar] [CrossRef] [PubMed]
- Schweitzer, J.K.; Burke, E.E.; Goodson, H.V.; D’Souza-Schorey, C. Endocytosis resumes during late mitosis and is required for cytokinesis. J. Biol. Chem. 2005, 280, 41628–41635. [Google Scholar] [CrossRef]
- Lanzetti, L. A novel function of Rab5 in mitosis. Small GTPases 2012, 3, 168–172. [Google Scholar] [CrossRef][Green Version]
- Warner, A.K.; Keen, J.H.; Wang, Y.L. Dynamics of membrane clathrin-coated structures during cytokinesis. Traffic 2006, 7, 205–215. [Google Scholar] [CrossRef]
- Thompson, H.M.; Skop, A.R.; Euteneuer, U.; Meyer, B.J.; McNiven, M.A. The large GTPase dynamin associates with the spindle midzone and is required for cytokinesis. Curr. Biol. CB 2002, 12, 2111–2117. [Google Scholar] [CrossRef]
- Wienke, D.C.; Knetsch, M.L.; Neuhaus, E.M.; Reedy, M.C.; Manstein, D.J. Disruption of a dynamin homologue affects endocytosis, organelle morphology, and cytokinesis in Dictyostelium discoideum. Mol. Biol. Cell 1999, 10, 225–243. [Google Scholar] [CrossRef]
- Gerald, N.J.; Damer, C.K.; O’Halloran, T.J.; De Lozanne, A. Cytokinesis failure in clathrin-minus cells is caused by cleavage furrow instability. Cell Motil Cytoskelet. 2001, 48, 213–223. [Google Scholar] [CrossRef]
- Niswonger, M.L.; O’Halloran, T.J. A novel role for clathrin in cytokinesis. Proc. Natl. Acad. Sci. USA 1997, 94, 8575–8578. [Google Scholar] [CrossRef]
- Okamoto, C.T.; McKinney, J.; Jeng, Y.Y. Clathrin in mitotic spindles. Am. J. Physiol. Cell Physiol. 2000, 279, C369–C374. [Google Scholar] [CrossRef] [PubMed]
- Royle, S.J.; Bright, N.A.; Lagnado, L. Clathrin is required for the function of the mitotic spindle. Nature 2005, 434, 1152–1157. [Google Scholar] [CrossRef] [PubMed]
- Royle, S.J. The role of clathrin in mitotic spindle organisation. J. Cell Sci. 2012, 125, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Booth, D.G.; Hood, F.E.; Prior, I.A.; Royle, S.J. A TACC3/ch-TOG/clathrin complex stabilises kinetochore fibres by inter-microtubule bridging. EMBO J. 2011, 30, 906–919. [Google Scholar] [CrossRef]
- Lin, C.H.; Hu, C.K.; Shih, H.M. Clathrin heavy chain mediates TACC3 targeting to mitotic spindles to ensure spindle stability. J. Cell Biol. 2010, 189, 1097–1105. [Google Scholar] [CrossRef]
- Zhao, J.; Wang, L.; Zhou, H.X.; Liu, L.; Lu, A.; Li, G.P.; Schatten, H.; Liang, C.G. Clathrin heavy chain 1 is required for spindle assembly and chromosome congression in mouse oocytes. Microsc. Microanal. Off. J. Microsc. Soc. Am. Microbeam Anal. Soc. Microsc. Soc. Can. 2013, 19, 1364–1373. [Google Scholar] [CrossRef]
- Holzenspies, J.J.; Roelen, B.A.; Colenbrander, B.; Romijn, R.A.; Hemrika, W.; Stoorvogel, W.; van Haeften, T. Clathrin is essential for meiotic spindle function in oocytes. Reproduction 2010, 140, 223–233. [Google Scholar] [CrossRef]
- Cahill, M.A.; Jazayeri, J.A.; Catalano, S.M.; Toyokuni, S.; Kovacevic, Z.; Richardson, D.R. The emerging role of progesterone receptor membrane component 1 (PGRMC1) in cancer biology. Biochim. Biophys. Acta 2016, 1866, 339–349. [Google Scholar] [CrossRef]
- Wang-Eckhardt, L.; Eckhardt, M. A progesterone receptor membrane component 1 antagonist induces large vesicles independent of progesterone receptor membrane component 1 expression. Biol. Chem. 2020, 401, 1093–1099. [Google Scholar] [CrossRef]
- Teakel, S.L.; Ludescher, M.; Thejer, B.M.; Poschmann, G.; Forwood, J.K.; Neubauer, H.; Cahill, M.A. Protein complexes including PGRMC1 and actin-associated proteins are disrupted by AG-205. Biochem. Biophys. Res. Commun. 2020, 524, 64–69. [Google Scholar] [CrossRef]
- Neubauer, H.; Ruan, X.; Schneck, H.; Seeger, H.; Cahill, M.A.; Liang, Y.; Mafuvadze, B.; Hyder, S.M.; Fehm, T.; Mueck, A.O. Overexpression of progesterone receptor membrane component 1: Possible mechanism for increased breast cancer risk with norethisterone in hormone therapy. Menopause 2013, 20, 504–510. [Google Scholar] [CrossRef] [PubMed]
- Ruan, X.; Neubauer, H.; Yang, Y.; Schneck, H.; Schultz, S.; Fehm, T.; Cahill, M.A.; Seeger, H.; Mueck, A.O. Progestogens and membrane-initiated effects on the proliferation of human breast cancer cells. Climacteric J. Int. Menopause Soc. 2012, 15, 467–472. [Google Scholar] [CrossRef]
- Bai, Y.; Ludescher, M.; Poschmann, G.; Stuhler, K.; Wyrich, M.; Oles, J.; Franken, A.; Rivandi, M.; Abramova, A.; Reinhardt, F.; et al. PGRMC1 Promotes Progestin-Dependent Proliferation of Breast Cancer Cells by Binding Prohibitins Resulting in Activation of ERalpha Signaling. Cancers 2021, 13, 5635. [Google Scholar] [CrossRef] [PubMed]
- Peluso, J.J.; Liu, X.; Saunders, M.M.; Claffey, K.P.; Phoenix, K. Regulation of ovarian cancer cell viability and sensitivity to cisplatin by progesterone receptor membrane component-1. J. Clin. Endocrinol. Metab. 2008, 93, 1592–1599. [Google Scholar] [CrossRef] [PubMed]
- Peluso, J.J.; Gawkowska, A.; Liu, X.; Shioda, T.; Pru, J.K. Progesterone receptor membrane component-1 regulates the development and Cisplatin sensitivity of human ovarian tumors in athymic nude mice. Endocrinology 2009, 150, 4846–4854. [Google Scholar] [CrossRef] [PubMed]
- Luciano, A.M.; Lodde, V.; Franciosi, F.; Ceciliani, F.; Peluso, J.J. Progesterone receptor membrane component 1 expression and putative function in bovine oocyte maturation, fertilization, and early embryonic development. Reproduction 2010, 140, 663–672. [Google Scholar] [CrossRef]
- Ahmed, I.S.; Rohe, H.J.; Twist, K.E.; Mattingly, M.N.; Craven, R.J. Progesterone receptor membrane component 1 (Pgrmc1): A heme-1 domain protein that promotes tumorigenesis and is inhibited by a small molecule. J. Pharm. Exp. 2010, 333, 564–573. [Google Scholar] [CrossRef]
- Lodde, V.; Peluso, J.J. A novel role for progesterone and progesterone receptor membrane component 1 in regulating spindle microtubule stability during rat and human ovarian cell mitosis. Biol. Reprod. 2011, 84, 715–722. [Google Scholar] [CrossRef]
- Peluso, J.J.; Griffin, D.; Liu, X.; Horne, M. Progesterone receptor membrane component-1 (PGRMC1) and PGRMC-2 interact to suppress entry into the cell cycle in spontaneously immortalized rat granulosa cells. Biol. Reprod. 2014, 91, 104. [Google Scholar] [CrossRef]
- Clark, N.C.; Friel, A.M.; Pru, C.A.; Zhang, L.; Shioda, T.; Rueda, B.R.; Peluso, J.J.; Pru, J.K. Progesterone receptor membrane component 1 promotes survival of human breast cancer cells and the growth of xenograft tumors. Cancer Biol. 2016, 17, 262–271. [Google Scholar] [CrossRef]
- Terzaghi, L.; Tessaro, I.; Raucci, F.; Merico, V.; Mazzini, G.; Garagna, S.; Zuccotti, M.; Franciosi, F.; Lodde, V. PGRMC1 participates in late events of bovine granulosa cells mitosis and oocyte meiosis. Cell Cycle 2016, 15, 2019–2032. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kabe, Y.; Nakane, T.; Koike, I.; Yamamoto, T.; Sugiura, Y.; Harada, E.; Sugase, K.; Shimamura, T.; Ohmura, M.; Muraoka, K.; et al. Haem-dependent dimerization of PGRMC1/Sigma-2 receptor facilitates cancer proliferation and chemoresistance. Nat. Commun. 2016, 7, 11030. [Google Scholar] [CrossRef] [PubMed]
- Peluso, J.J.; Pru, C.A.; Liu, X.; Kelp, N.C.; Pru, J.K. Progesterone receptor membrane component 1 and 2 regulate granulosa cell mitosis and survival through a NFKappaB-dependent mechanismdagger. Biol. Reprod. 2019, 100, 1571–1580. [Google Scholar] [CrossRef] [PubMed]
- Meyer, C.; Schmid, R.; Scriba, P.C.; Wehling, M. Purification and partial sequencing of high-affinity progesterone-binding site(s) from porcine liver membranes. Eur. J. Biochem. FEBS 1996, 239, 726–731. [Google Scholar] [CrossRef] [PubMed]
- Peluso, J.J.; Liu, X.; Gawkowska, A.; Johnston-MacAnanny, E. Progesterone activates a progesterone receptor membrane component 1-dependent mechanism that promotes human granulosa/luteal cell survival but not progesterone secretion. J. Clin. Endocrinol. Metab. 2009, 94, 2644–2649. [Google Scholar] [CrossRef]
- Peluso, J.J.; Liu, X.; Gawkowska, A.; Lodde, V.; Wu, C.A. Progesterone inhibits apoptosis in part by PGRMC1-regulated gene expression. Mol. Cell. Endocrinol. 2010, 320, 153–161. [Google Scholar] [CrossRef]
- Peluso, J.J.; Pappalardo, A.; Losel, R.; Wehling, M. Progesterone membrane receptor component 1 expression in the immature rat ovary and its role in mediating progesterone’s antiapoptotic action. Endocrinology 2006, 147, 3133–3140. [Google Scholar] [CrossRef][Green Version]
- Peluso, J.J.; Romak, J.; Liu, X. Progesterone receptor membrane component-1 (PGRMC1) is the mediator of progesterone’s antiapoptotic action in spontaneously immortalized granulosa cells as revealed by PGRMC1 small interfering ribonucleic acid treatment and functional analysis of PGRMC1 mutations. Endocrinology 2008, 149, 534–543. [Google Scholar]
- Peluso, J.J.; Yuan, A.; Liu, X.; Lodde, V. Plasminogen activator inhibitor 1 RNA-binding protein interacts with progesterone receptor membrane component 1 to regulate progesterone’s ability to maintain the viability of spontaneously immortalized granulosa cells and rat granulosa cells. Biol. Reprod. 2013, 88, 20. [Google Scholar] [CrossRef][Green Version]
- Galluzzi, L.; Vitale, I.; Abrams, J.M.; Alnemri, E.S.; Baehrecke, E.H.; Blagosklonny, M.V.; Dawson, T.M.; Dawson, V.L.; El-Deiry, W.S.; Fulda, S.; et al. Molecular definitions of cell death subroutines: Recommendations of the Nomenclature Committee on Cell Death 2012. Cell Death Differ. 2012, 19, 107–120. [Google Scholar] [CrossRef]
- Kroemer, G.; Galluzzi, L.; Vandenabeele, P.; Abrams, J.; Alnemri, E.S.; Baehrecke, E.H.; Blagosklonny, M.V.; El-Deiry, W.S.; Golstein, P.; Green, D.R.; et al. Classification of cell death: Recommendations of the Nomenclature Committee on Cell Death 2009. Cell Death Differ 2009, 16, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Vitale, I.; Galluzzi, L.; Castedo, M.; Kroemer, G. Mitotic catastrophe: A mechanism for avoiding genomic instability. Nat. Rev. Mol. Cell Biol. 2011, 12, 385–392. [Google Scholar] [CrossRef] [PubMed]
- Aizen, J.; Pang, Y.; Harris, C.; Converse, A.; Zhu, Y.; Aguirre, M.A.; Thomas, P. Roles of progesterone receptor membrane component 1 and membrane progestin receptor alpha in regulation of zebrafish oocyte maturation. Gen. Comp. Endocrinol. 2018, 263, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.J.; Thomas, P.; Zhu, Y. Pgrmc1 Knockout Impairs Oocyte Maturation in Zebrafish. Front. Endocrinol. 2018, 9, 560. [Google Scholar] [CrossRef]
- Wu, X.J.; Zhu, Y. Downregulation of nuclear progestin receptor (Pgr) and subfertility in double knockouts of progestin receptor membrane component 1 (pgrmc1) and pgrmc2 in zebrafish. Gen. Comp. Endocrinol. 2020, 285, 113275. [Google Scholar] [CrossRef]
- Lessman, C.A. Oocyte maturation: Converting the zebrafish oocyte to the fertilizable egg. Gen. Comp. Endocrinol. 2009, 161, 53–57. [Google Scholar] [CrossRef]
- Nagahama, Y.; Yamashita, M. Regulation of oocyte maturation in fish. Dev. Growth Differ. 2008, 50 (Suppl. S1), S195–S219. [Google Scholar] [CrossRef]
- Luciano, A.M.; Franciosi, F.; Lodde, V.; Tessaro, I.; Corbani, D.; Modina, S.C.; Peluso, J.J. Oocytes isolated from dairy cows with reduced ovarian reserve have a high frequency of aneuploidy and alterations in the localization of progesterone receptor membrane component 1 and aurora kinase B. Biol. Reprod. 2013, 88, 58. [Google Scholar] [CrossRef]
- Juhlen, R.; Landgraf, D.; Huebner, A.; Koehler, K. Identification of a novel putative interaction partner of the nucleoporin ALADIN. Biol. Open 2016, 5, 1697–1705. [Google Scholar] [CrossRef]
- Luciano, A.M.; Peluso, J.J. PGRMC1 and the faithful progression through mitosis and meiosis. Cell Cycle 2016, 15, 2239–2240. [Google Scholar] [CrossRef]
- Juhlen, R.; Landgraf, D.; Huebner, A.; Koehler, K. Triple A patient cells suffering from mitotic defects fail to localize PGRMC1 to mitotic kinetochore fibers. Cell Div. 2018, 13, 8. [Google Scholar] [CrossRef] [PubMed]
- Skop, A.R.; Liu, H.; Yates, J., 3rd; Meyer, B.J.; Heald, R. Dissection of the mammalian midbody proteome reveals conserved cytokinesis mechanisms. Science 2004, 305, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Sauer, G.; Korner, R.; Hanisch, A.; Ries, A.; Nigg, E.A.; Sillje, H.H. Proteome analysis of the human mitotic spindle. Mol. Cell. Proteom. MCP 2005, 4, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Nousiainen, M.; Sillje, H.H.; Sauer, G.; Nigg, E.A.; Korner, R. Phosphoproteome analysis of the human mitotic spindle. Proc. Natl. Acad. Sci. USA 2006, 103, 5391–5396. [Google Scholar] [CrossRef]
- Cheeseman, I.M.; Desai, A. Molecular architecture of the kinetochore-microtubule interface. Nat. Rev. Mol. Cell Biol. 2008, 9, 33–46. [Google Scholar] [CrossRef]
- Malik, R.; Lenobel, R.; Santamaria, A.; Ries, A.; Nigg, E.A.; Korner, R. Quantitative analysis of the human spindle phosphoproteome at distinct mitotic stages. J. Proteome Res. 2009, 8, 4553–4563. [Google Scholar] [CrossRef]
- Ohta, S.; Bukowski-Wills, J.C.; Sanchez-Pulido, L.; Alves Fde, L.; Wood, L.; Chen, Z.A.; Platani, M.; Fischer, L.; Hudson, D.F.; Ponting, C.P.; et al. The protein composition of mitotic chromosomes determined using multiclassifier combinatorial proteomics. Cell 2010, 142, 810–821. [Google Scholar] [CrossRef]
- Ozlu, N.; Monigatti, F.; Renard, B.Y.; Field, C.M.; Steen, H.; Mitchison, T.J.; Steen, J.J. Binding partner switching on microtubules and aurora-B in the mitosis to cytokinesis transition. Mol. Cell. Proteom. MCP 2010, 9, 336–350. [Google Scholar] [CrossRef]
- Bonner, M.K.; Poole, D.S.; Xu, T.; Sarkeshik, A.; Yates, J.R., 3rd; Skop, A.R. Mitotic spindle proteomics in Chinese hamster ovary cells. PLoS ONE 2011, 6, e20489. [Google Scholar] [CrossRef]
- Biro, M.; Romeo, Y.; Kroschwald, S.; Bovellan, M.; Boden, A.; Tcherkezian, J.; Roux, P.P.; Charras, G.; Paluch, E.K. Cell cortex composition and homeostasis resolved by integrating proteomics and quantitative imaging. Cytoskeleton 2013, 70, 741–754. [Google Scholar] [CrossRef]
- Bovellan, M.; Romeo, Y.; Biro, M.; Boden, A.; Chugh, P.; Yonis, A.; Vaghela, M.; Fritzsche, M.; Moulding, D.; Thorogate, R.; et al. Cellular control of cortical actin nucleation. Curr. Biol. CB 2014, 24, 1628–1635. [Google Scholar] [CrossRef] [PubMed]
- Gupta, G.D.; Coyaud, E.; Goncalves, J.; Mojarad, B.A.; Liu, Y.; Wu, Q.; Gheiratmand, L.; Comartin, D.; Tkach, J.M.; Cheung, S.W.; et al. A Dynamic Protein Interaction Landscape of the Human Centrosome-Cilium Interface. Cell 2015, 163, 1484–1499. [Google Scholar] [CrossRef] [PubMed]
- Samejima, I.; Spanos, C.; Alves Fde, L.; Hori, T.; Perpelescu, M.; Zou, J.; Rappsilber, J.; Fukagawa, T.; Earnshaw, W.C. Whole-proteome genetic analysis of dependencies in assembly of a vertebrate kinetochore. J. Cell Biol. 2015, 211, 1141–1156. [Google Scholar] [CrossRef] [PubMed]
- McCloy, R.A.; Parker, B.L.; Rogers, S.; Chaudhuri, R.; Gayevskiy, V.; Hoffman, N.J.; Ali, N.; Watkins, D.N.; Daly, R.J.; James, D.E.; et al. Global Phosphoproteomic Mapping of Early Mitotic Exit in Human Cells Identifies Novel Substrate Dephosphorylation Motifs. Mol. Cell. Proteom. MCP 2015, 14, 2194–2212. [Google Scholar] [CrossRef] [PubMed]
- Rao, S.R.; Flores-Rodriguez, N.; Page, S.L.; Wong, C.; Robinson, P.J.; Chircop, M. The Clathrin-dependent Spindle Proteome. Mol. Cell. Proteom. MCP 2016, 15, 2537–2553. [Google Scholar] [CrossRef] [PubMed]
- Gheiratmand, L.; Coyaud, E.; Gupta, G.D.; Laurent, E.M.; Hasegan, M.; Prosser, S.L.; Goncalves, J.; Raught, B.; Pelletier, L. Spatial and proteomic profiling reveals centrosome-independent features of centriolar satellites. EMBO J. 2019, 38, e101109. [Google Scholar] [CrossRef] [PubMed]
- Garcia, Y.A.; Velasquez, E.F.; Gao, L.W.; Gholkar, A.A.; Clutario, K.M.; Cheung, K.; Williams-Hamilton, T.; Whitelegge, J.P.; Torres, J.Z. Mapping Proximity Associations of Core Spindle Assembly Checkpoint Proteins. J. Proteome Res. 2021, 20, 3414–3427. [Google Scholar] [CrossRef]
- Borek, W.E.; Vincenten, N.; Duro, E.; Makrantoni, V.; Spanos, C.; Sarangapani, K.K.; de Lima Alves, F.; Kelly, D.A.; Asbury, C.L.; Rappsilber, J.; et al. The Proteomic Landscape of Centromeric Chromatin Reveals an Essential Role for the Ctf19(CCAN) Complex in Meiotic Kinetochore Assembly. Curr. Biol. CB 2021, 31, 283–296.e287. [Google Scholar] [CrossRef]
- Fontaine-Lenoir, V.; Chambraud, B.; Fellous, A.; David, S.; Duchossoy, Y.; Baulieu, E.E.; Robel, P. Microtubule-associated protein 2 (MAP2) is a neurosteroid receptor. Proc. Natl. Acad. Sci. USA 2006, 103, 4711–4716. [Google Scholar] [CrossRef]
- Murakami, K.; Fellous, A.; Baulieu, E.E.; Robel, P. Pregnenolone binds to microtubule-associated protein 2 and stimulates microtubule assembly. Proc. Natl. Acad. Sci. USA 2000, 97, 3579–3584. [Google Scholar] [CrossRef]
- Eckerle, S.; Ringler, M.; Lecaudey, V.; Nitschke, R.; Driever, W. Progesterone modulates microtubule dynamics and epiboly progression during zebrafish gastrulation. Dev. Biol. 2018, 434, 249–266. [Google Scholar] [CrossRef] [PubMed]
- Thejer, B.M.; Adhikary, P.P.; Kaur, A.; Teakel, S.L.; Van Oosterum, A.; Seth, I.; Pajic, M.; Hannan, K.M.; Pavy, M.; Poh, P.; et al. PGRMC1 phosphorylation affects cell shape, motility, glycolysis, mitochondrial form and function, and tumor growth. BMC Mol. Cell Biol. 2020, 21, 24. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.Y.; Chou, H.C.; Law, C.H.; Chang, W.T.; Wen, T.N.; Liao, E.C.; Lin, M.W.; Lin, L.H.; Wei, Y.S.; Tsai, Y.T.; et al. Progesterone receptor membrane component 1 is involved in oral cancer cell metastasis. J. Cell Mol. Med. 2020, 24, 9737–9751. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.R.; Lee, Y.H.; Jo, S.L.; Heo, J.H.; Kim, G.; Lee, G.S.; An, B.S.; Baek, I.J.; Hong, E.J. Absence of progesterone receptor membrane component 1 reduces migration and metastasis of breast cancer. Cell Commun. Signal 2021, 19, 42. [Google Scholar] [CrossRef]
- Xu, X.; Ruan, X.; Ju, R.; Wang, Z.; Yang, Y.; Cheng, J.; Gu, M.; Mueck, A.O. Progesterone Receptor Membrane Component-1 May Promote Survival of Human Brain Microvascular Endothelial Cells in Alzheimer’s Disease. Am. J. Alzheimers Dis. Other Demen. 2022, 37, 15333175221109749. [Google Scholar] [CrossRef]
- Salsano, S.; Quinonero, A.; Perez, S.; Garrido Gomez, T.; Simon, C.; Dominguez, F. Dynamic expression of PGRMC1 and SERBP1 in human endometrium: An implication in the human decidualization process. Fertil. Steril. 2017, 108, 832–842.e831. [Google Scholar] [CrossRef]
- Salsano, S.; Gonzalez-Martin, R.; Quinonero, A.; Lopez-Martin, S.; Gomez-Escribano, A.P.; Perez-Deben, S.; Yanez-Mo, M.; Dominguez, F. Novel nonclassic progesterone receptor PGRMC1 pulldown-precipitated proteins reveal a key role during human decidualization. Fertil. Steril. 2020, 113, 1050–1066.e1057. [Google Scholar] [CrossRef]
- Cahill, M.A. Choose your partners for the next dance: Implied PGRMC1 roles in membrane trafficking and mitochondrial modulation. Fertil. Steril. 2020, 113, 938–941. [Google Scholar] [CrossRef]
- Hehenberger, E.; Eitel, M.; Fortunato, S.A.V.; Miller, D.J.; Keeling, P.J.; Cahill, M.A. Early eukaryotic origins and metazoan elaboration of MAPR family proteins. Mol. Phylogenet. Evol. 2020, 148, 106814. [Google Scholar] [CrossRef]
- Bagchi, S.; Fredriksson, R.; Wallen-Mackenzie, A. In Situ Proximity Ligation Assay (Pla). Methods Mol. Biol. 2015, 1318, 149–159. [Google Scholar]
- Terzaghi, L.; Luciano, A.M.; Dall’Acqua, P.C.; Modina, S.C.; Peluso, J.J.; Lodde, V. PGRMC1 localization and putative function in the nucleolus of bovine granulosa cells and oocytes. Reproduction 2018, 155, 273–282. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.D.; Flamini, M.; Sanchez, A.M.; Goglia, L.; Giretti, M.S.; Genazzani, A.R.; Simoncini, T. Progestogens regulate endothelial actin cytoskeleton and cell movement via the actin-binding protein moesin. Mol. Hum. Reprod. 2008, 14, 225–234. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Stournaras, C.; Gravanis, A.; Margioris, A.N.; Lang, F. The actin cytoskeleton in rapid steroid hormone actions. Cytoskeleton 2014, 71, 285–293. [Google Scholar] [CrossRef] [PubMed]
- Wessel, L.; Balakrishnan-Renuka, A.; Henkel, C.; Meyer, H.E.; Meller, K.; Brand-Saberi, B.; Theiss, C. Long-term incubation with mifepristone (MLTI) increases the spine density in developing Purkinje cells: New insights into progesterone receptor mechanisms. Cell Mol. Life Sci. 2014, 71, 1723–1740. [Google Scholar] [CrossRef] [PubMed]
- Wessel, L.; Olbrich, L.; Brand-Saberi, B.; Theiss, C. New aspects of progesterone interactions with the actin cytoskeleton and neurosteroidogenesis in the cerebellum and the neuronal growth cone. J Histochem. Cytochem. 2014, 62, 835–845. [Google Scholar] [CrossRef]
- Ahmed, I.S.; Rohe, H.J.; Twist, K.E.; Craven, R.J. Pgrmc1 (progesterone receptor membrane component 1) associates with epidermal growth factor receptor and regulates erlotinib sensitivity. J. Biol. Chem. 2010, 285, 24775–24782. [Google Scholar] [CrossRef]
- Zhang, M.; Robitaille, M.; Showalter, A.D.; Huang, X.; Liu, Y.; Bhattacharjee, A.; Willard, F.S.; Han, J.; Froese, S.; Wei, L.; et al. Progesterone receptor membrane component 1 is a functional part of the glucagon-like peptide-1 (GLP-1) receptor complex in pancreatic beta cells. Mol. Cell. Proteom. MCP 2014, 13, 3049–3062. [Google Scholar] [CrossRef]
- Hampton, K.K.; Anderson, K.; Frazier, H.; Thibault, O.; Craven, R.J. Insulin Receptor Plasma Membrane Levels Increased by the Progesterone Receptor Membrane Component 1. Mol. Pharm. 2018, 94, 665–673. [Google Scholar] [CrossRef]
- Kaksonen, M.; Roux, A. Mechanisms of clathrin-mediated endocytosis. Nat. Rev. Mol. Cell Biol. 2018, 19, 313–326. [Google Scholar] [CrossRef]
- Riad, A.; Zeng, C.; Weng, C.C.; Winters, H.; Xu, K.; Makvandi, M.; Metz, T.; Carlin, S.; Mach, R.H. Sigma-2 Receptor/TMEM97 and PGRMC-1 Increase the Rate of Internalization of LDL by LDL Receptor through the Formation of a Ternary Complex. Sci. Rep. 2018, 8, 16845. [Google Scholar] [CrossRef]
- Furthauer, M.; Gonzalez-Gaitan, M. Endocytosis and mitosis: A two-way relationship. Cell Cycle 2009, 8, 3311–3318. [Google Scholar] [PubMed]
- Yu, H.; Li, Y.; Li, L.; Huang, J.; Wang, X.; Tang, R.; Jiang, Z.; Lv, L.; Chen, F.; Yu, C.; et al. Functional reciprocity of proteins involved in mitosis and endocytosis. FEBS J. 2021, 288, 5850–5866. [Google Scholar] [CrossRef]
- Unanue, E.R.; Ungewickell, E.; Branton, D. The binding of clathrin triskelions to membranes from coated vesicles. Cell 1981, 26 Pt 1, 439–446. [Google Scholar] [CrossRef] [PubMed]
- Reider, A.; Wendland, B. Endocytic adaptors—Social networking at the plasma membrane. J. Cell Sci. 2011, 124, 1613–1622. [Google Scholar] [CrossRef]
- Kiyomitsu, T.; Boerner, S. The Nuclear Mitotic Apparatus (NuMA) Protein: A Key Player for Nuclear Formation, Spindle Assembly, and Spindle Positioning. Front. Cell Dev. Biol. 2021, 9, 653801. [Google Scholar] [CrossRef]
- Ma, N.; Matsunaga, S.; Takata, H.; Ono-Maniwa, R.; Uchiyama, S.; Fukui, K. Nucleolin functions in nucleolus formation and chromosome congression. J. Cell Sci. 2007, 120, 2091–2105. [Google Scholar] [CrossRef]
- Kaluka, D.; Batabyal, D.; Chiang, B.Y.; Poulos, T.L.; Yeh, S.R. Spectroscopic and mutagenesis studies of human PGRMC1. Biochemistry 2015, 54, 1638–1647. [Google Scholar] [CrossRef]
- Cahill, M.A.; Jazayeri, J.A.; Kovacevic, Z.; Richardson, D.R. PGRMC1 regulation by phosphorylation: Potential new insights in controlling biological activity. Oncotarget 2016, 7, 50822–50827. [Google Scholar] [CrossRef]
- Sabbir, M.G.; Inoue, A.; Taylor, C.G.; Zahradka, P. Loss of beta-Arrestins or six Galpha proteins in HEK293 cells caused Warburg effect and prevented progesterone-induced rapid proteasomal degradation of progesterone receptor membrane component 1. J. Steroid. Biochem. Mol. Biol. 2021, 214, 105995. [Google Scholar] [CrossRef] [PubMed]
- Sabbir, M.G. Progesterone induced Warburg effect in HEK293 cells is associated with post-translational modifications and proteasomal degradation of progesterone receptor membrane component 1. J. Steroid. Biochem. Mol. Biol. 2019, 191, 105376. [Google Scholar] [CrossRef]
- Peluso, J.J.; Lodde, V.; Liu, X. Progesterone regulation of progesterone receptor membrane component 1 (PGRMC1) sumoylation and transcriptional activity in spontaneously immortalized granulosa cells. Endocrinology 2012, 153, 3929–3939. [Google Scholar] [CrossRef]
- Kim, J.Y.; Kim, S.Y.; Choi, H.S.; Kim, M.K.; Lee, H.M.; Jang, Y.-J.; Ryu, C.J. Progesterone Receptor Membrane Component 1 suppresses the p53 and Wnt/β-catenin pathways to promote human pluripotent stem cell self-renewal. Sci. Rep. 2018, 8, 3048. [Google Scholar] [CrossRef]
- Thomas, P.; Pang, Y.; Dong, J. Enhancement of cell surface expression and receptor functions of membrane progestin receptor alpha (mPRalpha) by progesterone receptor membrane component 1 (PGRMC1): Evidence for a role of PGRMC1 as an adaptor protein for steroid receptors. Endocrinology 2014, 155, 1107–1119. [Google Scholar] [CrossRef] [PubMed]
- Aizen, J.; Thomas, P. Role of Pgrmc1 in estrogen maintenance of meiotic arrest in zebrafish oocytes through Gper/Egfr. J. Endocrinol. 2015, 225, 59–68. [Google Scholar] [CrossRef]
- Griffin, D.; Liu, X.; Pru, C.; Pru, J.K.; Peluso, J.J. Expression of progesterone receptor membrane component-2 within the immature rat ovary and its role in regulating mitosis and apoptosis of spontaneously immortalized granulosa cells. Biol. Reprod. 2014, 91, 36. [Google Scholar] [CrossRef] [PubMed]
- Cahill, M.A. The evolutionary appearance of signaling motifs in PGRMC1. Biosci. Trends 2017, 11, 179–192. [Google Scholar] [CrossRef] [PubMed][Green Version]




{kind=link}
{kind=link}
{kind=link}
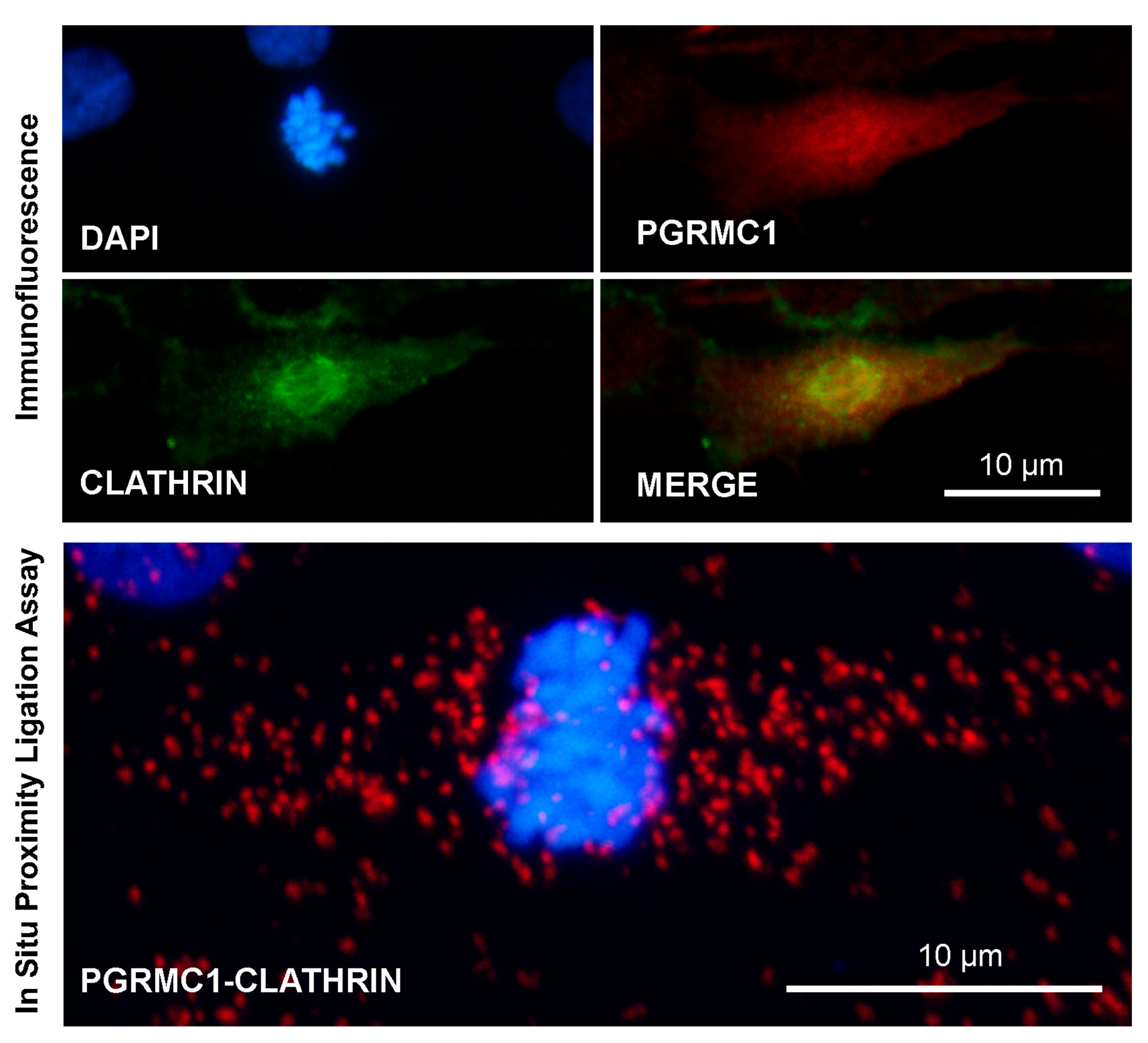{kind=link}
| Reference Title | Year | Cell Type | Experimental Approach Used to Disturb PGRMC1 Function | Effect on Cell Proliferation (Phenotype) | Proposed Mechanism (If Any) | Ref. |
|---|---|---|---|---|---|---|
| Regulation of ovarian cancer cell viability and sensitivity to cisplatin by progesterone receptor membrane component 1 | 2008 | Ovcar-3 cells | Overexpression of exogenous PGRMC1; siRNA-mediated gene silencing; transfection of antibody | Increased cell viability in response to cisplatin | Regulation of apoptosis and P4 antiapoptotic action | [106] |
| Progesterone receptor membrane component-1 regulates the development and Cisplatin sensitivity of human ovarian tumors in athymic nude mice | 2009 | Human ovarian cancer cells (SKOV-3 cells) | Gene silencing by short hairpin RNA knockdown approach; xenograft model of athymic nude mice | Lowered in vitro growth and reduced tumor xenograft growth | Regulation of apoptosis and P4 antiapoptotic action | [107] |
| Progesterone receptor membrane component 1 expression and putative function in bovine oocyte maturation, fertilization, and early embryonic development | 2010 | Bovine oocytes | Antibody injection | Impaired meiotic progression | Regulation of meiotic spindle function | [108] |
| Progesterone receptor membrane component 1 (Pgrmc1): a heme-1 domain protein that promotes tumorigenesis and is inhibited by a small molecule | 2010 | Human A549 non-small cell lung cancer cells and MDA- MB-468 breast cancer cells | siRNA-mediated gene silencing and short hairpin RNA knockdown approach; xenograft model of athymic nude mice | Reduced tumor xenograft growth | - | [109] |
| A novel role for progesterone and progesterone receptor membrane component 1 in regulating spindle microtubule stability during rat and human ovarian cell mitosis | 2011 | Rat spontaneously immortalized granulosa cells (SIGCs) and human ovarian cancer cells (SKOV-3 cells) | Antibody transfection; PGRMC1 downregulation | Lowered growth rate | Regulation of spindle function (microtubule-mediated process) | [110] |
| Progestogens and membrane-initiated effects on the proliferation of human breast cancer cells | 2012 | MCF-7 | PGRMC1 overexpression | Increased cell proliferation in response to progestin treatment | [104] | |
| Overexpression of progesterone receptor membrane component 1: possible mechanism for increased breast cancer risk with norethisterone in hormone therapy | 2013 | MCF-7 | PGRMC1 overexpression; xenograft model of athymic nude mice | Increased cell proliferation of PGRMC1-overexpressing breast cancer cells in response to E2/NET combination | [103] | |
| Progesterone receptor membrane component-1 (PGRMC1) and PGRMC-2 interact to suppress entry into the cell cycle in spontaneously immortalized rat granulosa cells | 2014 | Rat spontaneously immortalized granulosa cells (SIGCs) | siRNA-mediated gene silencing | Increased entry into the cell cycle without cell proliferation—PGRMC1- and/or PGRMC2-depleted cells accumulate in metaphase and undergo apoptosis | Regulation of entry into the G1 stage of the cell cycle through interaction with PGRMC2 and G3BP2 | [111] |
| Progesterone receptor membrane component 1 promotes survival of human breast cancer cells and the growth of xenograft tumors | 2016 | MDA-MB-468 breast cancer cells | Short hairpin RNA knockdown approach; xenograft model of athymic nude mice | Reduced tumor xenograft growth | Regulation of cell viability | [112] |
| PGRMC1 participates in late events of bovine granulosa cells mitosis and oocyte meiosis | 2016 | Primary bovine granulosa cell culture; bovine oocytes | siRNA-mediated gene silencing | Decreased cell proliferation; accumulation of M-phase cells that eventually die; defective meiotic maturation | Regulation of cytokinesis and of mitotic spindle function through association with AURKB | [113] |
| Haem-dependent dimerization of PGRMC1/Sigma-2 receptor facilitates cancer proliferation and chemoresistance | 2016 | HCT116 cells and derived tumors in a model of liver metastases of human colon cancer | Stable PGRMC1 knockdown and xenograft model of NOG mice | Reduced cell proliferation of spheroids grown in vitro and reduced tumor growth in vivo | Regulation of EGFR and cytochrome P450 signaling | [114] |
| Progesterone receptor membrane components 1 and 2 regulate granulosa cell mitosis and survival through a NFKappaB-dependent mechanism | 2019 | Mouse ovarian cells | Conditional knockout PGRMC1 mice; siRNA-mediated gene silencing | Increased follicular atresia—ovarian granulosa cells of PGRMC1 conditional KO mice enter the cell cycle more frequently compared to controls but then do not seem to progress, causing increased follicular atresia | Regulation of entry into the cell cycle by an NFkB-mediated action | [115] |
| PGRMC1 Promotes Progestin-Dependent Proliferation of Breast Cancer Cells by Binding Prohibitins Resulting in Activation of ERalpha Signaling | 2021 | Various breast cancer cell lines | PGRMC1 overexpression | Increased cell proliferation in response to progestin treatment | Regulation of ERα signaling | [105] |
| Title | Year | Cell Type | Cellular Fraction | Method | Presence of PGRMC1 (Y/N) | Ref. |
|---|---|---|---|---|---|---|
| Dissection of the mammalian midbody proteome reveals conserved cytokinesis mechanisms | 2004 | Chinese hamster ovaries cells | Midbody | Mass spectrometry | N | [134] |
| Proteome analysis of the human mitotic spindle | 2005 | HeLa S3 cells | Mitotic spindle | Mass spectrometry | N | [135] |
| Phosphoproteome analysis of the human mitotic spindle | 2006 | HeLa S3 cells | Mitotic spindle | Mass spectrometry | Y | [136] |
| Molecular architecture of the kinetochore–microtubule interface | 2008 | Various cell type | Kinetochore | Review paper | N | [137] |
| Quantitative analysis of the human spindle phosphoproteome at distinct mitotic stages | 2009 | HeLa S3 cells | Mitotic spindle | SILAC technology | N | [138] |
| The protein composition of mitotic chromosomes determined using multiclassifier combinatorial proteomics | 2010 | Chicken DT40 cells | Mitotic chromosome (kinetochore) | SILAC technology | Y | [139] |
| Binding Partner Switching on Microtubules and Aurora-B in the Mitosis to Cytokinesis Transition | 2010 | HeLa S3 cells | M/C phase (microtubules) | SILAC MS | Y | [140] |
| Mitotic spindle proteomics in Chinese hamster ovary cells | 2011 | Chinese hamster ovaries cells | Mitotic spindle | Mass spectrometry | Y | [141] |
| Cell cortex composition and homeostasis resolved by integrating proteomics and quantitative imaging | 2013 | Human melanoma cells/HeLa cells | Cell cortex (MII cells) | LC-MS/MS | N | [142] |
| Cellular control of cortical actin nucleation | 2014 | Human melanoma cells/HeLa cells | Cell cortex (MII cells) | LC-MS/MS | N | [143] |
| A dynamic protein interaction landscape of the human centrosome-cilium interface | 2015 | 293 T-REx cells | Centrioles | Mass spectrometry | Y | [144] |
| Whole-proteome genetic analysis of dependencies in assembly of a vertebrate kinetochore | 2015 | Chicken lymphoma B cell line DT40 | Mitotic chromosome (kinetochore) | Mass spectrometry | N | [145] |
| Global phosphoproteomic mapping of early mitotic exit in human cells identifies novel substrate dephosphorylation motifs | 2015 | HeLa Cells | Mitotic spindle | SILAC technology | Y | [146] |
| The clathrin-dependent spindle proteome | 2016 | Human HeLa cells | Mitotic spindle (KT and centrosomes) | LC-MS/MS | Y | [147] |
| Spatial and proteomic profiling reveals centrosome-independent features of centriolar satellites | 2019 | Flp-In T-REx 293 (human) | Microtubules | mass spectrometry | N | [148] |
| Mapping Proximity Associations of Core Spindle Assembly Checkpoint Proteins | 2021 | HeLa Flp-In T-Rex and retinal pigment epithelium (RPE) cells | Kinetochore (BUB1 BUB1B BUB3) association | LC-MS/MS | Y | [149] |
| The Proteomic Landscape of Centromeric Chromatin Reveals an Essential Role for the Ctf19CCAN Complex in Meiotic Kinetochore Assembly | 2021 | Yeast (S cerevisiae) | Meiotic centromeres and kinetochores | Label-Free Mass Spectrometry (LFQMS) | N | [150] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lodde, V.; Garcia Barros, R.; Terzaghi, L.; Franciosi, F.; Luciano, A.M. Insights on the Role of PGRMC1 in Mitotic and Meiotic Cell Division. Cancers 2022, 14, 5755. https://doi.org/10.3390/cancers14235755
Lodde V, Garcia Barros R, Terzaghi L, Franciosi F, Luciano AM. Insights on the Role of PGRMC1 in Mitotic and Meiotic Cell Division. Cancers. 2022; 14(23):5755. https://doi.org/10.3390/cancers14235755
Chicago/Turabian StyleLodde, Valentina, Rodrigo Garcia Barros, Laura Terzaghi, Federica Franciosi, and Alberto Maria Luciano. 2022. "Insights on the Role of PGRMC1 in Mitotic and Meiotic Cell Division" Cancers 14, no. 23: 5755. https://doi.org/10.3390/cancers14235755
APA StyleLodde, V., Garcia Barros, R., Terzaghi, L., Franciosi, F., & Luciano, A. M. (2022). Insights on the Role of PGRMC1 in Mitotic and Meiotic Cell Division. Cancers, 14(23), 5755. https://doi.org/10.3390/cancers14235755